
Genome-Wide Analysis of the Wall-Associated Kinase (WAK) Genes in Medicago truncatula and Functional Characterization of MtWAK24 in Response to Pathogen Infection
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of WAKs in M. truncatula
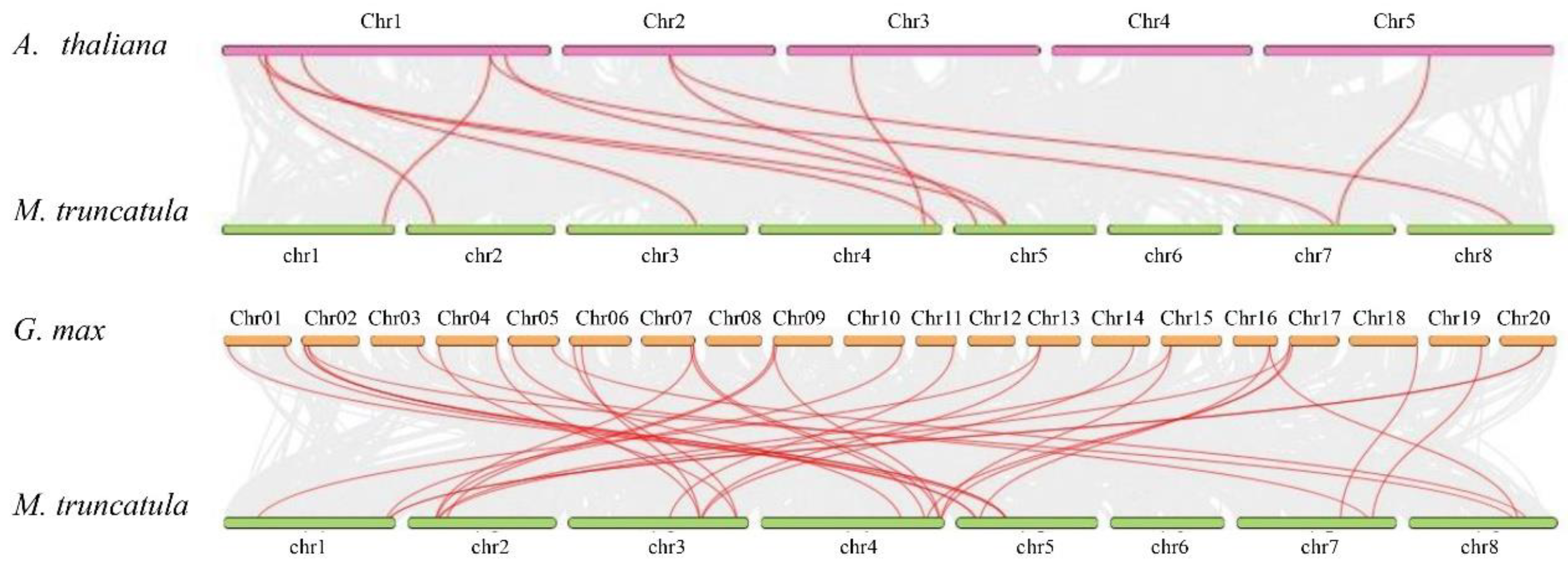
2.2. Chromosomal Location and Expansion Analysis of MtWAKs
2.3. Phylogeny, Gene Structure, Protein Domain and Motif Analysis of MtWAK-RLK Members
2.4. Analysis of Cis-Acting Elements in the Promoter Region of MtWAK-RLKs
2.5. Expression Analysis of MtWAK-RLKs across Different Tissues and Developmental Stages
2.6. Expression Analysis of MtWAK-RLKs in Response to Biotic and Abiotic Stresses
2.7. Gene Expression Validation of MtWAK-RLKs by qRT-PCR
2.8. Plasma Membrane Localized MtWAK24 Inhibited Phytophthora Infection in Tobacco
3. Discussion
4. Materials and Methods
4.1. Identification of MtWAK Genes
4.2. Chromosomal Location and Synteny Correlation Analysis
4.3. Phylogenetic Analysis and Gene Structure, Motif and Conserved Domain Analysis
4.4. Analysis of cis-Acting Regulatory Elements
4.5. Expression Pattern Analysis
4.6. Tissue Samples Collection and Cold and Salt Treatment
4.7. RNA Extraction and qRT-PCR
4.8. Subcellular Localization
4.9. Phytophthora Infection in N. benthamiana
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gish, L.A.; Clark, S.E. The RLK/Pelle family of kinases. Plant J. 2011, 66, 117–127. [Google Scholar] [CrossRef] [PubMed]
- He, Z.H.; Cheeseman, I.; He, D.; Kohorn, B.D. A cluster of five cell wall-associated receptor kinase genes, Wak1-5, are expressed in specific organs of Arabidopsis. Plant Mol. Biol. 1999, 39, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Lally, D.; Ingmire, P.; Tong, H.Y.; He, Z.H. Antisense expression of a cell wall-associated protein kinase, WAK4, inhibits cell elongation and alters morphology. Plant Cell 2001, 13, 1317–1331. [Google Scholar] [CrossRef] [PubMed]
- Kohorn, B.D.; Kobayashi, M.; Johansen, S.; Riese, J.; Huang, L.F.; Koch, K.; Fu, S.; Dotson, A.; Byers, N. An Arabidopsis cell wall-associated kinase required for invertase activity and cell growth. Plant J. 2006, 46, 307–316. [Google Scholar] [CrossRef]
- Yue, Z.L.; Liu, N.; Deng, Z.P.; Zhang, Y.; Wu, Z.M.; Zhao, J.L.; Sun, Y.; Wang, Z.Y.; Zhang, S.W. The receptor kinase OsWAK11 monitors cell wall pectin changes to fine-tune brassinosteroid signaling and regulate cell elongation in rice. Curr. Biol. 2022, 32, 2454–2466.e7. [Google Scholar] [CrossRef]
- Yin, W.; Yang, H.; Wang, Y.; Feng, P.; Deng, Y.; Zhang, L.; He, G.; Wang, N. Oryza sativa PECTIN DEFECTIVE TAPETUM1 affects anther development through a pectin-mediated signaling pathway in rice. Plant Physiol. 2022, 189, 1570–1586. [Google Scholar] [CrossRef]
- He, Z.H.; He, D.Z.; Kohorn, B.D. Requirement for the induced expression of a cell wall associated receptor kinase for survival during the pathogen response. Plant J. 1998, 14, 55–63. [Google Scholar] [CrossRef]
