
Effect of Dietary Supplementation of Lactiplantibacillus plantarum N-1 and Its Synergies with Oligomeric Isomaltose on the Growth Performance and Meat Quality in Hu Sheep

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. Sample Collection
2.3. Determination of Blood Lipids in Hu Sheep Serum
2.4. Meat Quality Analysis
2.5. Determination of the Chemical Composition of Hu Sheep Muscle
2.6. Determination of the Mineral Elements in Hu Sheep Tissue
2.7. Determination of the Texture and Shear Force of Hu Sheep Muscle Tissue
2.8. Determination of the Amino Acids in Hu Sheep Muscle Tissue
2.9. Determination of the Volatile Compounds in Hu Sheep Muscle Tissue
2.10. Determination of Carbonyl and Sulfhydryl Group Content in Myofibrillar Protein
2.11. Determination of TBARS in Hu Sheep Muscle Tissue
2.12. Determination of Surface Hydrophobicity, Circular Dichroism, and Fluorescence Spectroscopy of MP
2.13. Quantitative Real-Time PCR Analysis
2.14. Statistical Analysis
3. Results and Discussion
3.1. Growth and Slaughter Performance
3.2. Meat Quality
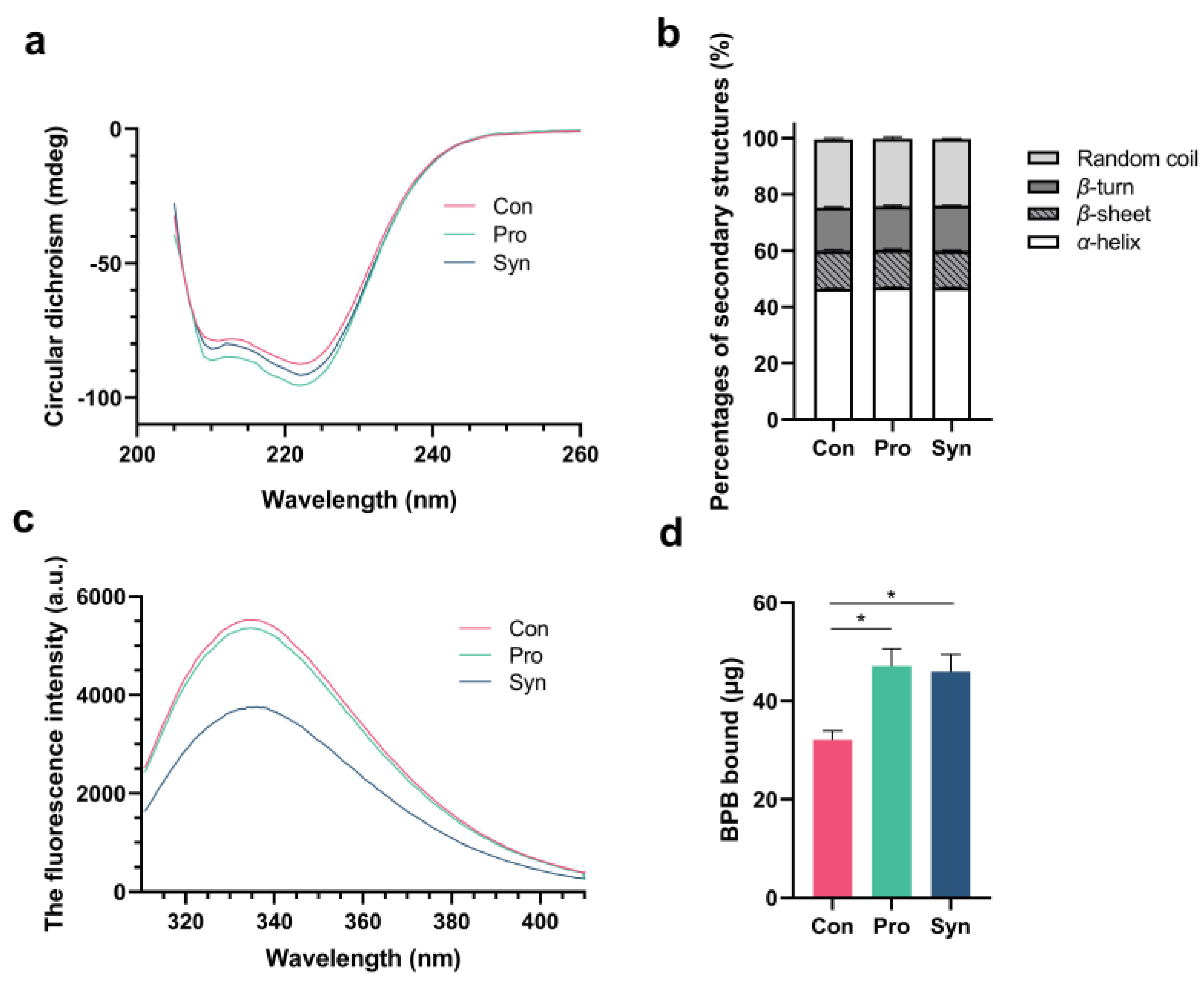
3.3. Protein Structure and Surface Hydrophobicity of MP
3.4. Amino Acids of LT Muscle
3.5. Volatile Compounds in LT Muscle
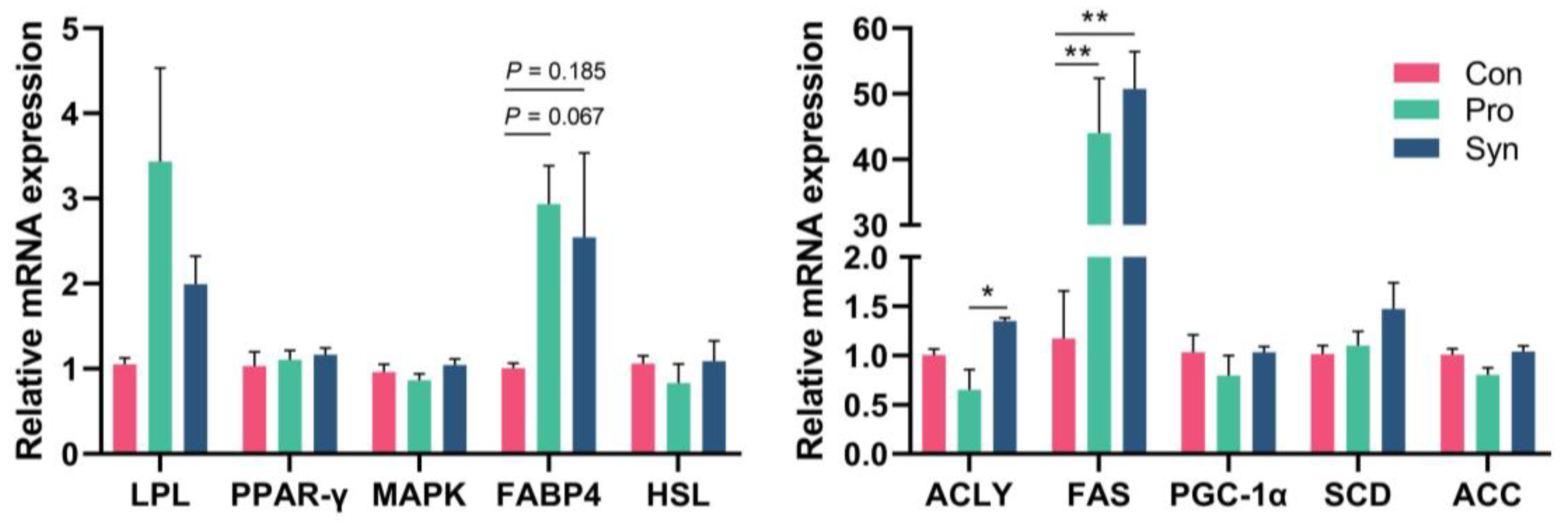
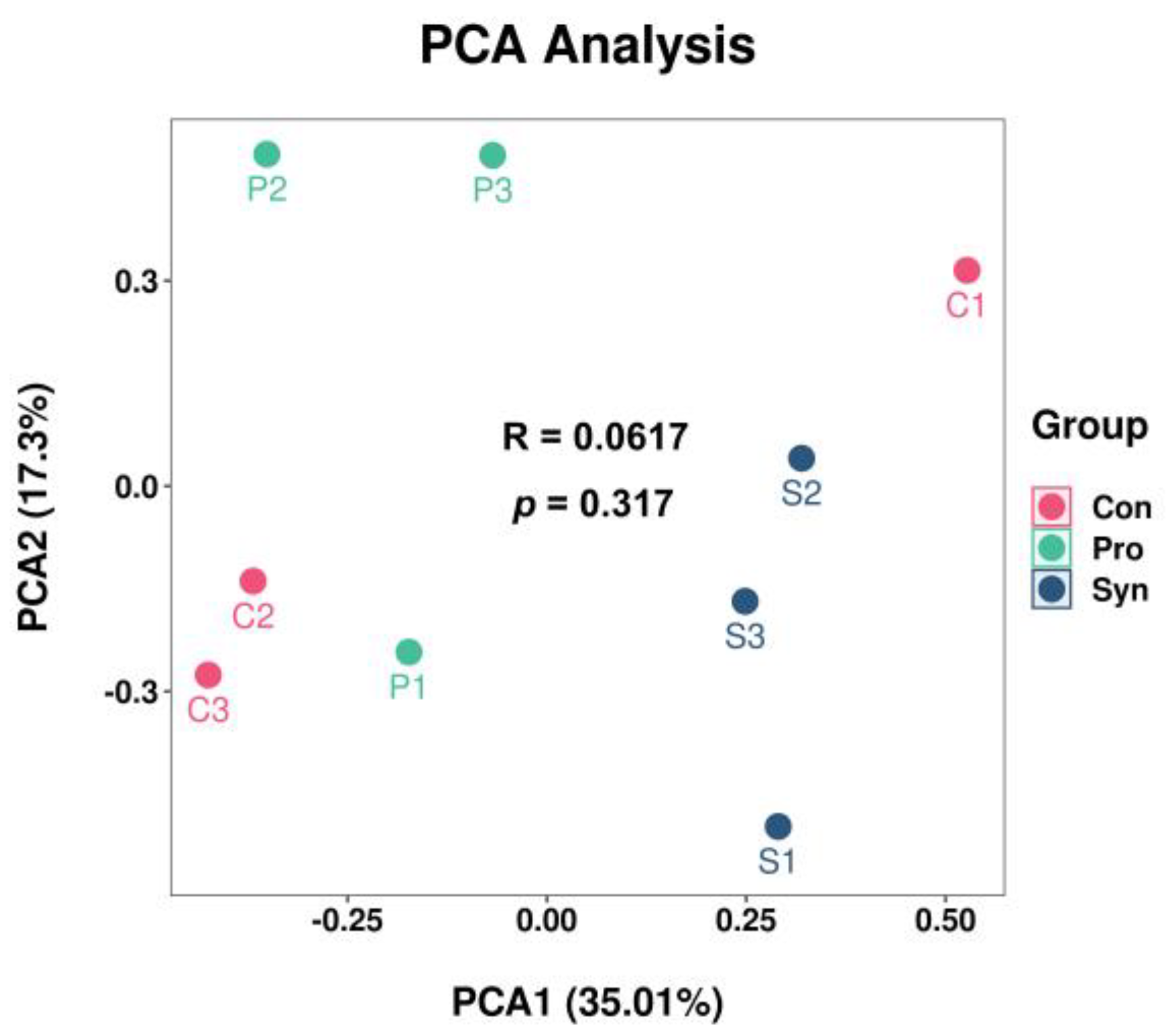
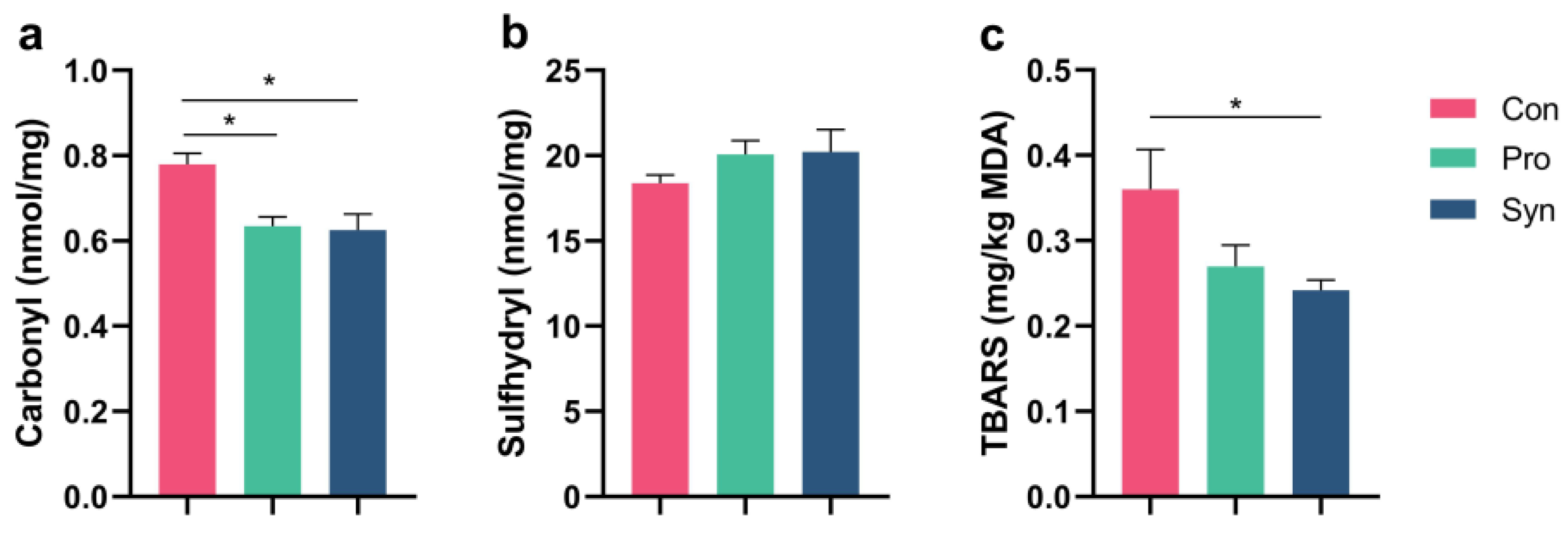
3.6. Antioxidant Capacity of Hu Sheep Tissue
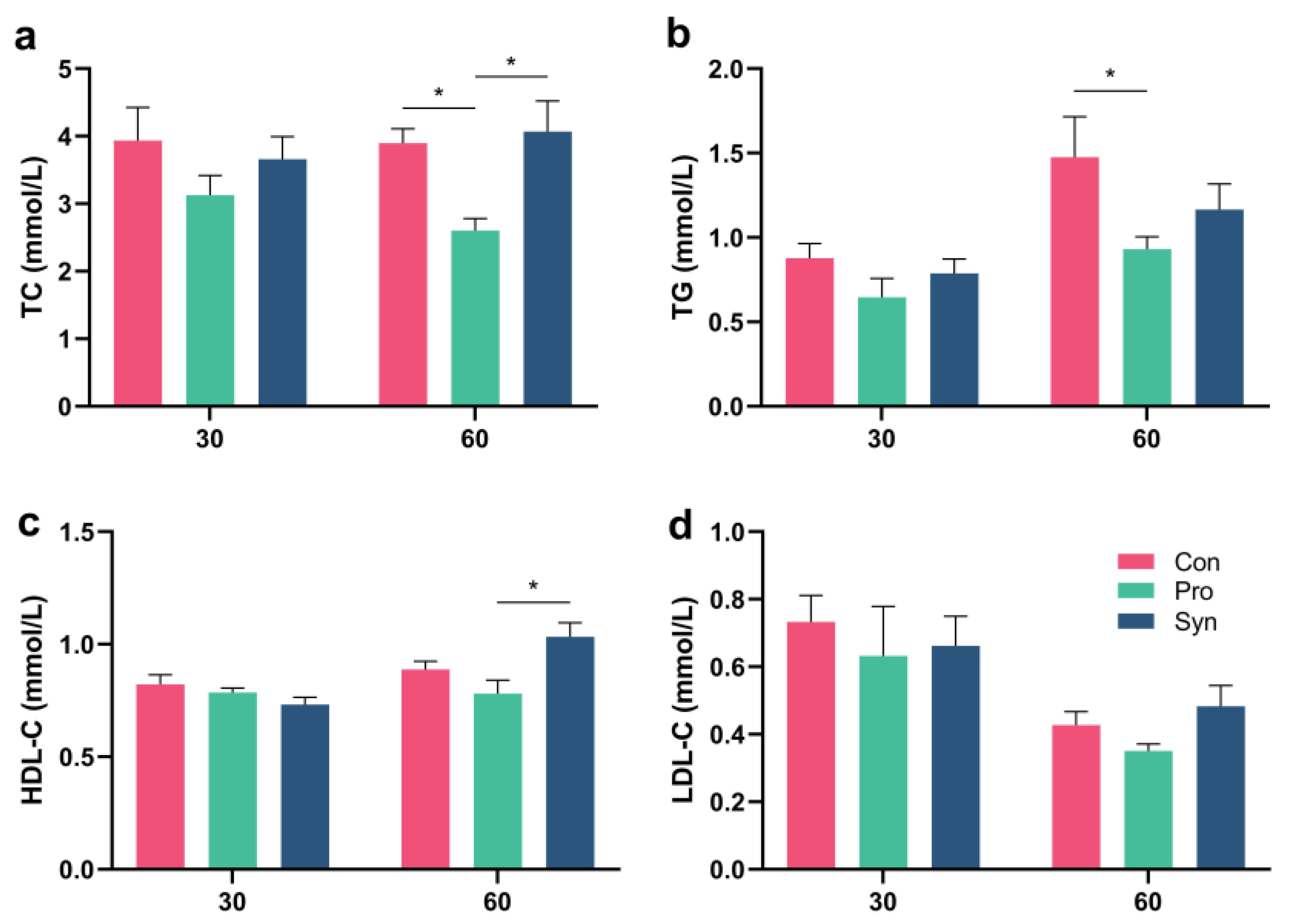
3.7. Blood Lipids in Hu Sheep Serum
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lu, Z.; Li, J.; Yuan, C.; Xi, B.; Yang, B.; Meng, X.; Guo, T.; Yue, Y.; Gao, Y.; Liu, J.; et al. Evaluation of mutton quality characteristics of dongxiang tribute sheep based on membership function and gas chromatography and Ion mobility spectrometry. Front. Nutr. 2022, 9, 852399. [Google Scholar] [CrossRef]
- Ripoll, G.; Joy, M.; Panea, B. Consumer Perception of the Quality of Lamb and Lamb Confit. Foods 2018, 7, 80. [Google Scholar] [CrossRef]
- Nie, C.; Hu, Y.; Chen, R.; Guo, B.; Li, L.; Chen, H.; Chen, H.; Song, X. Effect of probiotics and Chinese medicine polysaccharides on meat quality, muscle fibre type and intramuscular fat deposition in lambs. Ital. J. Anim. Sci 2022, 21, 811–820. [Google Scholar] [CrossRef]
- Chen, B.; Yue, Y.; Li, J.; Liu, J.; Yuan, C.; Guo, T.; Zhang, D.; Yang, B.; Lu, Z. Transcriptome-metabolome analysis reveals how sires affect meat quality in hybrid sheep populations. Front. Nutr. 2022, 9, 967985. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A. Dietary postbiotic Lactobacillus plantarum improves serum and ruminal antioxidant activity and upregulates hepatic antioxidant enzymes and ruminal barrier function in post-weaning lambs. Antioxidants 2020, 9, 250. [Google Scholar] [CrossRef]
- Liu, C.; Hou, Y.; Su, R.; Luo, Y.; Dou, L.; Yang, Z.; Yao, D.; Wang, B.; Zhao, L.; Su, L.; et al. Effect of dietary probiotics supplementation on meat quality, volatile flavor compounds, muscle fiber characteristics, and antioxidant capacity in lambs. Food Sci. Nutr. 2022, 10, 2646–2658. [Google Scholar] [CrossRef]
