
Knockout of mafba Causes Inner-Ear Developmental Defects in Zebrafish via the Impairment of Proliferation and Differentiation of Ionocyte Progenitor Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Maintenance
2.2. Generation of Mafba Mutant by CRISPR/Cas9 Technology
2.3. Whole-Mount In Situ Hybridization
2.4. Whole-Mount Immunofluorescence Assay
2.5. EDU and TUNEL Assay
2.6. Western Blot
2.7. Bioinformatics
2.8. Comet Assay
2.9. Startle Response Tests
2.10. Quantitative RT-PCR Analysis
2.11. Flow Cytometry Analysis for Cell Cycle
2.12. Statistical Analysis
3. Results
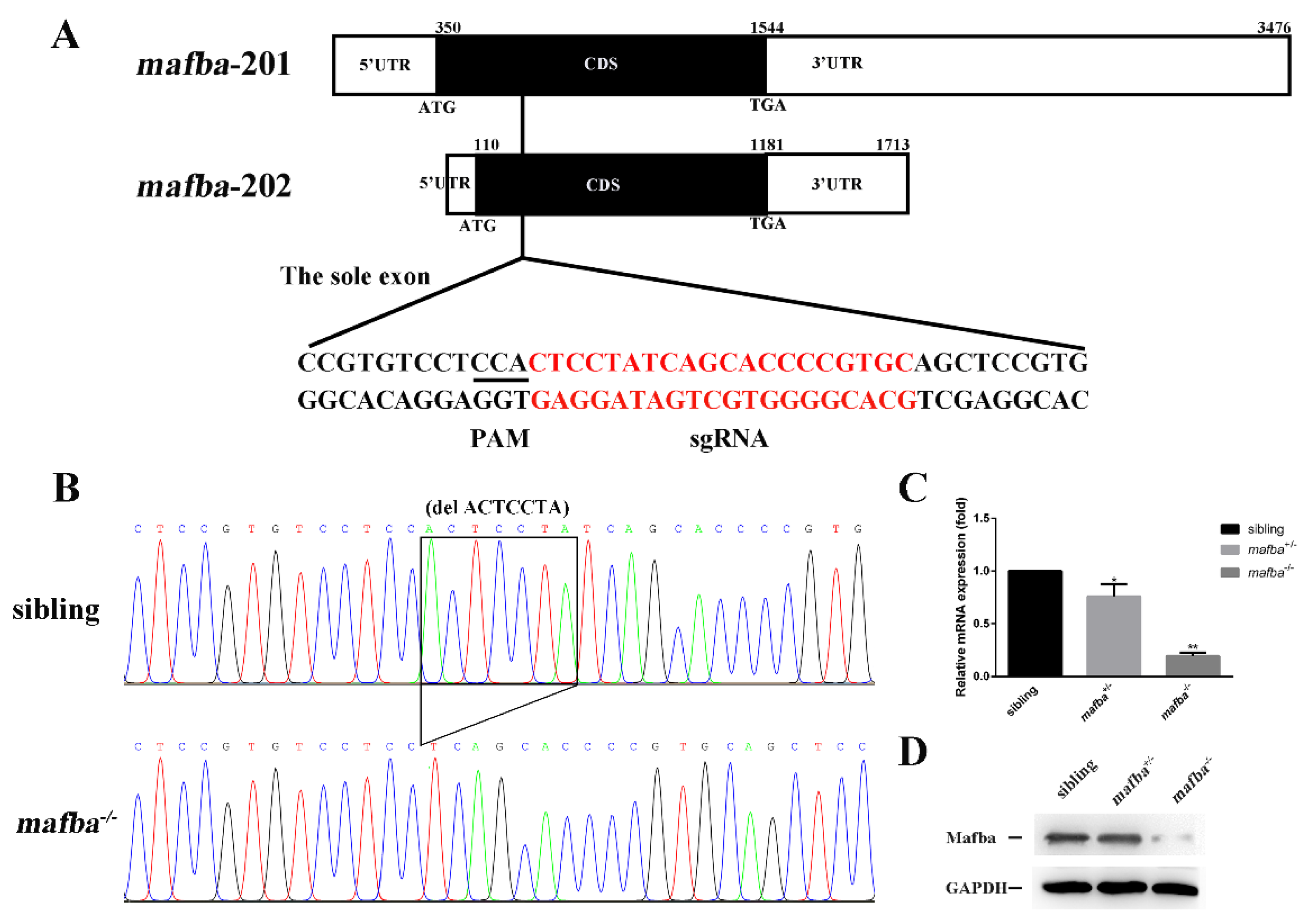
3.1. Generation of mafba−/− Mutant Zebrafish with CRISPR/Cas9 Technology
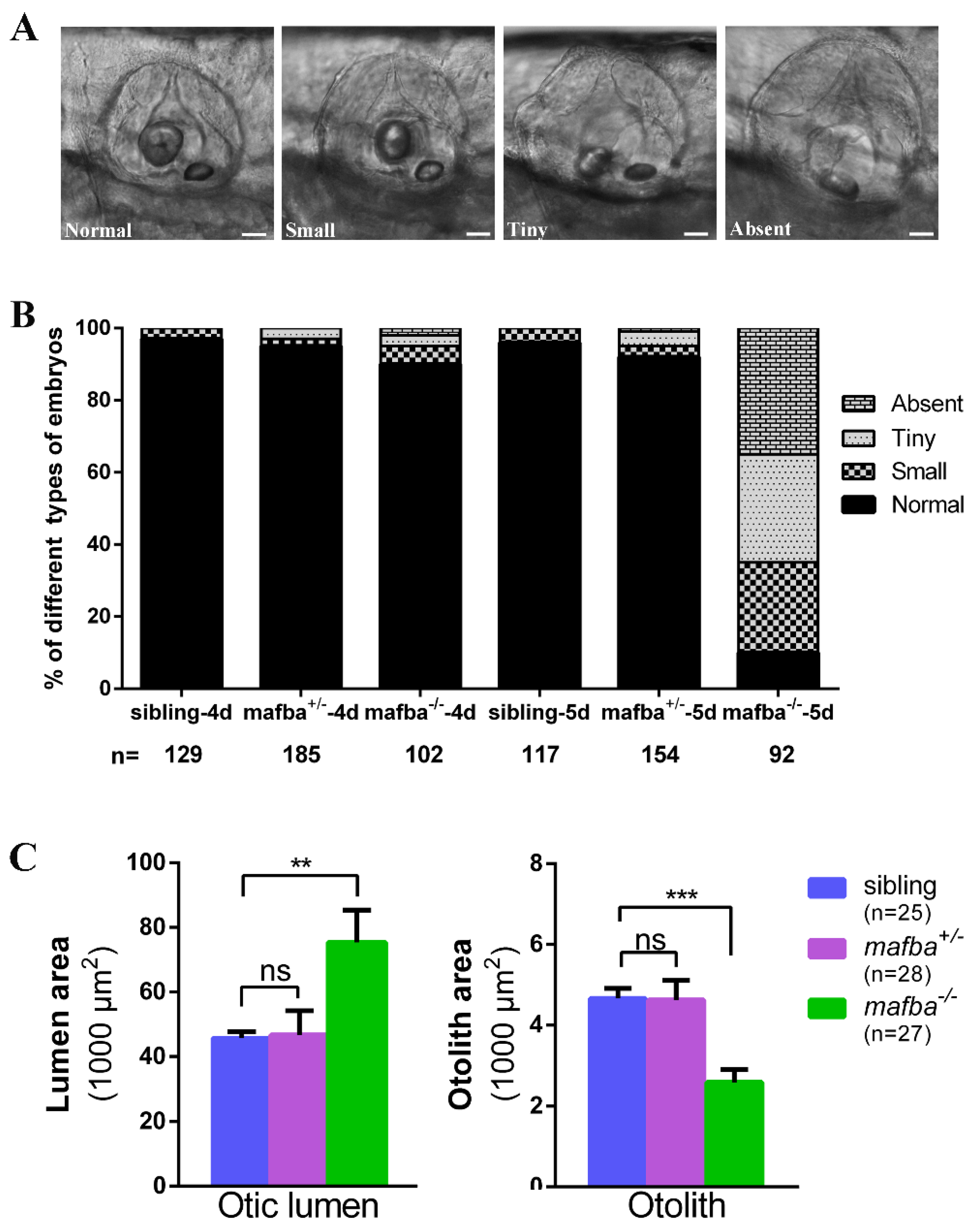
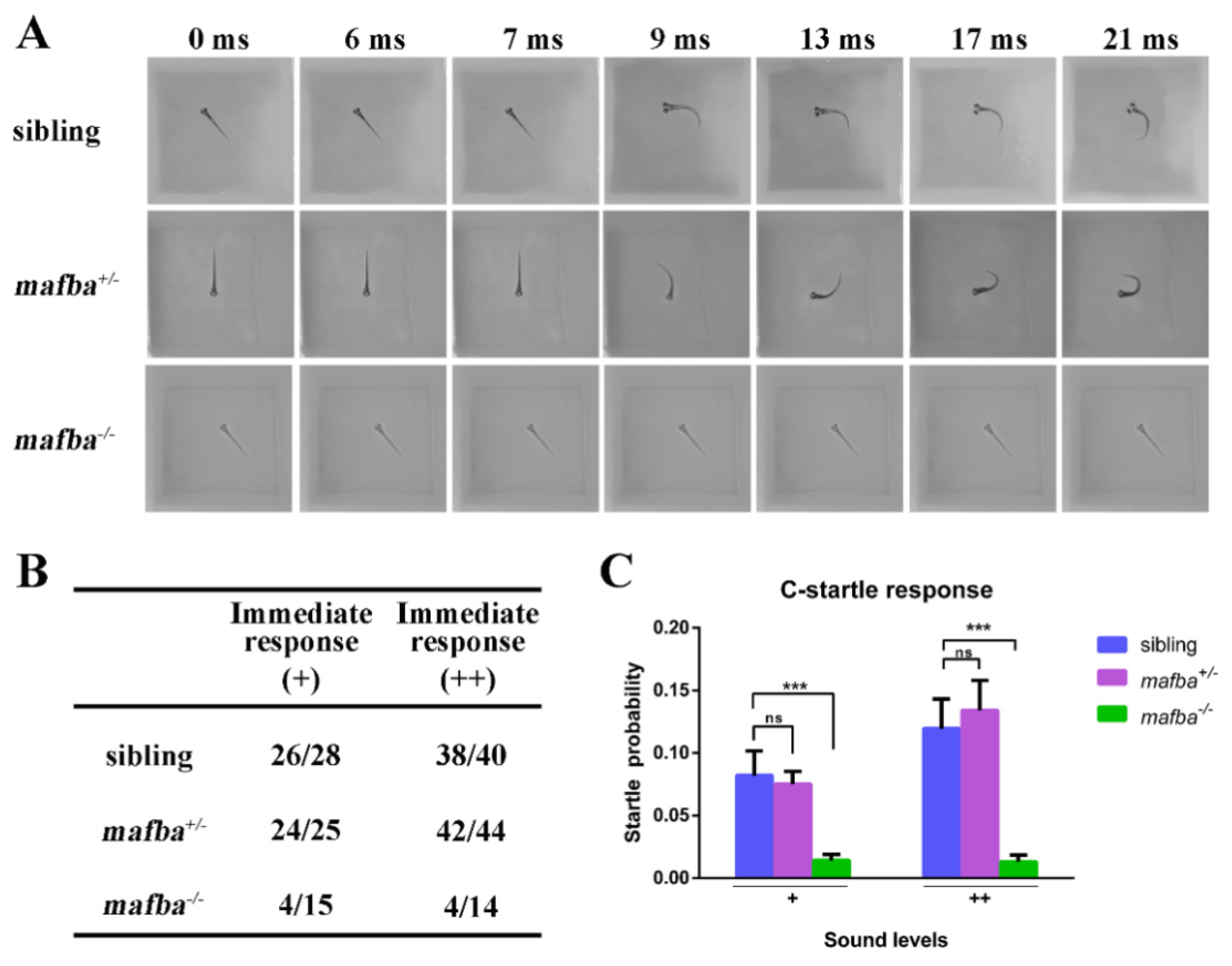
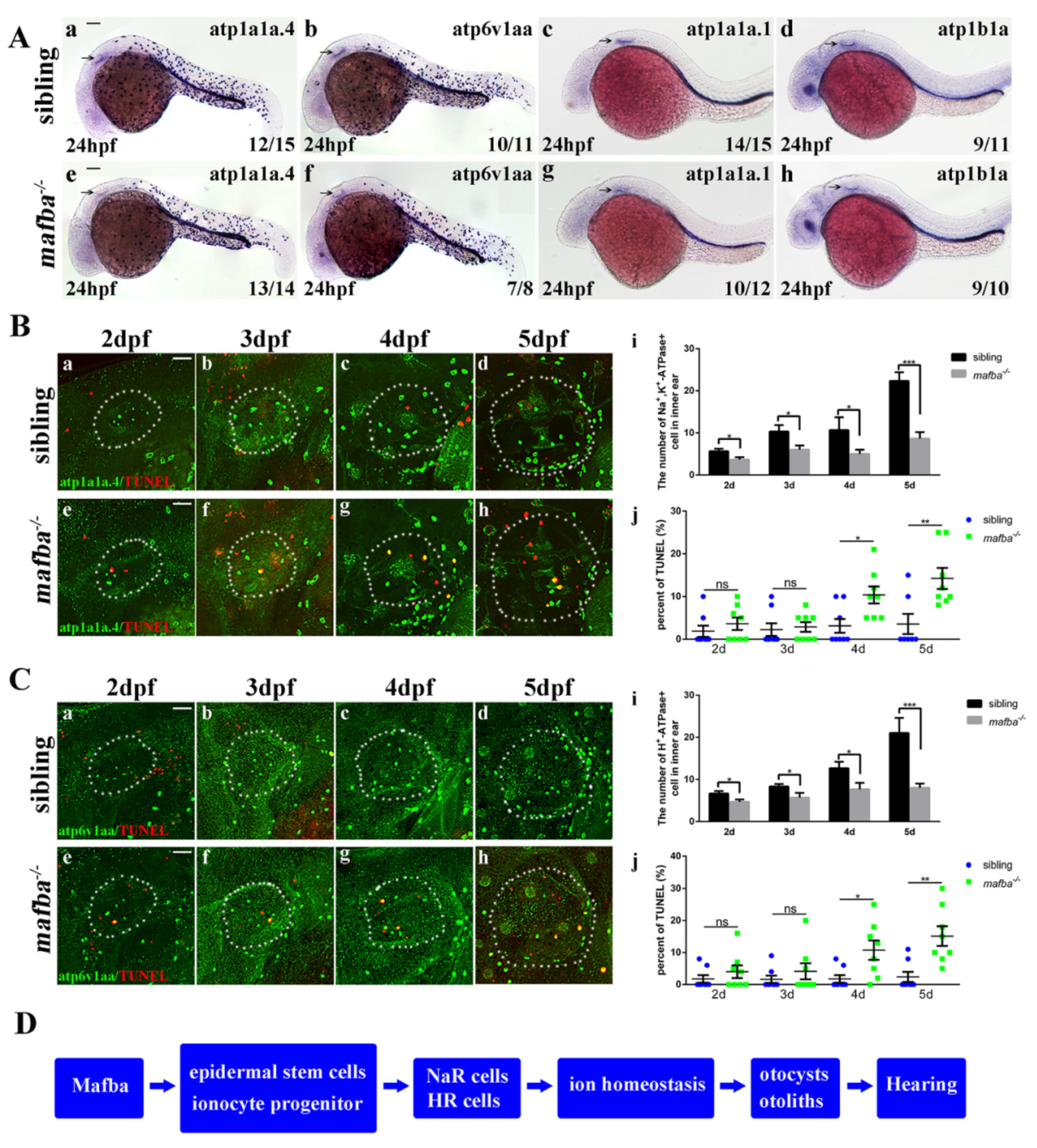
3.2. Depletion of Zebrafish mafba Results in Inner-Ear Defects and Hearing Loss
3.3. Deficiency of mafba Does Not Affect Otic Patterning or Hair Cell Development
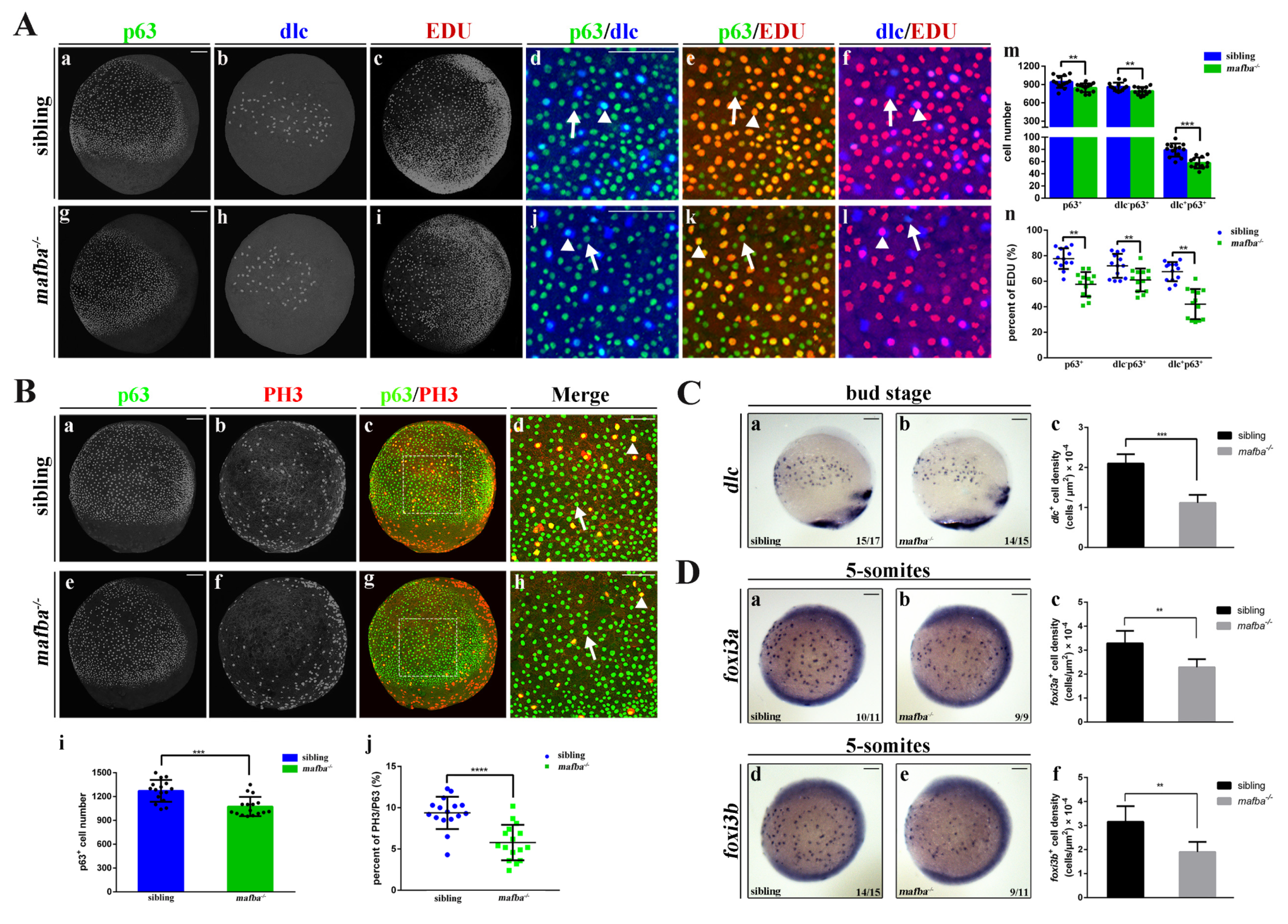
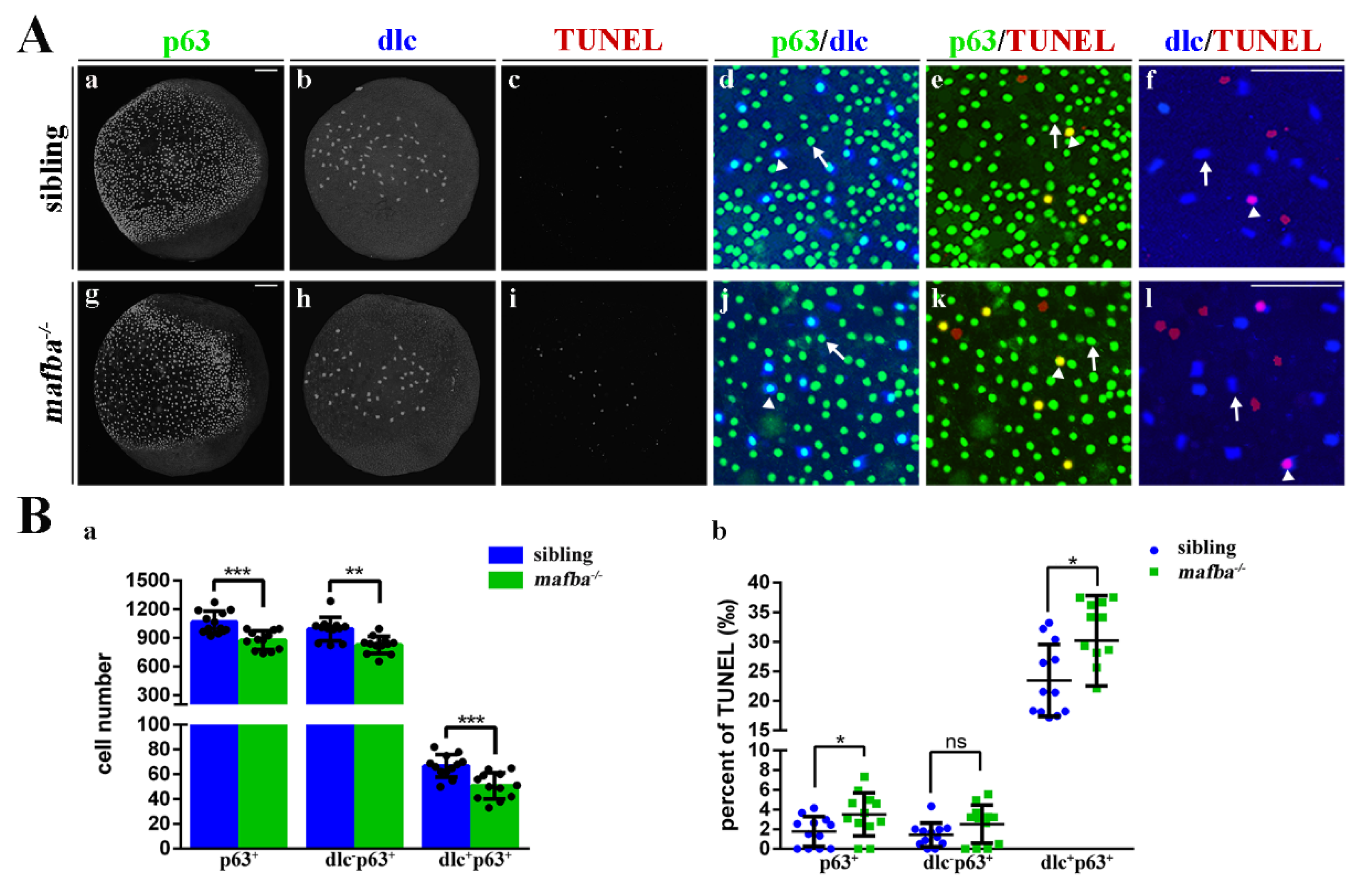
3.4. Knockout of mafba Suppresses the Proliferation of p63+ Epidermal Stem Cells and Reduces dlc+ Ionocyte Progenitor Cell Number
3.5. Apoptosis of the Differentiated Epidermal Ionocytes Is Increased in mafba−/− Embryos
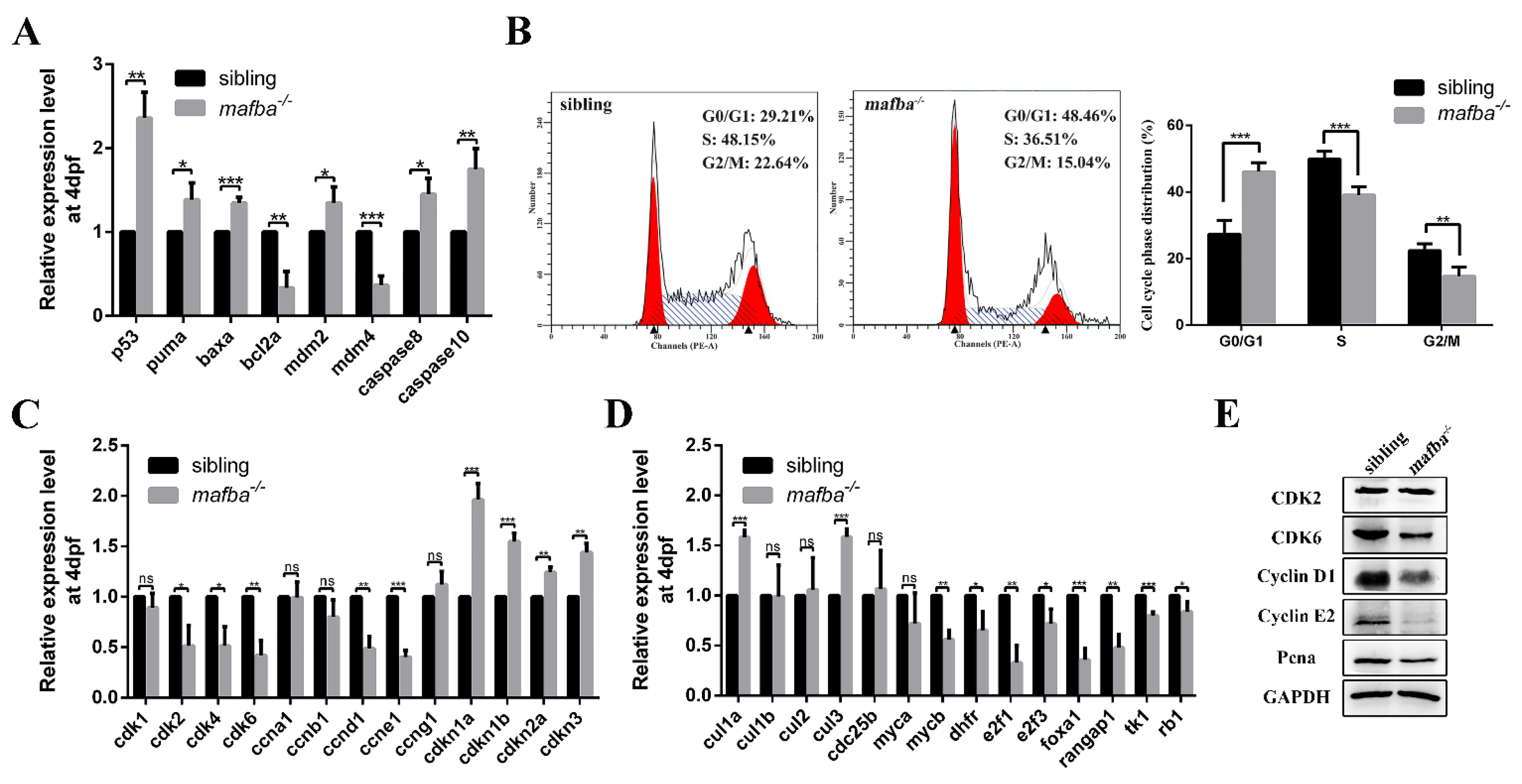
3.6. Deprivation of mafba Activates p53 Apoptosis Pathway and Arrests the Cell Cycle in the G0/G1 Phase
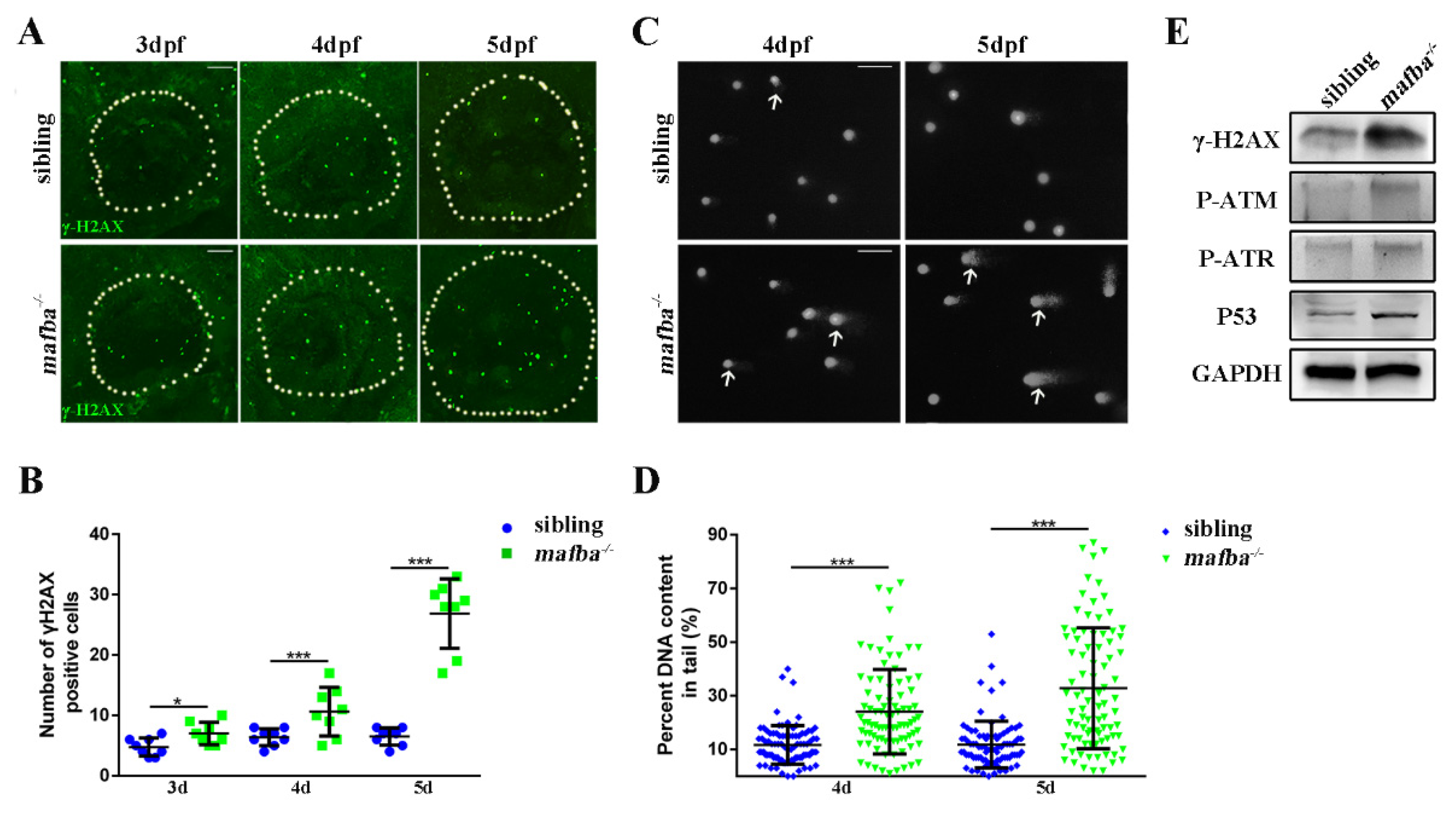
3.7. Accumulation of DNA Damages in mafba−/− Inner-Ear
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mittal, R.; Nguyen, D.; Patel, A.P.; Debs, L.H.; Mittal, J.; Yan, D.; Eshraghi, A.A.; Van De Water, T.R.; Liu, X.Z. Recent Advancements in the Regeneration of Auditory Hair Cells and Hearing Restoration. Front. Mol. Neurosci. 2017, 10, 236. [Google Scholar] [CrossRef] [Green Version]
- Ganapathy, A.; Pandey, N.; Srisailapathy, C.R.S.; Jalvi, R.; Malhotra, V.; Venkatappa, M.; Chatterjee, A.; Sharma, M.; Santhanam, R.; Chadha, S.; et al. Non-Syndromic Hearing Impairment in India: High Allelic Heterogeneity among Mutations in TMPRSS3, TMC1, USHIC, CDH23 and TMIE. PLoS ONE 2014, 9, e84773. [Google Scholar] [CrossRef] [Green Version]
- Nance, W.E.; Lim, B.G.; Dodson, K.M. Importance of congenital cytomegalovirus infections as a cause for pre-lingual hearing loss. J. Clin. Virol. 2006, 35, 221–225. [Google Scholar] [CrossRef]
