
The Role of miRNAs in Neuropathic Pain
 and
and
Abstract
:1. Introduction
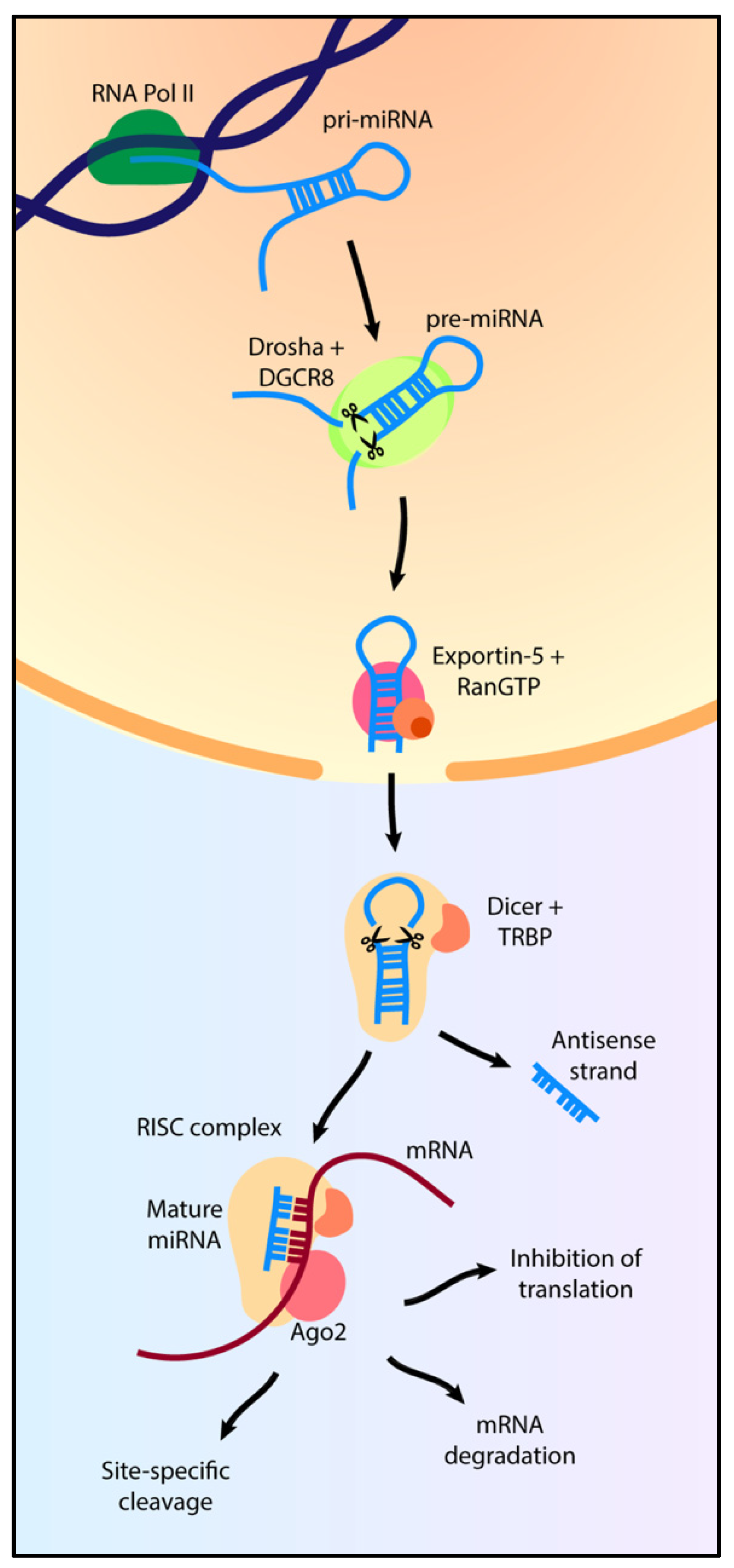
2. MicroRNA Biogenesis
3. MicroRNA Involvement in Pain Pathways
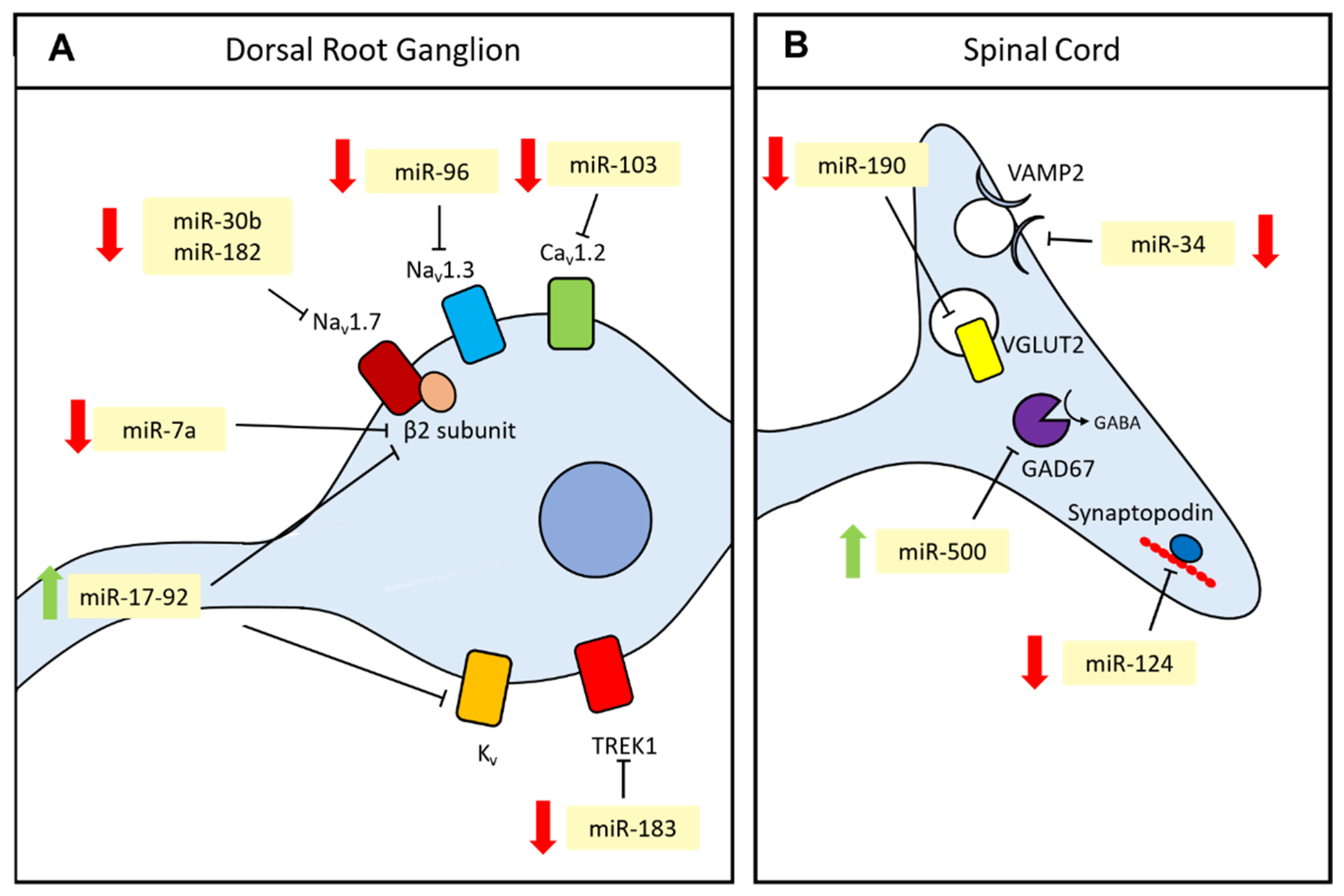
3.1. Neuronal Excitability
3.2. Synaptic Transmission
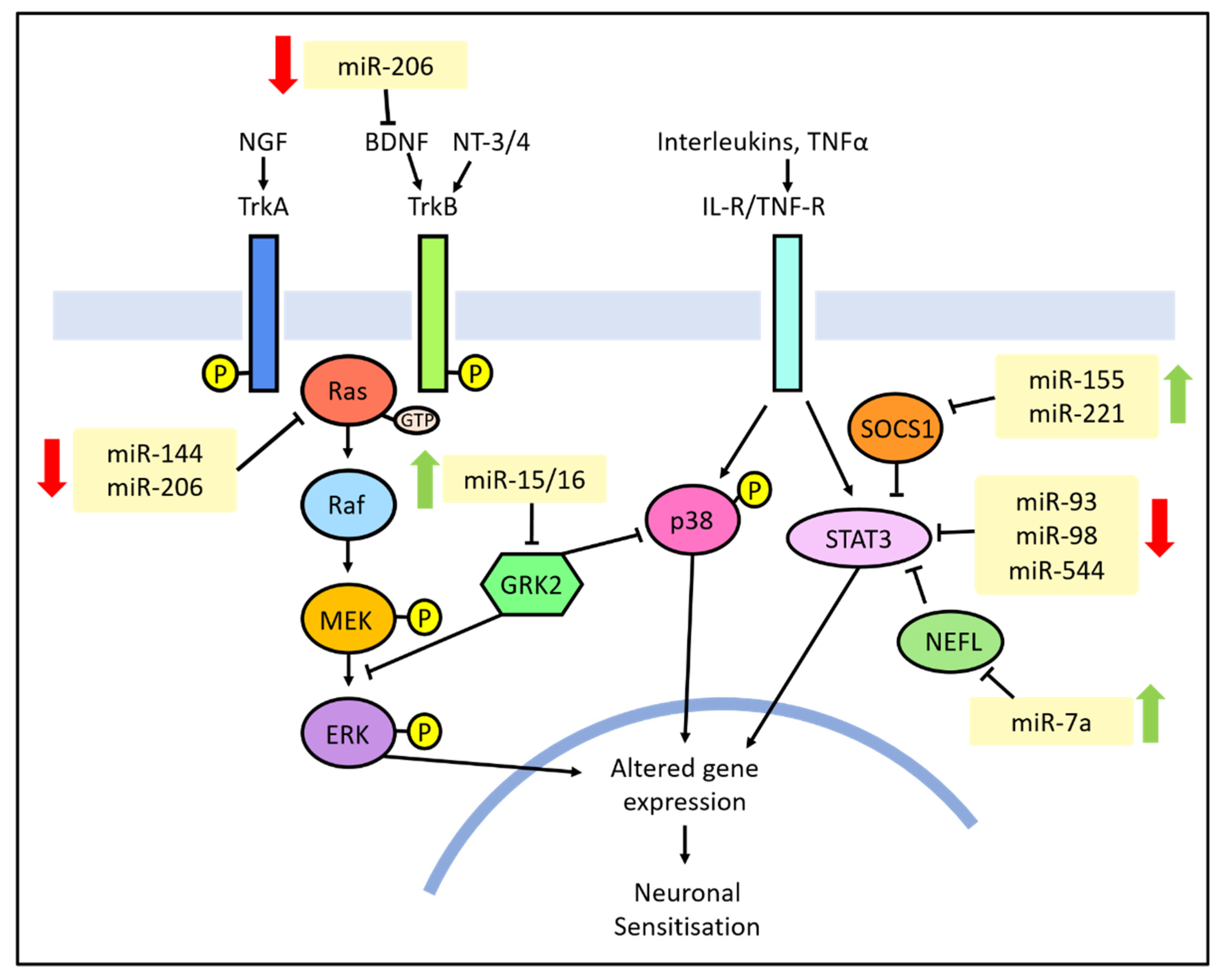
3.3. Intracellular Signalling
3.4. Interaction with Non-Neuronal Cell Types
3.4.1. Macrophages Communicate through miRNA-Containing Extracellular Vesicles
3.4.2. Schwann Cell Proliferation Is Directed by miRNA
3.4.3. Lymphocyte Differentiation Is Altered by miRNA Expression
3.4.4. Microglial Activation Is Regulated by miRNA
3.4.5. Astrocytic Activation and Cytokine Production Is Influenced by miRNA
4. A Closer Look at miRNA Regulation and Function
5. Potential Clinical Application of miRNA Studies
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpää, M.; Hansson, P.; Jensen, T.S.; et al. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol. 2015, 14, 162–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannini, S.; Coraci, D.; Brau, F.; Galluzzo, V.; Loreti, C.; Caliandro, P.; Padua, L.; Maccauro, G.; Biscotti, L.; Bernabei, R. Neuropathic Pain in the Elderly. Diagnostics 2021, 11, 613. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Kristen, A.V.; Ajroud-Driss, S.; Conceição, I.; Gorevic, P.; Kyriakides, T.; Obici, L. Patisiran, an RNAi therapeutic for the treatment of hereditary transthyretin-mediated amyloidosis. Neurodegener. Dis. Manag. 2019, 9, 5–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, H.H.; Duroux, M.; Gazerani, P. MicroRNAs as modulators and biomarkers of inflammatory and neuropathic pain conditions. Neurobiol. Dis. 2014, 71, 159–168. [Google Scholar] [CrossRef] [Green Version]
- López-González, M.J.; Landry, M.; Favereaux, A. MicroRNA and chronic pain: From mechanisms to therapeutic potential. Pharmacol. Ther. 2017, 180, 1–15. [Google Scholar] [CrossRef]
- Bai, G.; Ambalavanar, R.; Wei, D.; Dessem, D. Downregulation of Selective microRNAs in Trigeminal Ganglion Neurons Following Inflammatory Muscle Pain. Mol. Pain 2007, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Lee, M.-C.; Momin, A.; Cendan, C.-M.; Shepherd, S.T.; Baker, M.D.; Asante, C.; Bee, L.; Bethry, A.; Perkins, J.R.; et al. Small RNAs Control Sodium Channel Expression, Nociceptor Excitability, and Pain Thresholds. J. Neurosci. 2010, 30, 10860–10871. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, D.E.; Martin, M.M.; Feldman, D.S.; Terry, A.V.; Nuovo, G.J.; Elton, T.S. Experimental validation of miRNA targets. Methods 2008, 44, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Zhang, Y.; Liu, Y.; Liu, H.; Zhang, Z.; Su, Z. Effects of miR-150 on neuropathic pain process via targeting AKT3. Biochem. Biophys. Res. Commun. 2019, 517, 532–537. [Google Scholar] [CrossRef]
- You, H.; Zhang, L.; Chen, Z.; Liu, W.; Wang, H.; He, H. MiR-20b-5p relieves neuropathic pain by targeting Akt3 in a chronic constriction injury rat model. Synapse 2019, 73, e22125. [Google Scholar] [CrossRef]
- Shi, G.; Shi, J.; Liu, K.; Liu, N.; Wang, Y.; Fu, Z.; Ding, J.; Jia, L.; Yuan, W. Increased miR-195 aggravates neuropathic pain by inhibiting autophagy following peripheral nerve injury. Glia 2013, 61, 504–512. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Wang, X.; Zhang, J.; Xie, C. Extracellular vesicle-encapsulated microRNA-23a from dorsal root ganglia neurons binds to A20 and promotes inflammatory macrophage polarization following peripheral nerve injury. Aging 2021, 13, 6752–6764. [Google Scholar] [CrossRef]
- Li, X.; Wang, S.; Yang, X.; Chu, H. miR-142-3p targets AC9 to regulate sciatic nerve injury-induced neuropathic pain by regulating the cAMP/AMPK signalling pathway. Int. J. Mol. Med. 2021, 47, 561–572. [Google Scholar] [CrossRef]
- Gandla, J.; Lomada, S.K.; Lu, J.; Kuner, R.; Bali, K.K. miR-34c-5p functions as pronociceptive microRNA in cancer pain by targeting Cav2.3 containing calcium channels. Pain 2017, 158, 1765–1779. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Zhu, L.-J.; Li, Y.-Q.; Hao, L.-Y.; Yin, C.; Yang, J.-X.; Guo, Y.; Zhang, S.; Hua, L.; Xue, Z.-Y.; et al. Epigenetic Modification of Spinal miR-219 Expression Regulates Chronic Inflammation Pain by Targeting CaMKII gamma. J. Neurosci. 2014, 34, 9476–9483. [Google Scholar] [CrossRef] [Green Version]
- Favereaux, A.; Thoumine, O.; Bouali-Benazzouz, R.; Roques, V.; Papon, M.-A.; Salam, S.A.; Drutel, G.; Léger, C.; Calas, A.; Nagy, F.; et al. Bidirectional integrative regulation of Cav1.2 calcium channel by microRNA miR-103: Role in pain. EMBO J. 2011, 30, 3830–3841. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.-C.; Cao, D.-L.; Zhang, X.; Zhang, Z.-J.; He, L.-N.; Li, C.-H.; Zhang, W.-W.; Wu, X.-B.; Berta, T.; Ji, R.-R.; et al. CXCL13 drives spinal astrocyte activation and neuropathic pain via CXCR5. J. Clin. Investig. 2016, 126, 745–761. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Shan, Q.; Gu, P.; Wang, X.M.; Tai, L.W.; Sun, M.; Luo, X.; Sun, L.; Cheung, C.W. miRNA-23a/CXCR4 regulates neuropathic pain via directly targeting TXNIP/NLRP3 inflammasome axis. J. Neuroinflamm. 2018, 15, 29. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Zhang, X.; Wang, X.; Xu, F.; Zhang, J.; Li, L.; Xie, X.; Wang, L.; Yang, Y.; Xu, J. MicroRNA-124-3p attenuates the development of nerve injury–induced neuropathic pain by targeting early growth response 1 in the dorsal root ganglia and spinal dorsal horn. J. Neurochem. 2021, 158, 928–942. [Google Scholar] [CrossRef]