- Aluwong, T.; Kawu, M.; Raji, M.; Dzenda, T.; Govwang, F.; Sinkalu, V.; Ayo, J. Effect of yeast probiotic on growth, antioxidant enzyme activities and malondialdehyde concentration of broiler chickens. Antioxidants 2013, 2, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics—A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.F.; Shahzad, M.A.; Sarwar, M.; Rehman, A.U.; Sharif, M.; Mukhtar, N. Probiotics and lamb performance: A review. Afr. J. Agric. Res. 2011, 6, 5198–5203. [Google Scholar]
- Saleem, A.M.; Zanouny, A.I.; Singer, A.M. Growth performance, nutrients digestibility, and blood metabolites of lambs fed diets supplemented with probiotics during pre- and post-weaning period. Asian-Australas. J. Anim. Sci. 2017, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Dawson, K.A.; Newman, K.E.; Boling, J.A. Effects of microbial supplements containing yeast and lactobacilli on roughage-fed ruminal microbial activities. J. Anim. Sci. 1990, 68, 3392–3398. [Google Scholar] [CrossRef] [PubMed]
- Izuddin, W.I.; Loh, T.C.; Samsudin, A.A.; Foo, H.L.; Humam, A.M.; Shazali, N. Effects of postbiotic supplementation on growth performance, ruminal fermentation and microbial profile, blood metabolite and GHR, IGF-1 and MCT-1 gene expression in post-weaning lambs. BMC Vet. Res. 2019, 15, 315. [Google Scholar] [CrossRef]
- Tang, X.; Liu, X.; Liu, H. Effects of dietary probiotic (Bacillus subtilis) supplementation on carcass traits, meat quality, amino acid, and fatty acid profile of broiler chickens. Front. Vet. Sci. 2021, 8, 767802. [Google Scholar] [CrossRef]
- Tian, Z.; Deng, D.; Cui, Y.; Chen, W.; Yu, M.; Ma, X. Diet supplemented with fermented okara improved growth performance, meat quality, and amino acid profiles in growing pigs. Food Sci. Nutr. 2020, 8, 5650–5659. [Google Scholar] [CrossRef] [PubMed]
- Grela, E.R.; Swiatkiewicz, M.; Florek, M.; Bakowski, M.; Skiba, G. Effect of Inulin source and a probiotic supplement in pig diets on carcass traits, meat quality and fatty acid composition in finishing pigs. Animals 2021, 11, 2438. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Arif, M.; Sajjad, N.; Al-Ghadi, M.Q.; Alagawany, M.; Abd El-Hack, M.E.; Alhimaidi, A.R.; Elnesr, S.S.; Almutairi, B.O.; Amran, R.A.; et al. Dietary effect of probiotics and prebiotics on broiler performance, carcass, and immunity. Poult. Sci. 2020, 99, 6946–6953. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Liu, R.; Zhou, Z.; Xu, X.; Feng, S.; Kushmaro, A.; Marks, R.S.; Wang, D.; Sun, Q. Probiotic characteristics of Lactiplantibacillus plantarum N-1 and its cholesterol-lowering effect in hypercholesterolemic rats. Probiotics Antimicrob. Proteins 2022, 14, 337–348. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of the AOAC International, 16th ed.; AOAC International: Arlington, MA, USA, 1995. [Google Scholar]