- Fritzsch, B.; Pauley, S.; Beisel, K.W. Cells, molecules and morphogenesis: The making of the vertebrate ear. Brain Res. 2006, 1091, 151–171. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, B.L.; Johnston, M.; Sulik, K. Congenital Malformations of the External Middle, and Inner Ear Produced by Isotretinoin Exposure in Mouse Embryos. Otolaryngol. Head Neck Surg. 1990, 102, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A. Accelerating our understanding of the inner ear of sauropodomorpha: First global, statistical analysis of semicircular canals of diplodocid and macronarian dinosaurs and its implication for neck-posture. In Proceedings of the SVP 74th Annual Meeting, Berlin, Germany, 6 November 2014. [Google Scholar]
- Whitfield, T.T.; Mburu, P.; Hardisty-Hughes, R.E.; Brown, S.D. Models of congenital deafness: Mouse and zebrafish. Drug Discov. Today Dis. Model. 2005, 2, 85–92. [Google Scholar] [CrossRef]
- Han, Y.; Mu, Y.; Li, X.; Xu, P.; Tong, J.; Liu, Z.; Ma, T.; Zeng, G.; Yang, S.; Du, J.; et al. Grhl2 deficiency impairs otic development and hearing ability in a zebrafish model of the progressive dominant hearing loss DFNA28. Hum. Mol. Genet. 2011, 20, 3213–3226. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Song, G.; Zhao, Y.; Zhao, F.; Liu, C.; Liu, D.; Li, Q.; Cui, Z. Claudin7b is required for the formation and function of inner ear in zebrafish. J. Cell. Physiol. 2017, 233, 3195–3206. [Google Scholar] [CrossRef]
- Abbas, L.; Whitfield, T.T. The zebrafish inner ear. Fish Physiol. 2010, 29, 123–171. [Google Scholar] [CrossRef]
- Whitfield, T.T.; Riley, B.; Chiang, M.-Y.; Phillips, B. Development of the zebrafish inner ear. Dev. Dyn. 2002, 223, 427–458. [Google Scholar] [CrossRef]
- Whitfield, T.T.; Granato, M.; van Eeden, F.J.; Schach, U.; Brand, M.; Furutani-Seiki, M.; Haffter, P.; Hammerschmidt, M.; Heisenberg, C.P.; Jiang, Y.J.; et al. Mutations affecting development of the zebrafish inner ear and lateral line. Development 1996, 123, 241–256. [Google Scholar] [CrossRef]
- Trune, D.R. Ion homeostasis in the ear: Mechanisms, maladies, and management. Curr. Opin. Otolaryngol. Head Neck Surg. 2010, 18, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Ellertsdottir, E.; Ganz, J.; Dürr, K.; Loges, N.; Biemar, F.; Seifert, F.; Ettl, A.-K.; Kramer-Zucker, A.K.; Nitschke, R.; Driever, W. A mutation in the zebrafish Na,K-ATPase subunitatp1a1a.1provides genetic evidence that the sodium potassium pump contributes to left-right asymmetry downstream or in parallel to nodal flow. Dev. Dyn. 2006, 235, 1794–1808. [Google Scholar] [CrossRef] [PubMed]
- Blasiole, B.; Canfield, V.A.; Vollrath, M.A.; Huss, D.; Mohideen, M.-A.P.; Dickman, J.D.; Cheng, K.; Fekete, D.M.; Levenson, R. Separate Na,K-ATPase genes are required for otolith formation and semicircular canal development in zebrafish. Dev. Biol. 2006, 294, 148–160. [Google Scholar] [CrossRef] [Green Version]
