


Unraveling the Mechanism of StWRKY6 in Potato (Solanum tuberosum)’s Cadmium Tolerance for Ensuring Food Safety

 ,
,
Abstract
:1. Introduction
2. Results
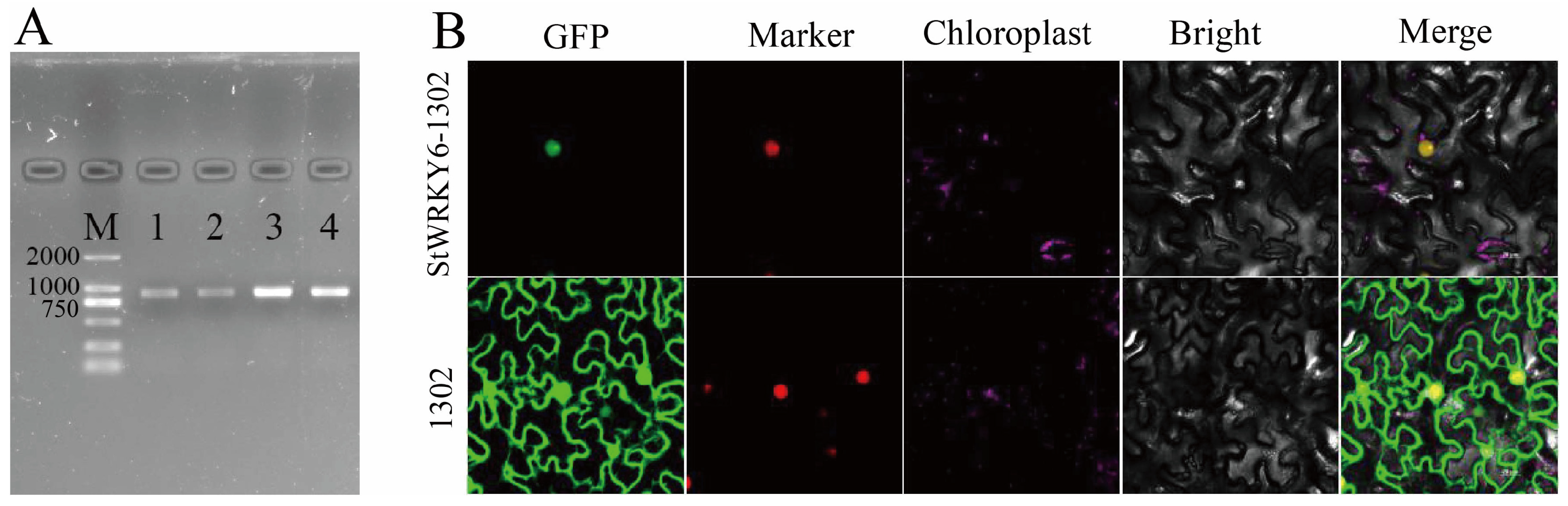
2.1. Subcellular Localization of StWRKY6 in Tobacco Leaves
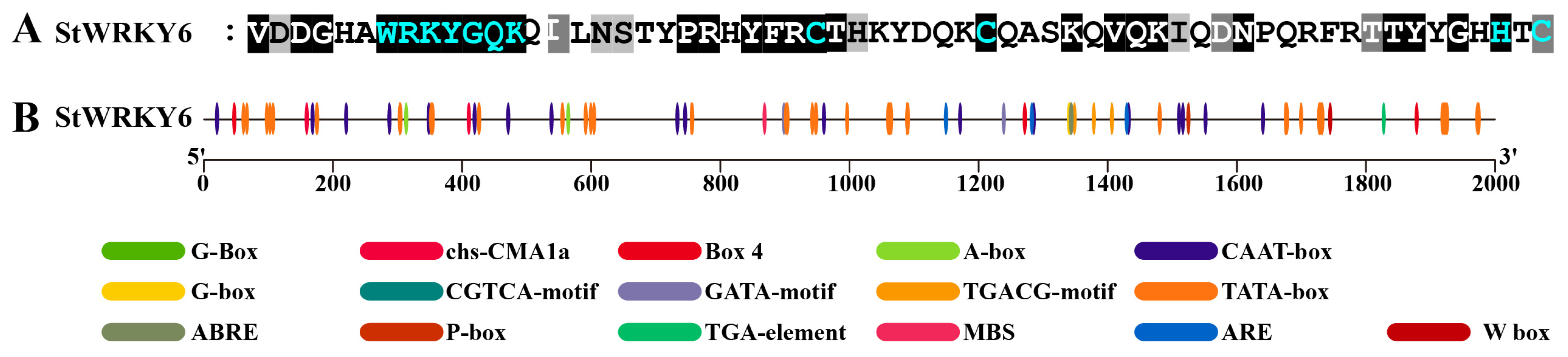
2.2. StWRKY6 Protein Sequence and Cis-Acting Element Analysis
2.3. Protein Interaction Analysis Revealed the Role of StWRKY6 in Potato Stress Adaptation
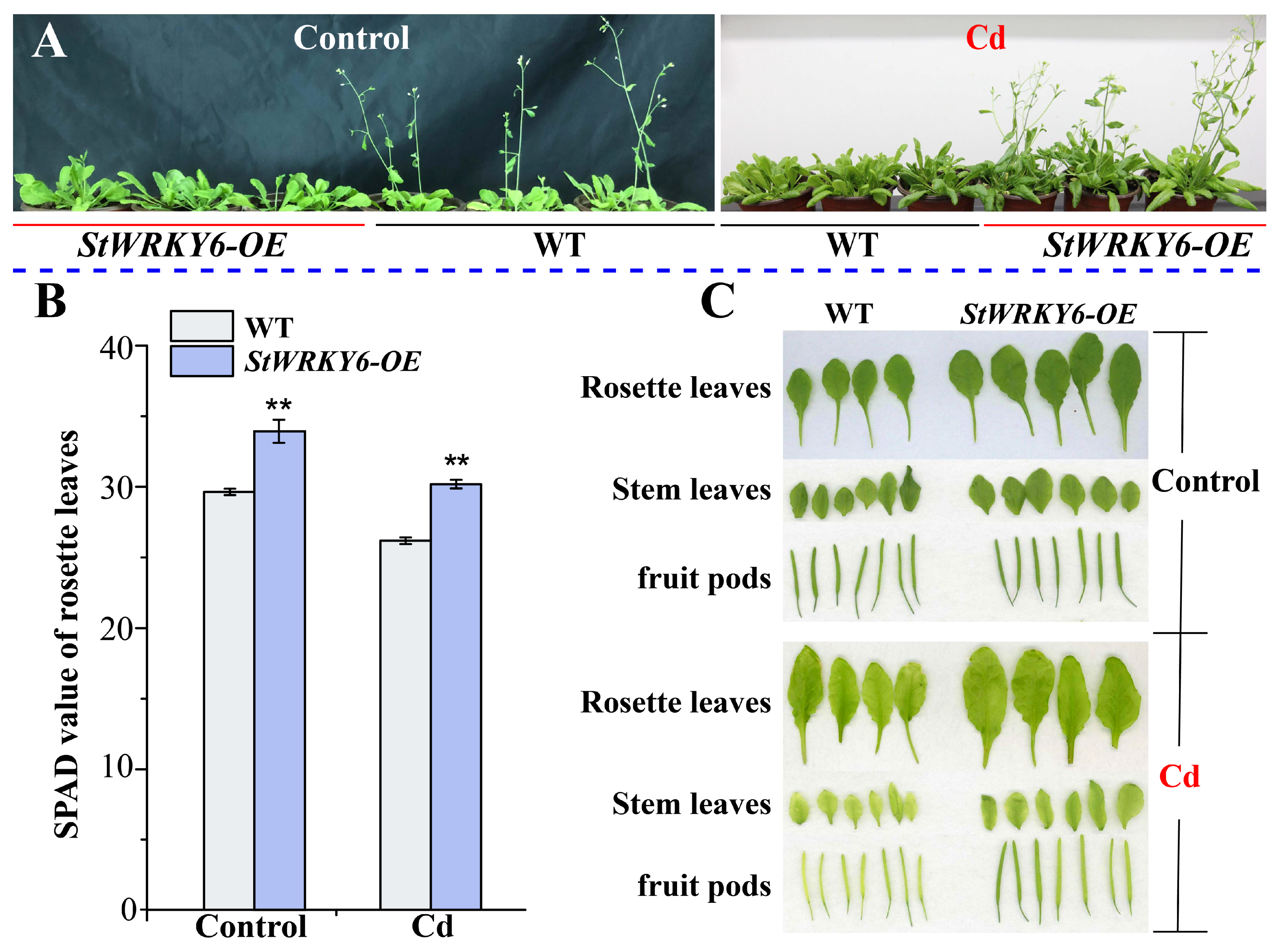
2.4. Improving Photosynthesis and Reducing Cd Toxicity-Induced Chlorosis through StWRKY6 Overexpression
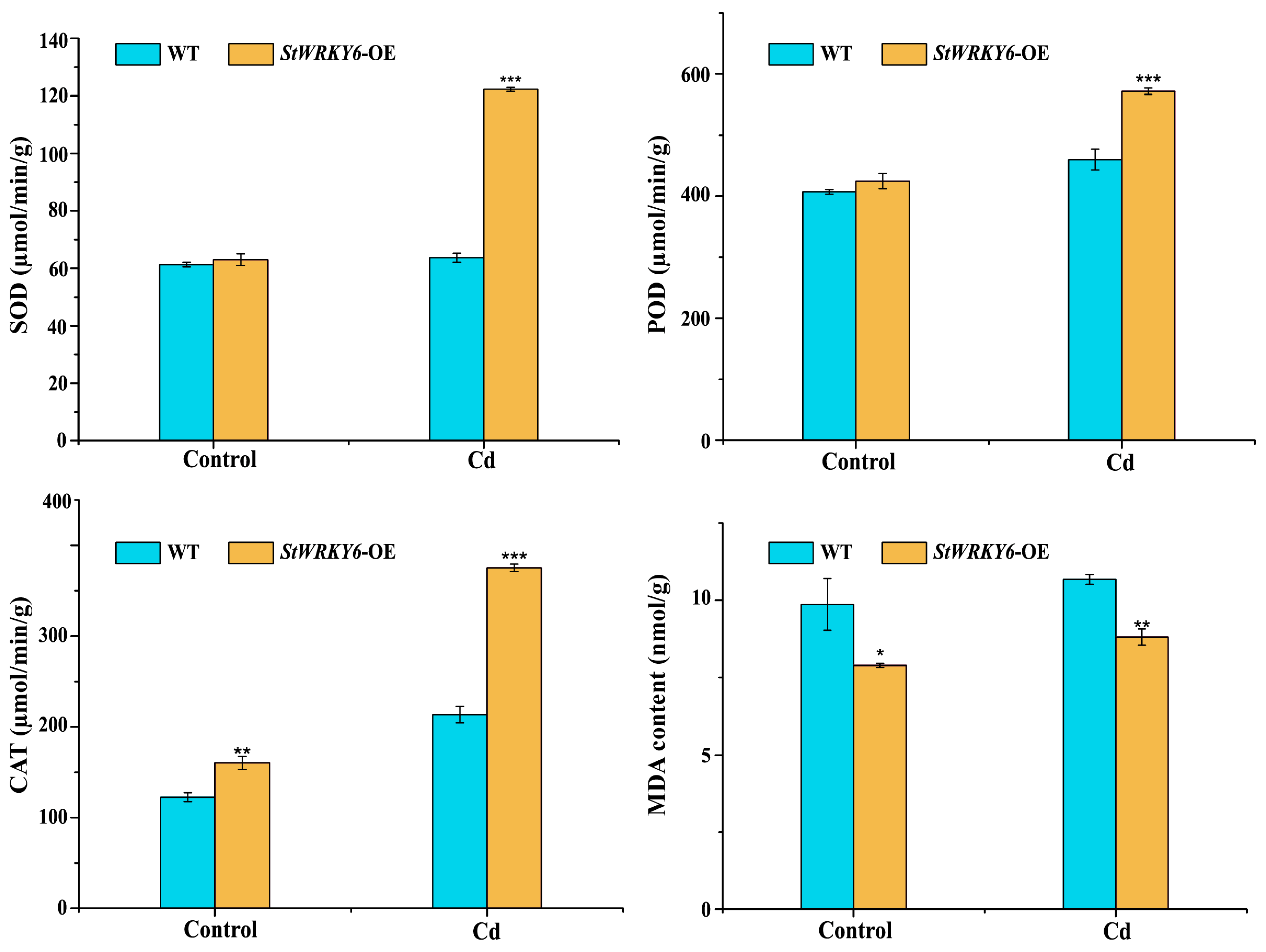
2.5. Enhanced Protective Enzyme Activity and Reduced MDA Content Contribute to StWRKY6-Mediated Cd Tolerance in Plants
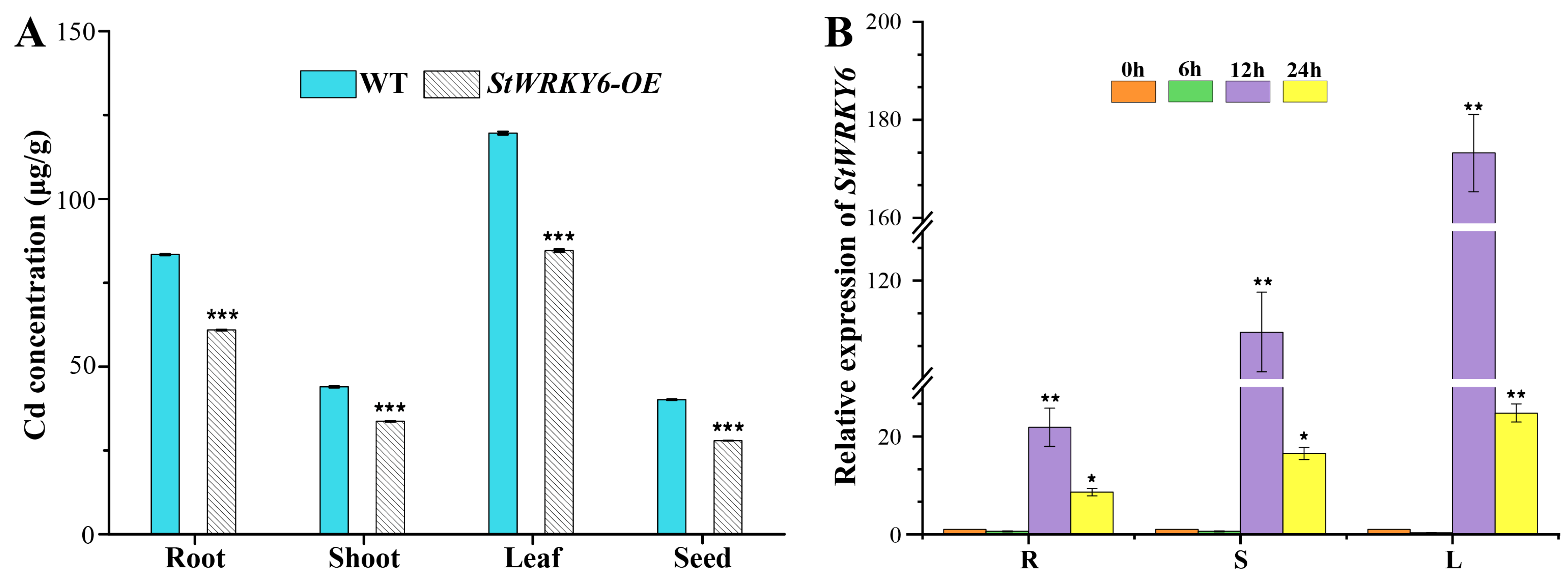
2.6. Transcriptional Regulation by StWRKY6 Reduces Cd Accumulation and Enhances Plant Tolerance
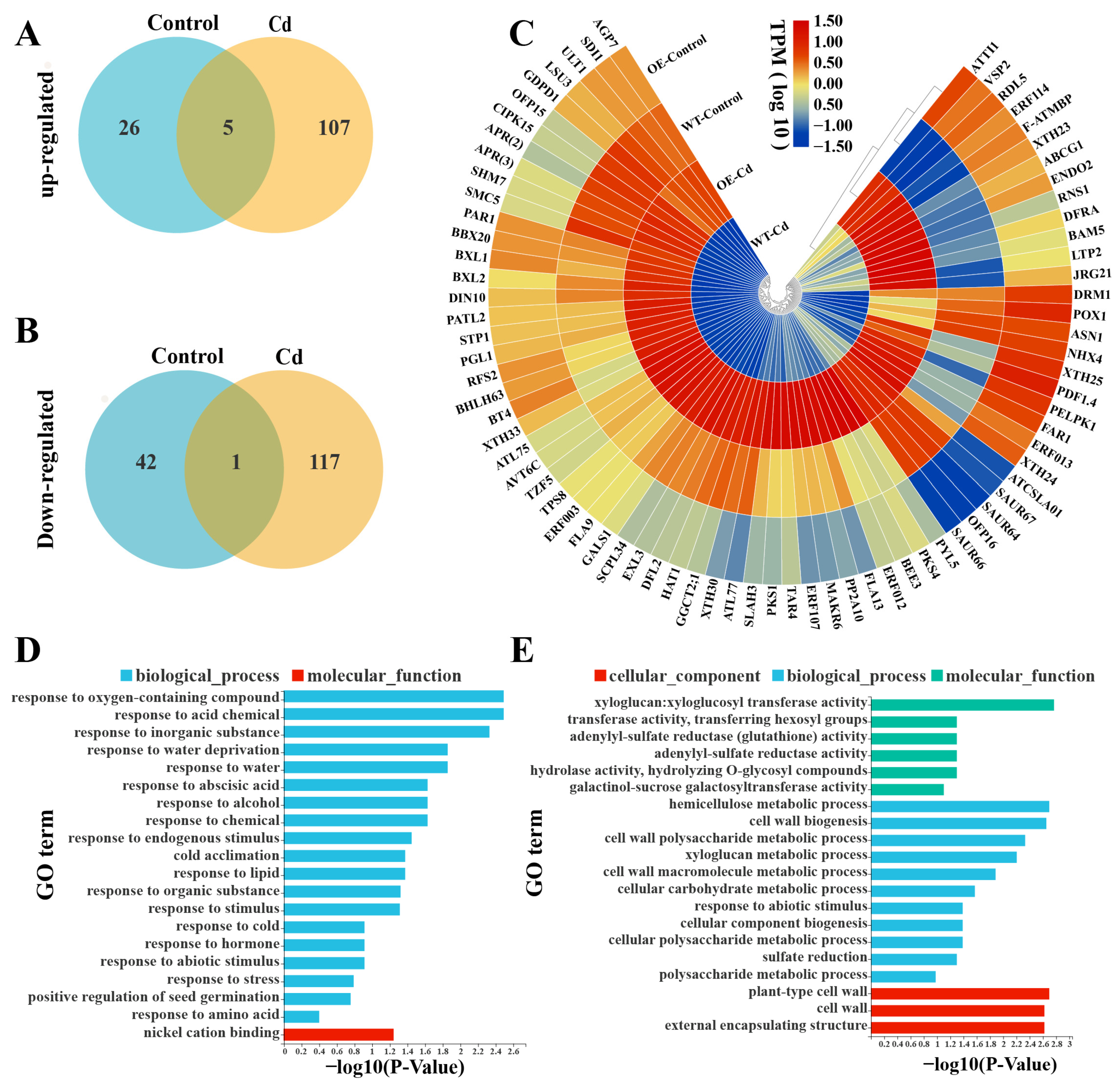
2.7. Elucidating the StWRKY6-Mediated Cd Detoxification Pathway in Transgenic Arabidopsis through RNA-seq and GO Enrichment Analysis
3. Discussion
3.1. StWRKY6 Regulates Cd Tolerance in Arabidopsis
3.2. StWRKY6 Reduces Cadmium-Induced Oxidative Stress by Enhancing Antioxidant Enzyme Activity
3.3. Alleviating Cd Toxicity in StWRKY6 Transgenic Arabidopsis by Regulating Photosynthesis
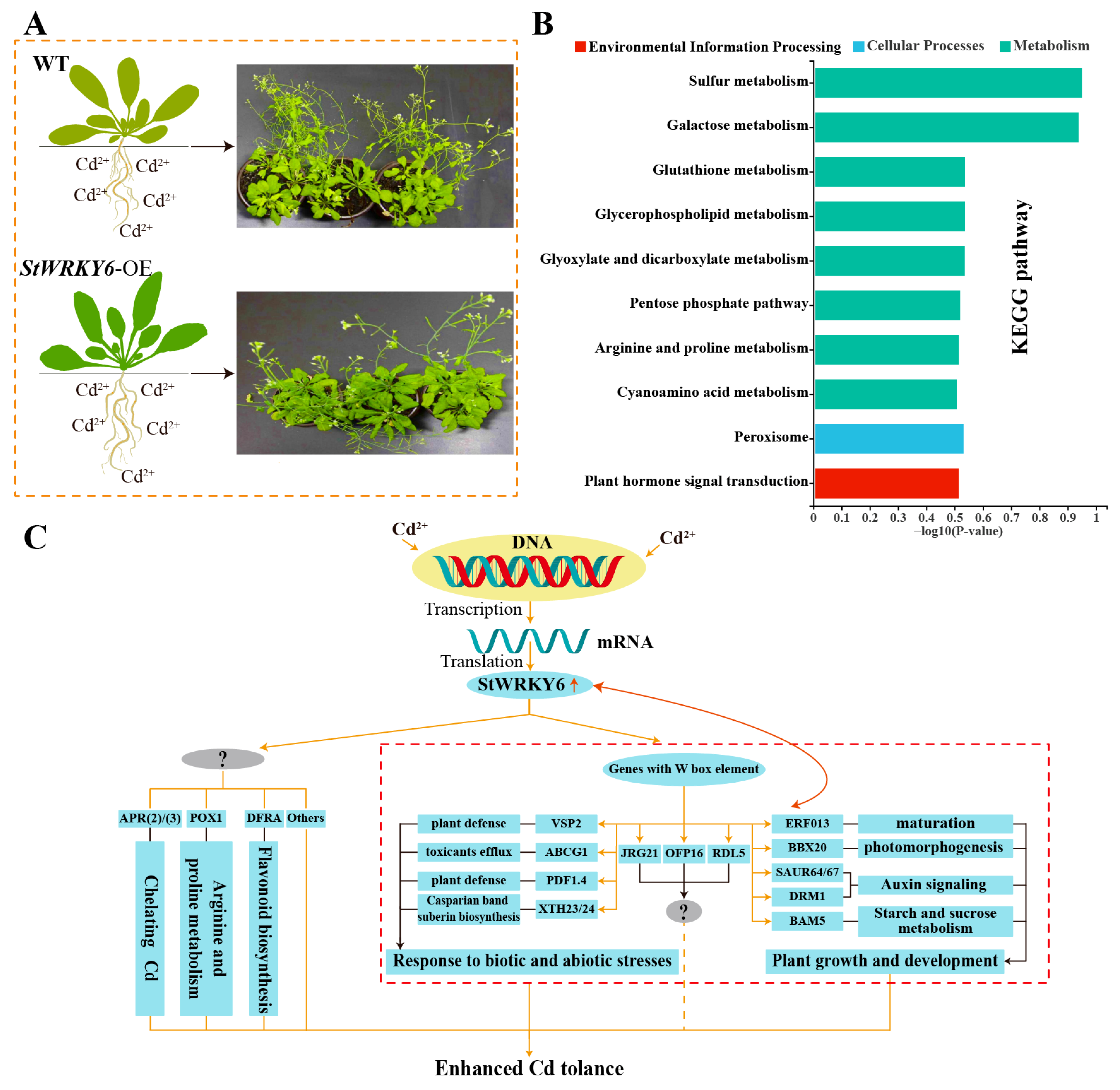
3.4. StWRKY6 as a Gene Regulatory Target to Enhance Plant Heavy Metal Tolerance
