Genetic and Epigenetic Stability in Rye Seeds under Different Storage Conditions: Ageing and Oxygen Effect


Abstract
:1. Introduction
2. Results
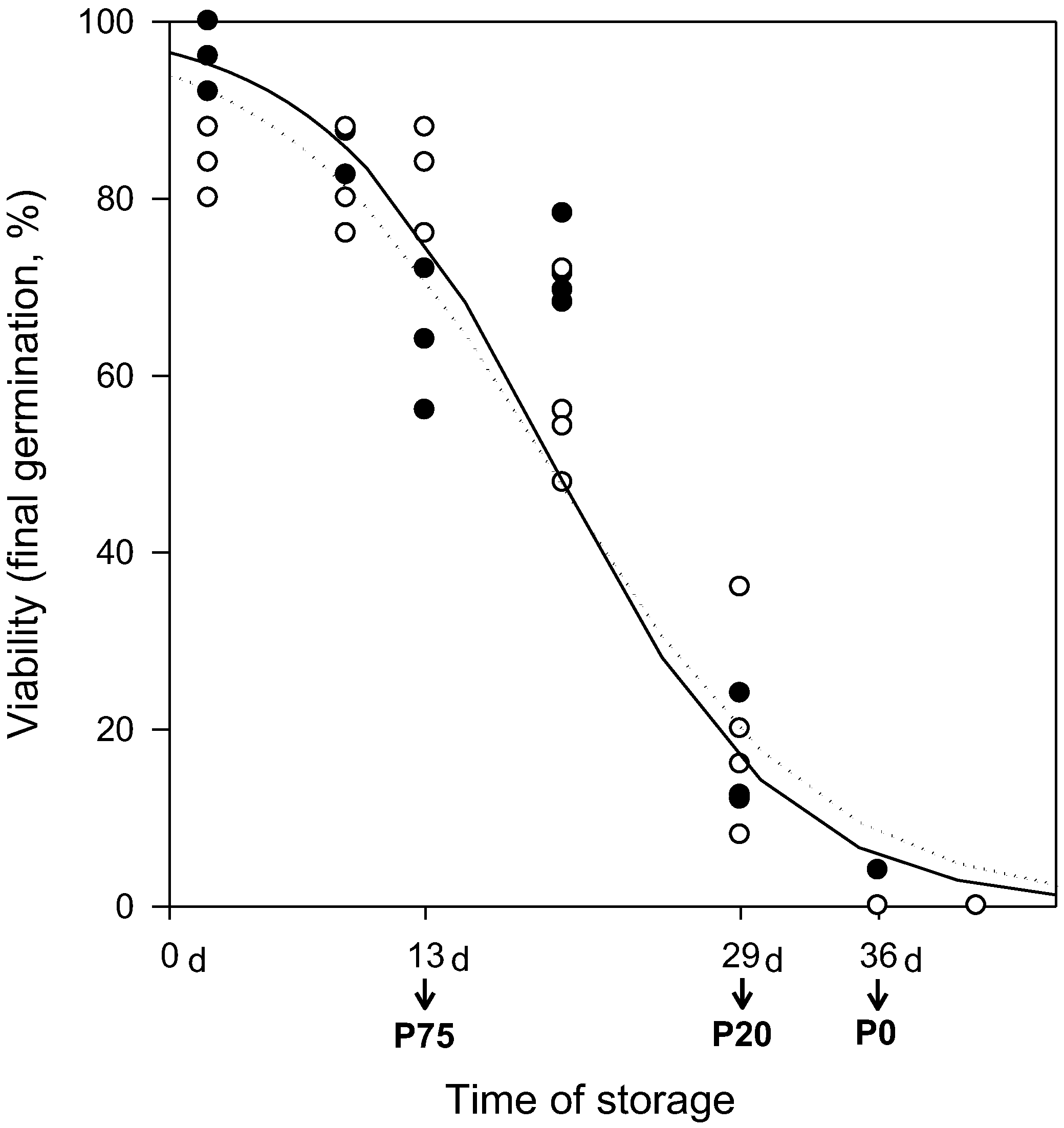
2.1. Seed Viability Loss during Storage
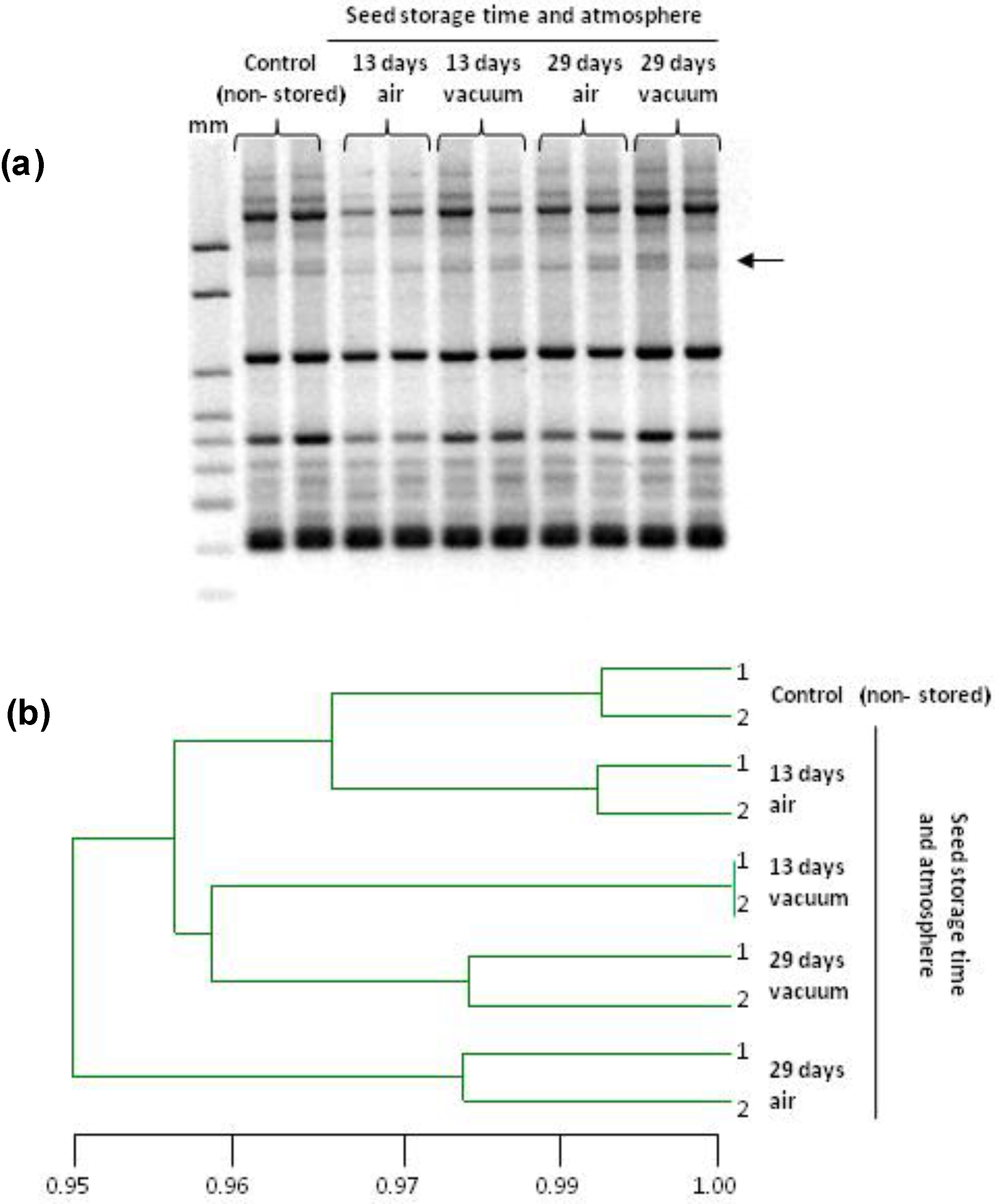
2.2. Genetic Stability of Stored Seeds and of Seedlings
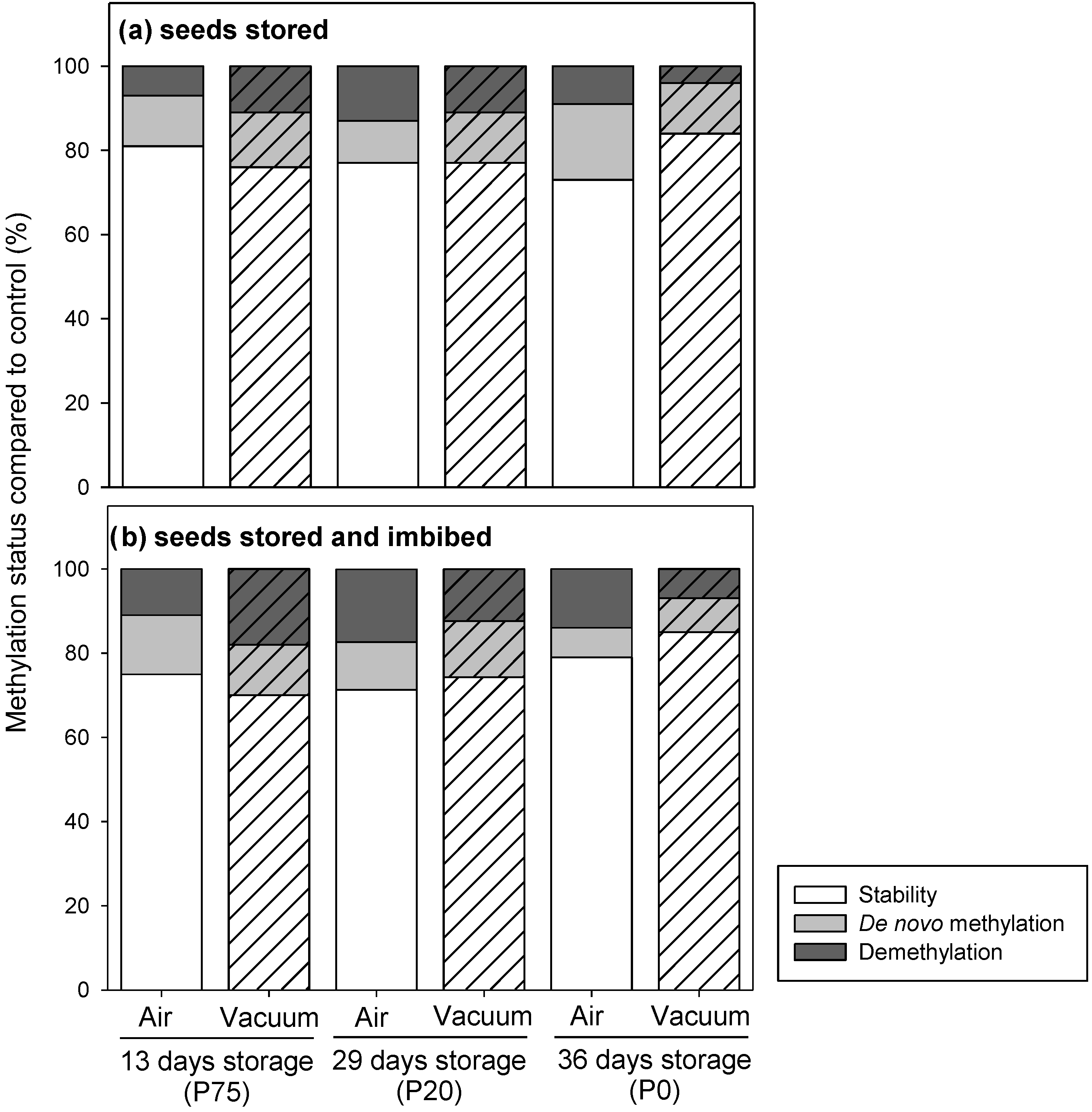
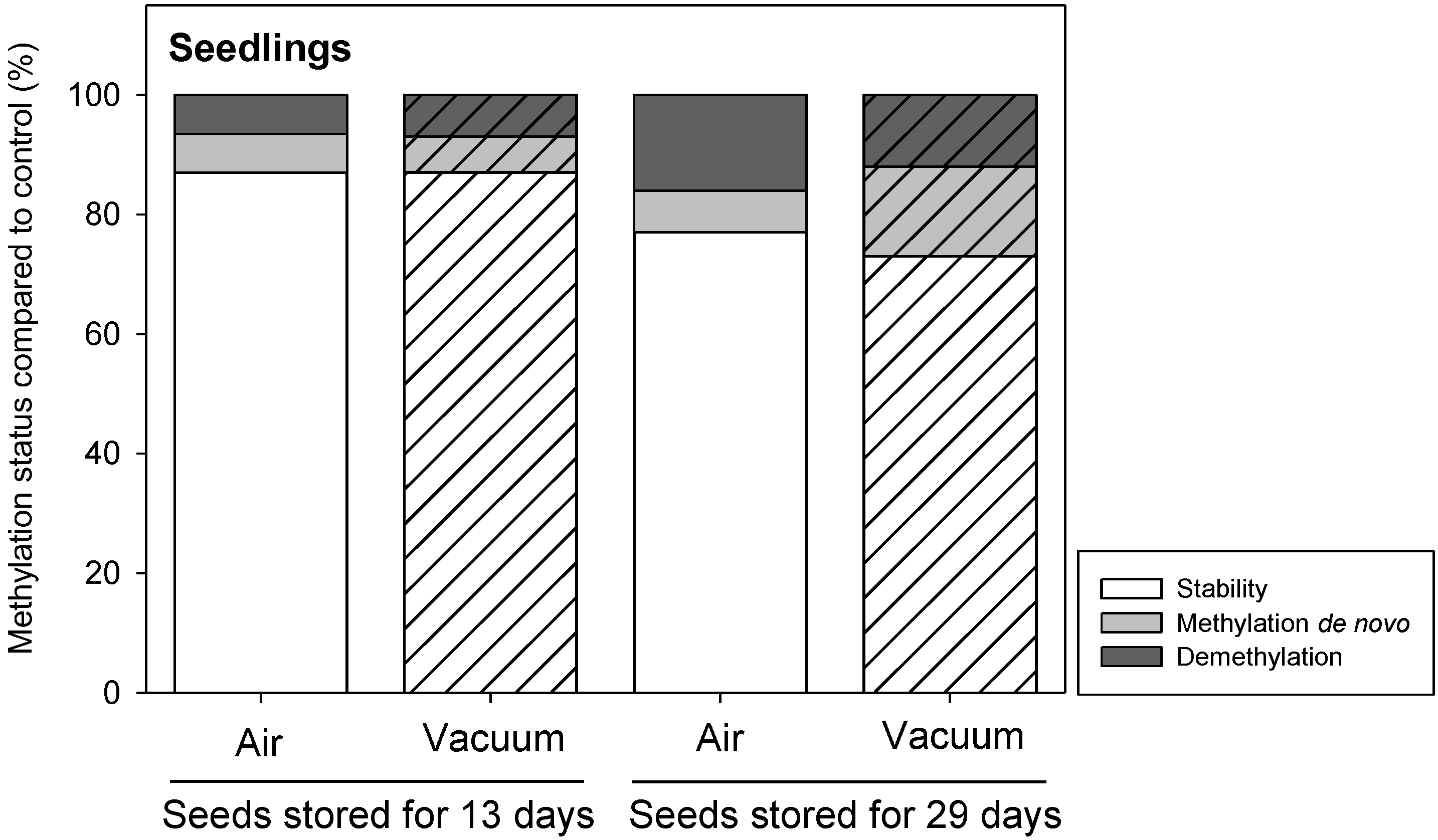
2.3. Epigenetic Stability of Stored Seeds and of Seedlings
3. Discussion
4. Materials and Methods
4.1. Seed Storage Experiment
4.2. Seed Viability and Water Content Determination
4.3. DNA Extraction and Analysis
4.4. Genetic Stability
4.5. Epigenetic Stability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walters, C.; Wheeler, L.M.; Grotenhuis, J.M. Longevity of seeds stored in a genebank: Species characteristics. Seed Sci. Res. 2005, 15, 1–20. [Google Scholar] [CrossRef]
- Probert, R.J.; Daws, M.I.; Hay, F.R. Ecological correlates of ex situ seed longevity: A comparative study on 195 species. Ann. Bot. 2009, 104, 57–69. [Google Scholar] [CrossRef] [Green Version]
- McDonald, M.B. Seed deterioration: Physiology, repair and assessment. Seed Sci. Technol. 1999, 27, 177–237. [Google Scholar]
- Barzali, M.; Lohwasser, U.; Niedzielski, M.; Borner, A. Effects of different temperatures and atmospheres on seed and seedling traits in a long-term storage experiment on rye (Secale cereale L.). Seed Sci. Technol. 2005, 33, 713–721. [Google Scholar] [CrossRef]
- Gonzalez-Benito, M.E.; Perez-Garcia, F.; Tejeda, G.; Gomez-Campo, C. Effect of the gaseous environment and water content on seed viability of four Brassicaceae species after 36 years storage. Seed Sci. Technol. 2011, 39, 443–451. [Google Scholar] [CrossRef] [Green Version]
