


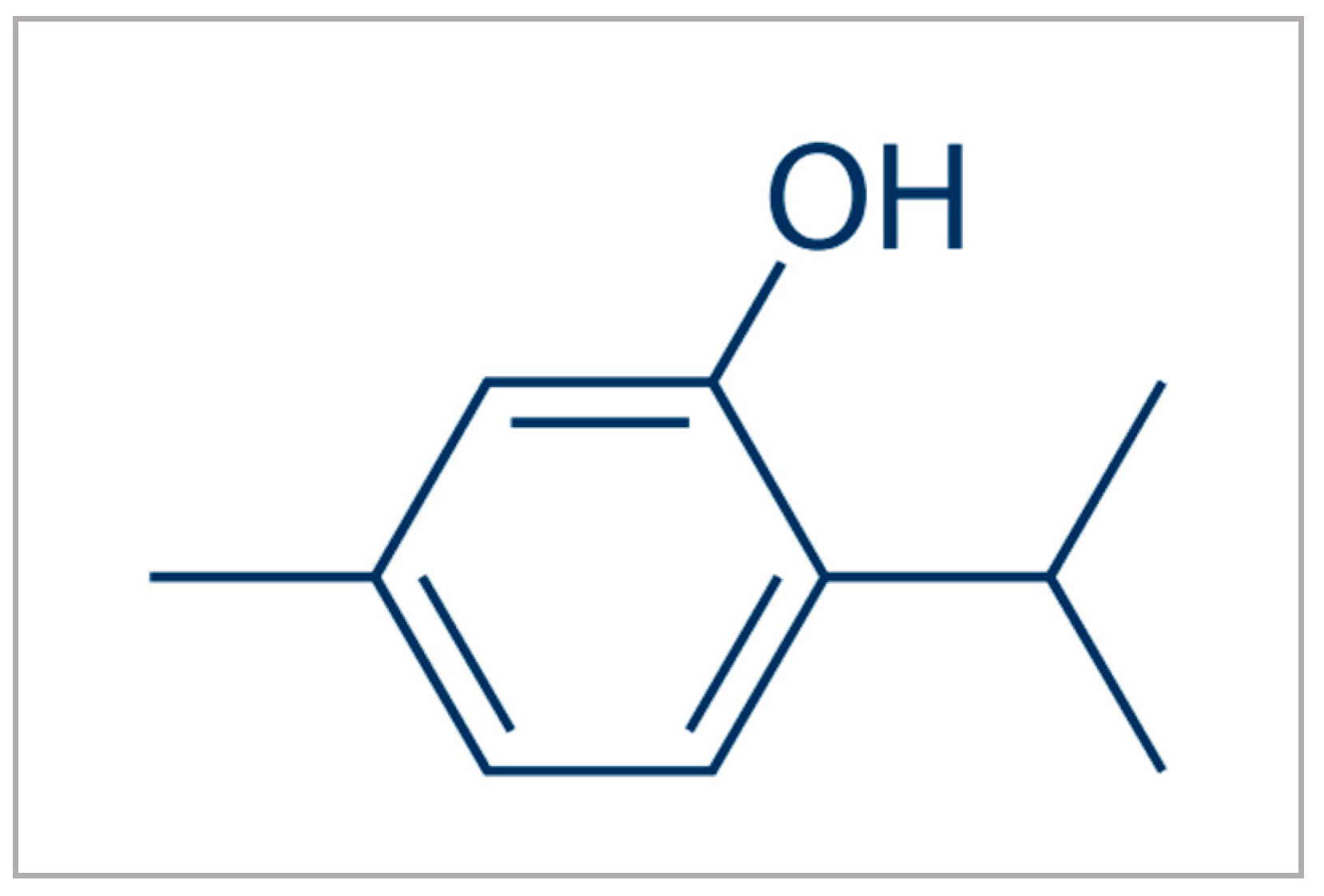
Nematicidal Potential of Thymol against Meloidogyne javanica (Treub) Chitwood
Abstract
:1. Introduction
- Assessment of the nematicidal and nematostatic activity of thymol on M. javanica J2s.
- Evaluation of thymol’s inhibition effect on undifferentiated eggs.
- Analysis of the hatch inhibition activity of thymol on egg masses.
- Examination of the contact and vapor activity of thymol in nematode-infested soil.
- Analysis of the effect of sublethal concentrations of thymol on the infectivity of M. javanica.