4. Conclusions
5. Materials and Methods
5.1. Plant Material, Treatment, and Analysis of Cd Accumulation and Enzyme Activity
5.2. Cloning and Subcellular Localization of StWRKY6
5.3. Sequence and Cis-Acting Element Analysis of StWRKY6
5.4. Interaction Proteins Analysis of StWRKY6
5.5. Plant Phenotypic Analysis between Treatments
5.6. RNA Extraction and RT-qPCR Analysis
5.7. Analysis of RNA Sequencing Data
5.8. Subsection
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, J.; Ma, S.; Ye, N.; Jiang, M.; Cao, J.; Zhang, J. WRKY transcription factors in plant responses to stresses. J. Integr. Plant Biol. 2017, 59, 86–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. WRKY transcription factors and plant defense responses: Latest discoveries and future prospects. Plant Cell Rep. 2021, 40, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D.; et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; He, G.; He, Y.; Tang, Y.; Zhao, F.; He, T. Discovery of cadmium-tolerant biomacromolecule (StCAX1/4 transportproteins) in potato and its potential regulatory relationship with WRKY transcription factors. Int. J. Biol. Macromol. 2023, 228, 385–399. [Google Scholar] [CrossRef]
- He, Y.; He, G.; Lou, F.; Zhou, Z.; Liu, Y.; Zhang, Y.; He, T. Identification of the Major Effector StSROs in Potato: A Potential StWRKY-SRO6 Regulatory Pathway Enhances Plant Tolerance to Cadmium Stress. Int. J. Mol Sci. 2022, 23, 14318. [Google Scholar] [CrossRef]
