
Functions of SMC2 in the Development of Zebrafish Liver


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry and Ethics
2.2. Generation of SMC2 Mutant Zebrafish Lines
2.3. Quantitative Real-Time PCR (qPCR)
2.4. Whole-Mount In Situ Hybridization (WISH)
2.5. TUNEL Assay
2.6. Immunofluorescence Staining
2.7. EdU Labeling
2.8. Synthesis of Capped SMC2 mRNA
2.9. Fluorescence-Activated Cell Sorting
2.10. Statistical Analysis
3. Results
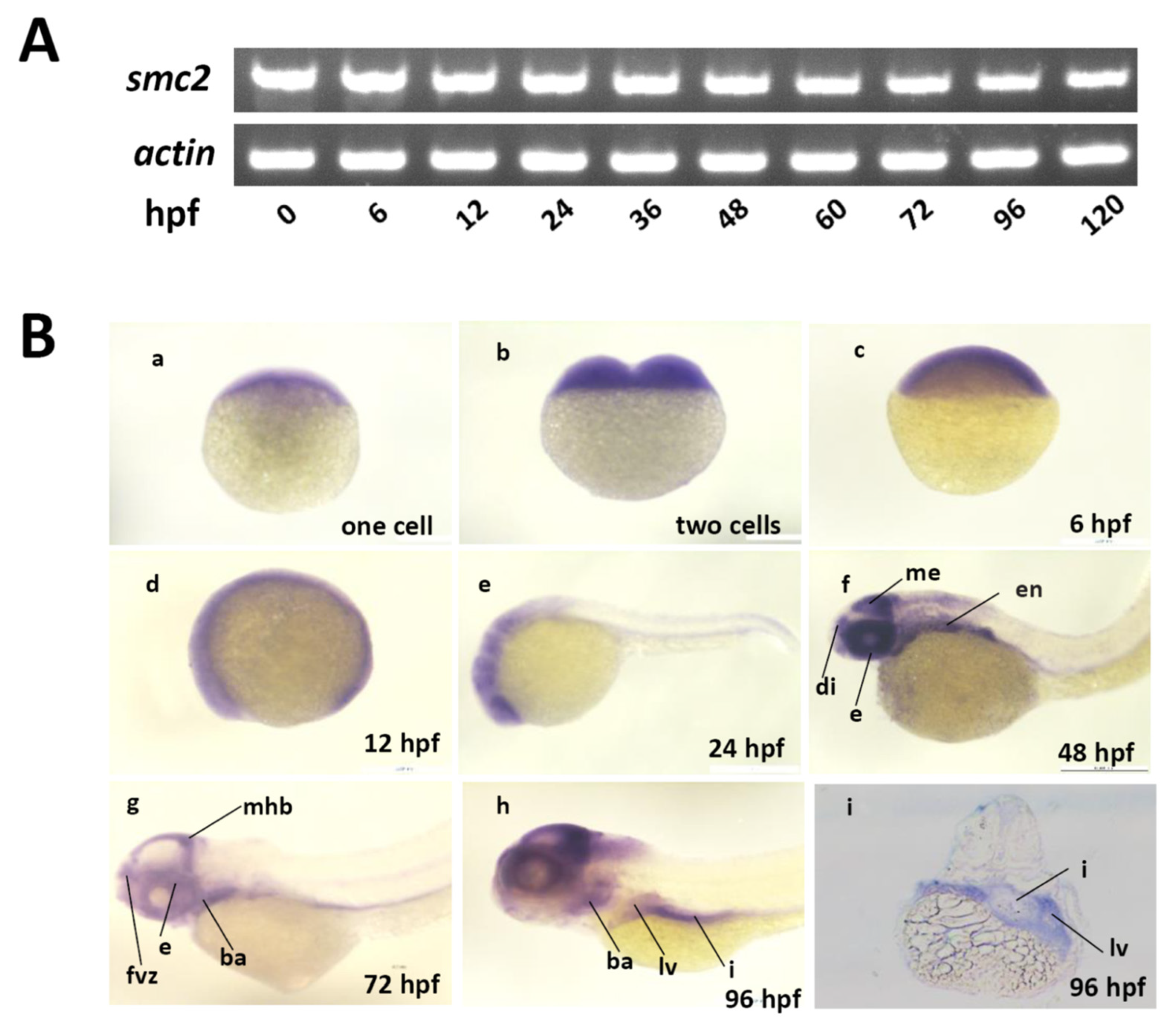
3.1. Conservation of SMC2-Containing Complexes among Vertebrates, and the Expression Patterns of SMC2 during Embryonic Development of Zebrafish
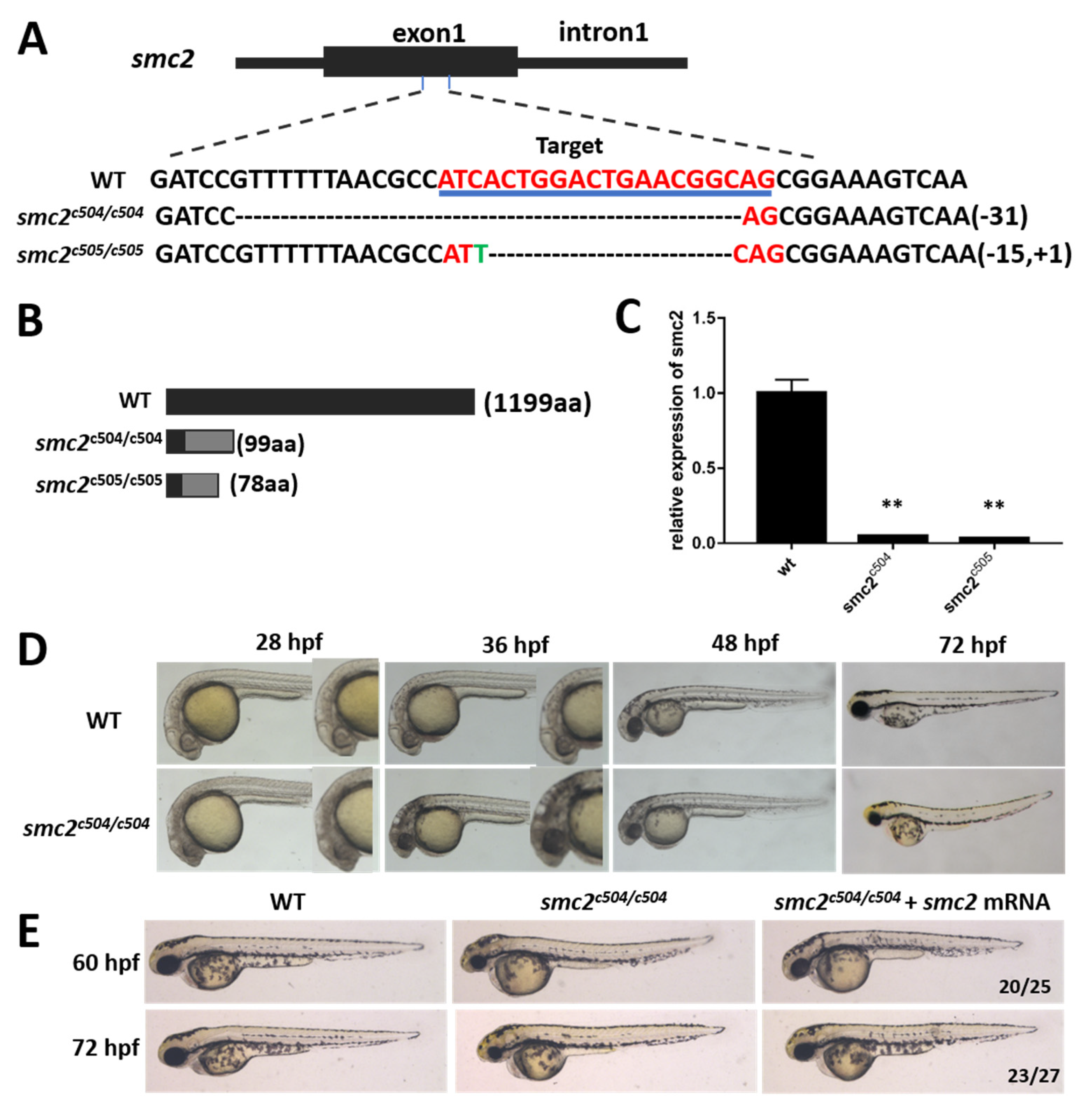
3.2. Knockout of SMC2 in Zebrafish
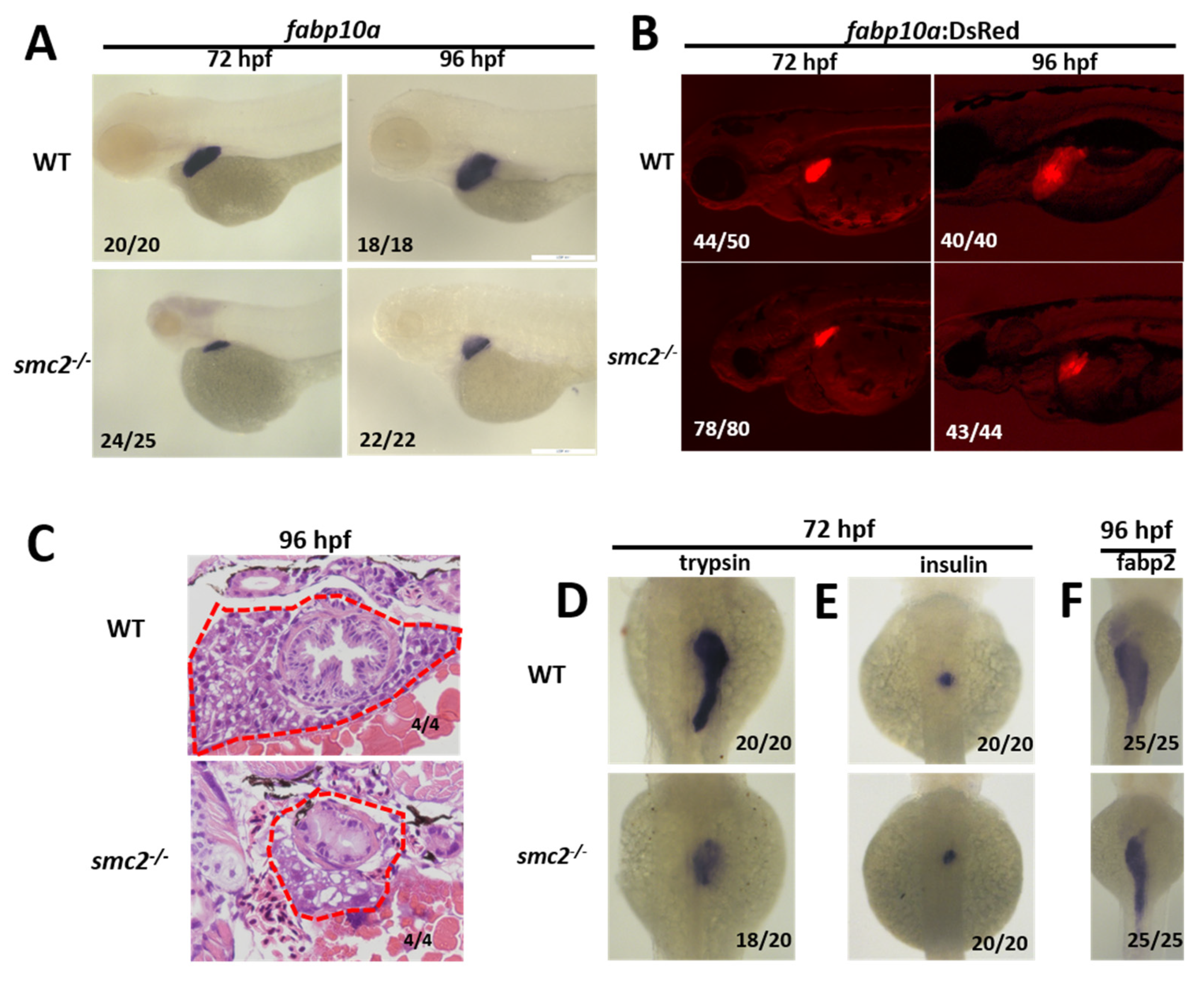
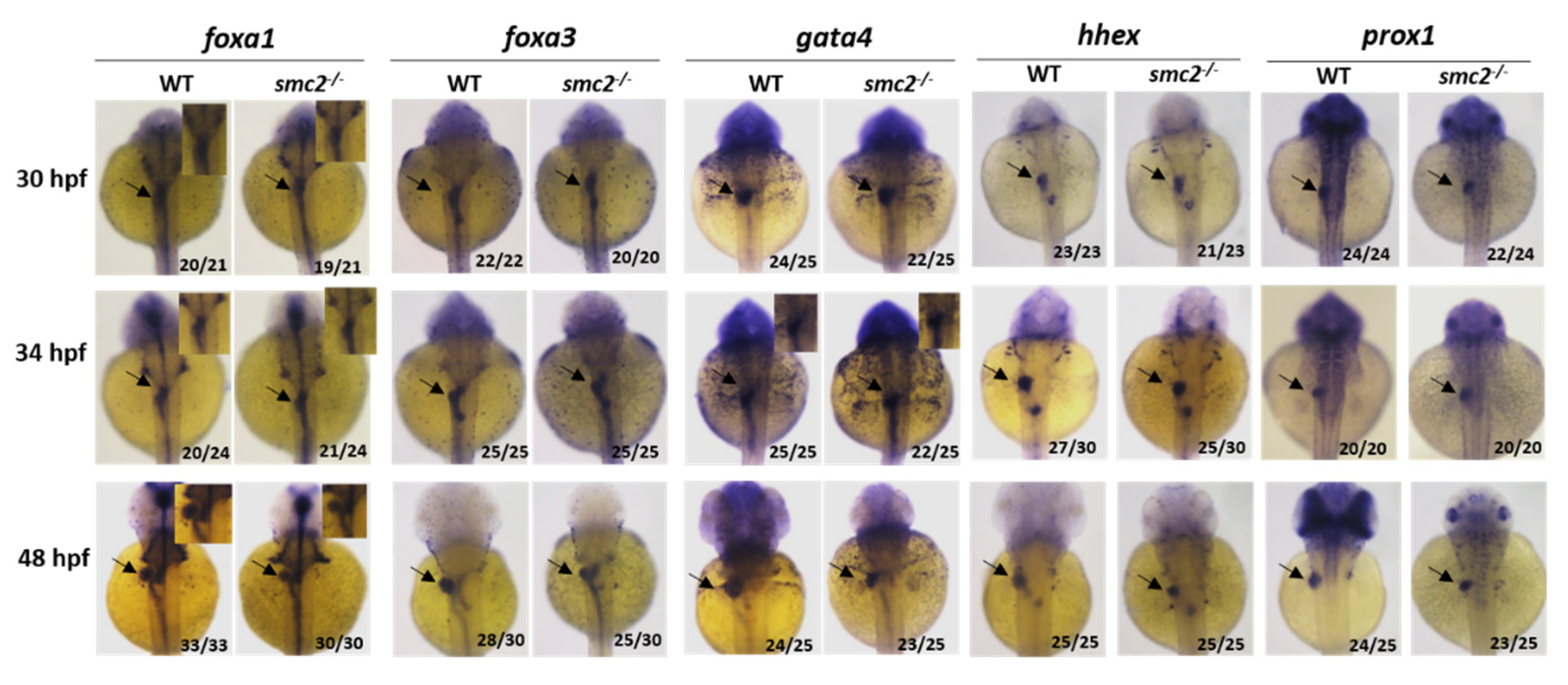
3.3. Loss of SMC2 Led to a Small Liver Phenotype in SMC2−/− Mutants
3.4. SMC2 Is Required for Liver Expansion
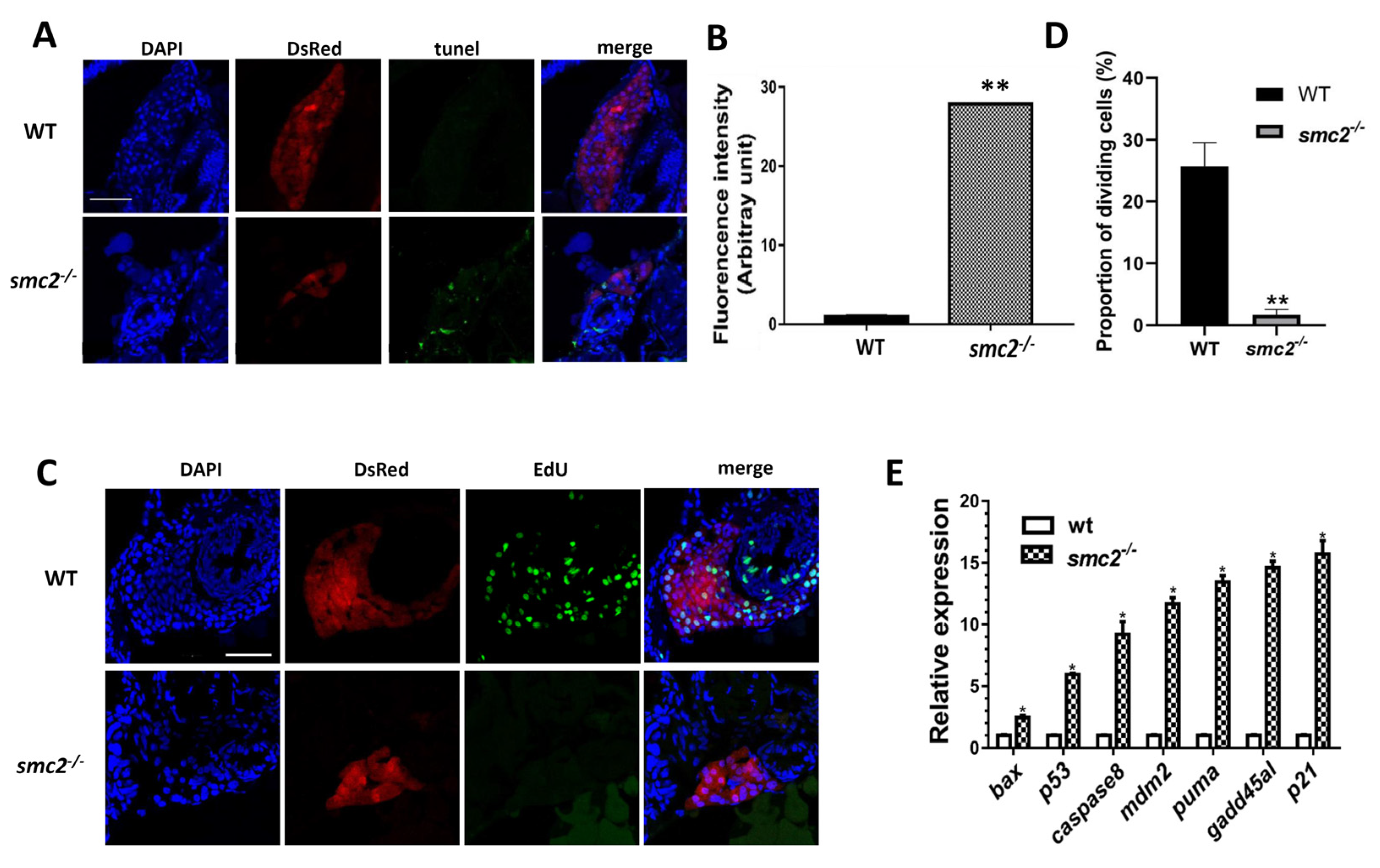
3.5. Hepatocellular Apoptosis Increased in SMC2−/− Mutants
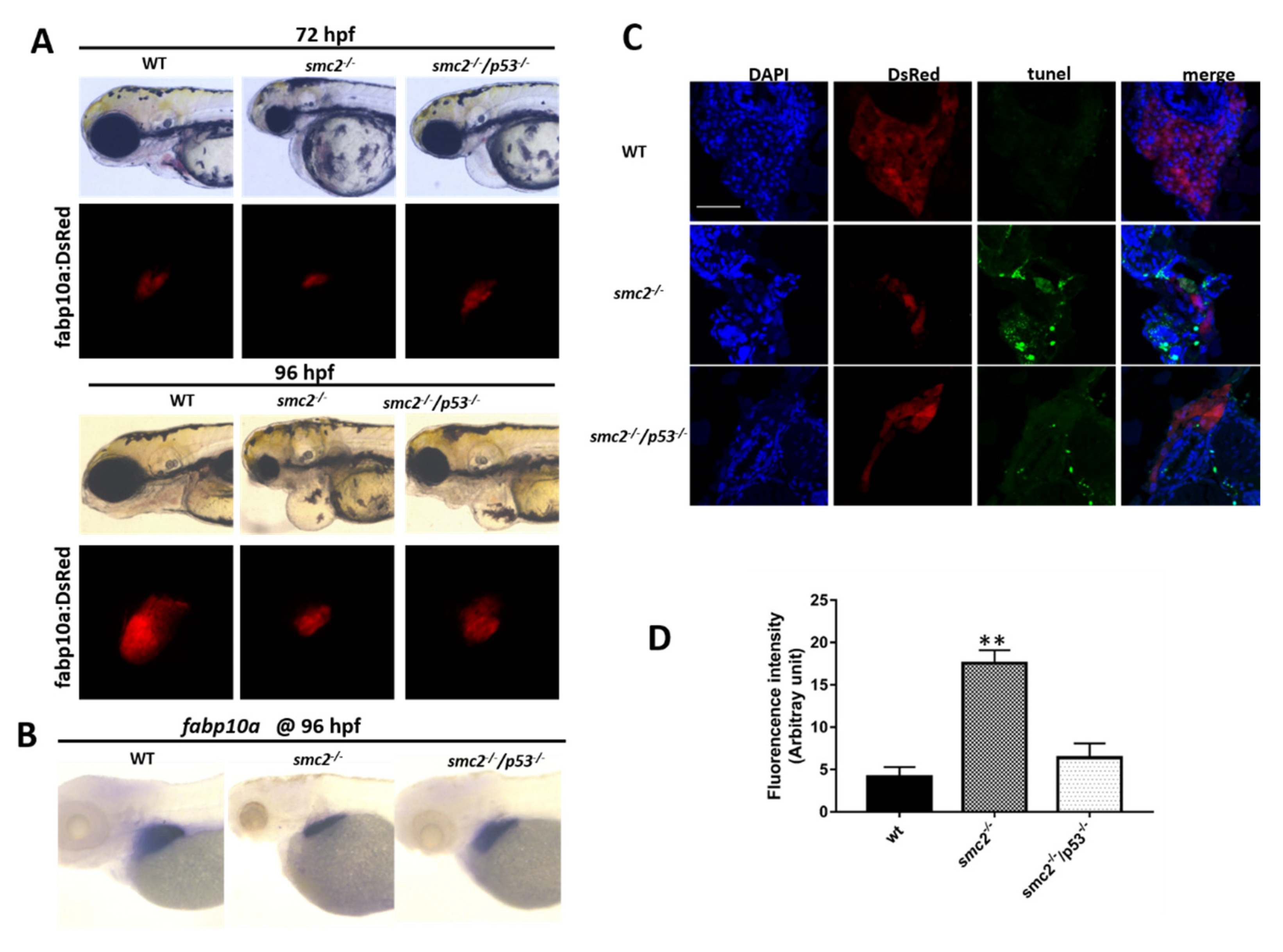
3.6. Activation of the p53-Driven Apoptotic Pathway Contributed to the Small Liver Phenotype in SMC2−/− Mutants
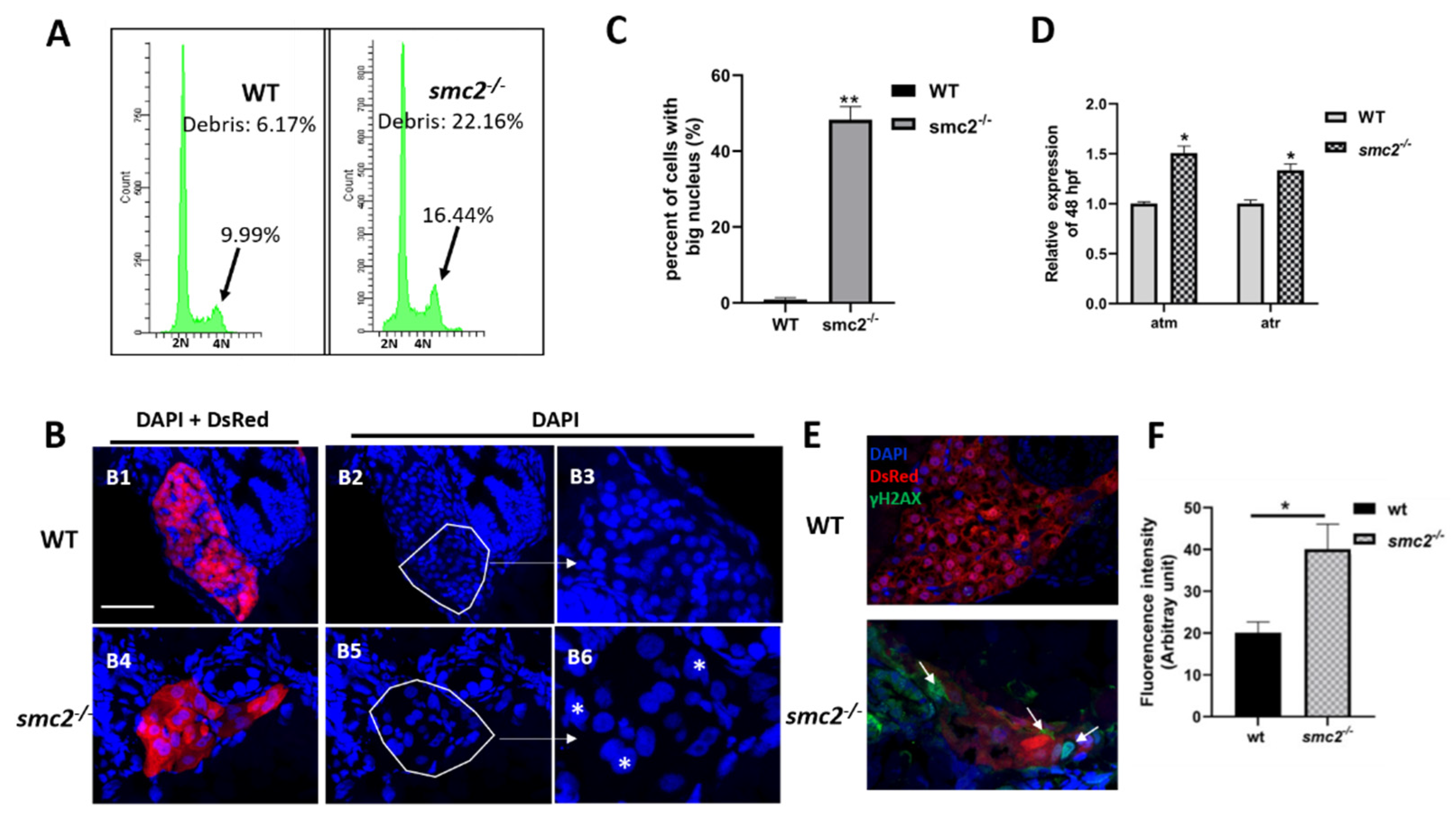