2. Results
2.1. Nematode Motility Bioassays
2.1.1. Nematicidal Activity
2.1.2. Nematostatic Activity
2.1.3. Effect of Thymol on Egg Development
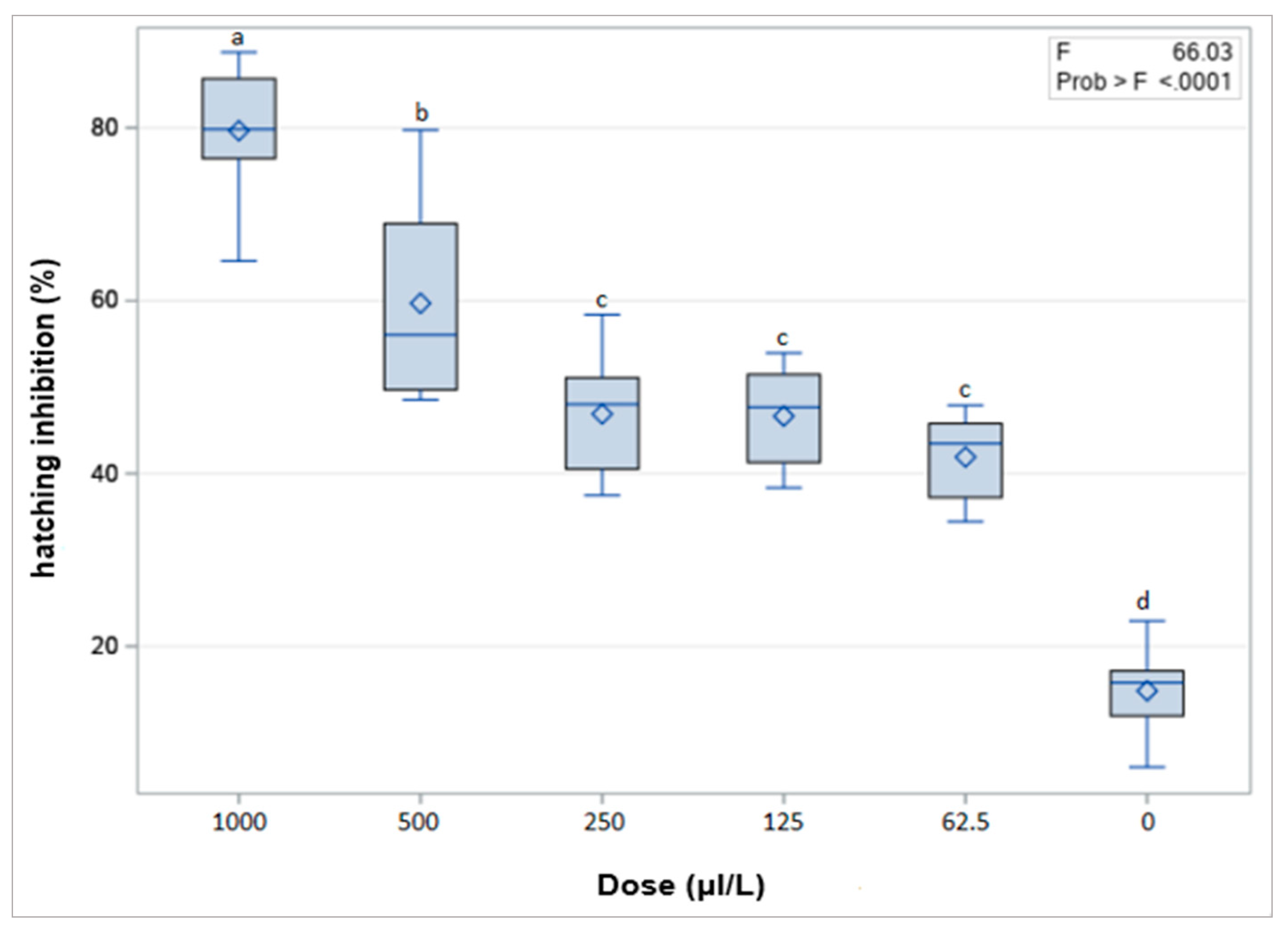
2.1.4. Effect of Thymol on Egg Hatching from Egg Masses
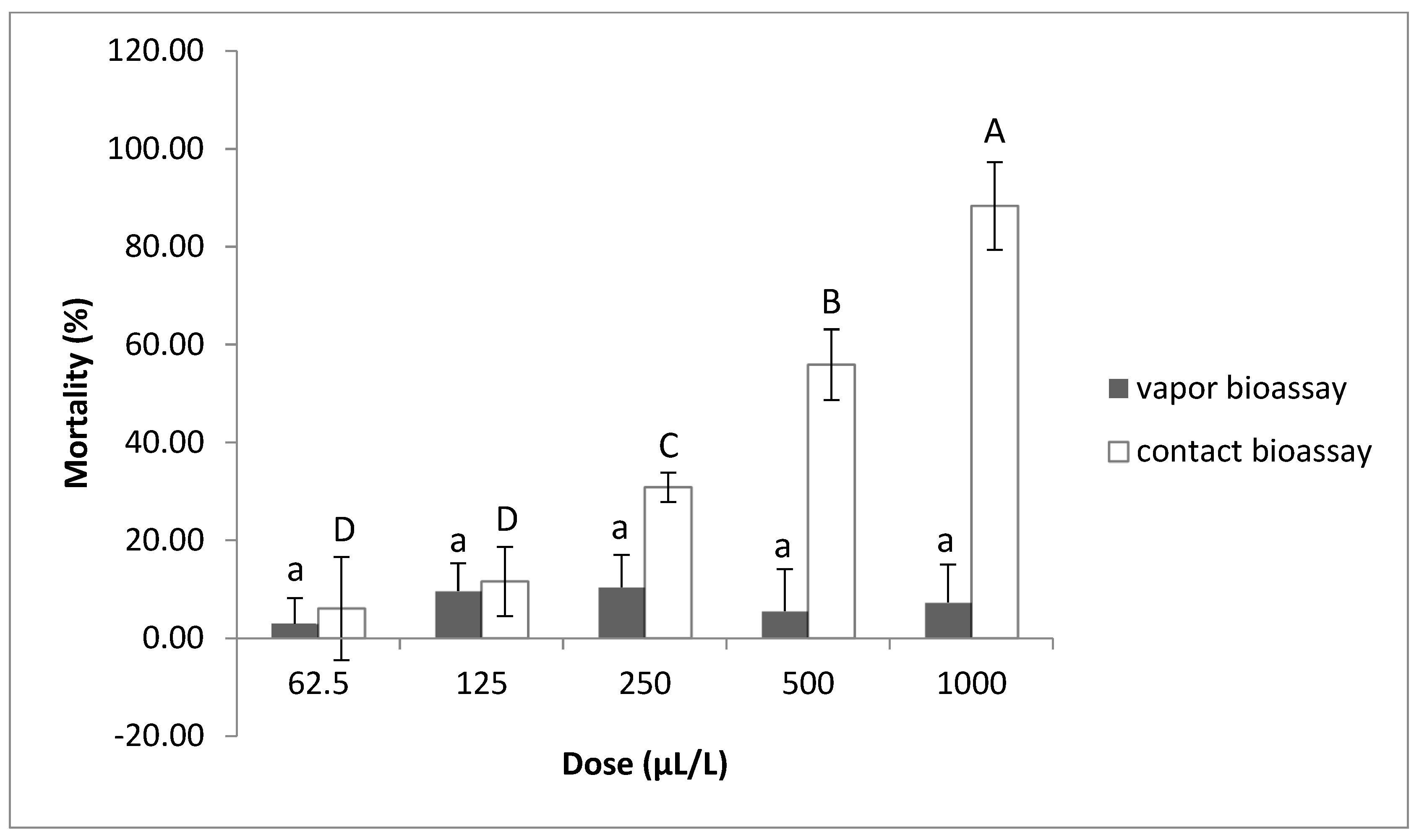
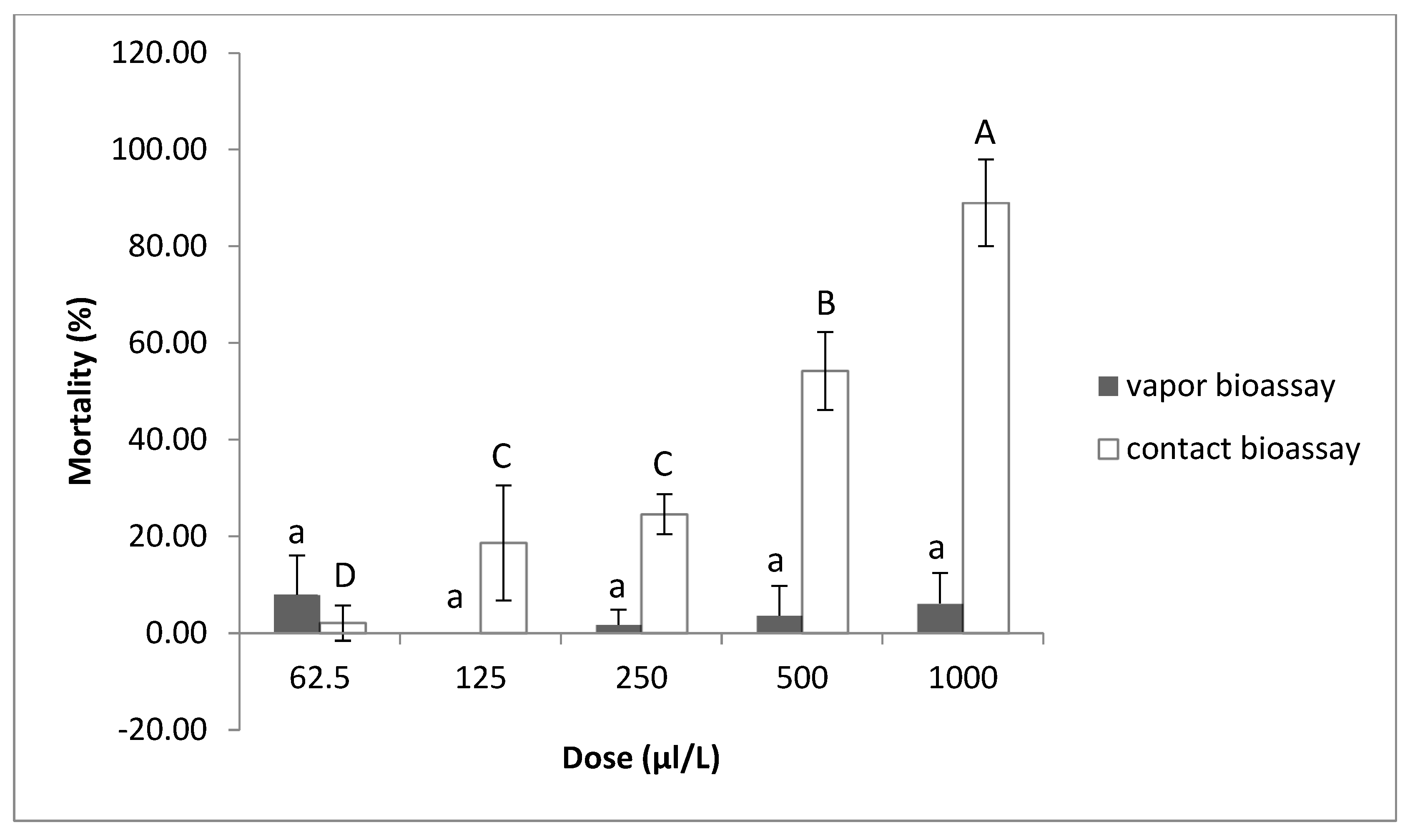
2.1.5. Contact and Vapor Effect of Thymol against M. javanica
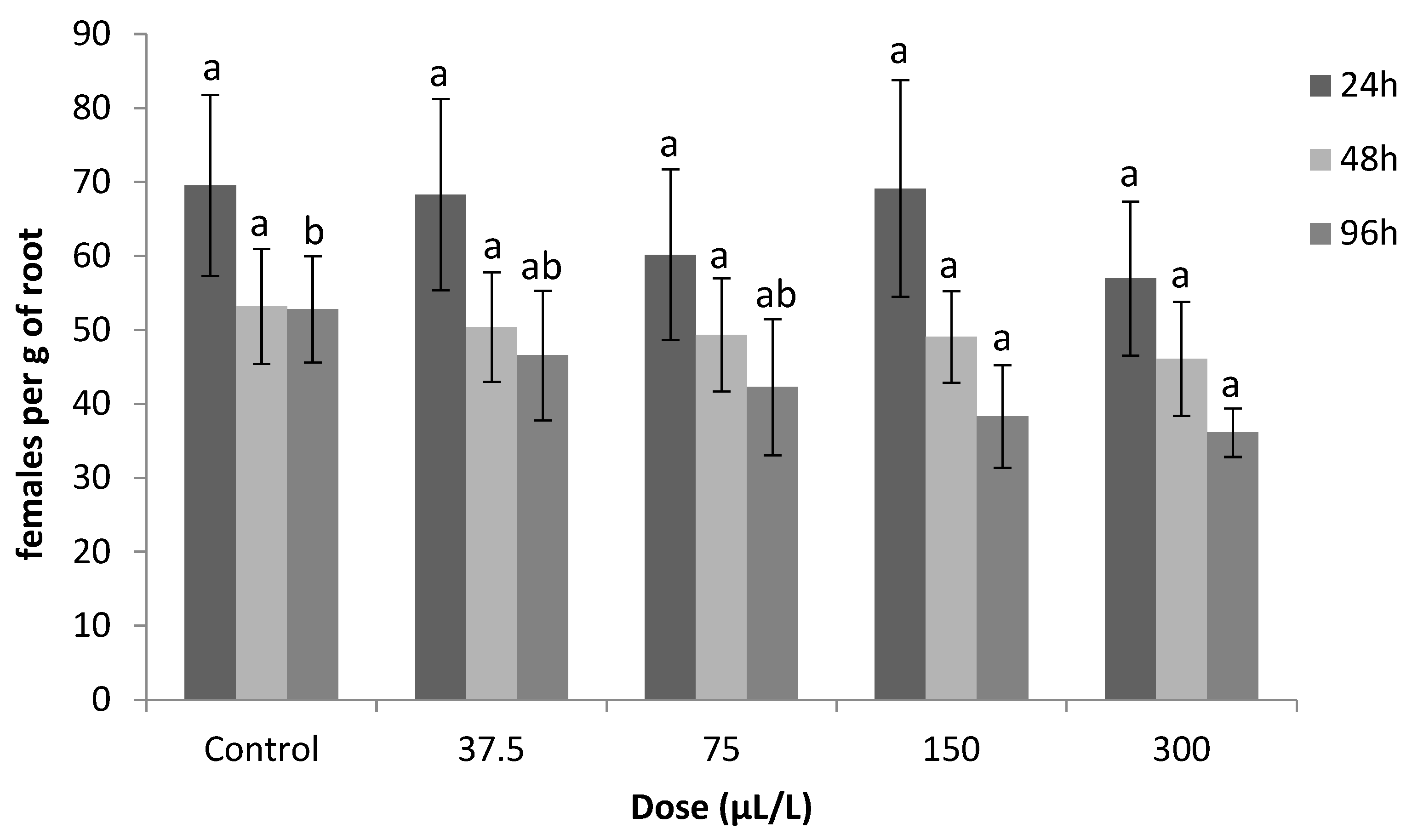
2.1.6. Sublethal Effect of Thymol on J2s Invasion (Pot Experiment)
3. Discussion
4. Materials and Methods
4.1. Nematode Cultures
4.2. Nematode Motility Bioassays
4.2.1. Nematicidal Activity
4.2.2. Nematostatic Activity
4.3. Effect of Thymol on Egg Development
4.4. Effect of Thymol on Egg Hatching from Egg Masses
4.5. Contact and Vapor Effect of Thymol against M. javanica
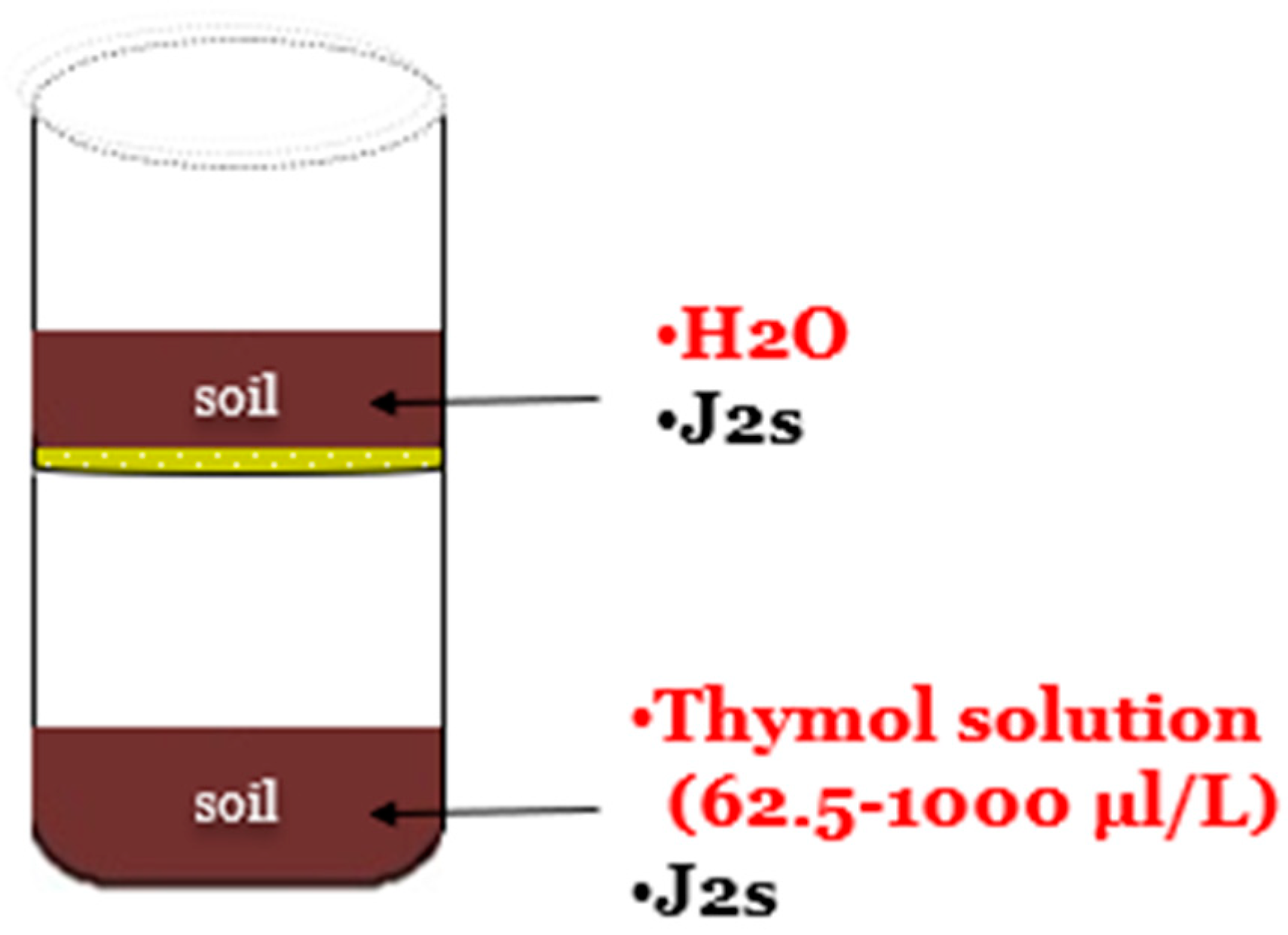
4.6. Sublethal Doses Effect of Thymol on Juvenile Invasion (Pot Experiment)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oka, Y.; Koltai, H.; Bar-Eyal, M.; Mor, M.; Sharon, E.; Chet, I.; Spiegel, Y. New strategies for the control of plant parasitic nematodes. Pest Manag. Sci. 2000, 56, 983–988. [Google Scholar] [CrossRef]
- Bleve-Zacheo, T.; Mellilo, M.T.; Castagnone-Sereno, P. The contribution of biotechnology to root-knot nematode control in tomato plants. Pest Technol. 2007, 1, 1–16. [Google Scholar]