- Zhou, Z.Q.; Xu, Q.D.; Chen, L.; Chen, N.; Gao, H.X.; Sun, Q.; Zeng, W.C. Interaction and action mechanism of quercetin and myofibrillar protein and its effects on the quality of cured meat. J. Food Process. Preserv. 2021, 45, e16020. [Google Scholar] [CrossRef]
- Wu, H.; Li, T.; Lv, J.; Chen, Z.; Wu, J.; Wang, N.; Wu, H.; Xiang, W. Growth and Biochemical Composition Characteristics of Arthrospira platensis Induced by Simultaneous Nitrogen Deficiency and Seawater-Supplemented Medium in an Outdoor Raceway Pond in Winter. Foods 2021, 10, 2974. [Google Scholar] [CrossRef]
- Cao, Y.; Ma, W.; Huang, J.; Xiong, Y.L. Effects of sodium pyrophosphate coupled with catechin on the oxidative stability and gelling properties of myofibrillar protein. Food Hydrocoll. 2020, 104, 105722. [Google Scholar] [CrossRef]
- Ellman, G.L. Reprint of: Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 2022, 726, 109245. [Google Scholar] [CrossRef]
- Kantono, K.; Hamid, N.; Ma, Q.; Oey, I.; Farouk, M. Changes in the physicochemical properties of chilled and frozen-thawed lamb cuts subjected to pulsed electric field processing. Food Res. Int. 2021, 141, 110092. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Liu, R.; Wang, H.; Hua, C.; Song, S.; Zhou, G.; Zhang, W. Effects of oxidation in vitro on structures and functions of myofibrillar protein from beef muscles. J. Agric. Food Chem. 2019, 67, 5866–5873. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, D.; Xu, X.; Dai, J.; Lao, G.; Zhang, S.; Xu, X.; Dinnyés, A.; Xiong, Y.; Sun, Q. Myofibrillar protein-chlorogenic acid complexes ameliorate glucose metabolism via modulating gut microbiota in a type 2 diabetic rat model. Food Chem. 2023, 409, 135195. [Google Scholar] [CrossRef] [PubMed]
- O’Dea, E.E.; Fasenko, G.M.; Allison, G.E.; Korver, D.R.; Tannock, G.W.; Guan, L.L. Investigating the effects of commercial probiotics on broiler chick quality and production efficiency. Poult. Sci. 2006, 85, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, X.; Han, D.; Pi, Y.; Tao, S.; Zhang, S.; Wang, S.; Zhao, J.; Chen, L.; Wang, J. Early life administration of milk fat globule membrane promoted SCFA-producing bacteria colonization, intestinal barriers and growth performance of neonatal piglets. Anim. Nutr. 2021, 7, 346–355. [Google Scholar] [CrossRef]
- Peng, Q.; Zeng, X.F.; Zhu, J.L.; Wang, S.; Liu, X.T.; Hou, C.L.; Thacker, P.A.; Qiao, S.Y. Effects of dietary Lactobacillus plantarum B1 on growth performance, intestinal microbiota, and short chain fatty acid profiles in broiler chickens. Poult. Sci. 2016, 95, 893–900. [Google Scholar] [CrossRef]
- Gao, M.R.; Xu, Q.D.; Zeng, W.C. Effect of tea polyphenols on the tenderness of yak meat. J. Food Process. Preserv. 2020, 44, e14433. [Google Scholar] [CrossRef]
- Zhong, R.Z.; Li, H.Y.; Fang, Y.; Sun, H.X.; Zhou, D.W. Effects of dietary supplementation with green tea polyphenols on digestion and meat quality in lambs infected with Haemonchus contortus. Meat Sci. 2015, 105, 1–7. [Google Scholar] [CrossRef]
