A Correlative Study of Sunflower Seed Vigor Components as Related to Genetic Background

 and
and
Abstract
:1. Introduction:
2. Results
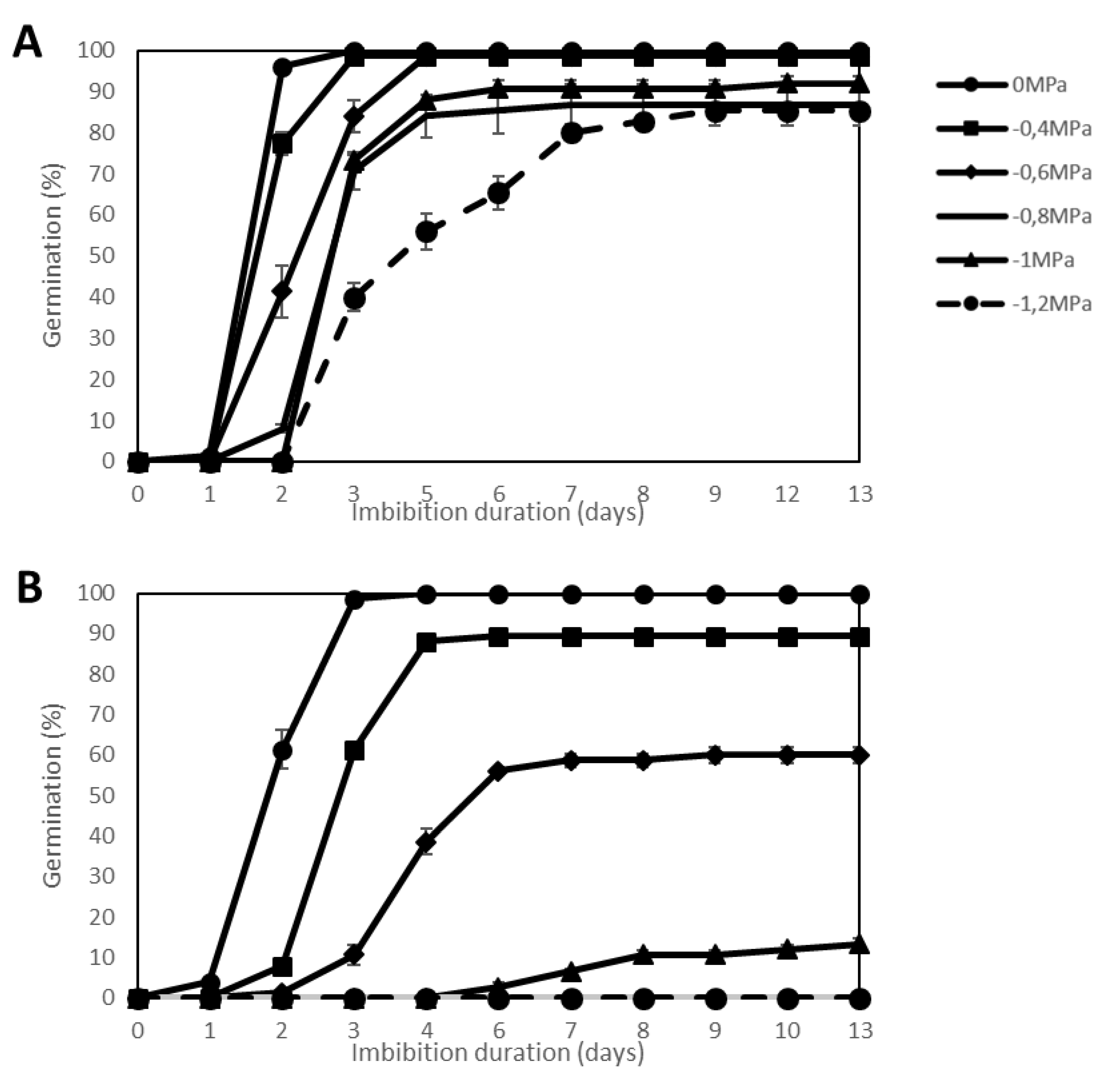
2.1. Effect of Water Stress
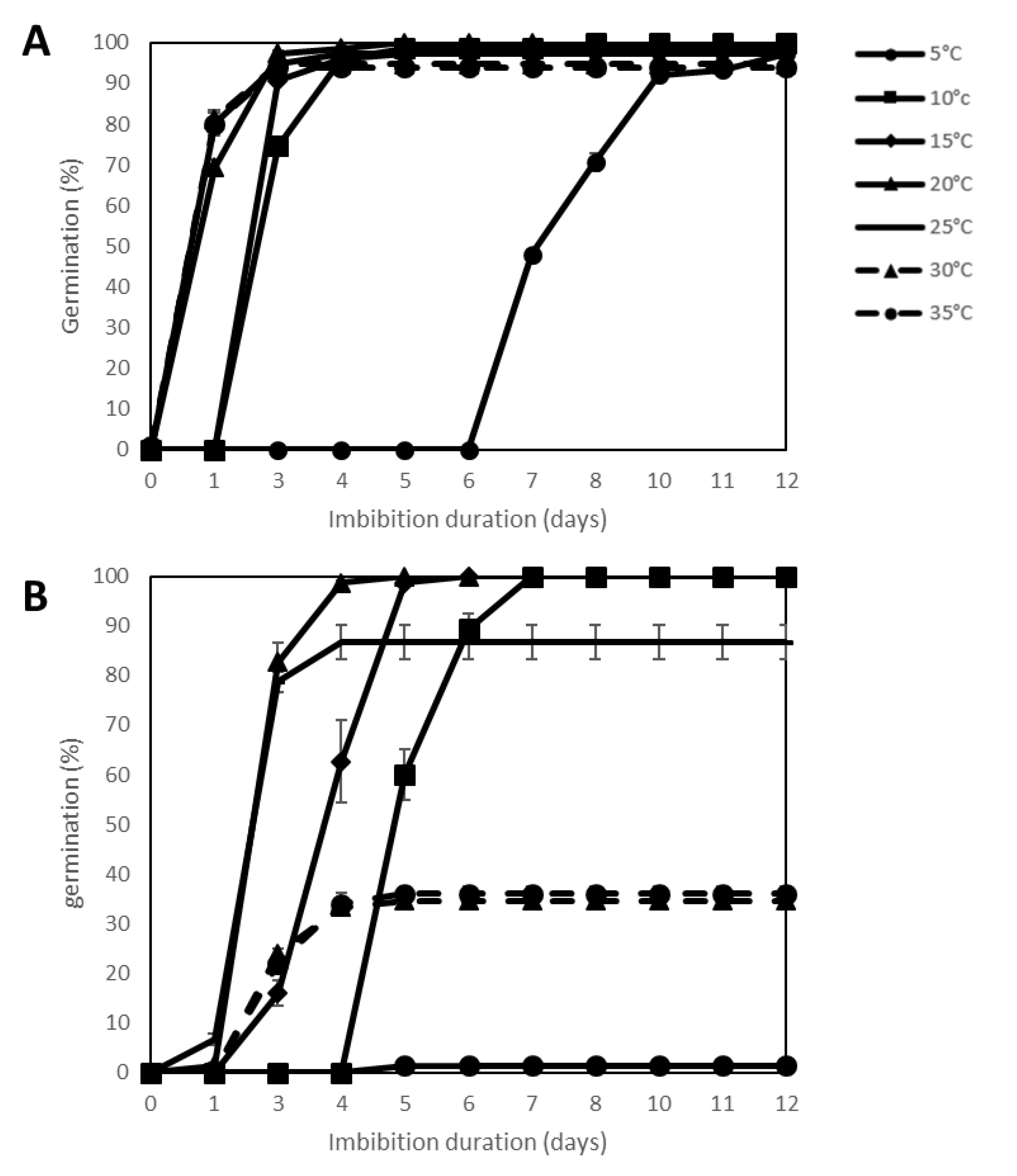
2.2. Effect of Temperature Stress
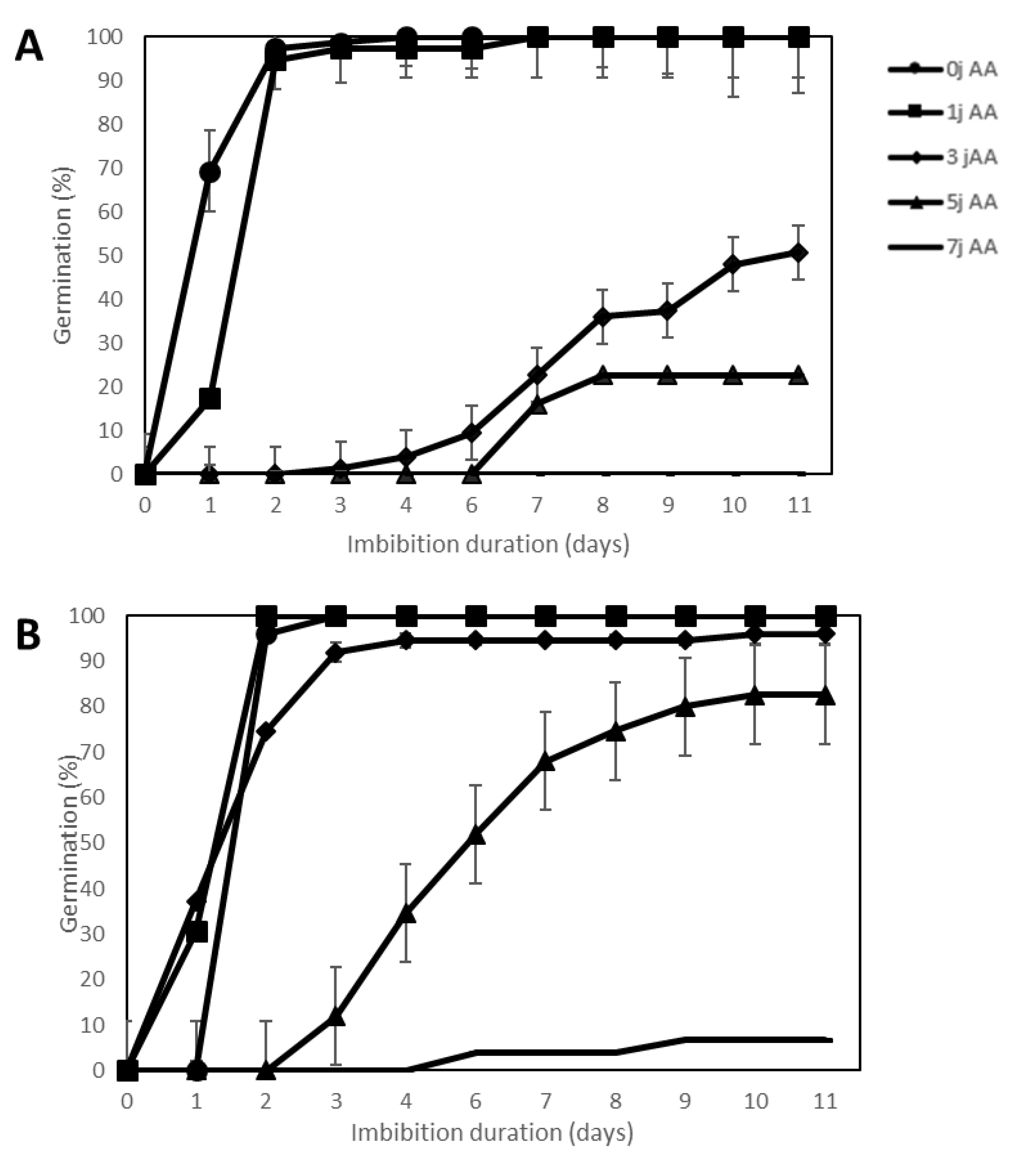
2.3. Effect of Accelerated Ageing
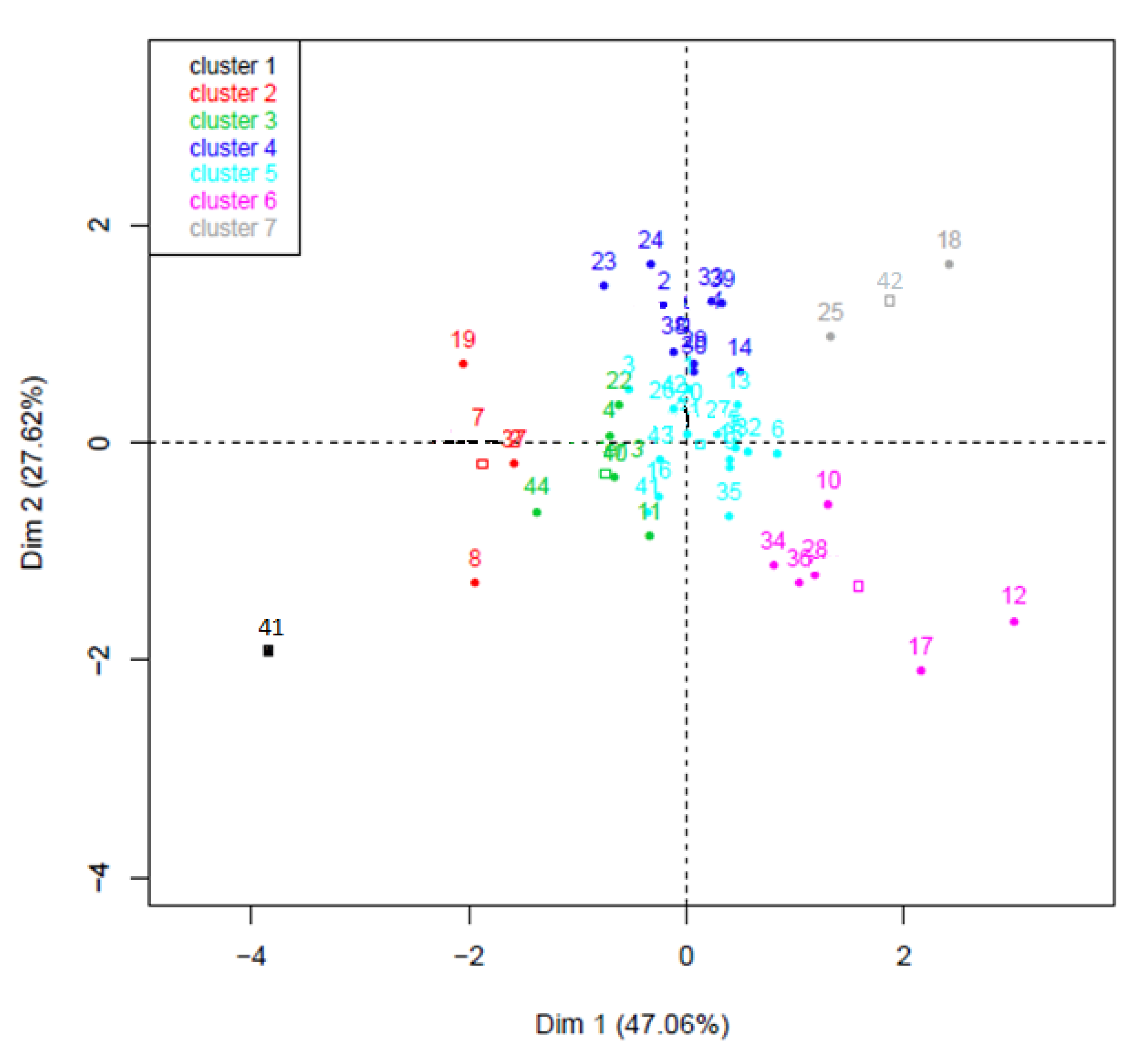
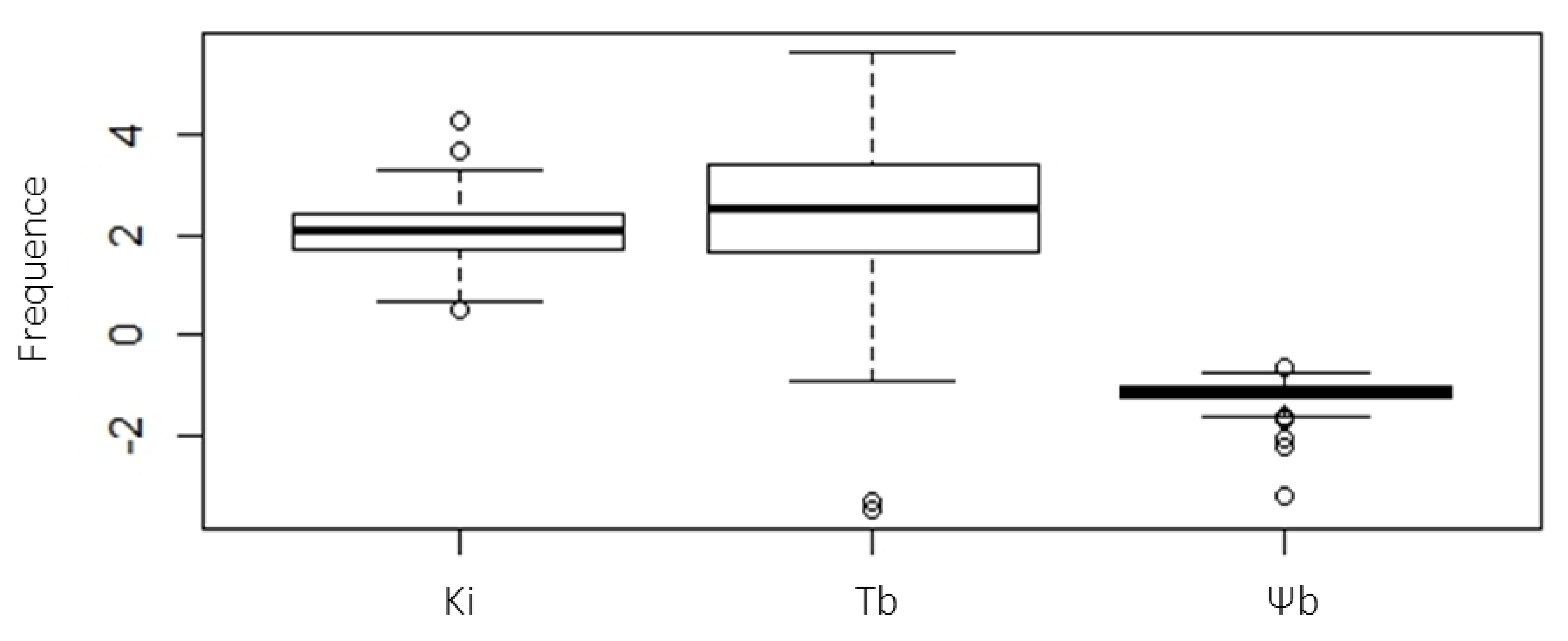
2.4. Determination of Seed Vigor Constants
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Seed Treatments and Germination
4.3. Germination Parameter Calculation
4.3.2. Base Water Potential
4.3.3. Base Temperature
4.3.4. Viability Constant Calculation (Ki)
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- David, L.D. Chapter 2: Seed Vigor. In Seed Quality. Basic Mechanisms and Agricultural Implications; Food Product Press: Basra, Iraq, 1995; pp. 45–80. [Google Scholar]
- Hampton, J.G. What is seed quality? Seed Sci. Technol. 2002, 30, 1–10. [Google Scholar]
- Ellis, R.H. Seed and seedling vigour in relation to crop growth and yield. Plant Growth Regul. 1992, 11, 249–255. [Google Scholar] [CrossRef]
- ISTA Vigour Test Committee. Understanding Seed Vigour. In Handbook of Vigour Test Methods; International Seed testing Association: Zurich, Switzerland, 1995. [Google Scholar]
- ISTA Vigour Test Committee. Chapter 15 Seed vigour testing. In International Rules for Seed Testing; International Seed testing Association: Zurich, Switzerland, 2015. [Google Scholar]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination, 2nd ed.; Plenum Press: New York, NY, USA, 1994. [Google Scholar]
