
Genetic Variability in Carotenoid Contents in a Panel of Genebank Accessions of Temperate Maize from Southeast Europe
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Introduction
2.2. Analysis of Variance
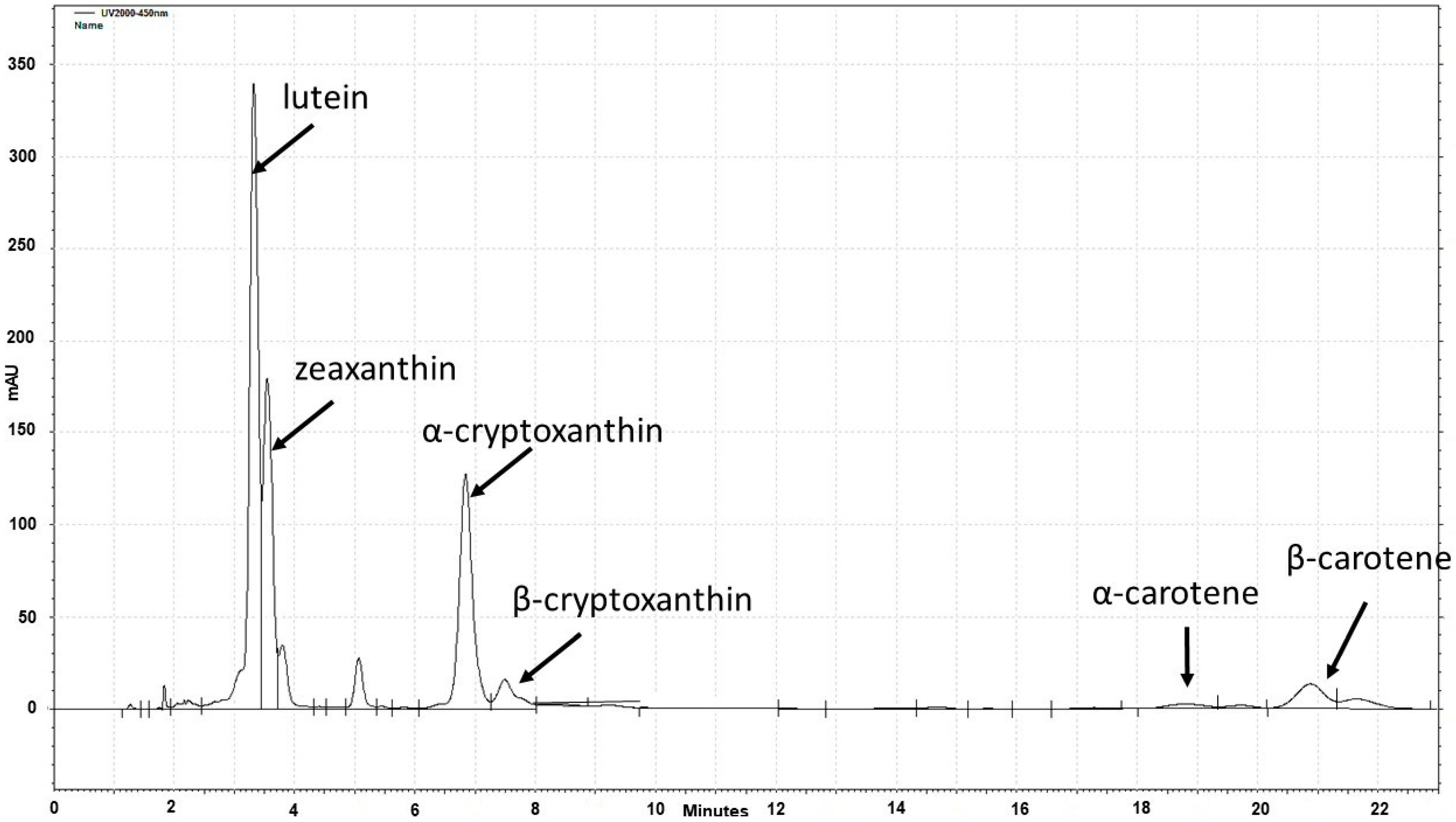
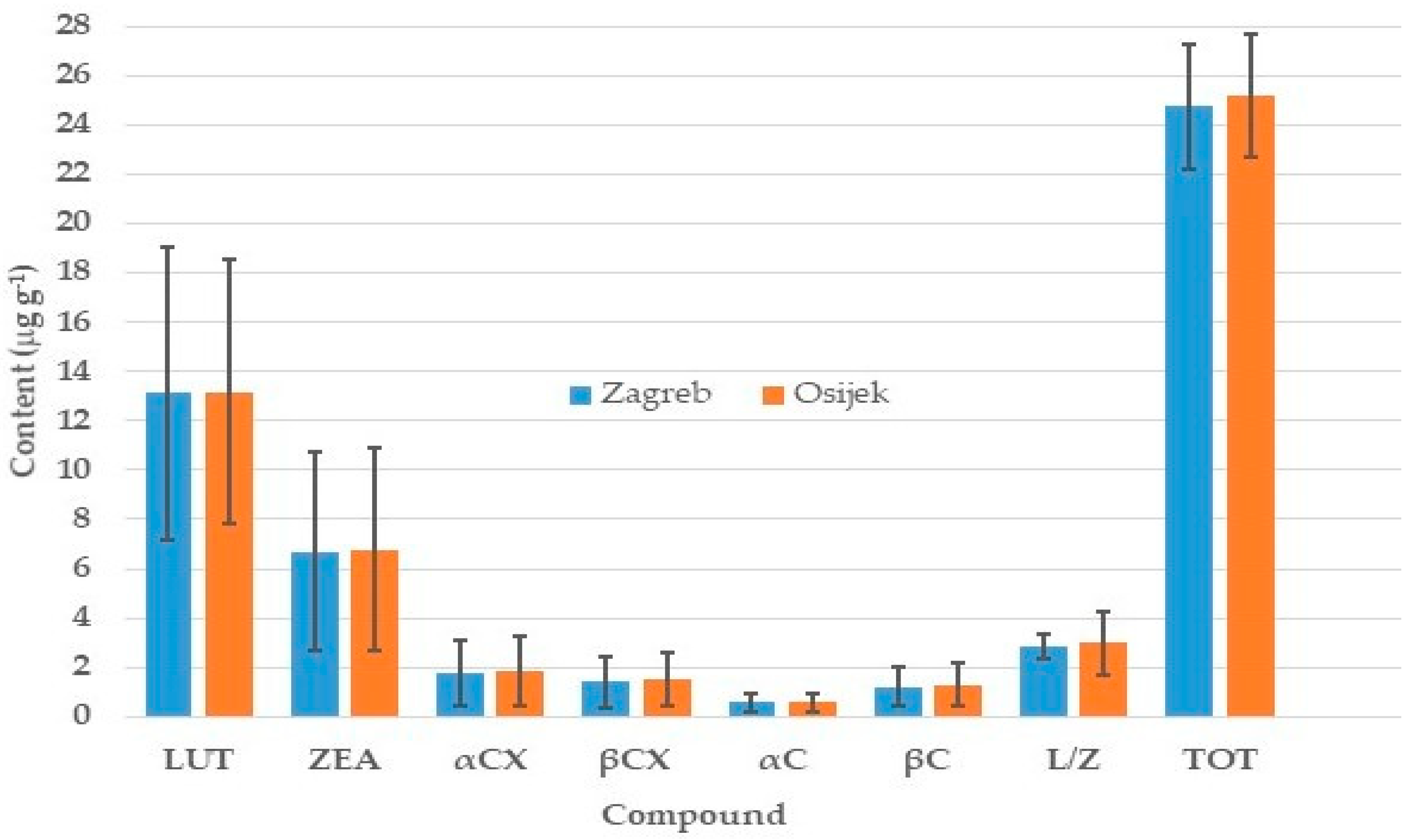
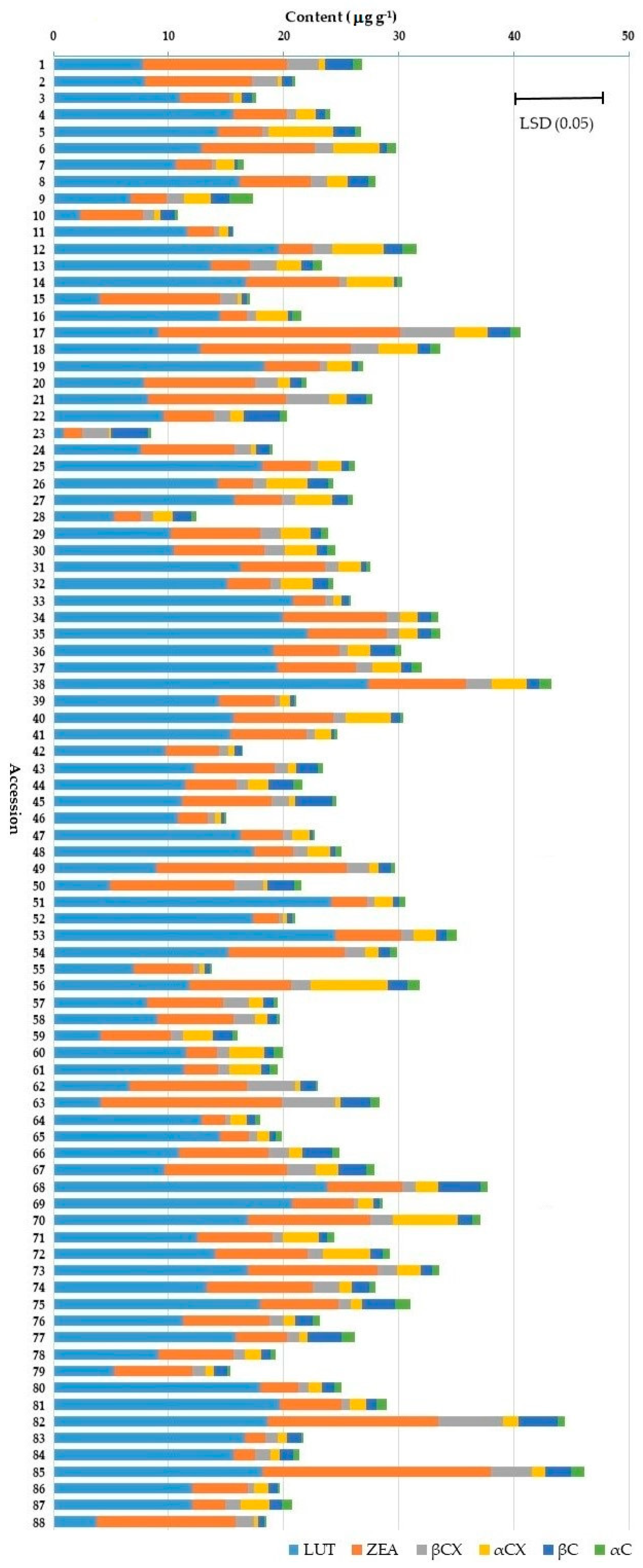
2.3. Mean and Variation in Carotenoid Contents
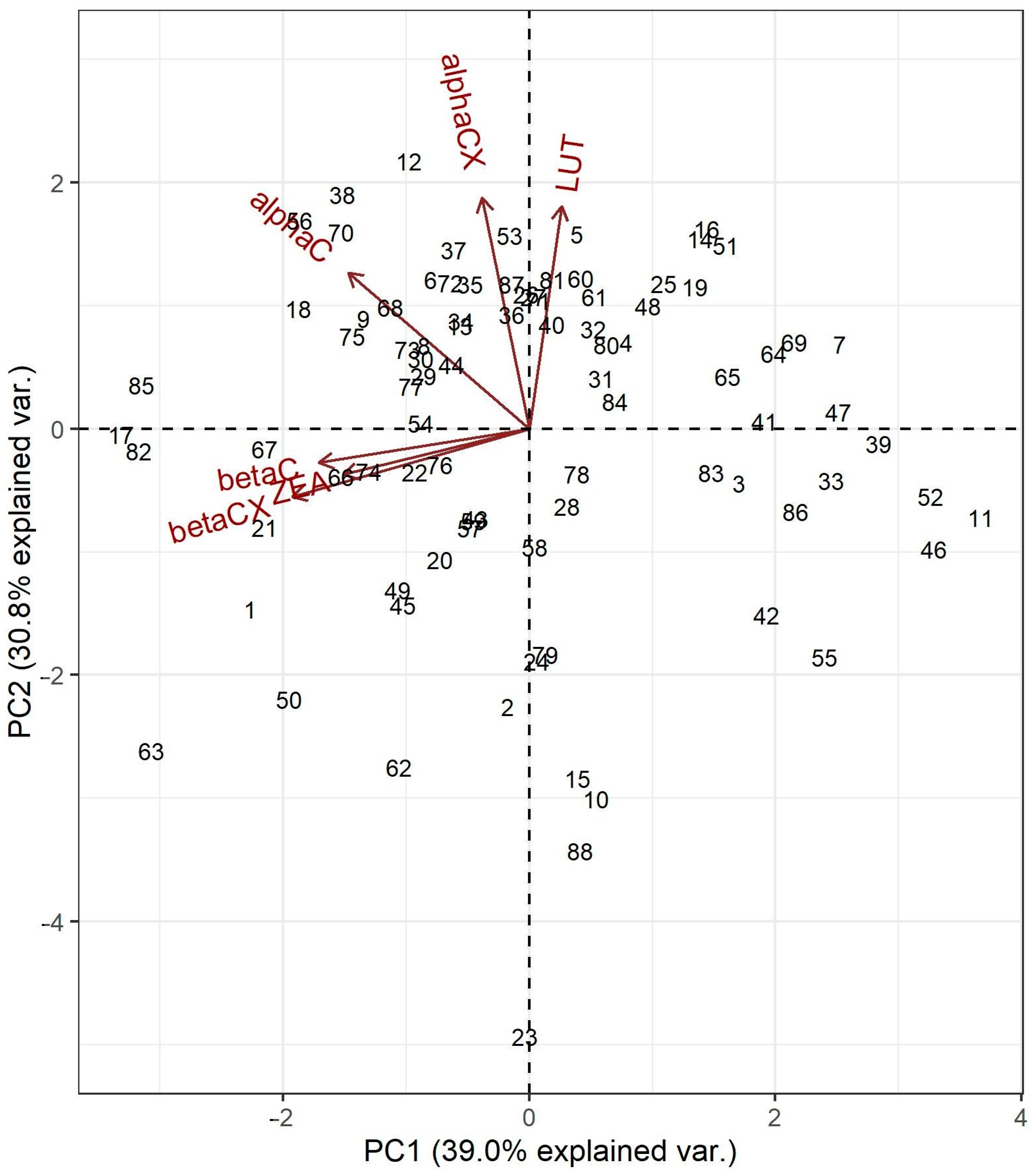
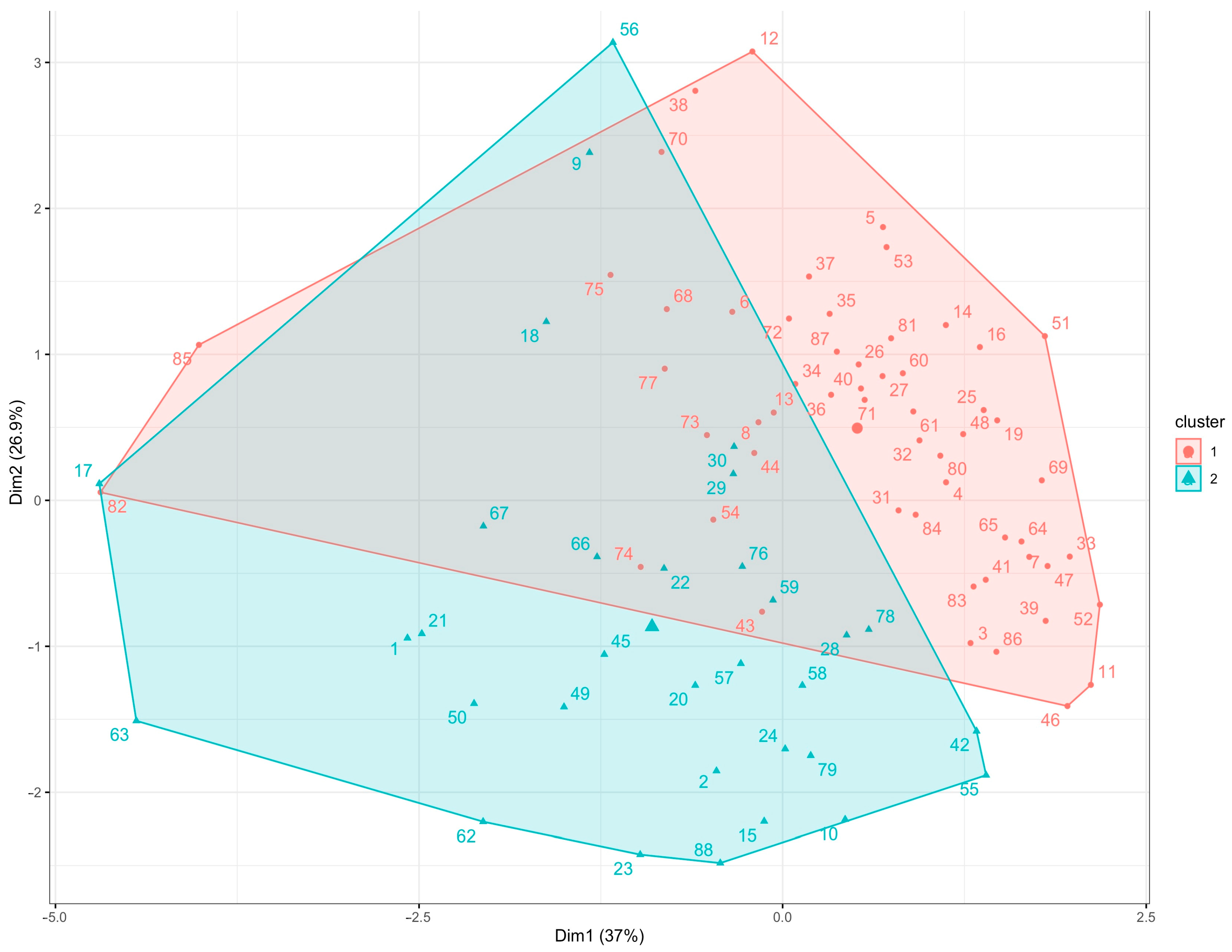
2.4. Multivariate Analyses
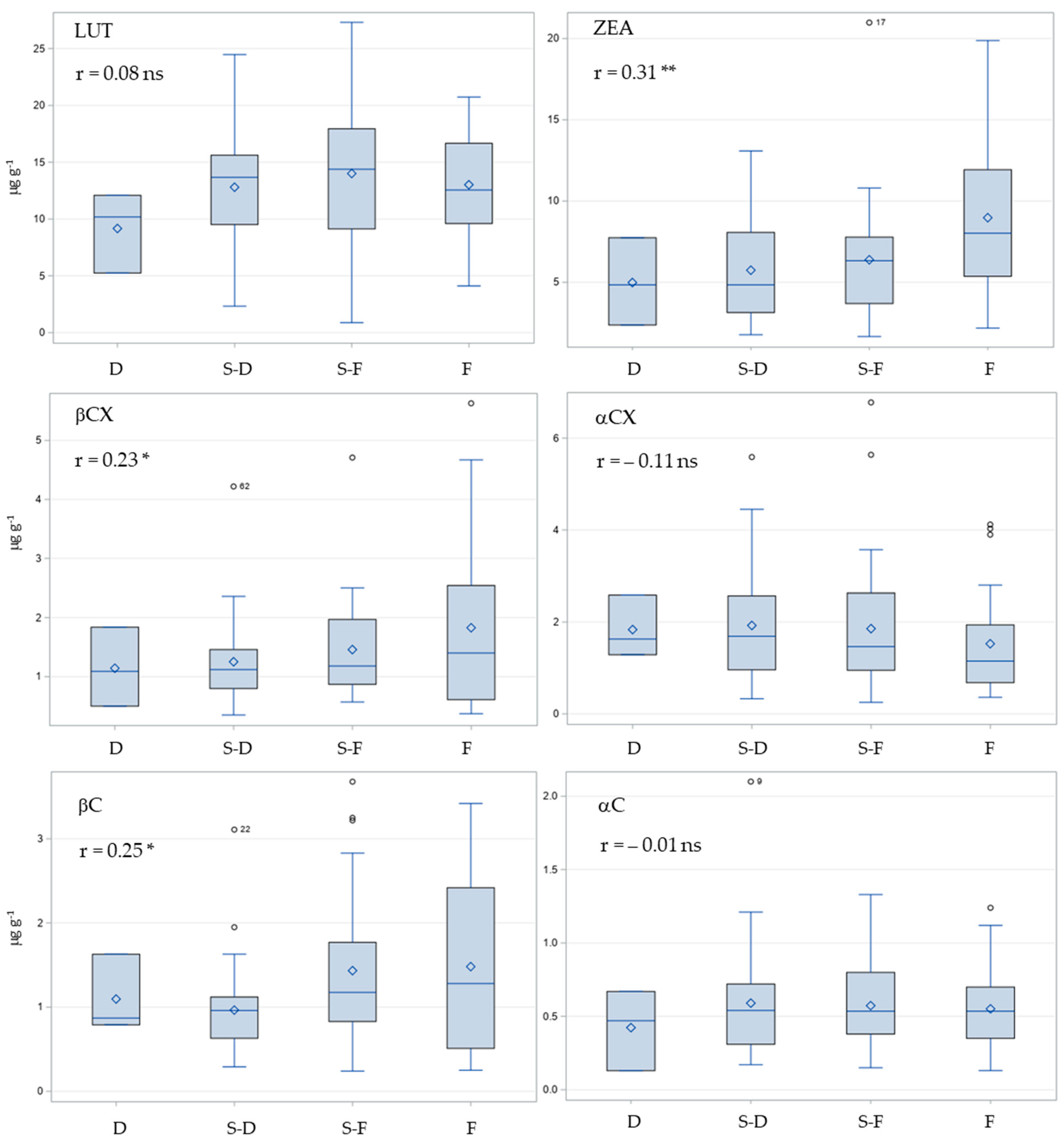
2.5. Correlations of Carotenoid Contents with Kernel Color and Kernel Hardness
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Demmig-Adams, B.; Gilmore, A.M.; Adams Iii, W.W. In vivo functions of carotenoids in higher plants. FASEB J. 1996, 10, 403–412. [Google Scholar] [CrossRef]