- Hurni, S.; Scheuermann, D.; Krattinger, S.G.; Kessel, B.; Wicker, T.; Herren, G.; Fitze, M.N.; Breen, J.; Presterl, T.; Ouzunova, M.; et al. The maize disease resistance gene Htn1 against northern corn leaf blight encodes a wall-associated receptor-like kinase. Proc. Natl. Acad. Sci. USA 2015, 112, 8780–8785. [Google Scholar] [CrossRef] [PubMed]
- Zuo, W.L.; Chao, Q.; Zhang, N.; Ye, J.R.; Tan, G.Q.; Li, B.L.; Xing, Y.X.; Zhang, B.Q.; Liu, H.J.; Fengler, K.A.; et al. A maize wall-associated kinase confers quantitative resistance to head smut. Nat. Genet. 2015, 47, 151–157. [Google Scholar] [CrossRef]
- Saintenac, C.; Lee, W.S.; Cambon, F.; Rudd, J.J.; King, R.C.; Marande, W.; Powers, S.J.; Berges, H.; Phillips, A.L.; Uauy, C.; et al. Wheat receptor-kinase-like protein Stb6 controls gene-for-gene resistance to fungal pathogen Zymoseptoria tritici. Nat. Genet. 2018, 50, 368–374. [Google Scholar] [CrossRef]
- Hu, K.; Cao, J.; Zhang, J.; Xia, F.; Ke, Y.; Zhang, H.; Xie, W.; Liu, H.; Cui, Y.; Cao, Y.; et al. Improvement of multiple agronomic traits by a disease resistance gene via cell wall reinforcement. Nat. Plants 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed]
- Delteil, A.; Gobbato, E.; Cayrol, B.; Estevan, J.; Michel-Romiti, C.; Dievart, A.; Kroj, T.; Morel, J.B. Several wall-associated kinases participate positively and negatively in basal defense against rice blast fungus. Bmc Plant Biol. 2016, 16, 17. [Google Scholar] [CrossRef] [PubMed]
- Denoux, C.; Galletti, R.; Mammarella, N.; Gopalan, S.; Werck, D.; De Lorenzo, G.; Ferrari, S.; Ausubel, F.M.; Dewdney, J. Activation of defense response pathways by OGs and Flg22 elicitors in Arabidopsis seedlings. Mol. Plant. 2008, 1, 423–445. [Google Scholar] [CrossRef]
- Sivaguru, M.; Ezaki, B.; He, Z.H.; Tong, H.; Osawa, H.; Baluska, F.; Volkmann, D.; Matsumoto, H. Aluminum-induced gene expression and protein localization of a cell wall-associated receptor kinase in Arabidopsis. Plant Physiol. 2003, 132, 2256–2266. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Tong, H.; Selby, J.; Dewitt, J.; Peng, X.; He, Z.H. Involvement of a cell wall-associated kinase, WAKL4, in Arabidopsis mineral responses. Plant Physiol. 2005, 139, 1704–1716. [Google Scholar] [CrossRef]
- Zhang, H.; Lian, C.; Shen, Z. Proteomic identification of small, copper-responsive proteins in germinating embryos of Oryza sativa. Ann. Bot. 2009, 103, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Wang, Y.H.; Liu, X.Y.; Shang, J.X.; Zhao, L.Q. OsWAK112, a wall-associated kinase, negatively regulates salt stress responses by inhibiting ethylene production. Front. Plant Sci. 2021, 12, 751965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, C.; Li, L.; Meng, L.; Singh, J.; Jiang, N.; Deng, X.W.; He, Z.H.; Lemaux, P.G. Evolutionary expansion, gene structure, and expression of the rice wall-associated kinase gene family. Plant Physiol. 2005, 139, 1107–1124. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Jiao, C.; Sun, H.H.; Rosli, H.G.; Pombo, M.A.; Zhang, P.F.; Banf, M.; Dai, X.B.; Martin, G.B.; Giovannoni, J.J.; et al. iTAK: A program for genome-wide prediction and classification of plant transcription factors, transcriptional regulators, and protein kinases. Mol. Plant 2016, 9, 1667–1670. [Google Scholar] [CrossRef]