3.7. Extensive Apoptosis Occurring in the Liver of SMC2 Mutants Is Attributable to DNA Damage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wood, A.J.; Severson, A.F.; Meyer, B.J. Condensin and cohesin complexity: The expanding repertoire of functions. Nat. Rev. Genet. 2010, 11, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Jessberger, R. The many functions of SMC proteins in chromosome dynamics. Nat. Rev. Mol. Cell Biol. 2002, 3, 767–778. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Yuan, J.; Zhang, J.; Han, F. The condensin subunits SMC2 and SMC4 interact for correct condensation and segregation of mitotic maize chromosomes. Plant J. Cell Mol. Biol. 2020, 102, 467–479. [Google Scholar] [CrossRef]
- Nolivos, S.; Sherratt, D. The bacterial chromosome: Architecture and action of bacterial SMC and SMC-like complexes. FEMS Microbiol. Rev. 2014, 38, 380–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matityahu, A.; Onn, I. A new twist in the coil: Functions of the coiled-coil domain of structural maintenance of chromosome (SMC) proteins. Curr. Genet. 2018, 64, 109–116. [Google Scholar] [CrossRef]
- Losada, A. Dynamic molecular linkers of the genome: The first decade of SMC proteins. Genes Dev. 2005, 19, 1269–1287. [Google Scholar] [CrossRef] [Green Version]
- Tsang, C.K.; Wei, Y.; Zheng, X.F. Compacting DNA during the interphase: Condensin maintains rDNA integrity. Cell Cycle 2007, 6, 2213–2218. [Google Scholar] [CrossRef] [PubMed]
- Fazzio, T.G.; Panning, B. Condensin complexes regulate mitotic progression and interphase chromatin structure in embryonic stem cells. J. Cell Biol. 2010, 188, 491–503. [Google Scholar] [CrossRef] [Green Version]
- Hartl, T.A.; Sweeney, S.J.; Knepler, P.J.; Bosco, G. Condensin II resolves chromosomal associations to enable anaphase I segregation in Drosophila male meiosis. PLoS Genet. 2008, 4, e1000228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagstrom, K.A.; Holmes, V.F.; Cozzarelli, N.R.; Meyer, B.J.C. elegans condensin promotes mitotic chromosome architecture, centromere organization, and sister chromatid segregation during mitosis and meiosis. Genes Dev. 2002, 16, 729–742. [Google Scholar] [CrossRef] [Green Version]
- Nishide, K.; Hirano, T. Overlapping and non-overlapping functions of condensins I and II in neural stem cell divisions. PLoS Genet. 2014, 10, e1004847. [Google Scholar] [CrossRef] [PubMed]