- Li, Y.; Ding, L.; Zhou, M.; Chen, Z.; Ding, Y.; Zhu, C. Transcriptional Regulatory Network of Plant Cadmium Stress Response. Int. J. Mol. Sci. 2023, 24, 4378. [Google Scholar] [CrossRef]
- Wang, H.; Zou, S.; Li, Y.; Lin, F.; Tang, D. An ankyrin-repeat and WRKY-domain-containing immune receptor confers stripe rust resistance in wheat. Nat. Commun. 2020, 11, 1353. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, D.; Yang, C.; Kong, N.; Shi, Z.; Zhao, P.; Nan, Y.; Nie, T.; Wang, R.; Ma, H.; et al. Genome-wide identification of the potato WRKY transcription factor family. PLoS ONE 2017, 12, e181573. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, P.; Mitra, M.; Banerjee, S.; Roy, S. MYB4 transcription factor, a member of R2R3-subfamily of MYB domain protein, regulates cadmium tolerance via enhanced protection against oxidative damage and increases expression of PCS1 and MT1C in Arabidopsis. Plant Sci. 2020, 297, 110501. [Google Scholar] [CrossRef]
- Himeno, S.; Sumi, D.; Fujishiro, H. Toxicometallomics of Cadmium, Manganese and Arsenic with Special Reference to the Roles of Metal Transporters. Toxicol. Res. 2019, 35, 311–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, W.; Huang, Y.; Li, D.; Meng, L.; He, T.; He, G. Identification of StAP2/ERF genes of potato (Solanum tuberosum) and their multiple functions in detoxification and accumulation of cadmium in yest: Implication for Genetic-based phytoremediation. Sci. Total Environ. 2022, 810, 152322. [Google Scholar] [CrossRef] [PubMed]