- Merritt, D.J.; Senaratna, T.; Touchell, D.H.; Dixon, K.W.; Sivasithamparam, K. Seed ageing of four Western Australian species in relation to storage environment and seed antioxidant activity. Seed Sci. Res. 2003, 13, 155–165. [Google Scholar] [CrossRef]
- Mira, S.; Estrelles, E.; Elena Gonzalez-Benito, M.; Corbineau, F. Biochemical changes induced in seeds of Brassicaceae wild species during ageing. Acta Physiol. Plant. 2011, 33, 1803–1809. [Google Scholar] [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Kurek, K.; Plitta-Michalak, B.; Ratajczak, E. Reactive oxygen species as potential drivers of the seed aging process. Plants Basel 2019, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Kimura, S.; Sakaguchi, K. DNA repair in plants. Chem. Rev. 2006, 106, 753–766. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Bray, C.M.; West, C.E. The importance of safeguarding genome integrity in germination and seed longevity. J. Exp. Bot. 2015, 66, 3549–3558. [Google Scholar] [CrossRef] [Green Version]
- Navashin, M.; Shkvarnikov, P. Process of mutation in resting seeds accelerated by increased temperature. Nature 1933, 132, 482–483. [Google Scholar] [CrossRef]
- Tuteja, N.; Singh, M.B.; Misra, M.K.; Bhalla, P.L.; Tuteja, R. Molecular mechanisms of DNA damage and repair: Progress in plants. Crit. Rev. Biochem. Mol. Biol. 2001, 36, 337–397. [Google Scholar] [CrossRef]