- Caillaud, M.C.; Dubreuil, G.; Quentin, M.; Perfus-Barbeoch, L.; Lecomte, P.; de Almeida Engler, J.; Abad, P.; Rosso, M.N.; Favery, B. Root-knot nematodes manipulate plant cell functions during a compatible interaction. J. Plant Physiol. 2008, 165, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, J.J.; Sevilla-Morán, B.; Sandín-España, P.; López-Goti, C.; Alonso-Prados, J.L. Biopesticides in the framework of the European Pesticide Regulation (EC) No. 1107/2009. Pest Manag. Sci. 2014, 70, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Nyczepir, A.P.; Thomas, S.H. Current and future management strategies in intensive crop production systems. In Root-Knot Nematodes; Perry, R.N., Moens, M., Starr, J.L., Eds.; CABI: Wallingford, UK, 2009; pp. 412–443. [Google Scholar]
- Sorribas, J.; Ornat, C. Estrategias de control integrado de nematodos fitoparásitos. In Enfermedades Causadas Por Nematodos Fitoparásitos En Espana; Andrés, M.F., Verdejo, S., Eds.; Phytoma-SEF: Valencia, Spain, 2011; pp. 115–127. [Google Scholar]
- Isman, M.B.; Miresmailli, S.; Machial, C. Commercial opportunities for pesticides based on plant EOs in agriculture, industry and consumer products. Phytochem. Rev. 2011, 10, 197–204. [Google Scholar] [CrossRef]
- Martin, F.N. Development of alternative strategies for management of soilborne pathogens currently controlled with methyl bromide. Ann. Rev. Phytopathol. 2003, 41, 325–350. [Google Scholar] [CrossRef]
- Fernandez, C.; Rodríguez-Kábana, R.; Warrior, P.; Kloepper, J. Induced soil suppressiveness to a root-knot nematode species by a nematicide. Biol. Control 2001, 22, 103–114. [Google Scholar] [CrossRef]
- Davies, K.; Spiegel, Y. Biological Control of Plant-Parasitic Nematodes: Building Coherence between Microbial Ecology and Molecular Mechanisms; Springer: Dordrecht, The Netherlands, 2011; Volume 11. [Google Scholar]