- Aprile, A.; Sabella, E.; Francia, E.; Milc, J.; Ronga, D.; Pecchioni, N.; Ferrari, E.; Luvisi, A.; Vergine, M.; De Bellis, L. Combined Effect of Cadmium and Lead on Durum Wheat. Int. J. Mol. Sci. 2019, 20, 5891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Fan, T.; Zhu, X.; Wu, X.; Ouyang, J.; Jiang, L.; Cao, S. WRKY12 represses GSH1 expression to negatively regulate cadmium tolerance in Arabidopsis. Plant Mol. Biol. 2019, 99, 149–159. [Google Scholar] [CrossRef]
- Wu, D.; Saleem, M.; He, T.; He, G. The Mechanism of Metal Homeostasis in Plants: A New View on the Synergistic Regulation Pathway of Membrane Proteins, Lipids and Metal Ions. Membranes 2021, 11, 984. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; He, T.; Saleem, M.; He, G. Metalloprotein-Specific or Critical Amino Acid Residues: Perspectives on Plant-Precise Detoxification and Recognition Mechanisms under Cadmium Stress. Int. J. Mol. Sci. 2022, 23, 1734. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, M.; Xu, D.; Xia, Z. The Arabidopsis APR2 positively regulates cadmium tolerance through glutathione-dependent pathway. Ecotoxicol. Environ. Saf. 2020, 187, 109819. [Google Scholar] [CrossRef]