- Osborne, D.J. Hazards of a germinating seed: Available water and the maintenance of genomic integrity. Isr. J. Plant Sci. 2000, 48, 173–179. [Google Scholar] [CrossRef]
- Kranner, I.; Chen, H.; Pritchard, H.W.; Pearce, S.R.; Birtic, S. Inter-nucleosomal DNA fragmentation and loss of RNA integrity during seed ageing. Plant Growth Regul. 2011, 63, 63–72. [Google Scholar] [CrossRef]
- Dona, M.; Balestrazzi, A.; Mondoni, A.; Rossi, G.; Ventura, L.; Buttafava, A.; Macovei, A.; Sabatini, M.E.; Valassi, A.; Carbonera, D. DNA profiling, telomere analysis and antioxidant properties as tools for monitoring ex situ seed longevity. Ann. Bot. 2013, 111, 987–998. [Google Scholar] [CrossRef] [Green Version]
- Gehring, M.; Henikoff, S. DNA methylation dynamics in plant genomes. Biochim. Biophys. Acta Gene Struct. Expr. 2007, 1769, 276–286. [Google Scholar] [CrossRef]
- Feng, S.; Jacobsen, S.E.; Reik, W. Epigenetic reprogramming in plant and animal development. Science 2010, 330, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Lister, R.; Ecker, J.R. Finding the fifth base: Genome-wide sequencing of cytosine methylation. Genome Res. 2009, 19, 959–966. [Google Scholar] [CrossRef] [Green Version]
- Vanyushin, B.F.; Ashapkin, V.V. DNA methylation in higher plants: Past, present and future. Biochim. Biophys. Acta Gene Regul. Mech. 2011, 1809, 360–368. [Google Scholar] [CrossRef]
- Jablonka, E.; Lamb, M.J. Epigenetic inheritance in evolution. J. Evol. Biol. 1998, 11, 159–183. [Google Scholar] [CrossRef]
- Lukens, L.N.; Zhan, S. The plant genome’s methylation status and response to stress: Implications for plant improvement. Curr. Opin. Plant Biol. 2007, 10, 317–322. [Google Scholar] [CrossRef]