- Fořtová, J.; del Mar Campo, M.; Valenta, J.; Needham, T.; Řehák, D.; Lebedová, N.; Bartoň, L.; Klouček, P.; Bureš, D. Preferences and acceptance of Czech and Spanish consumers regarding beef with varying intramuscular fat content. Meat Sci. 2022, 192, 108912. [Google Scholar] [CrossRef]
- Mwangi, F.W.; Charmley, E.; Gardiner, C.P.; Malau-Aduli, B.S.; Kinobe, R.T.; Malau-Aduli, A.E.O. Diet and genetics Influence beef cattle performance and meat quality characteristics. Foods 2019, 8, 648. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Urstad, A.P.; Semenkovich, C.F. Fatty acid synthase and liver triglyceride metabolism: Housekeeper or messenger? Biochim. Biophys. Acta 2012, 1821, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.Y.; Belal, S.A.; Kang, D.R.; Choi, Y.I.; Kim, Y.H.; Choe, H.S.; Heo, J.Y.; Shim, K.S. Influence of probiotics-friendly pig production on meat quality and physicochemical characteristics. Korean J. Food Sci. Anim. Resour. 2018, 38, 403–416. [Google Scholar] [CrossRef]
- Zhu, Q.; Song, M.; Azad, M.A.K.; Ma, C.; Yin, Y.; Kong, X. Probiotics and synbiotics addition to Bama mini-pigs’ diet Improve carcass traits and meat quality by altering plasma metabolites and related gene expression of offspring. Front. Vet. Sci. 2022, 9, 779745. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Zhang, Q.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. The synergistic effect of Lactobacillus plantarum CCFM242 and zinc on ulcerative colitis through modulating intestinal homeostasis. Food Funct. 2019, 10, 6147–6156. [Google Scholar] [CrossRef]
- Tan, C.; Xu, Q.-D.; Chen, N.; He, Q.; Sun, Q.; Zeng, W.-C. Cross-linking effects of EGCG on myofibrillar protein from common carp (Cyprinus carpio) and the action mechanism. J. Food Biochem. 2022, 46, e14416. [Google Scholar] [CrossRef]
- Diao, X.; Guan, H.; Zhao, X.; Diao, X.; Kong, B. Physicochemical and structural properties of composite gels prepared with myofibrillar protein and lard diacylglycerols. Meat Sci. 2016, 121, 333–341. [Google Scholar] [CrossRef]
- Guo, A.; Jiang, J.; True, A.D.; Xiong, Y.L. Myofibrillar protein cross-linking and gelling behavior modified by structurally relevant phenolic compounds. J. Agric. Food Chem. 2021, 69, 1308–1317. [Google Scholar] [CrossRef]
- Xia, W.; Ma, L.; Chen, X.; Li, X.; Zhang, Y. Physicochemical and structural properties of composite gels prepared with myofibrillar protein and lecithin at various ionic strengths. Food Hydrocoll. 2018, 82, 135–143. [Google Scholar] [CrossRef]
- Dong, L.; Zhong, Z.X.; Cui, H.H.; Wang, S.N.; Luo, Y.; Yu, L.H.; Loor, J.J.; Wang, H.R. Effects of rumen-protected betaine supplementation on meat quality and the composition of fatty and amino acids in growing lambs. Animal 2020, 14, 435–444. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Franco, D. Fat effect on physico-chemical, microbial and textural changes through the manufactured of dry-cured foal sausage lipolysis, proteolysis and sensory properties. Meat Sci. 2012, 92, 704–714. [Google Scholar] [CrossRef]