- Delouche, J.C.; Baskin, C.C. Accelerated aging techniques for predicting the relative storability of seed lots. Seed Sci. Technol. 1973, 1, 427–452. [Google Scholar]
- Bradford, K.J. Water relations in seed germination. In Seed Development and Germination; Jaime Kigel and Gad Galili Marcel Dekker Inc: New York, NY, USA, 1995; pp. 351–396. [Google Scholar]
- Alvarado, V.; Bradford, K.J. A hydrothermal time model explains the cardinal temperatures for seed germination. Plant Cell Env. 2002, 25, 1061–1069. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonogaki, H. Seed dormancy and germination emerging mechanisms and new hypotheses. Front. Plant Sci. 2014, 5, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonogaki, H. Seed Biology Updates—Highlights and New Discoveries in Seed Dormancy and Germination Research. Front. Plant Sci. 2017, 8, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Ayele, B.T. Functional genomics of seed dormancy in wheat: Advances and prospects. Front. Plant Sci. 2014, 5, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazin, J.; Langlade, N.; Vincourt, P.; Arribat, S.; Balzergue, S.; El-Maarouf-Bouteau, H.; Bailly, C. Targeted mRNA Oxidation Regulates Sunflower Seed Dormancy Alleviation during Dry After-Ripening. Plant Cell 2011, 23, 2196–2208. [Google Scholar] [CrossRef] [Green Version]