- Jänicke, M.; Carney, T.J.; Hammerschmidt, M. Foxi3 transcription factors and Notch signaling control the formation of skin ionocytes from epidermal precursors of the zebrafish embryo. Dev. Biol. 2007, 307, 258–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, C.-D.; You, M.-S.; Guh, Y.-J.; Ma, M.; Jiang, Y.-J.; Hwang, P.-P. A Positive Regulatory Loop between foxi3a and foxi3b Is Essential for Specification and Differentiation of Zebrafish Epidermal Ionocytes. PLoS ONE 2007, 2, e302. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Liao, B.-K.; Lu, Y.-F.; Liu, Y.-H.; Hsieh, F.-C.; Hwang, P.-P.; Hwang, S.-P.L. Zebrafish Klf4 maintains the ionocyte progenitor population by regulating epidermal stem cell proliferation and lateral inhibition. PLoS Genet. 2019, 15, e1008058. [Google Scholar] [CrossRef]
- Evans, D.H. Teleost fish osmoregulation: What have we learned since August Krogh, Homer Smith, and Ancel Keys. Am. J. Physiol. Integr. Comp. Physiol. 2008, 295, R704–R713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, P.-P.; Chou, M.-Y. Zebrafish as an animal model to study ion homeostasis. Pflüg. Arch. 2013, 465, 1233–1247. [Google Scholar] [CrossRef] [Green Version]
- Hunter, G.L.; Hadjivasiliou, Z.; Bonin, H.; He, L.; Perrimon, N.; Charras, G.; Baum, B. Coordinated control of Notch-Delta signalling and cell cycle progression drives lateral inhibition mediated tissue patterning. Development 2016, 143, 2305–2310. [Google Scholar] [CrossRef] [Green Version]
- Cruz, S.A.; Chao, P.-L.; Hwang, P.-P. Cortisol promotes differentiation of epidermal ionocytes through Foxi3 transcription factors in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 164, 249–257. [Google Scholar] [CrossRef]
- Chang, W.J.; Horng, J.L.; Yan, J.J.; Hsiao, C.D.; Hwang, P.P. The transcription factor, glial cell missing 2, is involved in differentiation and functional regulation of H+-ATPase-rich cells in zebrafish (Danio rerio). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1192–R1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, L.; Whitfield, T.T. Nkcc1 (Slc12a2) is required for the regulation of endolymph volume in the otic vesicle and swim bladder volume in the zebrafish larva. Development 2009, 136, 2837–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, S.; Shiao, J.-C.; Liao, B.-K.; Huang, C.-J.; Hwang, P.-P. Plasma membrane calcium ATPase required for semicircular canal formation and otolith growth in the zebrafish inner ear. J. Exp. Biol. 2009, 212, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, K.; Noda, M.; Nishizawa, M. Maf nuclear oncoprotein recognizes sequences related to an AP-1 site and forms heterodimers with both Fos and Jun. Mol. Cell Biol. 1994, 14, 