- Yuan, J.-L.; Sun, H.-M.; Guo, G.-P.; Yue, J.-J.; Gu, X.-P. Correlation between DNA methylation and chronological age of moso bamboo (Phyllostachys heterocycla var. pubescens). Bot. Stud. 2014, 55, 4. [Google Scholar] [CrossRef] [Green Version]
- Ogneva, Z.V.; Dubrovina, A.S.; Kiselev, K.V. Age-associated alterations in DNA methylation and expression of methyltransferase and demethylase genes in Arabidopsis thaliana. Biol. Plant. 2016, 60, 628–634. [Google Scholar] [CrossRef]
- Michalak, M.; Plitta-Michalak, B.P.; Naskret-Barciszewska, M.; Barciszewski, J.; Bujarska-Borkowska, B.; Chmielarz, P. Global 5-methylcytosine alterations in DNA during ageing of Quercus robur seeds. Ann. Bot. 2015, 116, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Plitta-Michalak, B.P.; Naskret-Barciszewska, M.Z.; Kotlarski, S.; Tomaszewski, D.; Tylkowski, T.; Barciszewski, J.; Chmielarz, P.; Michalak, M. Changes in genomic 5-methylcytosine level mirror the response of orthodox (Acer platanoides L.) and recalcitrant (Acer pseudoplatanus L.) seeds to severe desiccation. Tree Physiol. 2018, 38, 617–629. [Google Scholar] [CrossRef] [Green Version]
- Michalak, M.; Barciszewska, M.Z.; Barciszewski, J.; Plitta, B.P.; Chmielarz, P. Global changes in DNA Methylation in seeds and seedlings of Pyrus communis after seed desiccation and storage. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization. Hundred and Sixtieth Session of FAO Council. Proposal for an International Year of Rye. Rome, 3–7 December. 2018. Available online: http://www.fao.org/about/meetings/council/cl160/documents/en (accessed on 4 December 2019).