- Tan, M.; Shen, L.; Hou, Y. Epigenetic modification of BDNF mediates neuropathic pain via miR-30a-3p/EP300 axis in CCI rats. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhou, J.; Sun, C. MicroRNA-211-5p Enhances Analgesic Effect of Dexmedetomidine on Inflammatory Visceral Pain in Rats by Suppressing ERK Signaling. J. Mol. Neurosci. 2019, 68, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, H.; An, L.; Li, L.; Wei, M.; Ge, D.; Su, Z. miR-124-3p attenuates neuropathic pain induced by chronic sciatic nerve injury in rats via targeting EZH2. J. Cell. Biochem. 2018, 120, 5747–5755. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, Q.; Shen, J.; Wang, L.; Cai, Y.; Zhu, K. miR-194 relieve neuropathic pain and prevent neuroinflammation via targeting FOXA1. J. Cell. Biochem. 2020, 121, 3278–3285. [Google Scholar] [CrossRef]
- Huang, Z.-Z.; Wei, J.-Y.; Ou-Yang, H.-D.; Li, D.; Xu, T.; Wu, S.-L.; Zhang, X.-L.; Liu, C.-C.; Ma, C.; Xin, W.-J. mir-500-Mediated GAD67 Downregulation Contributes to Neuropathic Pain. J. Neurosci. 2016, 36, 6321–6331. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Wan, Y.; Sun, L.; Tao, S.; Chen, P.; Liu, C.; Wang, K.; Zhou, C.; Zhao, G. Inhibition of MicroRNA-15a/16 Expression Alleviates Neuropathic Pain Development through Upregulation of G Protein-Coupled Receptor Kinase 2. Biomol. Ther. 2019, 27, 414–422. [Google Scholar] [CrossRef]
- Ma, X.; Wang, H.; Song, T.; Wang, W.; Zhang, Z. lncRNA MALAT1 contributes to neuropathic pain development through regulating miR-129-5p/HMGB1 axis in a rat model of chronic constriction injury. Int. J. Neurosci. 2020, 130, 1215–1224. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Zi, T. Overexpression of microRNA-141 relieves chronic constriction injury-induced neuropathic pain via targeting high-mobility group box 1. Int. J. Mol. Med. 2015, 36, 1433–1439. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Mou, J.; Cao, L.; Zhen, S.; Huang, H.; Bao, H. MicroRNA-142-3p relieves neuropathic pain by targeting high mobility group box 1. Int. J. Mol. Med. 2017, 41, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Guo, Y.; Chen, Q.; Xiong, Q.; Min, S. MicroRNA-193a Downregulates HMGB1 to Alleviate Diabetic Neuropathic Pain in a Mouse Model. Neuroimmunomodulation 2019, 26, 250–257. [Google Scholar] [CrossRef]
- Xia, L.-X.; Ke, C.; Lu, J.-M. NEAT1 contributes to neuropathic pain development through targeting miR-381/HMGB1 axis in CCI rat models. J. Cell. Physiol. 2018, 233, 7103–7111. [Google Scholar] [CrossRef]
- Zhan, L.-Y.; Lei, S.-Q.; Zhang, B.-H.; Li, W.-L.; Wang, H.-X.; Zhao, B.; Cui, S.-S.; Ding, H.; Huang, Q.-M. Overexpression of miR-381 relieves neuropathic pain development via targeting HMGB1 and CXCR4. Biomed. Pharmacother. 2018, 107, 818–823. [Google Scholar] [CrossRef]
- Liu, C.C.; Cheng, J.T.; Li, T.Y.; Tan, P.H. Integrated analysis of microRNA and mRNA expression profiles in the rat spinal cord under inflammatory pain conditions. Eur. J. Neurosci. 2017, 46, 2713–2728. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, F.; Wei, M.; Qiu, Y.; Ma, C.; Shen, L.; Huang, Y. Chronic constriction injury-induced microRNA-146a-5p alleviates neuropathic pain through suppression of IRAK1/TRAF6 signaling pathway. J. Neuroinflamm. 2018, 15, 179. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Zeng, J.; Liu, Y.; Chen, J.; Zeng, Q.-C.; Chen, Y.-Q.; Tu, L.-L.; Chen, P.; Yang, F.; Zhang, M. microRNA-9 and -29a regulate the progression of diabetic peripheral neuropathy via ISL1-mediated sonic hedgehog signaling pathway. Aging 2020, 12, 11446–11465. [Google Scholar] [CrossRef]
- Li, H.; Fan, L.; Zhang, Y.; Cao, Y.; Liu, X. SNHG16 aggravates chronic constriction injury-induced neuropathic pain in rats via binding with miR-124-3p and miR-141-3p to upregulate JAG1. Brain Res. Bull. 2020, 165, 228–237. [Google Scholar] [CrossRef]
- Sakai, A.; Saitow, F.; Maruyama, M.; Miyake, N.; Miyake, K.; Shimada, T.; Okada, T.; Suzuki, H. MicroRNA cluster miR-17-92 regulates multiple functionally related voltage-gated potassium channels in chronic neuropathic pain. Nat. Commun. 2017, 8, 16079. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Rong, L.; Shao, J.; Zhang, Y.; Liu, Y.; Zhao, S.; Li, L.; Yu, W.; Zhang, M.; Ren, X.; et al. Epigenetic restoration of voltage-gated potassium channel Kv1.2 alleviates nerve injury-induced neuropathic pain. J. Neurochem. 2021, 156, 367–378. [Google Scholar] [CrossRef]
- Wang, W.; Li, R. MiR-216a-5p alleviates chronic constriction injury-induced neuropathic pain in rats by targeting KDM3A and inactivating Wnt/β-catenin signaling pathway. Neurosci. Res. 2020, 170, 255–264. [Google Scholar] [CrossRef]