- El-Maarouf-Bouteau, H.; Meimoun, P.; Job, C.; Job, D.; Bailly, C. Role of protein and mRNA oxidation in seed dormancy and germination. Front. Plant Sci. 2013, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Xia, Q.; Ponnaiah, M.; Cueff, G.; Rajjou, L.; Prodhomme, D.; Gibon, Y.; Bailly, C.; Corbineau, F.; Meimoun, P.; El-Maarouf-Bouteau, H. Integrating proteomics and enzymatic profiling to decipher seed metabolism affected by temperature in seed dormancy and germination. Plant Sci. 2018, 269, 118–125. [Google Scholar] [CrossRef]
- Xia, Q.; Saux, M.; Ponnaiah, M.; Gilard, F.; Perreau, F.; Huguet, S.; Balzergue, S.; Langlade, N.; Bailly, C.; Meimoun, P. One Way to Achieve Germination: Common Molecular Mechanism Induced by Ethylene and After-Ripening in Sunflower Seeds. IJMS 2018, 19, 2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilhorst, H.W.M.; Toorop, P.E. Review on dormancy, germinability and germination in crop and weed seeds. Adv. Agron. 1997, 61, 112–165. [Google Scholar]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed Germination and Vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [Green Version]
- Saux, M.; Ponnaiah, M.; Langlade, N.; Zanchetta, C.; Balliau, T.; El-Maarouf-Bouteau, H.; Bailly, C. A multiscale approach reveals regulatory players of water stress responses in seeds during germination. Plant Cell Environ. 2020, 13731. [Google Scholar] [CrossRef] [PubMed]