- Verica, J.A.; Chae, L.; Tong, H.Y.; Ingmire, P.; He, Z.H. Tissue-specific and developmentally regulated expression of a cluster of tandemly arrayed cell wall-associated kinase-like kinase genes in Arabidopsis. Plant Physiol. 2003, 133, 1732–1746. [Google Scholar] [CrossRef]
- Dou, L.; Li, Z.; Shen, Q.; Shi, H.; Li, H.; Wang, W.; Zou, C.; Shang, H.; Li, H.; Xiao, G. Genome-wide characterization of the WAK gene family and expression analysis under plant hormone treatment in cotton. BMC Genom. 2021, 22, 85. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Y.; Song, Y.P.; Chen, D.; Zang, Y.D.; Zhang, Q.L.; Yi, Y.T.; Qu, G.Q. Genome-wide identification, classification, characterization, and expression analysis of the wall-associated kinase family during fruit development and under wound stress in tomato (Solanum lycopersicum L.). Genes 2020, 11, 1186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, P.; Su, T.B.; Li, P.R.; Xin, X.Y.; Wang, W.H.; Zhao, X.Y.; Yu, Y.J.; Zhang, D.S.; Yu, S.C.; et al. Comprehensive analysis of wall-associated kinase genes and their expression under abiotic and biotic stress in chinese cabbage (Brassica rapa ssp. pekinensis). J. Plant Growth Regul. 2020, 39, 72–86. [Google Scholar] [CrossRef]
- Tripathi, R.K.; Aguirre, J.A.; Singh, J. Genome-wide analysis of wall associated kinase (WAK) gene family in barley. Genomics 2021, 113, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Lv, Y.Y.; Lei, W.R.; Li, X.; Chen, Y.H.; Zheng, L.Q.; Xia, Y.; Shen, Z.G. Cloning and characterization of the Oryza sativa wall-associated kinase gene OsWAK11 and its transcriptional response to abiotic stresses. Plant Soil 2014, 384, 335–346. [Google Scholar] [CrossRef]
- Tocquard, K.; Lafon-Placette, C.; Auguin, D.; Muries, B.; Bronner, G.; Lopez, D.; Fumanal, B.; Franchel, J.; Bourgerie, S.; Maury, S.; et al. In silico study of wall-associated kinase family reveals large-scale genomic expansion potentially connected with functional diversification in Populus. Tree Genet. Genom. 2014, 10, 1135–1147. [Google Scholar] [CrossRef]
- Li, M.; Ma, J.; Liu, H.; Ou, M.; Ye, H.; Zhao, P. Identification and characterization of wall-associated kinase (WAK) and WAK-like (WAKL) gene family in Juglans regia and its wild related species Juglans mandshurica. Genes 2022, 13, 134. [Google Scholar] [CrossRef]
- Xia, X.; Zhang, X.; Zhang, Y.; Wang, L.; An, Q.; Tu, Q.; Wu, L.; Jiang, P.; Zhang, P.; Yu, L.; et al. Characterization of the WAK gene family reveals genes for FHB resistance in bread wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2022, 23, 7157. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, D.C.; Zhang, H.Y.; Gao, H.B.; Guo, X.L.; Fu, X.D.; Zhang, A.M. Isolation and characterisation of six putative wheat cell wall-associated kinases. Funct. Plant Biol. 2006, 33, 811–821. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Plant cell walls: Wall-associated kinases and cell expansion. Curr. Biol. 2001, 11, R558–R559. [Google Scholar] [CrossRef]