700–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriguchi, T.; Hamada, M.; Morito, N.; Terunuma, T.; Hasegawa, K.; Zhang, C.; Yokomizo, T.; Esaki, R.; Kuroda, E.; Yoh, K.; et al. MafB Is Essential for Renal Development and F4/80 Expression in Macrophages. Mol. Cell. Biol. 2006, 26, 5715–5727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, A.; Vanhille, L.; Mohideen, P.; Kelly, L.M.; Otto, C.; Bakri, Y.; Mossadegh, N.; Sarrazin, S.; Sieweke, M.H. Development of Macrophages with Altered Actin Organization in the Absence of MafB. Mol. Cell. Biol. 2006, 26, 6808–6818. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Kim, J.H.; Lee, J.; Jin, H.M.; Kook, H.; Kim, K.K.; Lee, S.Y.; Kim, N. MafB negatively regulates RANKL-mediated osteoclast differentiation. Blood 2006, 109, 3253–3259. [Google Scholar] [CrossRef]
- Miyai, M.; Hamada, M.; Moriguchi, T.; Hiruma, J.; Kamitani-Kawamoto, A.; Watanabe, H.; Hara-Chikuma, M.; Takahashi, K.; Takahashi, S.; Kataoka, K. Transcription Factor MafB Coordinates Epidermal Keratinocyte Differentiation. J. Investig. Dermatol. 2016, 136, 1848–1857. [Google Scholar] [CrossRef] [Green Version]
- Artner, I.; Le Lay, J.; Hang, Y.; Elghazi, L.; Schisler, J.; Henderson, E.; Sosa-Pineda, B.; Stein, R. MafB: An activator of the glucagon gene expressed in developing islet alpha- and beta-cells. Diabetes 2006, 55, 297–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamitani-Kawamoto, A.; Hamada, M.; Moriguchi, T.; Miyai, M.; Saji, F.; Hatamura, I.; Nishikawa, K.; Takayanagi, H.; Hitoshi, S.; Ikenaka, K.; et al. MafB interacts with Gcm2 and regulates parathyroid hormone expression and parathyroid development. J. Bone Miner. Res. 2011, 26, 2463–2472. [Google Scholar] [CrossRef]
- Sato, Y.; Tsukaguchi, H.; Morita, H.; Higasa, K.; Tran, M.T.N.; Hamada, M.; Usui, T.; Morito, N.; Horita, S.; Hayashi, T.; et al. A mutation in transcription factor MAFB causes Focal Segmental Glomerulosclerosis with Duane Retraction Syndrome. Kidney Int. 2018, 94, 396–407. [Google Scholar] [CrossRef]
- Park, J.G.; Tischfield, M.A.; Nugent, A.A.; Cheng, L.; Di Gioia, S.A.; Chan, W.-M.; Maconachie, G.; Bosley, T.M.; Summers, C.G.; Hunter, D.; et al. Loss of MAFB Function in Humans and Mice Causes Duane Syndrome, Aberrant Extraocular Muscle Innervation, and Inner-Ear Defects. Am. J. Hum. Genet. 2016, 98, 1220–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez-Echeverría, C.; Dominguez-Frutos, E.; Charnay, P.; Schimmang, T.; Pujades, C. Analysis of mouse kreisler mutants reveals new roles of hindbrain-derived signals in the establishment of the otic neurogenic domain. Dev. Biol. 2008, 322, 167–178. [Google Scholar] [CrossRef]