- Bettey, M.; Finch-Savage, W.E.; King, G.J.; Lynn, J.R. Quantitative genetic analysis of seed vigour and pre-emergence seedling growth traits in Brassica oleracea: RESEARCH QTL analysis of seed vigour in Brassica. New Phytol. 2000, 148, 277–286. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Clay, H.A.; Lynn, J.R.; Morris, K. Towards a genetic understanding of seed vigour in small-seeded crops using natural variation in Brassica oleracea. Plant Sci. 2010, 179, 582–589. [Google Scholar] [CrossRef]
- Morris, K.; Barker, G.C.; Walley, P.G.; Lynn, J.R.; Finch-Savage, W.E. Trait to gene analysis reveals that allelic variation in three genes determines seed vigour. New Phytol. 2016, 212, 964–976. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.H.; Roberts, E.H. Improved equations for the prediction of seed longevity. Ann. Bot. 1980, 45, 13–30. [Google Scholar] [CrossRef]
- Balesevic-Tubic, S.; Malencic, D.; Tatic, M.; Miladinovic, J. Influence of aging process on biochemical changes in sunflower seed. Helia 2005, 28, 107–114. [Google Scholar] [CrossRef]
- Castillo-Lorenzo, E.; Finch-Savage, W.E.; Seal, C.E.; Pritchard, H.W. Adaptive significance of functional germination traits in crop wild relatives of Brassica. Agric. For. Meteorol. 2018, 264, 343–350. [Google Scholar] [CrossRef]
- Belo, R.G.; Tognetti, J.; Benech-Arnold, R.; Izquierdo, N.G. Germination responses to temperature and water potential as affected by seed oil composition in sunflower. Ind. Crop. Prod. 2014, 62, 537–544. [Google Scholar] [CrossRef]