- Kaur, R.; Singh, K.; Singh, J. A root-specific wall-associated kinase gene, HvWAK1, regulates root growth and is highly divergent in barley and other cereals. Funct. Integr. Genom. 2013, 13, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Van Spronsen, P.C.; Bakhuizen, R.; van Brussel, A.A.; Kijne, J.W. Cell wall degradation during infection thread formation by the root nodule bacterium Rhizobium leguminosarum is a two-step process. Eur. J. Cell Biol. 1994, 64, 88–94. [Google Scholar] [PubMed]
- Wilmowicz, E.; Kucko, A.; Alche, J.D.; Czeszewska-Rosiak, G.; Florkiewicz, A.B.; Kapusta, M.; Karwaszewski, J. Remodeling of cell wall components in root nodules and flower abscission zone under drought in yellow lupine. Int. J. Mol. Sci. 2022, 23, 1680. [Google Scholar] [CrossRef] [PubMed]
- Diener, A.C.; Ausubel, F.M. RESISTANCE TO FUSARIUM OXYSPORUM 1, a dominant Arabidopsis disease-resistance gene, is not race specific. Genetics 2005, 171, 305–321. [Google Scholar] [CrossRef]
- Li, H.; Zhou, S.Y.; Zhao, W.S.; Su, S.C.; Peng, Y.L. A novel wall-associated receptor-like protein kinase gene, OsWAK1, plays important roles in rice blast disease resistance. Plant Mol. Biol. 2009, 69, 337–346. [Google Scholar] [CrossRef]
- Cayrol, B.; Delteil, A.; Gobbato, E.; Kroj, T.; Morel, J.B. Three wall-associated kinases required for rice basal immunity form protein complexes in the plasma membrane. Plant Signal. Behav. 2016, 11, e1149676. [Google Scholar] [CrossRef]
- Harkenrider, M.; Sharma, R.; De Vleesschauwer, D.; Tsao, L.; Zhang, X.T.; Chern, M.; Canlas, P.; Zuo, S.M.; Ronald, P.C. Overexpression of rice wall-associated kinase 25 (OsWAK25) alters resistance to bacterial and fungal pathogens. PLoS ONE 2016, 11, e0147310. [Google Scholar] [CrossRef]
- Rosli, H.G.; Zheng, Y.; Pombo, M.A.; Zhong, S.; Bombarely, A.; Fei, Z.; Collmer, A.; Martin, G.B. Transcriptomics-based screen for genes induced by flagellin and repressed by pathogen effectors identifies a cell wall-associated kinase involved in plant immunity. Genom. Biol. 2013, 14, R139. [Google Scholar] [CrossRef]
- Dmochowska-Boguta, M.; Kloc, Y.; Zielezinski, A.; Werecki, P.; Nadolska-Orczyk, A.; Karlowski, W.M.; Orczyk, W. TaWAK6 encoding wall-associated kinase is involved in wheat resistance to leaf rust similar to adult plant resistance. PLoS ONE 2020, 15, e0227713. [Google Scholar] [CrossRef]
- Wang, P.; Zhou, L.; Jamieson, P.; Zhang, L.; Zhao, Z.X.; Babilonia, K.; Shao, W.Y.; Wu, L.Z.; Mustafa, R.; Amin, I.; et al. The Cotton Wall-associated kinase GhWAK7A mediates responses to fungal wilt pathogens by complexing with the chitin sensory receptors. Plant Cell 2020, 32, 3978–4001. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hu, A.H.; Qi, J.J.; Dou, W.F.; Qin, X.J.; Zou, X.P.; Xu, L.Z.; Chen, S.C.; He, Y.R. CsWAKL08, a pathogen-induced wall-associated receptor-like kinase in sweet orange, confers resistance to citrus bacterial canker via ROS control and JA signaling. Hortic. Res. 2020, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Haddadi, P.; Larkan, N.J.; Van DeWouw, A.; Zhang, Y.Q.; Neik, T.X.; Beynon, E.; Bayer, P.; Edwards, D.; Batley, J.; Borhan, M.H. Brassica napus genes Rlm4 and Rlm7, conferring resistance to Leptosphaeria maculans, are alleles of the Rlm9 wall-associated kinase-like resistance locus. Plant Biotechnol. J. 2022, 20, 1229–1231. [Google Scholar] [CrossRef]