- Giannakou, I.O. Efficacy of a formulated product containing Quillaja Saponaria plant extracts for the control of root-knot nematodes. E. J. Plant Pathol. 2011, 130, 587–596. [Google Scholar] [CrossRef]
- Pardavella, I.; Daferera, D.; Tselios, T.; Skiada, P.; Giannakou, I. The use of essential oil and hydrosol extracted from Cuminum cyminum seeds for the control of Meloidogyne incognita and Meloidogyne javanica. Plants 2021, 10, 46. [Google Scholar] [CrossRef]
- Wink, M. Production and Application of Phytochemicals from an agricultural perspective. In Phytochemistry and Agriculture Clarendon; Beek, V.T.A., Breteler, H., Eds.; Clarendon Press: Oxford, UK, 1993; Volume 34, pp. 171–213. [Google Scholar]
- Akhtar, M.; Mahmood, I. Potentiality of phytochemicals in nematode control: A review. Bioresour. Technol. 1994, 48, 189–201. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Ann. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Kabera, J.N.; Semana, E.; Mussa, A.R.; He, X. Plant secondary metabolites: Biosynthesis, classification, Function and Pharmacological Properties. J. Pharm. Pharmacol. 2014, 2, 377–392. [Google Scholar]
- Sousa, R.; Rosa, J.; Silva, C.; Almeida, M.; Novo, M.; Cunha, A.; Fernandes-Ferreira, M. Larvicidal, molluscicidal and nematicidal activities of essential oils and compounds from Foeniculum vulgare. J. Pest Sci. 2015, 88, 413–426. [Google Scholar] [CrossRef]