- Kwak, S.J.; Phillips, B.T.; Heck, R.; Riley, B.B. An expanded domain of fgf3 expression in the hindbrain of zebrafish valentino mutants results in mis-patterning of the otic vesicle. Development 2002, 129, 5279–5287. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.-S.; Zhang, X.-J.; Xie, Y.-Y.; Sun, X.-J.; Zhao, R.; Huang, Q.-H. SUMOylated MAFB promotes colorectal cancer tumorigenesis. Oncotarget 2016, 7, 83488–83501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suda, N.; Itoh, T.; Nakato, R.; Shirakawa, D.; Bando, M.; Katou, Y.; Kataoka, K.; Shirahige, K.; Tickle, C.; Tanaka, M. Dimeric combinations of MafB, cFos and cJun control the apoptosis-survival balance in limb morphogenesis. Development 2014, 141, 2885–2894. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Huang, Y.; Wang, L.; Huang, M.; Liu, F.; Liao, S.; Yu, S.; Lu, Z.; Han, S.; Hu, X.; et al. HSF4 regulates lens fiber cell differentiation by activating p53 and its downstream regulators. Cell Death Dis. 2017, 8, e3082. [Google Scholar] [CrossRef] [Green Version]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2007, 3, 59–69. [Google Scholar] [CrossRef]
- Malicki, J.; Avanesov, A.; Li, J.; Yuan, S.; Sun, Z. Analysis of Cilia Structure and Function in Zebrafish. Method Cell Biol. 2011, 101, 39–74. [Google Scholar] [CrossRef]
- Hu, X.; Lu, Z.; Yu, S.; Reilly, J.; Liu, F.; Jia, D.; Qin, Y.; Han, S.; Liu, X.; Qu, Z.; et al. CERKL regulates autophagy via the NAD-dependent deacetylase SIRT1. Autophagy 2018, 15, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Preibisch, S.; Saalfeld, S.; Tomancak, P. Globally optimal stitching of tiled 3D microscopic image acquisitions. Bioinformatics 2009, 25, 1463–1465. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Jiang, T.; Jia, D.; Han, Y.; Liu, F.; Huang, Y.; Qu, Z.; Zhao, Y.; Tu, J.; Lv, Y.; et al. BCAS2 is essential for hematopoietic stem and progenitor cell maintenance during zebrafish embryogenesis. Blood 2019, 133, 805–815. [Google Scholar] [CrossRef]
- Li, J.; Liu, F.; Lv, Y.; Sun, K.; Zhao, Y.; Reilly, J.; Zhang, Y.; Tu, J.; Yu, S.; Liu, X.; et al. Prpf31 is essential for the survival and differentiation of retinal progenitor cells by modulating alternative splicing. Nucleic Acids Res. 2021, 49, 2027–2043. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Qin, Y.; Yu, S.; Soares, D.; Yang, L.; Weng, J.; Li, C.; Gao, M.; Lu, Z.; Hu, X.; et al. Pathogenic mutations in retinitis pigmentosa 2 predominantly result in loss of RP2 protein stability in humans and zebrafish. J. Biol. Chem. 2017, 292, 6225–6239. [Google Scholar] [CrossRef] [Green Version]
- Końca, K.; Lankoff, A.; Banasik, A.; Lisowska, H.; Kuszewski, T.; Gozdz, S.; Koza, Z.; Wojcik, A. A cross-platform public domain PC image-analysis program for the comet assay. Mutat. Res. Toxicol. Environ. Mutagen. 2002, 534, 15–20. [Google Scholar] [CrossRef]