- Zaret, K.S. Regulatory phases of early liver development: Paradigms of organogenesis. Nat. Rev. Genet. 2002, 3, 499–512. [Google Scholar] [CrossRef]
- Field, H.A.; Ober, E.A.; Roeser, T.; Stainier, D.Y.R. Formation of the digestive system in zebrafish. I. Liver morphogenesis. Dev. Biol. 2003, 253, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Ober, E.A.; Verkade, H.; Field, H.A.; Stainier, D.Y. Mesodermal Wnt2b signalling positively regulates liver specification. Nature 2006, 442, 688–691. [Google Scholar] [CrossRef]
- Shin, D.; Shin, C.H.; Tucker, J.; Ober, E.A.; Rentzsch, F.; Poss, K.D.; Hammerschmidt, M.; Mullins, M.C.; Stainier, D.Y. Bmp and Fgf signaling are essential for liver specification in zebrafish. Development 2007, 134, 2041–2050. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Sadler, K.C. New school in liver development: Lessons from zebrafish. Hepatology 2009, 50, 1656–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, Z.; Oliver, G. Prox1 is an early specific marker for the developing liver and pancreas in the mammalian foregut endoderm. Mech. Dev. 2002, 118, 147–155. [Google Scholar] [CrossRef]
- Zhang, W.; Yatskievych, T.A.; Baker, R.K.; Antin, P.B. Regulation of Hex gene expression and initial stages of avian hepatogenesis by Bmp and Fgf signaling. Dev. Biol. 2004, 268, 312–326. [Google Scholar] [CrossRef] [Green Version]
- Berghmans, S.; Murphey, R.D.; Wienholds, E.; Neuberg, D.; Kutok, J.L.; Fletcher, C.D.; Morris, J.P.; Liu, T.X.; Schulte-Merker, S.; Kanki, J.P.; et al. tp53 mutant zebrafish develop malignant peripheral nerve sheath tumors. Proc. Natl. Acad. Sci. USA 2005, 102, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Korzh, S.; Pan, X.; Garcia-Lecea, M.; Winata, C.L.; Pan, X.; Wohland, T.; Korzh, V.; Gong, Z. Requirement of vasculogenesis and blood circulation in late stages of liver growth in zebrafish. BMC Dev. Biol. 2008, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Chang, N.; Sun, C.; Gao, L.; Zhu, D.; Xu, X.; Zhu, X.; Xiong, J.W.; Xi, J.J. Genome editing with RNA-guided Cas9 nuclease in zebrafish embryos. Cell Res. 2013, 23, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitramuthu, B.P.; Bennett, H.P. High resolution whole mount in situ hybridization within zebrafish embryos to study gene expression and function. J. Vis. Exp. JoVE 2013, 80, e50644. [Google Scholar] [CrossRef] [Green Version]