- Zhang, X.; Han, L.; Hou, S.; Raza, S.H.A.; Gui, L.; Sun, S.; Wang, Z.; Yang, B.; Yuan, Z.; Simal-Gandara, J.; et al. Metabolomics approach reveals high energy diet improves the quality and enhances the flavor of black Tibetan sheep meat by altering the composition of rumen microbiota. Front. Nutr. 2022, 9, 915558. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Wang, Y.; Zhao, G.; Wen, J.; Cui, H. Metabolomics-Based Analysis of the Major Taste Contributors of Meat by Comparing Differences in Muscle Tissue between Chickens and Common Livestock Species. Foods 2022, 11, 3586. [Google Scholar] [CrossRef]
- Dragan, M.; Nguyen, M.U.; Guzman, S.; Goertzen, C.; Brackstone, M.; Dhillo, W.S.; Bech, P.R.; Clarke, S.; Abbara, A.; Tuck, A.B.; et al. G protein-coupled kisspeptin receptor induces metabolic reprograming and tumorigenesis in estrogen receptor-negative breast cancer. Cell Death Dis. 2020, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Dashdorj, D.; Amna, T.; Hwang, I. Influence of specific taste-active components on meat flavor as affected by intrinsic and extrinsic factors: An overview. Eur. Food Res. Technol. 2015, 241, 157–171. [Google Scholar] [CrossRef]
- Yin, H.D.; Gilbert, E.R.; Chen, S.Y.; Wang, Y.; Zhang, Z.C.; Zhao, X.L.; Zhang, Y.; Zhu, Q. Effect of hybridization on carcass traits and meat quality of erlang mountainous chickens. Asian-Australas. J. Anim. Sci. 2013, 26, 1504–1510. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, H.; Liu, M.; Zhao, X.; Luo, H. Effect of Breed on the Volatile Compound Precursors and Odor Profile Attributes of Lamb Meat. Foods 2020, 9, 1178. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, Y.; Tang, C.; Yue, S.; Zhao, Q.; Li, F.; Zhang, J. Changes in lipids and aroma compounds in intramuscular fat from Hu sheep. Food Chem. 2022, 383, 132611. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, T.; Zhang, Y.; Yang, L.; Duan, Y.; Lin, S.; Tian, J.; Sun, L.; Wang, B.; Ye, J. Impact of feeding probiotics on blood parameters, tail fat metabolites, and volatile flavor components of Sunit sheep. Foods 2022, 11, 2644. [Google Scholar] [CrossRef]
- Estevez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Guo, A.; Xiong, Y.L. Glucose oxidase promotes gallic acid-myofibrillar protein interaction and thermal gelation. Food Chem. 2019, 293, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Zhang, D.; Wen, X.; Li, X.; Chen, L.; Zheng, X.; Fang, F.; Li, J.; Hou, C. Effects of chilling rate on the freshness and microbial community composition of lamb carcasses. Lwt 2022, 153, 112559. [Google Scholar] [CrossRef]
- Tang, W.; Xing, Z.; Li, C.; Wang, J.; Wang, Y. Molecular mechanisms and in vitro antioxidant effects of Lactobacillus plantarum MA2. Food Chem. 2017, 221, 1642–1649. [Google Scholar] [CrossRef]
- Chen, H.; Peng, T.; Shang, H.; Shang, X.; Zhao, X.; Qu, M.; Song, X. RNA-Seq analysis reveals the potential molecular mechanisms of puerarin on intramuscular fat deposition in heat-stressed beef cattle. Front. Nutr. 2022, 9, 817557. [Google Scholar] [CrossRef] [PubMed]