- Han, S.; Liu, X.; Xie, S.; Gao, M.; Liu, F.; Yu, S.; Sun, P.; Wang, C.; Archacki, S.; Lu, Z.; et al. Knockout of ush2a gene in zebrafish causes hearing impairment and late onset rod-cone dystrophy. Qual. Life Res. 2018, 137, 779–794. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.-C.; Wu, M.; Zhang, D.-C.; Guo, H.-Y.; Zhang, N.; Guo, L.; Liu, B.-S.; Jiang, S.-G. Toll-Like Receptor 5 of Golden Pompano Trachinotus ovatus (Linnaeus 1758): Characterization, Promoter Activity and Functional Analysis. Int. J. Mol. Sci. 2020, 21, 5916. [Google Scholar] [CrossRef]
- Yang, Y.; Hongyan, L.; Shicui, Z. Phylogenetic analysis of genes in the Maf family. Ludong Univ. J. Nat. Sci. Ed. 2014, 30, 330–335. [Google Scholar] [CrossRef]
- Kajihara, M.; Kawauchi, S.; Kobayashi, M.; Ogino, H.; Takahashi, S.; Yasuda, K. Isolation, characterization, and expression analysis of zebrafish large Mafs. J. Biochem. 2001, 129, 139–146. [Google Scholar] [CrossRef]
- Nicolson, T. The Genetics of Hearing and Balance in Zebrafish. Annu. Rev. Genet. 2005, 39, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Baxendale, S.; Whitfield, T.T. Zebrafish Inner Ear Development and Function. In Development of Auditory and Vestibular Systems, 4th ed.; Romand, R., Varela-Nieto, I., Eds.; Academic Press: Amsterdam, The Netherlands, 2014; pp. 63–105. [Google Scholar] [CrossRef]
- Riley, B.B.; Moorman, S.J. Development of utricular otoliths, but not saccular otoliths, is necessary for vestibular function and survival in zebrafish. J. Neurobiol. 2015, 43, 329–337. [Google Scholar] [CrossRef]
- Miyai, M.; Tsunekage, Y.; Saito, M.; Kohno, K.; Takahashi, K.; Kataoka, K. Ectopic expression of the transcription factor MafB in basal keratinocytes induces hyperproliferation and perturbs epidermal homeostasis. Exp. Dermatol. 2017, 26, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Hamze, Z.; Bonnavion, R.; Herath, N.; Pouponnot, C.; Assade, F.; Fontaniere, S.; Bertolino, P.; Cordierbussat, M.; Zhang, C.X. Reexpression of oncoprotein MafB in proliferative β-cells and Men1 insulinomas in mouse. Oncogene 2011, 31, 3647–3654. [Google Scholar] [CrossRef] [PubMed]
- Karbaschi, M.; Ji, Y.; Abdulwahed, A.M.S.; Alohaly, A.; Bedoya, J.F.; Burke, S.L.; Boulos, T.M.; Tempest, H.G.; Cooke, M. Evaluation of the Major Steps in the Conventional Protocol for the Alkaline Comet Assay. Int. J. Mol. Sci. 2019, 20, 6072. [Google Scholar] [CrossRef] [Green Version]
- Cordes, S.P.; Barsh, G.S. The mouse segmentation gene kr encodes a novel basic domain-leucine zipper transcription factor. Cell 1994, 79, 1025–1034. [Google Scholar] [CrossRef]