- Miedaner, T.; Korzun, V. Marker-assisted selection for disease resistance in wheat and barley breeding. Phytopathology 2012, 102, 560–566. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Herrera, L.A.; Garkava-Gustavsson, L.; Ahman, I. A systematic review of rye (Secale cereale L.) as a source of resistance to pathogens and pests in wheat (Triticum aestivum L.). Hereditas 2017, 154, 1–9. [Google Scholar] [CrossRef]
- Niedzielski, M.; Walters, C.; Luczak, W.; Hill, L.M.; Wheeler, L.J.; Puchalski, J. Assessment of variation in seed longevity within rye, wheat and the intergeneric hybrid triticale. Seed Sci. Res. 2009, 19, 213–224. [Google Scholar] [CrossRef]
- Mira, S.; Pirredda, M.; Martín-Sánchez, M.; Marchessi, J.E.; Martín, C. DNA methylation and integrity in aged seeds and regenerated plants. Seed Sci. Res. 2020, in press. [Google Scholar]
- Marcos-Filho, J.; McDonald, M.B.; Tekrony, D.M.; Zhang, J. RAPD fragment profiles from deteriorating soybean seeds. Seed Technol. 1997, 19, 33–44. [Google Scholar]
- Vijay, D.; Dadlani, M.; Kumar, P.A.; Panguluri, S.K. Molecular marker analysis of differentially aged seeds of soybean and safflower. Plant Mol. Biol. Rep. 2009, 27, 282–291. [Google Scholar] [CrossRef]
- El-Maarouf-Bouteau, H.; Mazuy, C.; Corbineau, F.; Bailly, C. DNA alteration and programmed cell death during ageing of sunflower seed. J. Exp. Bot. 2011, 62, 5003–5011. [Google Scholar] [CrossRef] [Green Version]
- Shatters, R.G.; Schweder, M.E.; West, S.H.; Abdelghany, A.; Smith, R.L. Environmentally-induced polymorphism detected by RAPD analysis of soyabean seed DNA. Seed Sci. Res. 1995, 5, 109–116. [Google Scholar] [CrossRef]