- Sudun; Liu, S.; Xiao, C.; Peng, C.; Liang, L.; He, X.; Zhao, S.; Zhang, G. Probiotic strains improve high-fat diet-induced hypercholesterolemia through modulating gut microbiota in ways different from atorvastatin. Food Funct. 2019, 10, 6098–6109. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | Probiotics | Synbiotics | SEM | p-Value |
|---|---|---|---|---|---|
| Initial body wight (kg) | 4.36 | 4.83 | 4.95 | 0.200 | 0.479 |
| Final body wight (kg) | 13.46 b | 15.15 ab | 18.39 a | 0.962 | 0.093 |
| Initial body length (cm) | 33.50 | 35.00 | 35.00 | 0.748 | 0.683 |
| Final body length (cm) | 53.45 b | 55.97 ab | 59.20 a | 1.093 | 0.085 |
| Initial body height (cm) | 38.08 | 39.45 | 38.75 | 0.588 | 0.678 |
| Final body height (cm) | 51.90 b | 53.80 ab | 57.95 a | 0.874 | 0.003 |
| Initial chest circumference (cm) | 39.48 | 39.18 | 41.96 | 0.872 | 0.387 |
| Final chest circumference (cm) | 55.82 b | 58.20 ab | 64.00 a | 1.430 | 0.027 |
| Average daily gain (g/d) | 151.71 | 171.92 | 223.85 | 14.933 | 0.119 |
| Carcass weight (kg) | 7.18 | 7.60 | 9.86 | 0.542 | 0.079 |
| Slaughter rate (%) | 53.20 | 50.26 | 53.58 | 0.660 | 0.063 |
| Muscle Part | Parameters | Control | Probiotics | Synbiotics | SEM | p-Value |
|---|---|---|---|---|---|---|
| Longissimus thoracis | pH1h | 6.49 | 6.47 | 6.26 | 0.055 | 0.157 |
| pH24h | 5.60 | 5.52 | 5.63 | 0.031 | 0.334 | |
| L1h | 45.04 | 42.87 | 43.01 | 0.627 | 0.304 | |
| a1h | 7.22 ab | 6.10 b | 8.12 a | 0.347 | 0.045 | |
| b1h | 11.07 | 9.99 | 10.44 | 0.237 | 0.180 | |
| L24h | 37.42 | 37.60 | 35.03 | 1.307 | 0.701 | |
| a24h | 5.58 | 4.39 | 5.46 | 0.346 | 0.322 | |
| b24h | 9.27 | 7.94 | 8.44 | 0.446 | 0.504 | |
| Cooking loss (%) | 41.34 | 42.65 | 41.74 | 0.346 | 0.339 | |
| Dripping loss (%) | 5.66 | 5.38 | 5.88 | 0.529 | 0.939 | |
| Water-holding capacity (%) | 95.85 | 95.57 | 95.29 | 0.664 | 0.950 | |
| Biceps brachii | pH1h | 6.35 | 6.48 | 6.48 | 0.034 | 0.234 |
| pH24h | 5.59 | 5.58 | 5.58 | 0.014 | 0.945 | |
| L1h | 48.75 | 49.19 | 45.27 | 0.851 | 0.114 | |
| a1h | 6.90 | 5.72 | 7.86 | 0.611 | 0.386 | |
| b1h | 11.66 | 12.01 | 11.37 | 0.380 | 0.812 | |
| L24h | 36.72 | 39.31 | 41.17 | 1.106 | 0.273 | |
| a24h | 5.32 | 5.48 | 6.70 | 0.385 | 0.300 | |
| b24h | 9.05 b | 10.27 ab | 11.11 a | 0.367 | 0.059 | |
| Cooking loss (%) | 41.82 | 42.38 | 41.95 | 0.577 | 0.931 | |
| Dripping loss (%) | 4.13 | 4.62 | 3.54 | 0.264 | 0.297 | |
| Water-holding capacity (%) | 96.95 | 95.78 | 96.97 | 0.350 | 0.303 |
| Parameters (μg/g) | Control | Probiotics | Synbiotics | SEM | p-Value |
|---|---|---|---|---|---|
| K | 5038.10 a | 4321.66 b | 4830.10 ab | 108.635 | 0.009 |
| Ca | 37.68 | 42.28 | 46.54 | 2.724 | 0.446 |
| Na | 631.10 | 660.54 | 559.42 | 21.426 | 0.139 |
| Mg | 246.80 | 248.10 | 256.44 | 3.063 | 0.408 |
| Cu | 1.16 | 1.10 | 1.40 | 0.086 | 0.348 |
| Fe | 13.66 | 16.20 | 13.54 | 1.285 | 0.668 |
| Zn | 25.78 b | 27.32 ab | 33.06 a | 1.442 | 0.085 |
| Cr | ND | ND | ND | ||
| Mn | ND | ND | ND | ||