- Tozaki-Saitoh, H.; Masuda, J.; Kawada, R.; Kojima, C.; Yoneda, S.; Masuda, T.; Inoue, K.; Tsuda, M. Transcription factor MafB contributes to the activation of spinal microglia underlying neuropathic pain development. Glia 2018, 67, 729–740. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, Z.; Liu, H.-L.; Li, L.; Wei, M.; Ge, D.-J.; Zhang, Z.-J. Effects of miR-26a-5p on neuropathic pain development by targeting MAPK6 in in CCI rat models. Biomed. Pharmacother. 2018, 107, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Guo, S.; Zhang, Y.; Lin, P.; Lin, C.; Chen, M.; Zhu, S.; Huang, L.; He, J.; Zhang, L.; et al. MiR-223-3p alleviates trigeminal neuropathic pain in the male mouse by targeting MKNK2 and MAPK/ERK signaling. Brain Behav. 2022, 12, e2634. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Liu, B.; Mo, Y.; Wang, X.; Zhong, L.; Han, X.; Mi, F. MiR-101 promotes pain hypersensitivity in rats with chronic constriction injury via the MKP-1 mediated MAPK pathway. J. Cell. Mol. Med. 2020, 24, 8986–8997. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Ma, L.; Xi, K.; Zhang, W.; Fan, D. MicroRNA-183 Suppresses Neuropathic Pain and Expression of AMPA Receptors by Targeting mTOR/VEGF Signaling Pathway. Cell. Physiol. Biochem. 2017, 41, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Li, P.; Ni, Y.; Zhao, J.; Liu, Z. Decreased MicroRNA-125a-3p Contributes to Upregulation of p38 MAPK in Rat Trigeminal Ganglions with Orofacial Inflammatory Pain. PLoS ONE 2014, 9, e111594. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, H.; Zhang, T.; He, M.; Liang, H.; Wang, H.; Xu, L.; Chen, S.; Xu, M. Inhibition of MicroRNA-195 Alleviates Neuropathic Pain by Targeting Patched1 and Inhibiting SHH Signaling Pathway Activation. Neurochem. Res. 2019, 44, 1690–1702. [Google Scholar] [CrossRef]
- Wan, L.; Su, Z.; Li, F.; Gao, P.; Zhang, X. MiR-122-5p suppresses neuropathic pain development by targeting PDK4. Neurochem. Res. 2021, 46, 957–963. [Google Scholar] [CrossRef]
- Pan, Z.; Li, G.-F.; Sun, M.-L.; Xie, L.; Liu, D.; Zhang, Q.; Yang, X.-X.; Xia, S.; Liu, X.; Zhou, H.; et al. MicroRNA-1224 Splicing CircularRNA-Filip1l in an Ago2-Dependent Manner Regulates Chronic Inflammatory Pain via Targeting Ubr5. J. Neurosci. 2019, 39, 2125–2143. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Guo, S.; Wang, S. MicroRNA-16 Alleviates Inflammatory Pain by Targeting Ras-Related Protein 23 (RAB23) and Inhibiting p38 MAPK Activation. Experiment 2016, 22, 3894–3901. [Google Scholar] [CrossRef] [Green Version]
- Fang, B.; Wei, L.; Dong, K.; Niu, X.; Sui, X.; Zhang, H. miR-202 modulates the progression of neuropathic pain through targeting RAP1A. J. Cell. Biochem. 2018, 120, 2973–2982. [Google Scholar] [CrossRef]
- Wu, Y.; Gu, Y.; Shi, B. miR-590-3p Alleviates diabetic peripheral neuropathic pain by targeting RAP1A and suppressing infiltration by the T cells. Acta Biochim. Pol. 2020, 67, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Guo, H.; Xie, A.; Liao, O.; Ju, F.; Zhou, Y. MicroRNA-144 relieves chronic constriction injury-induced neuropathic pain via targeting RASA. Biotechnol. Appl. Biochem. 2019, 67, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, Y.; Ma, Z.; Liu, Y.; Sun, Z.; Wu, Y. miR-140 ameliorates neuropathic pain in CCI rats by targeting S1PR1. J. Recept. Signal Transduct. Res. 2020, 41, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-P.; Zhou, W.; Kang, L.-M.; Yan, H.; Zhang, L.; Xu, B.-H.; Cai, W.-H. Intrathecal miR-96 Inhibits Nav1.3 Expression and Alleviates Neuropathic Pain in Rat Following Chronic Construction Injury. Neurochem. Res. 2013, 39, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Zhang, Y.; Zhao, J.; Chen, Y.; Kong, L.; Sheng, C.; Yuan, L. miR-384-5p ameliorates neuropathic pain by targeting SCN3A in a rat model of chronic constriction injury. Neurol. Res. 2020, 42, 299–307. [Google Scholar] [CrossRef]
- Jia, Q.; Dong, W.; Zhang, L.; Yang, X. Activating Sirt1 by resveratrol suppresses Nav1.7 expression in DRG through miR-182 and alleviates neuropathic pain in rats. Channels 2020, 14, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Zhao, Q.; Shao, J.; Zhang, J.; Li, L.; Ren, X.; Su, S.; Bai, Q.; Li, M.; Chen, X.; et al. MicroRNA-182 Alleviates Neuropathic Pain by Regulating Nav1.7 Following Spared Nerve Injury in Rats. Sci. Rep. 2018, 8, 16750. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.; Cao, J.; Wang, J.; Ren, X.; Su, S.; Li, M.; Li, Z.; Zhao, Q.; Zang, W. MicroRNA-30b regulates expression of the sodium channel Nav1.7 in nerve injury-induced neuropathic pain in the rat. Mol. Pain 2016, 12, 671523. [Google Scholar] [CrossRef]
- Chen, S.; Gu, Y.; Dai, Q.; He, Y.; Wang, J. Spinal miR-34a regulates inflammatory pain by targeting SIRT1 in complete Freund’s adjuvant mice. Biochem. Biophys. Res. Commun. 2019, 516, 1196–1203. [Google Scholar] [CrossRef]
- Chu, Y.; Ge, W.; Wang, X. MicroRNA-448 modulates the progression of neuropathic pain by targeting sirtuin 1. Exp. Ther. Med. 2019, 18, 4665–4672. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Yang, Q.; Wei, X.; Liu, Y.; Ma, D.; Li, J.; Wan, Y.; Luo, Y. The role of miR-190a-5p contributes to diabetic neuropathic pain via targeting SLC17A6. J. Pain Res. 2017, 10, 2395–2403. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.G.; He, H.; Wang, P. A critical role for miR-135a-5p-mediated regulation of SLC24A2 in neuropathic pain. Mol. Med. Rep. 2020, 22, 2115–2122. [Google Scholar] [CrossRef]
- Tan, Y.; Yang, J.; Xiang, K.; Tan, Q.; Guo, Q. Suppression of MicroRNA-155 Attenuates Neuropathic Pain by Regulating SOCS1 Signalling Pathway. Neurochem. Res. 2014, 40, 550–560. [Google Scholar] [CrossRef]