- Wątły, J.; Łuczkowski, M.; Padjasek, M.; Krężel, A. Phytochelatins as a Dynamic System for Cd(II) Buffering from the Micro- to Femtomolar Range. Inorg. Chem. 2021, 60, 4657–4675. [Google Scholar] [CrossRef]
- Song, W.Y.; Martinoia, E.; Lee, J.; Kim, D.; Kim, D.Y.; Vogt, E.; Shim, D.; Choi, K.S.; Hwang, I.; Lee, Y. A novel family of cys-rich membrane proteins mediates cadmium resistance in Arabidopsis. Plant Physiol. 2004, 135, 1027–1039. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Feng, S.J.; Chen, J.; Zhao, W.T.; Yang, Z.M. A cadmium stress-responsive gene AtFC1 confers plant tolerance to cadmium toxicity. BMC Plant Biol. 2017, 17, 187. [Google Scholar] [CrossRef] [Green Version]
- He, G.; Tian, W.; Qin, L.; Meng, L.; Wu, D.; Huang, Y.; Li, D.; Zhao, D.; He, T. Identification of novel heavy metal detoxification proteins in Solanum tuberosum: Insights to improve food security protection from metal ion stress. Sci. Total Environ. 2021, 779, 146197. [Google Scholar] [CrossRef]
- Podolec, R.; Wagnon, T.B.; Leonardelli, M.; Johansson, H.; Ulm, R. Arabidopsis B-box transcription factors BBX20-22 promote UVR8 photoreceptor-mediated UV-B responses. Plant J. 2022, 111, 422–439. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Klejchová, M.; Snipes, S.A.; Nagpal, P.; Bak, G.; Wang, B.; Dunlap, S.; Park, M.Y.; Kunkel, E.N.; Trinidad, B.; et al. SAUR proteins and PP2C.D phosphatases regulate H+-ATPases and K+ channels to control stomatal movements. Plant Physiol. 2021, 185, 256–273. [Google Scholar] [CrossRef] [PubMed]