- Mahizan, N.A.; Yang, S.K.; Moo, C.L.; Song, A.A.L.; Chong, C.M.; Chong, C.W.; Abushelaibi, A.; Lim, S.E.; Lai, K.S. Terpene Derivatives as a Potential Agent against Antimicrobial Resistance (AMR) Pathogens. Molecules 2019, 24, 2631. [Google Scholar] [CrossRef]
- Xu, Y.; Tong, Z.; Zhang, X.; Wang, Y.; Fang, Y.; Li, L.; Luo, Z. Unveiling the Mechanisms for the Plant Volatile Organic Compound Linalool to Control Gray Mold on Strawberry Fruits. J. Agric. Food Chem. 2019, 67, 9265–9276. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Nasiou, E.; Skiada, P.; Giannakou, I.O. Effects of four terpenes on the mortality of Ditylenchus dipsaci (Kühn). E J. Plant Pathol. 2021, 160, 137–146. [Google Scholar] [CrossRef]
- Nasiou, E.; Giannakou, I.O. The potential of eugenol as a nematicidal agent against Meloidogyne javanica (Treub) Chitwood. J. Nematol. 2020, 52, e2020-103. [Google Scholar] [CrossRef]
- Jyoti-Dheer, D.; Singh, D.; Kumar, G.; Karnatak, S.C.; Prakash-Verma, V.P.; Shankar, R. Thymol chemistry: A medicinal toolbox. Cur. Bioact. Comp. 2019, 15, 456–475. [Google Scholar]
- Andrés, M.F.; González-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematicidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef]
- Chillet, M.; Minier, J.; Hoarau, M.; Meile, J.-C. Potential use of thymol to control anthracnose development in mango. E. J. Plant Pathol. 2019, 155, 943–952. [Google Scholar] [CrossRef]
- Mongiano, G.; Zampieri, E.; Morcia, C.; Titone, P.; Volante, A.; Terzi, V.; Valé, G.; Monaco, S. Application of plant-derived bioactive compounds as seed treatments to manage the rice pathogen Fusarium fujikuroi. Crop Prot. 2021, 148, 105739. [Google Scholar] [CrossRef]