- Xia, L.; Zhang, Y.; Dong, T. Inhibition of MicroRNA-221 Alleviates Neuropathic Pain Through Targeting Suppressor of Cytokine Signaling 1. J. Mol. Neurosci. 2016, 59, 411–420. [Google Scholar] [CrossRef]
- Li, L.; Zhao, G. Downregulation of microRNA-218 relieves neuropathic pain by regulating suppressor of cytokine signaling International 3. J. Mol. Med. 2016, 37, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.-T.; Ji, L.-J.; Wang, Z.; Wu, X.; Wang, Q.; Sun, S.; Lu, J.-M.; Zhang, Y. MicroRNA-93 alleviates neuropathic pain through targeting signal transducer and activator of transcription 3. Int. Immunopharmacol. 2017, 46, 156–162. [Google Scholar] [CrossRef]
- Zhong, L.; Fu, K.; Xiao, W.; Wang, F.; Shen, L. Overexpression of miR-98 attenuates neuropathic pain development via targeting STAT3 in CCI rat models. J. Cell. Biochem. 2018, 120, 7989–7997. [Google Scholar] [CrossRef]
- Jin, H.; Du, X.; Zhao, Y.; Xia, D. XIST/miR-544 axis induces neuropathic pain by activating STAT3 in a rat model. J. Cell. Physiol. 2018, 233, 5847–5855. [Google Scholar] [CrossRef]
- Elramah, S.; López-González, M.J.; Bastide, M.; Dixmérias, F.; Roca-Lapirot, O.; Wielanek-Bachelet, A.C.; Vital, A.; Leste-Lasserre, T.; Brochard, A.; Landry, M.; et al. Spinal miRNA-124 regulates synaptopodin and nociception in an animal model of bone cancer pain. Sci. Rep. 2017, 7, 10949. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Wang, S.; Xie, Y.; Jin, K.; Bai, Y.; Shan, S. MiR-28-5p relieves neuropathic pain by targeting Zeb1 in CCI rat models. J. Cell. Biochem. 2018, 119, 8555–8563. [Google Scholar] [CrossRef]
- Tramullas, M.; Francés, R.; de la Fuente, R.; Velategui, S.; Carcelén, M.; García, R.; Llorca, J.; Hurlé, M.A. MicroRNA-30c-5p modulates neuropathic pain in rodents. Sci. Transl. Med. 2018, 10, eaao6299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Zhang, H. miR-451 elevation relieves inflammatory pain by suppressing microglial activation-evoked inflammatory response via targeting TLR4. Cell Tissue Res. 2018, 374, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Li, L.; Zhang, Y.; Zhang, Z.; Liu, H.; Bao, H. LncRNA X inactive specific transcript contributes to neuropathic pain development by sponging miR-154-5p via inducing toll-like receptor 5 in CCI rat models. J. Cell. Biochem. 2018, 120, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-J.; Guo, J.-S.; Li, S.-S.; Wu, X.-B.; Cao, D.-L.; Jiang, B.-C.; Jing, P.-B.; Bai, X.-Q.; Li, C.-H.; Wu, Z.-H.; et al. TLR8 and its endogenous ligand miR-21 contribute to neuropathic pain in murine DRG. J. Exp. Med. 2018, 215, 3019–3037. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.-N.; Yuan, Y.-T.; Ye, D.; Kang, L.-M.; Wen, J.; Chen, H.-P. MiR-183-5p Alleviates Chronic Constriction Injury-Induced Neuropathic Pain Through Inhibition of TREK-1. Neurochem. Res. 2018, 43, 1143–1149. [Google Scholar] [CrossRef]
- Miao, J.; Zhou, X.; Ji, T.; Chen, G. NF-κB p65-dependent transcriptional regulation of histone deacetylase 2 contributes to the chronic constriction injury-induced neuropathic pain via the microRNA-183/TXNIP/NLRP3 axis. J Neuroinflamm. 2020, 17, 225. [Google Scholar] [CrossRef]
- Ji, L.-J.; Su, J.; Xu, A.-L.; Pang, B.; Huang, Q.-M. MiR-134-5p attenuates neuropathic pain progression through targeting Twist1. J. Cell. Biochem. 2019, 120, 1694–1701. [Google Scholar] [CrossRef]
- Brandenburger, T.; Johannsen, L.; Prassek, V.; Kuebart, A.; Raile, J.; Wohlfromm, S.; Köhrer, K.; Huhn, R.; Hollmann, M.W.; Hermanns, H. MiR-34a is differentially expressed in dorsal root ganglia in a rat model of chronic neuropathic pain. Neurosci. Lett. 2019, 708, 134365. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Cai, W.; Liu, Y.; Liu, H.; Zhang, Z.; Su, Z. MicroRNA-128-3p Alleviates Neuropathic Pain Through Targeting ZEB1. Neurosci. Lett. 2020, 729, 134946. [Google Scholar] [CrossRef]
- Yan, X.; Lu, J.; Wang, Y.; Cheng, X.; He, X.; Zheng, W.; Chen, H.; Wang, Y. XIST accelerates neuropathic pain progression through regulation of miR-150 and ZEB1 in CCI rat models. J. Cell. Physiol. 2018, 233, 6098–6106. [Google Scholar] [CrossRef]
- Chen, Z.-L.; Liu, J.-Y.; Wang, F.; Jing, X. Suppression of MALAT1 ameliorates chronic constriction injury-induced neuropathic pain in rats via modulating miR-206 and ZEB2. J. Cell. Physiol. 2019, 234, 15647–15653. [Google Scholar] [CrossRef]
- Łuczkowska, K.; Rogińska, D.; Ulańczyk, Z.; Safranow, K.; Paczkowska, E.; Baumert, B.; Milczarek, S.; Osękowska, B.; Górska, M.; Borowiecka, E.; et al. microRNAs as the biomarkers of chemotherapy-induced peripheral neuropathy in patients with multiple myeloma. Leuk Lymphoma 2021, 62, 2768–2776. [Google Scholar] [CrossRef]
- McDonald, M.K.; Tian, Y.; Qureshi, R.A.; Gormley, M.; Ertel, A.; Gao, R.; Lopez, E.A.; Alexander, G.M.; Sacan, A.; Fortina, P.; et al. Functional significance of macrophage-derived exosomes in inflammation and pain. Pain 2014, 155, 1527–1539. [Google Scholar] [CrossRef] [Green Version]
- Orlova, I.A.; Alexander, G.M.; Qureshi, R.A.; Sacan, A.; Graziano, A.; Barrett, J.E.; Schwartzman, R.J.; Ajit, S.K. MicroRNA modulation in complex regional pain syndrome. J. Transl. Med. 2011, 9, 195. [Google Scholar] [CrossRef] [Green Version]