- Andreev, I.O.; Spiridonova, E.V.; Kunakh, V.A.; Solovyan, V.T. Aging and loss of germination in rye seeds is accompanied by a decreased fragmentation of nuclear DNA at loop domain boundaries. Russ. J. Plant Physiol. 2004, 51, 241–248. [Google Scholar] [CrossRef]
- Boubriak, I.; Kargiolaki, H.; Lyne, L.; Osborne, D.J. The requirement for DNA repair in desiccation tolerance of germinating embryos. Seed Sci. Res. 1997, 7, 97–105. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Masnavi, G.; Bhardwaj, R.M.; Jiang, Q.; Bray, C.M.; West, C.E. A plant DNA ligase is an important determinant of seed longevity. Plant J. 2010, 63, 848–860. [Google Scholar] [CrossRef]
- Zhang, X.; Yazaki, J.; Sundaresan, A.; Cokus, S.; Chan, S.W.L.; Chen, H.; Henderson, I.R.; Shinn, P.; Pellegrini, M.; Jacobsen, S.E.; et al. Genome-wide high-resolution mapping and functional analysis of DNA methylation in Arabidopsis. Cell 2006, 126, 1189–1201. [Google Scholar] [CrossRef] [Green Version]
- Kaeppler, S.M.; Kaeppler, H.F.; Rhee, Y. Epigenetic aspects of somaclonal variation in plants. Plant Mol. Biol. 2000, 43, 179–188. [Google Scholar] [CrossRef]
- Kim, J.-M.; Sasaki, T.; Ueda, M.; Sako, K.; Seki, M. Chromatin changes in response to drought, salinity, heat, and cold stresses in plants. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoeven, K.J.F.; Jansen, J.J.; Van Dijk, P.J.; Biere, A. Stress-induced DNA methylation changes and their heritability in asexual dandelions. New Phytol. 2010, 185, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Schwember, A.R.; Bradford, K.J. Oxygen interacts with priming, moisture content and temperature to affect the longevity of lettuce and onion seeds. Seed Sci. Res. 2011, 21, 175–185. [Google Scholar] [CrossRef]