- Strzałka, K.; Kostecka-Gugała, A.; Latowski, D. Carotenoids and environmental stress in plants: Significance of carotenoid-mediated modulation of membrane physical properties. Russ. J. Plant Physiol. 2003, 50, 168–173. [Google Scholar] [CrossRef]
- Tanumihardjo, S.A. (Ed.) Carotenoids and Human Health; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Pixley, K.; Palacios Rojas, N.; Babu, R.; Mutale, R.; Surles, R.; Simpungwe, E. Biofortification of maize with provitamin A carotenoids. In Carotenoids and Human Health; Tanumihardjo, S.A., Ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; pp. 271–292. [Google Scholar]
- Ma, L.; Dou, H.L.; Wu, Y.Q.; Huang, Y.M.; Huang, Y.B.; Xu, X.R.; Zou, Z.Y.; Lin, X.M. Lutein and zeaxanthin intake and the risk of age-related macular degeneration: A systematic review and meta-analysis. Br. J. Nutr. 2012, 107, 350–359. [Google Scholar] [CrossRef]
- Bone, R.A.; Landrum, J.T.; Friedes, L.M.; Gomez, C.M.; Kilburn, M.D.; Menendez, E.; Vidal, I.; Wang, W. Distribution of lutein and zeaxanthin stereoisomers in the human retina. Exp. Eye Res. 1997, 64, 211–218. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S.G. Role of vitamin A in the immune system. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef] [PubMed]
- Burri, B.J. Beta-cryptoxanthin as a source of vitamin A. J. Sci. Food Agric. 2015, 95, 1786–1794. [Google Scholar] [CrossRef]
- Song, J.; Li, D.; He, M.; Chen, J.; Liu, C. Comparison of carotenoid composition in immature and mature grains of corn (Zea mays L.) varieties. Int. J. Food Prop. 2016, 19, 351–358. [Google Scholar] [CrossRef]