- Ciccacci, C.; Latini, A.; Colantuono, A.; Politi, C.; D’Amato, C.; Greco, C.; Rinaldi, M.E.; Lauro, D.; Novelli, G.; Spallone, V.; et al. Expression study of candidate miRNAs and evaluation of their potential use as biomarkers of diabetic neuropathy. Epigenomics 2020, 12, 575–585. [Google Scholar] [CrossRef]
- Li, Y.B.; Wu, Q.; Liu, J.; Fan, Y.Z.; Yu, K.F.; Cai, Y. miR-199a-3p is involved in the pathogenesis and progression of diabetic neuropathy through downregulation of SerpinE2. Mol. Med. Rep. 2017, 16, 2417–2424. [Google Scholar] [CrossRef] [Green Version]
- Asahchop, E.L.; Branton, W.G.; Krishnan, A.; Chen, P.A.; Yang, D.; Kong, L.L.; Zochodne, D.W.; Brew, B.; John, G.M.; Power, D.C. HIV-associated sensory polyneuropathy and neuronal injury are associated with miRNA–455-3p induction. J. Clin. Investig. 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Tavares-Ferreira, D.; Lawless, N.; Bird, E.V.; Atkins, S.; Collier, D.; Sher, E.; Malki, K.; Lambert, D.W.; Boissonade, F.M. Correlation of miRNA expression with intensity of neuropathic pain in man. Mol. Pain 2019, 15. [Google Scholar] [CrossRef] [Green Version]
- Dayer, C.F.; Luthi, F.; Le Carré, J.; Vuistiner, P.; Terrier, P.; Benaim, C.; Giacobino, J.-P.; Léger, B. Differences in the miRNA signatures of chronic musculoskeletal pain patients from neuropathic or nociceptive origins. PLoS ONE 2019, 14, e0219311. [Google Scholar] [CrossRef] [Green Version]
- Heyn, J.; Luchting, B.; Hinske, L.C.; Hübner, M.; Azad, S.C.; Kreth, S. miR-124a and miR-155 enhance differentiation of regulatory T cells in patients with neuropathic pain. J. Neuroinflamm. 2016, 13, 248. [Google Scholar] [CrossRef] [Green Version]
- Leinders, M.; Üçeyler, N.; Pritchard, R.; Sommer, C.; Sorkin, L. Increased miR-132-3p expression is associated with chronic neuropathic pain. Exp. Neurol. 2016, 283, 276–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.C.; Xue, D.F.; Wang, X.Q.; Ai, D.B.; Qin, P.J. MiR-101 relates to chronic peripheral neuropathic pain through targeting KPNB1 and regulating NF-κB signaling. Kaohsiung J. Med. Sci. 2019, 35, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Leinders, M.; Üçeyler, N.; Thomann, A.; Sommer, C. Aberrant microRNA expression in patients with painful peripheral neuropathies. J. Neurol. Sci. 2017, 380, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, D.; Zhou, J.; Yan, Y.; Chen, L. Evaluation of circulating microRNA expression in patients with trigeminal neuralgia: An observational study. Medicine 2020, 99, e22972. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, Y.; Li, Z. NFKB1 Signalling Activation Contributes to TRPV1 Over-expression via Repressing MiR-375 and MiR-455: A Study on Neuropathic Low Back Pain. Folia Biol. (Praha) 2022, 68, 105–111. [Google Scholar]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Kosik, K.S. The neuronal microRNA system. Nat. Rev. Neurosci. 2006, 7, 911–920. [Google Scholar] [CrossRef]
- Stavast, C.; Erkeland, S. The Non-Canonical Aspects of MicroRNAs: Many Roads to Gene Regulation. Cells 2019, 8, 1465. [Google Scholar] [CrossRef] [Green Version]
- Sakai, A.; Saitow, F.; Miyake, N.; Miyake, K.; Shimada, T.; Suzuki, H. miR-7a alleviates the maintenance of neuropathic pain through regulation of neuronal excitability. Brain 2013, 136, 2738–2750. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-J.; Li, Y. KCNQ potassium channels in sensory system and neural circuits. Acta Pharmacol. Sin. 2015, 37, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Alloui, A.; Zimmermann, K.; Mamet, J.; Duprat, F.; Noël, J.; Chemin, J.; Guy, N.; Blondeau, N.; Voilley, N.; Rubat-Coudert, C.; et al. TREK-1, a K+ channel involved in polymodal pain perception. EMBO J. 2006, 25, 2368–2376. [Google Scholar] [CrossRef] [Green Version]
- Perret, D.; Luo, Z.D. Targeting voltage-gated calcium channels for neuropathic pain management. Neurotherapeutics 2009, 6, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Willemen, H.L.D.M.; Huo, X.-J.; Mao-Ying, Q.-L.; Zijlstra, J.; Heijnen, C.J.; Kavelaars, A. MicroRNA-124 as a novel treatment for persistent hyperalgesia. J. Neuroinflamm. 2012, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Scholz, J.; Broom, D.C.; Youn, D.-H.; Mills, C.D.; Kohno, T.; Suter, M.R.; Moore, K.A.; Decosterd, I.; Coggeshall, R.E.; Woolf, C.J. Blocking Caspase Activity Prevents Transsynaptic Neuronal Apoptosis and the Loss of Inhibition in Lamina II of the Dorsal Horn after Peripheral Nerve Injury. J. Neurosci. 2005, 25, 7317–7323. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, S.M.; Krüger, C.; Park, B.; Derkow, K.; Rosenberger, K.; Baumgart, J.; Trimbuch, T.; Eom, G.; Hinz, M.; Kaul, D.; et al. An unconventional role for miRNA: Let-7 activates Toll-like receptor 7 and causes neurodegeneration. Nat. Neurosci. 2012, 15, 827–835. [Google Scholar] [CrossRef]
- Chen, Y.; Stallings, R.L. Differential Patterns of MicroRNA Expression in Neuroblastoma Are Correlated with Prognosis, Differentiation, and Apoptosis. Cancer Res. 2007, 67, 976–983. [Google Scholar] [CrossRef] [Green Version]
- Yin, K.-J.; Deng, Z.; Huang, H.; Hamblin, M.; Xie, C.; Zhang, J.; Chen, Y.E. miR-497 regulates neuronal death in mouse brain after transient focal cerebral ischemia. Neurobiol. Dis. 2010, 38, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.-K.; Ji, R.-R. Intracellular Signaling in Primary Sensory Neurons and Persistent Pain. Neurochem. Res. 2008, 33, 1970–1978. [Google Scholar] [CrossRef] [Green Version]