| Se | ND | ND | ND | ||
| Moisture content (%) | 75.30 | 76.15 | 75.78 | 0.347 | 0.443 |
| Crude fat content (%) | 3.32 b | 4.16 b | 5.00 a | 0.303 | 0.065 |
| Crude protein content (%) | 18.16 | 18.14 | 18.74 | 0.223 | 0.493 |
| Ash content (%) | 1.08 | 1.02 | 1.06 | 0.016 | 0.344 |
| Muscle Part | Parameters | Control | Probiotics | Synbiotics | SEM | p-Value |
|---|---|---|---|---|---|---|
| Longissimus thoracis | Hardness (g) | 2013.48 b | 3199.24 a | 2158.32 b | 142.476 | <0.001 |
| Springiness | 0.777 a | 0.771 ab | 0.721 b | 0.010 | 0.035 | |
| Cohesiveness | 0.644 | 0.639 | 0.619 | 0.006 | 0.240 | |
| Gumminess | 1422.99 b | 2035.86 a | 1340.55 b | 86.333 | <0.001 | |
| Chewiness | 990.64 b | 1575.67 a | 982.36 b | 75.694 | <0.001 | |
| Resilience | 0.247 a | 0.229 ab | 0.218 b | 0.005 | 0.046 | |
| Shear force (N) | 114.17 a | 108.38 a | 90.63 b | 4.019 | 0.031 | |
| Biceps brachii | Hardness (g) | 1704.82 | 1891.09 | 1761.08 | 84.076 | 0.666 |
| Springiness | 0.799 a | 0.695 b | 0.777 a | 0.011 | <0.001 | |
| Cohesiveness | 0.663 | 0.684 | 0.657 | 0.008 | 0.370 | |
| Gumminess | 1121.83 | 1291.87 | 1149.06 | 54.398 | 0.405 | |
| Chewiness | 897.08 | 892.21 | 889.49 | 38.186 | 0.997 | |
| Resilience | 0.287 | 0.281 | 0.267 | 0.006 | 0.365 | |
| Shear force (N) | 122.01 | 124.17 | 114.99 | 3.655 | 0.591 |
| Parameters (g/100 g) | Control | Probiotics | Synbiotics | SEM | p-Value |
|---|---|---|---|---|---|
| Asp | 1.764 b | 1.782 ab | 1.806 a | 0.007 | 0.029 |
| Thr | 0.896 b | 0.903 ab | 0.924 a | 0.006 | 0.043 |
| Ser | 0.763 b | 0.764 b | 0.794 a | 0.006 | 0.015 |
| Glu | 3.059 | 3.114 | 3.123 | 0.017 | 0.306 |
| Gly | 0.802 | 0.810 | 0.827 | 0.016 | 0.613 |
| Ala | 1.099 | 1.104 | 1.119 | 0.005 | 0.307 |
| Val | 0.929 | 0.938 | 0.946 | 0.003 | 0.100 |
| Ile | 0.867 | 0.887 | 0.888 | 0.004 | 0.046 |
| Leu | 1.562 | 1.572 | 1.592 | 0.006 | 0.070 |
| Tyr | 0.722 b | 0.727 b | 0.751 a | 0.005 | 0.016 |
| Phe | 0.820 b | 0.838 a | 0.843 a | 0.004 | 0.003 |
| Lys | 1.739 | 1.761 | 1.772 | 0.007 | 0.144 |
| His | 0.628 | 0.668 | 0.645 | 0.011 | 0.413 |
| Arg | 1.289 | 1.278 | 1.295 | 0.005 | 0.426 |
| Pro | 0.685 | 0.676 | 0.692 | 0.005 | 0.443 |
| Met | 0.521 b | 0.537 a | 0.537 a | 0.003 | 0.028 |
| FAA | 8.534 | 8.625 | 8.706 | 0.038 | 0.196 |
| TEAA | 7.333 b | 7.435 ab | 7.503 a | 0.030 | 0.039 |
| TNEAA | 10.810 | 10.923 | 11.051 | 0.046 | 0.084 |
| TAA | 18.142 b | 18.357 ab | 18.554 a | 0.075 | 0.050 |
| Species | Volatile Compounds | Relative Abundance (%) | SEM | p-Value | Odorant Descriptors | ||
|---|---|---|---|---|---|---|---|
| Con | Pro | Syn | |||||
| Aldehydes | Hexanal | 4.41 | 5.82 | 6.19 | 0.488 | 0.332 | Herbal, grassy |
| 2-Hexenal | ND | 0.35 | ND | Green apple-like, bitter almond-like | |||
| Heptanal | 2.31 | 3.61 | 2.34 | 0.365 | 0.281 | Jasmine, mint, burnt fat, green | |
| Octanal | 9.02 | 3.43 | 13.36 | 3.355 | 0.545 | Citrus, floral | |
| (E)-2-Octenal | 1.43 | 1.82 | 0.64 | 0.223 | 0.062 | Fatty, wet ground, grass, coffee | |