- Wittkopp, N.; Huntzinger, E.; Weiler, C.; Saulière, J.; Schmidt, S.; Sonawane, M.; Izaurralde, E. Nonsense-mediated mRNA decay effectors are essential for zebrafish embryonic development and survival. Mol. Cell. Biol. 2009, 29, 3517–3528. [Google Scholar] [CrossRef] [Green Version]
- Tao, T.; Peng, J. Liver development in zebrafish (Danio rerio). J. Genet. Genom. 2009, 36, 325–334. [Google Scholar] [CrossRef]
- Ober, E.A.; Field, H.A.; Stainier, D.Y.R. From endoderm formation to liver and pancreas development in zebrafish. Mech. Dev. 2003, 120, 5–18. [Google Scholar] [CrossRef]
- Wallace, K.N.; Pack, M. Unique and conserved aspects of gut development in zebrafish. Dev. Biol. 2003, 255, 12–29. [Google Scholar] [CrossRef] [Green Version]
- Poulain, M.; Ober, E.A. Interplay between Wnt2 and Wnt2bb controls multiple steps of early foregut-derived organ development. Development 2011, 138, 3557–3568. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Li, X.; Gu, Q.; Li, Q.; Cui, Z. Nucleoporin 62-Like Protein is Required for the Development of Pharyngeal Arches through Regulation of Wnt/beta-Catenin Signaling and Apoptotic Homeostasis in Zebrafish. Cells 2019, 8, 1038. [Google Scholar] [CrossRef] [Green Version]
- Price, B.D.; D’Andrea, A.D. Chromatin remodeling at DNA double-strand breaks. Cell 2013, 152, 1344–1354. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T. Condensin-Based Chromosome Organization from Bacteria to Vertebrates. Cell 2016, 164, 847–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, L.C.; Kalitsis, P.; Chang, T.M.; Cipetic, M.; Kim, J.H.; Marshall, O.; Turnbull, L.; Whitchurch, C.B.; Vagnarelli, P.; Samejima, K.; et al. Contrasting roles of condensin I and condensin II in mitotic chromosome formation. J. Cell Sci. 2012, 125, 1591–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houlard, M.; Godwin, J.; Metson, J.; Lee, J.; Hirano, T.; Nasmyth, K. Condensin confers the longitudinal rigidity of chromosomes. Nat. Cell Biol. 2015, 17, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Hafner, A.; Bulyk, M.L.; Jambhekar, A.; Lahav, G. The multiple mechanisms that regulate p53 activity and cell fate. Nat. Rev. Mol. Cell Biol. 2019, 20, 199–210. [Google Scholar] [CrossRef]