- Ji, R.-R.; Gereau, R.W.; Malcangio, M.; Strichartz, G.R. MAP kinase and pain. Brain Res. Rev. 2009, 60, 135–148. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, Z.; Chang, Y. Downregulation of miR-206 contributes to neuropathic pain in rats by enhancing RASA1 expression. Int. J. Clin. Exp. Med. 2016, 9, 3146–3152. [Google Scholar]
- Sun, W.; Zhang, L.; Li, R. Overexpression of miR-206 ameliorates chronic constriction injury-induced neuropathic pain in rats via the MEK/ERK pathway by targeting brain-derived neurotrophic factor. Neurosci. Lett. 2017, 646, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.-R.; Suter, M. p38 MAPK, microglial signaling, and neuropathic pain. Mol. Pain 2007, 3, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Z.; Zhang, J.; Li, J.; Pang, X.; Wang, H. Inhibition of microRNA-155 Reduces Neuropathic Pain During Chemotherapeutic Bortezomib via Engagement of Neuroinflammation. Front. Oncol. 2020, 10, 416. [Google Scholar] [CrossRef]
- Gao, Y.-J.; Ji, R.-R. Activation of JNK pathway in persistent pain. Neurosci. Lett. 2008, 437, 180–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.-R.; Chen, J.; Yi, H.; Peng, L.-Y.; Hu, X.-L.; Guo, Q.-L. MicroRNA-7a ameliorates neuropathic pain in a rat model of spinal nerve ligation via the neurofilament light polypeptide-dependent signal transducer and activator of transcription signaling pathway. Mol. Pain 2019, 15, 842464. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Yu, B.; Qian, T.; Yao, D.; Wang, Y.; Ding, F.; Gu, X. Early changes of microRNAs expression in the dorsal root ganglia following rat sciatic nerve transection. Neurosci. Lett. 2011, 494, 89–93. [Google Scholar] [CrossRef]
- Ji, R.-R.; Chamessian, A.; Zhang, Y.-Q. Pain regulation by non-neuronal cells and inflammation. Science 2016, 354, 572–577. [Google Scholar] [CrossRef] [Green Version]
- Calvo, M.; Dawes, J.M.; Bennett, D.L. The role of the immune system in the generation of neuropathic pain. Lancet Neurol. 2012, 11, 629–642. [Google Scholar] [CrossRef]
- Simeoli, R.; Montague, K.; Jones, H.R.; Castaldi, L.; Chambers, D.; Kelleher, J.H.; Vacca, V.; Pitcher, T.; Grist, J.; Al-Ahdal, H.; et al. Exosomal cargo including microRNA regulates sensory neuron to macrophage communication after nerve trauma. Nat. Commun. 2017, 8, 1778. [Google Scholar] [CrossRef] [Green Version]
- Bhatheja, K.; Field, J. Schwann cells: Origins and role in axonal maintenance and regeneration. Int. J. Biochem. Cell Biol. 2006, 38, 1995–1999. [Google Scholar] [CrossRef]
- Scholz, J.; Woolf, C.J. The neuropathic pain triad: Neurons, immune cells and glia. Nat. Neurosci. 2007, 10, 1361–1368. [Google Scholar] [CrossRef]
- Sohn, E.J.; Park, H.T. MicroRNA Mediated Regulation of Schwann Cell Migration and Proliferation in Peripheral Nerve Injury. BioMed Res. Int. 2018, 2018, 8198365. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, X.; Gu, Y.; Chen, C.; Wang, Y.; Liu, J.; Hu, W.; Yu, B.; Wang, Y.; Ding, F.; et al. Let-7 microRNAs Regenerate Peripheral Nerve Regeneration by Targeting Nerve Growth Factor. Mol. Ther. 2015, 23, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.; Yuan, Y.; Chen, Q.; Wang, X.; Gong, L.; Liu, J.; Gu, X.; Li, S. Regulation of Schwann cell proliferation and migration by miR-1 targeting brain-derived neurotrophic factor after peripheral nerve injury. Sci. Rep. 2016, 6, 29121. [Google Scholar] [CrossRef] [Green Version]
- Viader, A.; Chang, L.-W.; Fahrner, T.; Nagarajan, R.; Milbrandt, J. MicroRNAs Modulate Schwann Cell Response to Nerve Injury by Reinforcing Transcriptional Silencing of Dedifferentiation-Related Genes. J. Neurosci. 2011, 31, 17358–17369. [Google Scholar] [CrossRef] [Green Version]
- Norcini, M.; Sideris, A.; Hernandez, L.A.M.; Zhang, J.; Blanck, T.J.J.; Recio-Pinto, E. An approach to identify microRNAs involved in neuropathic pain following a peripheral nerve injury. Front. Neurosci. 2014, 8, 266. [Google Scholar] [CrossRef] [Green Version]
- Bali, K.K.; Hackenberg, M.; Lubin, A.; Kuner, R.; Devor, M. Sources of Individual Variability: Mirnas That Predispose to Neuropathic Pain Identified Using Genome-Wide Sequencing. Mol. Pain 2014, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Ponomarev, E.D.; Veremeyko, T.; Barteneva, N.; Krichevsky, A.M.; Weiner, H.L. MicroRNA-124 promotes microglia quiescence and suppresses EAE by deactivating macrophages via the C/EBP-α–PU.1 pathway. Nat. Med. 2011, 17, 64–70. [Google Scholar] [CrossRef]
- Lu, Y.; Cao, D.-L.; Jiang, B.-C.; Yang, T.; Gao, Y.-J. MicroRNA-146a-5p attenuates neuropathic pain via suppressing TRAF6 signaling in the spinal cord. Brain Behav. Immun. 2015, 49, 119–129. [Google Scholar] [CrossRef]