- Groot, S.P.C.; De Groot, L.; Kodde, J.; Van Treuren, R. Prolonging the longevity of ex situ conserved seeds by storage under anoxia. Plant Genet. Resour. 2015, 13, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Roberts, E.H.; Ellis, R.H. Water and seed survival. Ann. Bot. 1989, 63, 39–52. [Google Scholar] [CrossRef]
- Walters, C.; Hill, L.M.; Wheeler, L.J. Dying while dry: Kinetics and mechanisms of deterioration in desiccated organisms. Integr. Comp. Boil. 2005, 45, 751–758. [Google Scholar] [CrossRef]
- Chwedorzewska, K.J.; Bednarek, P.T.; Lewandowska, R.; Krajewski, P.; Puchalski, J. Studies on genetic changes in rye samples (Secale cereale L.) maintained in a seed bank. Cell. Mol. Biol. Lett. 2006, 11, 338–347. [Google Scholar] [CrossRef]
- Schoen, D.J.; David, J.L.; Bataillon, T.M. Deleterious mutation accumulation and the regeneration of genetic resources. Proc. Natl. Acad. Sci. USA 1998, 95, 394–399. [Google Scholar] [CrossRef] [Green Version]
- Vertucci, C.W.; Roos, E.E. Theoretical basis of protocols for seed storage II. The influence of temperature on optimal moisture levels. Seed Sci. Res. 1993, 3, 201–213. [Google Scholar] [CrossRef]
- International Seed Testing Association. International Rules for Seed Testing; ISTA: Bassersdorf, Switzerland, 2017. [Google Scholar]
- Team R.C. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org (accessed on 27 February 2020).