- Ambika, S.; Manonmani, V.; Somasundar, G. Review on Effect of Seed Size on Seedling Vigour and Seed Yield. Res. J. Seed Sci. 2014, 7, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Moshatati, A.; Gharineh, M.H. Effect of grain weight on germination and seed vigor of wheat. Int. J. Agric. Crop Sci. 2012, 4, 458–460. [Google Scholar]
- Zareian, A.; Hamidi, A.; Sadeghi, H.; Jazaeri, M.R. Effect of seed size on some germination characteristics, seedling emergence percentage and yield of three wheat (Triticum aestivum L.) cultivars in laboratory and field. Middle-East J. Sci. Res. 2013, 13, 1126–1131. [Google Scholar]
- Ries, S.K.; Everson, E.H. Protein Content and Seed Size Relationships with Seedling Vigor of Wheat Cultivars 1. Agron. J. 1973, 65, 884–886. [Google Scholar] [CrossRef]
- Naylor, R.E.L. The effect of parent plant nutrition on seed size, viability and vigour and on germination of wheat and triticale at different temperatures. Ann. Appl. Biol. 1993, 123, 379–390. [Google Scholar] [CrossRef]
- Farahani, H.A.; Moaveni, P.; Maroufi, K. Effect of seed size on seedling vigour in sunflower (Helianthus annuus L.). Adv. Environ. Biol. 2011, 5, 1701–1705. [Google Scholar]
- Saranga, Y.; Levi, A.; Horcicka, P.; Wolf, S. Large sunflower seeds are characterized by low embryo vigor. J. Am. Soc. Hortic. Sci. 1998, 123, 470–474. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The Osmotic Potential of Polyethylene Glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef]