- Zhang, S.; Ji, J.; Zhang, S.; Guan, C.; Wang, G. Effects of three cooking methods on content changes and absorption efficiencies of carotenoids in maize. Food Funct. 2020, 11, 944–954. [Google Scholar] [CrossRef]
- De Moura, F.F.; Miloff, A.; Boy, E. Retention of provitamin A carotenoids in staple crops targeted for biofortification in Africa: Cassava, maize and sweet potato. Crit. Rev. Food Sci. Nutr. 2015, 55, 1246–1269. [Google Scholar] [CrossRef]
- Muzhingi, T.; Palacios-Rojas, N.; Miranda, A.; Cabrera, M.L.; Yeum, K.J.; Tang, G. Genetic variation of carotenoids, vitamin E and phenolic compounds in Provitamin A biofortified maize. J. Sci. Food Agric. 2017, 97, 793–801. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Chilungo, S.; Memba, A.; Mponela, P. Biofortification of maize and sweetpotatoes with provitamin A carotenoids and implication on eradicating vitamin A deficiency in developing countries. J. Agric. Food Res. 2020, 2, 100068. [Google Scholar] [CrossRef]
- Egesel, C.O.; Wong, J.C.; Lambert, R.J.; Rocheford, T.R. Combining ability of maize inbreds for carotenoids and tocopherols. Crop Sci. 2003, 43, 818–823. [Google Scholar] [CrossRef]
- Burt, A.J.; Grainger, C.M.; Smid, M.P.; Shelp, B.J.; Lee, E.A. Allele mining of exotic maize germplasm to enhance macular carotenoids. Crop Sci. 2011, 51, 991–1004. [Google Scholar] [CrossRef]
- Zheng, X.; Giuliano, G.; Al-Babili, S. Carotenoid biofortification in crop plants: Citius, altius, fortius. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158664. [Google Scholar] [CrossRef]
- Berardo, N.; Mazzinelli, G.; Valoti, P.; Laganà, P.; Redaelli, R. Characterisation of maize germplasm for the chemical composition of the grain. J. Agric. Food Chem. 2009, 57, 2378–2384. [Google Scholar] [CrossRef]