- Gottlieb, T.M.; Oren, M. P53 in growth control and neoplasia. Biochim. Biophys. Acta 1996, 1287, 77–102. [Google Scholar] [CrossRef]
- Levine, A.J. p53, the cellular gatekeeper for growth and division. Cell 1997, 88, 323. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA damage, repair, and mutagenesis. Environ. Mol. Mutagenesis 2017, 58, 235–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.-B.S.; Elledge, S.J. The DNA damage response: Putting checkpoints in perspective. Nature 2000, 408, 433–439. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Song, G.; Zhao, Y.; Ren, J.; Li, Q.; Cui, Z. Functions of SMC2 in the Development of Zebrafish Liver. Biomedicines 2021, 9, 1240. https://doi.org/10.3390/biomedicines9091240
Li X, Song G, Zhao Y, Ren J, Li Q, Cui Z. Functions of SMC2 in the Development of Zebrafish Liver. Biomedicines. 2021; 9(9):1240. https://doi.org/10.3390/biomedicines9091240
Chicago/Turabian StyleLi, Xixi, Guili Song, Yasong Zhao, Jing Ren, Qing Li, and Zongbin Cui. 2021. "Functions of SMC2 in the Development of Zebrafish Liver" Biomedicines 9, no. 9: 1240. https://doi.org/10.3390/biomedicines9091240