- Leung, A.K.; Vyas, S.; Rood, J.E.; Bhutkar, A.; Sharp, P.A.; Chang, P. Poly(ADP-Ribose) Regulates Stress Responses and MicroRNA Activity in the Cytoplasm. Mol. Cell 2011, 42, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Aldrich, B.; Frakes, E.; Kasuya, J.; Hammond, D.; Kitamoto, T. Changes in expression of sensory organ-specific microRNAs in rat dorsal root ganglia in association with mechanical hypersensitivity induced by spinal nerve ligation. Neuroscience 2009, 164, 711–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ergun, S.; Oztuzcu, S. Oncocers: ceRNA-mediated cross-talk by sponging miRNAs in oncogenic pathways. Tumour Biol. 2015, 36, 3129–3136. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Yang, Z.; Guo, J.; Zheng, Y.; Su, X.; Wang, X. Interactions Among lncRNAs/circRNAs, miRNAs, and mRNAs in Neuropathic Pain. Neurotherapeutics 2020, 17, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, X.; Chen, J.; Kang, Y.; Guo, J.; Duscher, D.; Yang, X.; Guo, G.; Ren, S.; Xiong, H.; et al. The Construction and Analysis of lncRNA-miRNA-mRNA Competing Endogenous RNA Network of Schwann Cells in Diabetic Peripheral Neuropathy. Front. Bioeng. Biotechnol. 2020, 8, 490. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.H.; Liao, H.L.; Tang, Q.F.; Liu, Y.J.; Zhang, Y.Y.; Lin, J.; Yu, H.P.; Zhou, C.; Li, C.J.; Liu, F.; et al. Interactions among Non-Coding RNAs and mRNAs in the Trigeminal Ganglion Associated with Neuropathic Pain. J. Pain Res. 2022, 15, 2967–2988. [Google Scholar] [CrossRef]
- Park, C.-K.; Xu, Z.-Z.; Berta, T.; Han, Q.; Chen, G.; Liu, X.-J.; Ji, R.-R. Extracellular microRNAs activate nociceptor neurons to elicit pain via TLR7 and TRPA1. Neuron 2014, 82, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, M. TLRs as miRNA Receptors. Cancer Res. 2012, 72, 6333–6337. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Liu, D.; Convertino, M.; Wang, Z.; Jiang, C.; Kim, Y.H.; Luo, X.; Zhang, X.; Nackley, A.; Dokholyan, N.V.; et al. miRNA-711 Binds and Activates TRPA1 Extracellularly to Evoke Acute and Chronic Pruritus. Neuron 2018, 99, 449–463. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.-B.; Zhu, Y.; Chen, B.-L.; Song, G.; Peng, M.-S.; Hu, H.-Y.; Zheng, Y.-L.; Chen, C.-C.; Yang, J.-Z.; Chen, P.-J.; et al. Network and pathway-based analysis of microRNA role in neuropathic pain in rat models. J. Cell. Mol. Med. 2019, 23, 4534–4544. [Google Scholar] [CrossRef]
- Linnstaedt, S.D.; Riker, K.D.; Rueckeis, C.A.; Kutchko, K.M.; Lackey, L.; McCarthy, K.R.; Tsai, Y.-H.; Parker, J.S.; Kurz, M.C.; Hendry, P.L.; et al. A Functional riboSNitch in the 3′ Untranslated Region of FKBP5 Alters MicroRNA-320a Binding Efficiency and Mediates Vulnerability to Chronic Post-Traumatic Pain. J. Neurosci. 2018, 38, 8407–8420. [Google Scholar] [CrossRef] [Green Version]
- Atkins, S.; Kyriakidou, E. Clinical outcomes of lingual nerve repair. Br. J. Oral Maxillofac. Surg. 2021, 59, 39–45. [Google Scholar] [CrossRef]
- Dasgupta, I.; Chatterjee, A. Recent Advances in miRNA Delivery Systems. Methods Protoc. 2021, 4, 10. [Google Scholar] [CrossRef]
- Paunovska, K.; Loughrey, D.; Dahlman, J.E. Drug delivery systems for RNA therapeutics. Nat. Rev. Genet. 2022, 23, 265–280. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| MicroRNA | Change | Injury Model | Localisation | Validated Target/s | Reference |
|---|---|---|---|---|---|
| miR-150 | Down | CCI | DRG | AKT3 | Cai, et al. [10] |
| miR-20b | Spinal cord | You, et al. [11] | |||
| miR-195 | Up | SNL | Spinal microglia | ATG14 | Shi, et al. [12] |
| miR-23 | Up | SNI | DRG | A20 | Zhang, et al. [13] |
| miR-142 | Up | CCI | Sciatic nerve | AC9 | Li, et al. [14] |
| miR-34c | Up | BCP | DRG | Cacna1e (Cav2.3) | Gandla, et al. [15] |
| miR-219 | Down | CFA | Spinal cord | CAMKII | Pan, et al. [16] |
| miR-103 | Down | SNL | Spinal cord | Cav1.2 | Favereaux, et al. [17] |
| miR-186-5p | Down | SNL | Spinal cord | CXCL13 | Jiang, et al. [18] |
| miR-23a | Down | pSNL | Spinal cord | CXCR4 | Pan, et al. [19] |
| miR-124 | Down | SNL | Spinal cord and DRG | EGR1 | Jiang, et al. [20] |
| miR-30a-5p | Down | CCI | Spinal cord | EP300 | Tan, et al. [21] |
| miR-211 | Down | CIVP | Spinal cord | ERK | Sun, et al. [22] |
| miR-124-3p | Down | CCI | Spinal cord | EZH2 | Zhang, et al. [23] |
| miR-194 | Down | CCI | Spinal cord | FOXA1 | Zhang, et al. [24] |
| miR-500 | Up | CIP | Spinal cord | GAD67 | Huang, et al. [25] |
| miR-15a/16 | Up | CCI | Spinal cord | GRK2 | Li, et al. [26] |
| miR-129 | Down | CCI | Spinal cord | HMGB1 | Ma, et al. [27] |
| miR-141 | DRG | Zhang, et al. [28] | |||
| miR-142-3p | SNL | DRG | Zhang, et al. [29] | ||
| miR-193 | DNP | Spinal cord | Wu, et al. [30] | ||
| miR-381 | CCI | DRG | Xia, et al. [31] | ||
| miR-381 | Down | CCI | Spinal cord | HMGB1, CXCR4 | Zhan, et al. [32] |
| miR-124-3p | Down | CFA | Spinal cord | IL6R | Liu, et al. [33] |
| miR-146 | Up | CCI | DRG, spinal cord | IRAK1, TRAF6 | Wang, et al. [34] |
| miR-9 | Up | DNP | Sciatic nerve | ISL1 | Sun, et al. [35] |
| miR-29a | |||||
| miR-124 | Down | CCI | Spinal cord | JAG1 | Li, et al. [36] |
| miR-141 | |||||
| miR-17-92 cluster | Up | SNL | DRG | KCNA1, KCNA4, KCNC4, KCND3, KCNQ5, DPP10, SCN1B | Sakai, et al. [37] |
| miR-137a | Up | CCI | DRG, spinal cord | KCNA2 | Zhang, et al. [38] |
| miR-216 | Down | CCI | DRG, spinal cord | KDM3A | Wang and Li [39] |
| miR-152 | Down | PNI | Spinal cord | MafB | Tozaki-Saitoh, et al. [40] |
| miR-26a | Down | CCI | Spinal cord | MAPK6 | Zhang, et al. [41] |
| miR-223 | Down | iCCI | TG | MKNK2 | Huang, et al. [42] |
| miR-101 | Up | CCI | Spinal cord and microglia | MKP4 | Qiu, et al. [43] |
| miR-183 | Down | CCI | Spinal cord | mTOR | Xie, et al. [44] |
| miR-125a-3p | Down | CFA | TG | p38 MAPK | Dong, et al. [45] |