- Schvarzstein, M.; Kirn, A.; Haffter, P.; Cordes, S.P. Expression of Zkrml2, a homologue of the Krml1/val segmentation gene, during embryonic patterning of the zebrafish (Danio rerio). Mech. Dev. 1999, 80, 223–226. [Google Scholar] [CrossRef]
- Katzenback, B.A.; Karpman, M.; Belosevic, M. Distribution and expression analysis of transcription factors in tissues and progenitor cell populations of the goldfish (Carassius auratus L.) in response to growth factors and pathogens. Mol. Immunol. 2011, 48, 1224–1235. [Google Scholar] [CrossRef]
- Sadl, V.S.; Jin, F.; Yu, J.; Cui, S.; Holmyard, D.; Quaggin, S.E.; Barsh, G.S.; Cordes, S.P. The Mouse Kreisler (Krml1/MafB) Segmentation Gene Is Required for Differentiation of Glomerular Visceral Epithelial Cells. Dev. Biol. 2002, 249, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Koltowska, K.; Paterson, S.; Bower, N.I.; Baillie, G.J.; Lagendijk, A.K.; Astin, J.W.; Chen, H.; Francois, M.; Crosier, P.S.; Taft, R.J. mafba is a downstream transcriptional effector of Vegfc signaling essential for embryonic lymphangiogenesis in zebrafish. Genes Dev. 2015, 29, 1618–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, F.-S.; Abbas, L.; Baxendale, S.; Holdsworth, C.J.; Swanson, A.G.; Slanchev, K.; Hammerschmidt, M.; Topczewski, J.; Whitfield, T.T. Semicircular canal morphogenesis in the zebrafish inner ear requires the function of gpr126 (lauscher), an adhesion class G protein-coupled receptor gene. Development 2013, 140, 4362–4374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clément, A.; Blanco-Sánchez, B.; Peirce, J.L.; Westerfield, M. Cog4 is required for protrusion and extension of the epithelium in the developing semicircular canals. Mech. Dev. 2018, 155, 1–7. [Google Scholar] [CrossRef]
- Navis, A.; Marjoram, L.; Bagnat, M. Cftr controls lumen expansion and function of Kupffer’s vesicle in zebrafish. Development 2013, 140, 1703–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosaliganti, K.R.; Swinburne, I.A.; Chan, C.U.; Obholzer, N.D.; A Green, A.; Tanksale, S.; Mahadevan, L.; Megason, S.G. Size control of the inner ear via hydraulic feedback. eLife 2019, 8. [Google Scholar] [CrossRef]
- Choo, D.; Ward, J.; Reece, A.; Dou, H.; Lin, Z.; Greinwald, J. Molecular mechanisms underlying inner ear patterning defects in kreisler mutants. Dev. Biol. 2006, 289, 308–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zannino, D.A.; Sagerström, C.G.; Appel, B. olig2-expressing hindbrain cells are required for migrating facial motor neurons. Dev. Dyn. 2011, 241, 315–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, P.; Maurer, J.M.; Sagerström, C.G. Analysis of novel caudal hindbrain genes reveals different regulatory logic for gene expression in rhombomere 4 versus 5/6 in embryonic zebrafish. Neural Dev. 2018, 13, 1–24. [Google Scholar] [CrossRef]