- Kachur, K.; Suntres, Z. The antibacterial properties of phenolic isomers, carvacrol and thymol. Crit. Rev. Food Sci. Nutr. 2020, 60, 3042–3053. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; El-Arami, S.A.A.; Abdelgaleil, S.A.M. Acaricidal and quantitative structure activity relationship of monoterpenes against the two-spotted spider mite, Tetranychus urticae. Exp. Appl. Acarol. 2010, 52, 261–274. [Google Scholar] [CrossRef]
- Katiki, L.M.; Barbieri, A.M.E.; Araujo, R.C.; Veríssimo, C.J.; Louvandini, H.; Ferreira, J.F.S. Synergistic interaction of ten essential oils against Haemonchus contortus in vitro. Vet. Parasitol. 2017, 243, 47–51. [Google Scholar] [CrossRef]
- Ibrahim, S.K.; Traboulsi, A.F.; El-Haj, S. Effect of essential oils and plant extracts on hatching, migration and mortality of Meloidogyne incognita. Phytop. Medit 2006, 45, 238–246. [Google Scholar]
- Choi, I.H.; Kim, J.; Shin, S.C.; Park, I.K. Nematicidal activity of monoterpenoids against the pine wood nematode (Bursaphelenchus xylophilus). R. J. Nematol. 2007, 15, 35–40. [Google Scholar]
- Kong, J.O.; Park, I.K.; Choi, K.S.; Shin, S.C.; Ahn, Y.J. Nematicidal and propagation activities of thyme red and white oil compounds toward Bursaphelenchus xylophilus (Nematoda: Parasitaphelenchidae). J. Nematol. 2007, 39, 237–242. [Google Scholar]
- Oka, Y.; Nacar, S.; Putievsky, E.; Ravid, U.; Yaniv, Z.; Spiegel, Y. Nematicidal activity of essential oils and their components against the root-knot nematode. Phytopathology 2000, 90, 710–715. [Google Scholar] [CrossRef]