| miR-195 | Up | iCCI | Caudal medulla and CSF | Patched1 | Wang, et al. [46] |
| miR-122 | Down | CCI | Spinal cord | PDK4 | Wan, et al. [47] |
| miR-1224 | Down | CFA | Spinal cord | pre-circ-Filip1I | Pan, et al. [48] |
| miR-16 | Up | CCI | Spinal cord | RAB23 | Chen, et al. [49] |
| miR-202 | Down | bCCI | Spinal cord | RAP1A | Fang, et al. [50] |
| miR-590 | DNP | DRG | Wu, et al. [51] | ||
| miR-144 | Down | CCI | DRG | RASA1 | Zhang, et al. [52] |
| miR-140 | Down | CCI | DRG | S1PR1 | Li, et al. [53] |
| miR-96 | Down | CCI | DRG | SCN3A | Chen, et al. [54] |
| miR-384 | Ye, et al. [55] | ||||
| miR-182 | Down | CCI | DRG | SCN9A | Jia, et al. [56] |
| SNI | Cai, et al. [57] | ||||
| miR-30b | Shao, et al. [58] | ||||
| miR-34 | Up | CFA | Spinal cord | SIRT1 | Chen, et al. [59] |
| miR-448 | CCI | Chu, et al. [60] | |||
| miR-190a | Down | DNP | Spinal cord | SLC17A6, (VGLUT2) | Yang, et al. [61] |
| miR-135a | Up | CCI | Spinal cord | SLC24A2 | Zhou, et al. [62] |
| miR-155 | Up | CCI | Spinal cord | SOCS1 | Tan, et al. [63] |
| miR-221 | Spinal cord and microglia | Xia, et al. [64] | |||
| miR-218 | Up | CCI | Spinal cord and microglia | SOCS3 | Li and Zhao [65] |
| miR-93 | Down | CCI | Spinal cord | STAT3 | Yan, et al. [66] |
| miR-98 | Zhong, et al. [67] | ||||
| miR-544 | Jin, et al. [68] | ||||
| miR-124a | Down | BCP | Spinal cord | Synpo | Elramah, et al. [69] |
| miR-28 | Down | CCI | Spinal cord | TF Zeb1 | Bao, et al. [70] |
| miR-30c | Up | SNI | Spinal cord | TGFβ | Tramullas, et al. [71] |
| miR-451 | Down | CFA | Spinal cord and microglia | TLR4 | Sun and Zhang [72] |
| miR-154 | Down | CCI | Spinal cord | TLR5 | Wei, et al. [73] |
| miR-21 | Up | SNL | DRG | TLR8 | Zhang, et al. [74] |
| miR-183 | Down | CCI | DRG | TREK1 | Shi, et al. [75] |
| miR-183 | Down | CCI | Spinal cord | TXNIP | Miao, et al. [76] |
| miR-134 | Down | CCI | Spinal cord | Twist1 | Ji, et al. [77] |
| miR-34a | Down | CCI | DRG | VAMP2 | Brandenburger, et al. [78] |
| miR-128 | Down | CCI | Spinal cord and microglia | ZEB1 | Zhang, et al. [79] |
| miR-150 | Down | CCI | Spinal cord and microglia | Zeb1 | Yan, et al. [80] |
| miR-206 | Down | CCI | Spinal microglia | ZEB2 | Chen, et al. [81] |
| Clinical Condition | Control | Sample Types | Significance | Reference |
|---|---|---|---|---|
| CiPN in patients with multiple myeloma | Myeloma patients without CiPN | Plasma | miR-22, -23a and -24a are clinically relevant biomarkers for CiPN. | Łuczkowska, et al. [82] |
| CRPS | Healthy | Whole blood | Exosomal miRNA signature is altered in CPRS patients. | McDonald, et al. [83] Orlova, et al. [84] |
| Diabetic neuropathy | Diabetic patients without neuropathy | WBCs | miR-128 is upregulated in diabetic neuropathy; miR-155 and -499 are downregulated. | Ciccacci, et al. [85] |
| Diabetic neuropathy | Healthy | Plasma and skin biopsy | miR-199-3p is increased in patients with DN and targets SERPINE2. | Li, et al. [86] |
| HIV-associated sDSP | Non-DSP and HIV | Plasma | Increased miR-455 expression is associated with reduced neurite growth. | Asahchop, et al. [87] |
| Lingual nerve injury | Lingual nerve injury without pain | Neuroma | miR-29a and miR-500a are inversely correlated with clinical VAS scores. | Tavares-Ferreira, et al. [88] |
| Musculoskeletal pain | Healthy | Plasma | miR-320 and miR-98 successfully distinguish the origin of chronic pain in 70% of the patients. | Dayer, et al. [89] |
| Neuropathic pain of various origins | Healthy | Primary human CD4+ T cells | Increased miR-124a and miR-155 levels promote Tregs differentiation. | Heyn, et al. [90] |
| Painful peripheral neuropathy | Painless peripheral neuropathy and healthy | WBCs and sural nerve biopsy | miR-132-3p is overexpressed in WBC and sural nerve of patients affected by peripheral neuropathies. | Leinders, et al. [91] |
| Painful peripheral neuropathy | Painless peripheral neuropathy and healthy | Plasma and sural nerve biopsy | miR-101 and -132 are altered in plasma and sural nerve of DN patients. miR-101 targets KPNB1. | Liu, et al. [92] |
| Painful peripheral neuropathy | Painless peripheral neuropathy and healthy | WBCs, sural nerve and skin biopsy | miR-21, -146 and -155 are differentially expressed in WBC, skin and sural nerve of patients affected by peripheral neuropathies. | Leinders, et al. [93] |
| Trigeminal neuralgia | Healthy | Serum | miR-132, -146, -155 and -384 are upregulated in patients with trigeminal neuralgia. | Li, et al. [94] |
| Neuropathic low back pain | Patients without neuropathic pain | Sinuvertebral nerve biopsy | TRPV1 upregulation is inversely correlated with miR-375 and -455 expression. | Li, et al. [95] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morchio, M.; Sher, E.; Collier, D.A.; Lambert, D.W.; Boissonade, F.M. The Role of miRNAs in Neuropathic Pain. Biomedicines 2023, 11, 775. https://doi.org/10.3390/biomedicines11030775
Morchio M, Sher E, Collier DA, Lambert DW, Boissonade FM. The Role of miRNAs in Neuropathic Pain. Biomedicines. 2023; 11(3):775. https://doi.org/10.3390/biomedicines11030775
Chicago/Turabian StyleMorchio, Martina, Emanuele Sher, David A. Collier, Daniel W. Lambert, and Fiona M. Boissonade. 2023. "The Role of miRNAs in Neuropathic Pain" Biomedicines 11, no. 3: 775. https://doi.org/10.3390/biomedicines11030775