- Holland, C.K.; Jez, J.M. Arabidopsis: The original plant chassis organism. Plant Cell Rep. 2018, 37, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Amos, R.A.; Mohnen, D. Critical Review of Plant Cell Wall Matrix Polysaccharide Glycosyltransferase Activities Verified by Heterologous Protein Expression. Front. Plant Sci. 2019, 10, 915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodward, A.W.; Bartel, B. Biology in Bloom: A Primer on the Arabidopsis thaliana Model System. Genetics 2018, 208, 1337–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, K.; Kohli, S.K.; Ohri, P.; Bhardwaj, R.; Ahmad, P. Agroecotoxicological Aspect of Cd in Soil-Plant System: Uptake, Translocation and Amelioration Strategies. Environ. Sci. Pollut. Res. Int. 2022, 29, 30908–30934. [Google Scholar] [CrossRef]
- Dang, F.; Li, Y.; Wang, Y.; Lin, J.; Du, S.; Liao, X. ZAT10 plays dual roles in cadmium uptake and detoxification in Arabidopsis. Front. Plant Sci. 2022, 13, 994100. [Google Scholar] [CrossRef]
- Cuypers, A.; Vanbuel, I.; Iven, V.; Kunnen, K.; Vandionant, S.; Huybrechts, M.; Hendrix, S. Cadmium-induced oxidative stress responses and acclimation in plants require fine-tuning of redox biology at subcellular level. Free Radic. Biol. Med. 2023, 199, 81–96. [Google Scholar] [CrossRef]