- Ameen, G.; Solanki, S.; Drader, T.; Sager-Bittara, L.; Steffenson, B.; Kleinhofs, A.; Vogiatzis, C.; Brueggeman, R.S. rcs5-mediated spot blotch resistance in barley is conferred by wall-associated kinases that resist pathogen manipulation. BioRxiv 2020. [Google Scholar] [CrossRef]
- Glawe, D.A. The powdery mildews: A review of the world’s most familiar (yet poorly known) plant pathogens. Annu. Rev. Phytopathol. 2008, 46, 27–51. [Google Scholar] [CrossRef] [PubMed]
- Meco, V.; Egea, I.; Ortiz-Atienza, A.; Drevensek, S.; Esch, E.; Yuste-Lisbona, F.J.; Barneche, F.; Vriezen, W.; Bolarin, M.C.; Lozano, R.; et al. The salt sensitivity induced by disruption of cell wall-associated kinase 1 (SlWAK1) tomato gene is linked to altered osmotic and metabolic homeostasis. Int. J. Mol. Sci. 2020, 21, 6308. [Google Scholar] [CrossRef] [PubMed]
- Giarola, V.; Krey, S.; von den Driesch, B.; Bartels, D. The Craterostigma plantagineum glycine-rich protein CpGRP1 interacts with a cell wall-associated protein kinase 1 (CpWAK1) and accumulates in leaf cell walls during dehydration. New Phytol. 2016, 210, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Sharma, G.; Saxena, D.; Budhwar, R.; Vasudevan, M.; Gupta, V.; Gupta, A.; Gupta, R.; Chandran, D. Dual RNA-Seq analysis of Medicago truncatula and the pea powdery mildew Erysiphe pisi uncovers distinct host transcriptional signatures during incompatible and compatible interactions and pathogen effector candidates. Genomics 2020, 112, 2130–2145. [Google Scholar] [CrossRef]
- Qiu, X.; Kong, L.; Chen, H.; Lin, Y.; Tu, S.; Wang, L.; Chen, Z.; Zeng, M.; Xiao, J.; Yuan, P.; et al. The Phytophthora sojae nuclear effector PsAvh110 targets a host transcriptional complex to modulate plant immunity. Plant Cell 2022, 35, 574–597. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, W.; Shi, J.; Yang, B.; Yu, S.; Zhao, P.; Guo, Z.; Zhu, H. Genome-Wide Analysis of the Wall-Associated Kinase (WAK) Genes in Medicago truncatula and Functional Characterization of MtWAK24 in Response to Pathogen Infection. Plants 2023, 12, 1849. https://doi.org/10.3390/plants12091849
Kong W, Shi J, Yang B, Yu S, Zhao P, Guo Z, Zhu H. Genome-Wide Analysis of the Wall-Associated Kinase (WAK) Genes in Medicago truncatula and Functional Characterization of MtWAK24 in Response to Pathogen Infection. Plants. 2023; 12(9):1849. https://doi.org/10.3390/plants12091849
Chicago/Turabian StyleKong, Weiyi, Jia Shi, Bo Yang, Shuhan Yu, Pengcheng Zhao, Zhenfei Guo, and Haifeng Zhu. 2023. "Genome-Wide Analysis of the Wall-Associated Kinase (WAK) Genes in Medicago truncatula and Functional Characterization of MtWAK24 in Response to Pathogen Infection" Plants 12, no. 9: 1849. https://doi.org/10.3390/plants12091849