- Avato, P.; Laquale, S.; Argentieri, M.P.; Lamiri, A.; Radicci, V.; D’Addabbo, T. Nematicidal activity of essential oils from aromatic plants of Morocco. J. Pest Sci. 2017, 90, 711–722. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Ferrari, F.; Giannakou, I.; Menkissoglu-Spiroudi, U. Phytochemistry and nematicidal activity of the essential oils from 8 greek lamiaceae aromatic plants and 13 terpene components. J. Agric. Food Chem. 2010, 58, 7856–7863. [Google Scholar] [CrossRef] [PubMed]
- Wharton, D. Nematode eggshells. Parasitology 1980, 81, 447–463. [Google Scholar] [CrossRef] [PubMed]
- Nasiou, E.; Giannakou, I.O. The potential use of carvacrol for the control of Meloidogyne javanica. E. J. Plant Pathol. 2017, 149, 415–424. [Google Scholar] [CrossRef]
- Nasiou, E.; Giannakou, I.O. Effect of geraniol, a plant based alcohol monoterpene oil, against Meloidogyne javanica. E. J. Plant Pathol. 2018, 152, 701–710. [Google Scholar] [CrossRef]
- Hussey, R.S.; Barker, K.R. A comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Tzortzakakis, E.A.; Trudgill, D.L. A comparative study of the thermal time requirements for embryogenesis in Meloidogyne javanica and M. incognita. Nematology 2005, 7, 313–315. [Google Scholar]
- Pantelelis, I.; Karpouzas, D.G.; Menkissoglu-Spiroudi, U.; Tsiropoulos, N.G. Influence of soil physicochemical and biological properties on the degradation and adsorption of the nematicide fosthiazate. J. Agric. Food Chem. 2006, 54, 6783–6789. [Google Scholar] [CrossRef]
- Flegg, J.J.M. Extraction of Xiphinema and Longidorus species from soil by a modification of Cobb’s decanting and sieving technique. Ann. Appl. Biol. 1967, 60, 429–437. [Google Scholar] [CrossRef]