- ReynaLopez, G.E.; Simpson, J.; RuizHerrera, J. Differences in DNA methylation patterns are detectable during the dimorphic transition of fungi by amplification of restriction polymorphisms. Mol. Gen. Genet. 1997, 253, 703–710. [Google Scholar] [CrossRef]
- Xiong, L.Z.; Xu, C.G.; Maroof, M.A.S.; Zhang, Q.F. Patterns of cytosine methylation in an elite rice hybrid and its parental lines, detected by a methylation-sensitive amplification polymorphism technique. Mol. Gen. Genet. 1999, 261, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Bardini, M.; Labra, M.; Winfield, M.; Sala, F. Antibiotic-induced DNA methylation changes in calluses of Arabidopsis thaliana. Plant Cell Tissue Organ. Cult. 2003, 72, 157–162. [Google Scholar] [CrossRef]
- Fulnecek, J.; Kovarik, A. How to interpret Methylation Sensitive Amplified Polymorphism (MSAP) profiles? BMC Genet. 2014, 15, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Restriction Enzyme Database. Available online: http://rebase.neb.com (accessed on 27 February 2020).
- Agresti, A. An Introduction to Categorical Data Analysis; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Ibañez, M.A.; Alvarez-Mari, A.; Rodríguez-Sanz, H.; Kremer, C.; González-Benito, M.; Martín, C. Genetic and epigenetic stability of recovered mint apices after several steps of a cryopreservation protocol by encapsulation-dehydration. A new approach for epigenetic analysis. Plant Physiol. Biochem. 2019, 143, 299–307. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Atmosphere | WC (% dw) | Seed Longevity Parameters (Days) | ||
|---|---|---|---|---|
| P75 | P50 | P20 | ||
| Air | 15.2 ± 0.1 | 14 ± 2 | 20 ± 2 | 28 ± 2 |
| Vacuum | 15.1 ± 0.1 | 12 ± 2 | 19 ± 2 | 29 ± 3 |
| Storage Atmosphere | Number of Polymorphic Markers | |||||||
|---|---|---|---|---|---|---|---|---|
| Stored Seeds | Stored-Imbibed Seeds | Seedlings | ||||||
| Storage Time (Days) | Storage Time (Days) | Seed Storage Time (Days) | ||||||
| 13 (P75) | 29 (P20) | 36 (P0) | 13 (P75) | 29 (P20) | 36 (P0) | 13 (P75) | 29 (P20) | |
| Air | 3 | 4 | 5 | 0 | 1 | 1 | 4 | 5 |
| Vacuum | 0 | 2 | 3 | 2 | 1 | 1 | 4 | 5 |
| Total | 134 | 134 | 134 | 134 | 134 | 134 | 104 | 104 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pirredda, M.; González-Benito, M.E.; Martín, C.; Mira, S. Genetic and Epigenetic Stability in Rye Seeds under Different Storage Conditions: Ageing and Oxygen Effect. Plants 2020, 9, 393. https://doi.org/10.3390/plants9030393
Pirredda M, González-Benito ME, Martín C, Mira S. Genetic and Epigenetic Stability in Rye Seeds under Different Storage Conditions: Ageing and Oxygen Effect. Plants. 2020; 9(3):393. https://doi.org/10.3390/plants9030393
Chicago/Turabian StylePirredda, Michela, M. Elena González-Benito, Carmen Martín, and Sara Mira. 2020. "Genetic and Epigenetic Stability in Rye Seeds under Different Storage Conditions: Ageing and Oxygen Effect" Plants 9, no. 3: 393. https://doi.org/10.3390/plants9030393