| (E)-2-Decenal | 1.19 | 1.23 | 0.76 | 0.906 | 0.025 | Chicken fat, fried notes, waxy | |
| (Z)-4-Decenal | 0.57 | 0.22 | 0.66 | 0.124 | 0.354 | Orange, chicken fat | |
| (E, E)-2,4-Decadienal | 0.37 | 0.62 | 0.41 | 0.057 | 0.169 | Chicken fat, poultry | |
| (Z)-2-Nonenal | 1.64 | 2.05 | 1.92 | 0.182 | 0.704 | Potato peel | |
| Undecanal | 0.10 | 0.19 | 1.73 | 0.567 | 0.482 | Grassy, rain, dirt | |
| (E)-2-Undecenal | 1.02 | 1.30 | 1.16 | 0.170 | 0.833 | Waxy, fatty | |
| Dodecanal | 9.20 | 8.10 | 6.51 | 1.198 | 0.714 | Onion, green, yeast, vomit | |
| (E, E)-2,4-Dodecadienal | 0.28 | 0.15 | 0.09 | 0.099 | 0.778 | ||
| Hexadecanal | 18.79 | 23.54 | 21.24 | 2.118 | 0.718 | Sweet | |
| E-9-Tetradecenal | 0.22 | 0.42 | 0.61 | 0.088 | 0.215 | ||
| 9-Octadecenal | 0.20 | ND | 0.43 | 0.086 | 0.109 | ||
| (E)-1,4-Undecadiene | 0.79 | 0.38 | 0.23 | 0.159 | 0.387 | ||
| Benzaldehyde | 1.97 | 0.86 | 3.00 | 0.420 | 0.096 | Almond, caramel, nutty | |
| 4-Pentylbenzaldehyde | 0.57 | 0.49 | 0.47 | 0.031 | 0.414 | ||
| 2-Methylundecane | 0.27 | 0.16 | 0.43 | 0.100 | 0.600 | ||
| Alcohols | 2-Nonen-1-ol | 35.61 | 35.11 | 25.71 | 2.113 | 0.074 | |
| Esters | Sulfurous acid, decyl 2-propyl ester | 0.75 | 0.22 | 0.27 | 0.174 | 0.449 | |
| Isooctylvinyl ether | ND | 0.15 | 0.46 | 0.082 | 0.026 | ||
| Alkanes | 2-Methyldecane | ND | 0.78 | 0.74 | 0.218 | 0.291 | |
| 3-Methyltridecane | 0.13 | 0.59 | ND | 0.097 | 0.004 | ||
| 4,8-Dimethyltridecane | 1.12 | 3.05 | 0.47 | 1.008 | 0.620 | ||
| 6-Propyltridecane | 0.69 | 5.10 | 4.81 | 1.094 | 0.191 | ||
| Octadecane | ND | 0.21 | ND | ||||
| 2-Methyltricosane | 0.38 | ND | 0.21 | 0.090 | 0.237 | ||
| Ketones | 2,3-Octanedione | 3.12 | 0.62 | 1.99 | 0.762 | 0.467 | |
| Alkenes | 1,3-Octadiene | 1.88 | 1.13 | 4.09 | 0.913 | 0.444 | |
| 2,4-Octadiene | 1.38 | ND | 0.79 | 0.501 | 0.595 | ||
| (Z, Z)-3,5-Octadiene | 0.51 | 0.48 | 2.18 | 0.354 | 0.050 | ||
| Heterocyclics | 2-Pentylfuran | 0.20 | ND | 0.62 | 0.142 | 0.208 | Roast, buttery |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Z.; Xu, X.; Luo, D.; Zhou, Z.; Zhang, S.; He, R.; An, T.; Sun, Q. Effect of Dietary Supplementation of Lactiplantibacillus plantarum N-1 and Its Synergies with Oligomeric Isomaltose on the Growth Performance and Meat Quality in Hu Sheep. Foods 2023, 12, 1858. https://doi.org/10.3390/foods12091858
Zhou Z, Xu X, Luo D, Zhou Z, Zhang S, He R, An T, Sun Q. Effect of Dietary Supplementation of Lactiplantibacillus plantarum N-1 and Its Synergies with Oligomeric Isomaltose on the Growth Performance and Meat Quality in Hu Sheep. Foods. 2023; 12(9):1858. https://doi.org/10.3390/foods12091858
Chicago/Turabian StyleZhou, Zhiqiang, Xinyi Xu, Dongmei Luo, Zhiwei Zhou, Senlin Zhang, Ruipeng He, Tianwu An, and Qun Sun. 2023. "Effect of Dietary Supplementation of Lactiplantibacillus plantarum N-1 and Its Synergies with Oligomeric Isomaltose on the Growth Performance and Meat Quality in Hu Sheep" Foods 12, no. 9: 1858. https://doi.org/10.3390/foods12091858