- Modareszadeh, M.; Bahmani, R.; Kim, D.; Hwang, S. CAX3 (cation/proton exchanger) mediates a Cd tolerance by decreasing ROS through Ca elevation in Arabidopsis. Plant Mol. Biol. 2021, 105, 115–132. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Jiang, W.; Wang, X.; Hussain, S.; Ahmad, M.; Maqsood, M.F.; Ali, N.; Ishfaq, M.; Kaleem, M.; Haider, F.U.; et al. Cadmium Phytotoxicity, Tolerance, and Advanced Remediation Approaches in Agricultural Soils; A Comprehensive Review. Front. Plant Sci. 2022, 13, 773815. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, H.; Jiang, H.; Wang, H.; Chen, K.; Duan, J.; Feng, S.; Wu, G. Regulation of cadmium tolerance and accumulation by miR156 in Arabidopsis. Chemosphere 2020, 242, 125168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, R.; Ju, Q.; Li, W.; Tran, L.P.; Xu, J. The R2R3-MYB Transcription Factor MYB49 Regulates Cadmium Accumulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Lou, F.; Fu, T.; He, G.; Tian, W.; Wen, J.; Yang, M.; Wei, X.; He, Y.; He, T. Different composites inhibit Cd accumulation in grains under the rice-oilseed rape rotation mode of karst area: A field study. Ecotoxicol. Environ. Saf. 2023, 256, 114884. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2021, 49, D10–D17. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Wu, D.; He, G.; Tian, W.; Saleem, M.; Li, D.; Huang, Y.; Meng, L.; He, Y.; Liu, Y.; He, T. OPT gene family analysis of potato (Solanum tuberosum) responding to heavy metal stress: Comparative omics and co-expression networks revealed the underlying core templates and specific response patterns. Int. J. Biol. Macromol. 