- Coolbear, P.; Francis, A.; Grierson, D. The effect of low temperature pre-sowing treatment on the germination performance and membrane integrity of artificially aged tomato seeds. J. Exp. Bot. 1984, 35, 1609–1617. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Ahmad, N.; Hafeez, K. Thermal hardening: A new seed vigor enhancement tool in rice. J. Integr. Plant Biol. 2005, 47, 187–193. [Google Scholar] [CrossRef]
- Bradford, K.J. Applications of hydrothermal time to quantifying and modeling seed germination and dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Genotypes | Ψb (50%) (MPa) | Tb (°C) | Ki |
|---|---|---|---|---|
| 1 | SOLQUA-003 | −1.066 | 3.07 | 2.0823 |
| 2 | SOLQUA-006 | −1.2284 | 2.15 | 2.3716 |
| 3 | SOLQUA-007 | −1.1135 | 1.58 | 2.3209 |
| 4 | SOLQUA-011 | −0.9311 | 4.53 | 2.0814 |
| 5 | SOLQUA-015-3 | −0.9404 | 4.17 | 2.4945 |
| 6 | SOLQUA-020 | −1.0896 | 3.62 | 1.6434 |
| 7 | SOLQUA-023 | −0.8251 | 3.06 | 0.7898 |
| 8 | SOLQUA-024 | −0.9446 | 1.72 | 1.9408 |
| 9 | SOLQUA-028 | −0.9856 | 1.27 | 2.3179 |
| 10 | SOLQUA-030 | −1.0049 | −0.9 | 2.034 |
| 11 | SOLQUA-031 | −2.217 | 4.39 | 2.169 |
| 12 | SOLQUA-037 | −1.0511 | 2.4 | 1.8161 |
| 13 | SOLQUA-037-2 | −1.1699 | −0.36 | 2.0397 |
| 14 | SOLQUA-040 | −2.0158 | 2.27 | 1.0646 |
| 15 | SOLQUA-047 | −1.1548 | 5.42 | 2.0977 |
| 16 | SOLQUA-048 | −0.9483 | 1.5 | 1.8038 |
| 17 | SOLQUA-050 | −0.9966 | 2.93 | 1.6187 |
| 18 | SOLQUA-055 | −1.1191 | 0.05 | 2.6992 |
| 19 | SOLQUA-056-3 | −1.1189 | 0.79 | 1.8438 |