- Abbot, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Byrd, D.W.; Krickpatrick, T.; Barker, K.R. An improved technique for cleaning and staining plant tissue for detection of nematodes. J. Nematol. 1983, 15, 142–143. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose (μL/L) | Exposure Time (hours) | |||
|---|---|---|---|---|
| 12 | 24 | 48 | 96 | |
| Paralyzed J2s (%) | Paralyzed J2s (%) | Paralyzed J2s (%) | Paralyzed J2s (%) | |
| 0 | 0 d | 0.18 f | 1.1 f | 2.0 f |
| 62.5 | 0.4 d | 1.6 e | 5.1 e | 10.0 e |
| 125 | 0.6 d | 5.7 d | 9.1 d | 15.2 d |
| 250 | 14.2 c | 13.9 c | 16.5 c | 31.5 c |
| 500 | 23.5 b | 39.7 b | 71.8 b | 96.4 b |
| 1000 | 81.2 a | 98.0 a | 100 a | 100 a |
| Concentration (μL/L) | Exposure Time (21 Days) |
|---|---|
| Eggs Differentiation (%) | |
| 0 | 92.2 a 1 |
| 62.5 | 84.1 ab |
| 125 | 81.2 bc |
| 250 | 74.9 c |
| 500 | 75.0 c |
| 1000 | 60.4 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasiou, E.; Giannakou, I.O. Nematicidal Potential of Thymol against Meloidogyne javanica (Treub) Chitwood. Plants 2023, 12, 1851. https://doi.org/10.3390/plants12091851
Nasiou E, Giannakou IO. Nematicidal Potential of Thymol against Meloidogyne javanica (Treub) Chitwood. Plants. 2023; 12(9):1851. https://doi.org/10.3390/plants12091851
Chicago/Turabian StyleNasiou, Eleni, and Ioannis O. Giannakou. 2023. "Nematicidal Potential of Thymol against Meloidogyne javanica (Treub) Chitwood" Plants 12, no. 9: 1851. https://doi.org/10.3390/plants12091851