- Asakawa, K.; Kawakami, K. Protocadherin-Mediated Cell Repulsion Controls the Central Topography and Efferent Projections of the Abducens Nucleus. Cell Rep. 2018, 24, 1562–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatch, E.P.; Noyes, C.A.; Wang, X.; Wright, T.J.; Mansour, S.L. Fgf3 is required for dorsal patterning and morphogenesis of the inner ear epithelium. Development 2007, 134, 3615–3625. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Pajares, V.; Qu, K.; Zhang, J.; Webster, D.E.; Barajas, B.C.; Siprashvili, Z.; Zarnegar, B.J.; Boxer, L.; Rios, E.J.; Tao, S.; et al. A LncRNA-MAF:MAFB Transcription Factor Network Regulates Epidermal Differentiation. Dev. Cell 2015, 32, 693–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labott, A.T.; Lopez-Pajares, V. Epidermal differentiation gene regulatory networks controlled by MAF and MAFB. Cell Cycle 2016, 15, 1405–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.-J.; Hwang, P.-P. Development of zebrafish epidermis. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 205–214. [Google Scholar] [CrossRef]
- Sancar, A.; Lindsey-Boltz, L.A.; Ünsal-Kaçmaz, K.; Linn, S. Molecular Mechanisms of Mammalian DNA Repair and the DNA Damage Checkpoints. Annu. Rev. Biochem. 2004, 73, 39–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Lan, X.; Li, D.; Li, T.; Lu, S. MiR-223 targeting MAFB suppresses proliferation and migration of nasopharyngeal carcinoma cells. BMC Cancer 2015, 15, 461. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Huang, Y.; Gao, P.; Lv, Y.; Jia, D.; Sun, K.; Han, Y.; Hu, H.; Tang, Z.; Ren, X.; et al. Knockout of mafba Causes Inner-Ear Developmental Defects in Zebrafish via the Impairment of Proliferation and Differentiation of Ionocyte Progenitor Cells. Biomedicines 2021, 9, 1699. https://doi.org/10.3390/biomedicines9111699
Chen X, Huang Y, Gao P, Lv Y, Jia D, Sun K, Han Y, Hu H, Tang Z, Ren X, et al. Knockout of mafba Causes Inner-Ear Developmental Defects in Zebrafish via the Impairment of Proliferation and Differentiation of Ionocyte Progenitor Cells. Biomedicines. 2021; 9(11):1699. https://doi.org/10.3390/biomedicines9111699
Chicago/Turabian StyleChen, Xiang, Yuwen Huang, Pan Gao, Yuexia Lv, Danna Jia, Kui Sun, Yunqiao Han, Hualei Hu, Zhaohui Tang, Xiang Ren, and et al. 2021. "Knockout of mafba Causes Inner-Ear Developmental Defects in Zebrafish via the Impairment of Proliferation and Differentiation of Ionocyte Progenitor Cells" Biomedicines 9, no. 11: 1699. https://doi.org/10.3390/biomedicines9111699