| 20 | SOLQUA-056-4 | −1.1355 | −0.86 | 2.0945 |
| 21 | SOLQUA-057 | −1.1903 | 1.23 | 1.2352 |
| 22 | SOLQUA-068-5 | −1.6805 | 3.67 | 1.4013 |
| 23 | SOLQUA-073 | −0.966 | 3.76 | 1.9841 |
| 24 | SOLQUA-075 | −0.9354 | −3.44 | 3.2565 |
| 25 | SOLQUA-088 | −1.1602 | 4.08 | 2.9622 |
| 26 | SOLQUA-096 | −1.2488 | 5.33 | 2.7511 |
| 27 | SOLQUA-107 | −1.0686 | 2.38 | 2.3946 |
| 28 | SOLQUA-109 | −1.1588 | 2.57 | 3.6534 |
| 29 | SOLQUA-110 | −1.1102 | 2.42 | 1.1141 |
| 30 | SOLQUA-110-2 | −1.5864 | 1.69 | nd |
| 31 | SOLQUA-113 | −0.9967 | 2.86 | 2.4624 |
| 32 | SOLQUA-114 | −0.9953 | 3.39 | 2.1343 |
| 33 | SOLQUA-123 | −1.2608 | 3.33 | 2.5701 |
| 34 | SOLQUA-127 | −1.2095 | 2.64 | 0.4968 |
| 35 | SOLQUA-132 | −0.6323 | 3.4 | 0.6833 |
| 36 | SOLQUA-133 | −1.3559 | 2.95 | 2.373 |
| 37 | SOLQUA-138 | −1.2303 | 1.67 | 2.3454 |
| 38 | SOLQUA-143 | −1.3089 | 1.78 | 1.7256 |
| 39 | SOLQUA-147 | −1.244 | 2.29 | 1.8077 |
| 40 | SOLQUA-148 | −1.1306 | −3.3 | 2.6047 |
| 41 | SOLQUA-160 | −3.1991 | 3.35 | 1.1666 |
| 42 | SOLQUA-165 | −0.7507 | 2.5 | 4.2442 |
| 43 | SOLQUA-168 | −1.6257 | 5.62 | 1.4051 |
| 44 | SOLQUA-169 | −1.094 | 2.64 | 1.9944 |
| Variables | Ψb (50%) (MPa) | Tb (°C) | Ki |
|---|---|---|---|
| Ψb (50%) (MPa) | 1 (p = 0) | 0.035 (p = 0.226) | 0.058 (p = 0.116) |
| Tb (°C) | 0.035 (p = 0.226) | 1 (p = 0) | 0.036 (p = 0.218) |
| Ki | 0.058 (p = 0.116) | 0.036 (p = 0.218) | 1 (p = 0) |