2021, 188, 892–903. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Krishnakumar, V.; Chan, A.P.; Thibaud-Nissen, F.; Schobel, S.; Town, C.D. Araport11: A complete reannotation of the Arabidopsis thaliana reference genome. Plant J. 2017, 89, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Reverse Primer (5′–3′) | Forward Primer (5′–3′) |
|---|---|---|
| StWRKY6 (qRT-PCR) | CTTGACCTCAACCGGCGTAGTG | AACCTCTTGTTGGTGGTGGTGAAG |
| StWRKY6 (1302) | agttcttctcctttactagtCACTTGATCAAAATTCCAAAGACC | acgggggactcttgaccatggATGGATAACTCATCGACTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, G.; Saleem, M.; Deng, T.; Zhong, Z.; He, T.; Wu, J. Unraveling the Mechanism of StWRKY6 in Potato (Solanum tuberosum)’s Cadmium Tolerance for Ensuring Food Safety. Foods 2023, 12, 2303. https://doi.org/10.3390/foods12122303
He G, Saleem M, Deng T, Zhong Z, He T, Wu J. Unraveling the Mechanism of StWRKY6 in Potato (Solanum tuberosum)’s Cadmium Tolerance for Ensuring Food Safety. Foods. 2023; 12(12):2303. https://doi.org/10.3390/foods12122303
Chicago/Turabian StyleHe, Guandi, Muhammad Saleem, Tingfei Deng, Zhuoyan Zhong, Tengbing He, and Jiahai Wu. 2023. "Unraveling the Mechanism of StWRKY6 in Potato (Solanum tuberosum)’s Cadmium Tolerance for Ensuring Food Safety" Foods 12, no. 12: 2303. https://doi.org/10.3390/foods12122303