| Cluster Number | Description of Seed Lot Response | Number of Genotypes | Number of Genotypes Whose 1000 SW > 50 | Number of Genotypes Whose 1000 SW < 50 |
|---|---|---|---|---|
| 1 | Tolerant to high temperatures Tolerant to water stress Low initial quality | 1 | 0 | 1 |
| 2 | Tolerant to low temperatures Tolerant to water stress Sensitive to high temperatures Low initial quality | 4 | 2 | 2 |
| 3 | Tolerant to water stress Low initial quality | 6 | 2 | 4 |
| 4 | Sensitive to low or high temperatures Tolerant to water stress Low initial quality | 8 | 5 | 3 |
| 5 | None | 17 | 11 | 6 |
| 6 | Tolerant to low temperature Sensitive to water stress High initial quality | 6 | 5 | 1 |
| 7 | Tolerant to low temperature Sensitive to water stress Sensitive to high temperatures High initial quality | 2 | 0 | 2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saux, M.; Bleys, B.; André, T.; Bailly, C.; El-Maarouf-Bouteau, H. A Correlative Study of Sunflower Seed Vigor Components as Related to Genetic Background. Plants 2020, 9, 386. https://doi.org/10.3390/plants9030386
Saux M, Bleys B, André T, Bailly C, El-Maarouf-Bouteau H. A Correlative Study of Sunflower Seed Vigor Components as Related to Genetic Background. Plants. 2020; 9(3):386. https://doi.org/10.3390/plants9030386
Chicago/Turabian StyleSaux, Marine, Benoît Bleys, Thierry André, Christophe Bailly, and Hayat El-Maarouf-Bouteau. 2020. "A Correlative Study of Sunflower Seed Vigor Components as Related to Genetic Background" Plants 9, no. 3: 386. https://doi.org/10.3390/plants9030386