- Alfieri, M.; Hidalgo, A.; Berardo, N.; Redaelli, R. Carotenoid composition and heterotic effect in selected Italian maize germplasm. J. Cereal Sci. 2014, 59, 181–188. [Google Scholar] [CrossRef]
- Revilla, P.; Alves, M.L.; Anđelković, V.; Balconi, C.; Dinis, I.; Mendes-Moreira, P.; Redaelli, R.; Ruiz de Galarreta, J.I.; Vaz Patto, M.C.; Žilić, S.; et al. Traditional foods from maize (Zea mays L.) in Europe. Front. Nutr. 2022, 8, 683399. [Google Scholar] [CrossRef]
- Calugar, R.E.; Muntean, E.; Varga, A.; Vana, C.D.; Has, V.V.; Tritean, N.; Ceclan, L.A. Improving the Carotenoid Content in Maize by Using Isonuclear Lines. Plants 2022, 11, 1632. [Google Scholar] [CrossRef]
- Niaz, M.; Zhang, B.; Zhang, Y.; Yan, X.; Yuan, M.; Cheng, Y.; Lv, G.; Fadlalla, T.; Zhao, L.; Sun, C.; et al. Genetic and molecular basis of carotenoid metabolism in cereals. Theor. Appl. Genet. 2023, 136, 63. [Google Scholar] [CrossRef]
- Zurak, D.; Grbeša, D.; Duvnjak, M.; Kiš, G.; Međimurec, T.; Kljak, K. Carotenoid content and bioaccessibility in commercial maize hybrids. Agriculture 2021, 11, 586. [Google Scholar] [CrossRef]
- Mangelsdorf, P.C.; Fraps, G.S. A direct quantitative relationship between vitamin A in corn and the number of genes for yellow pigmentation. Science 1931, 73, 241–242. [Google Scholar] [CrossRef] [PubMed]
- Hallauer, A.R.; Carena, M.J.; Filho, J.B.M. Quantitative Genetics in Maize Breeding; Springer: New York, NY, USA, 2010. [Google Scholar] [CrossRef]
- Diepenbrock, C.H.; Ilut, D.C.; Magallanes-Lundback, M.; Kandianis, C.B.; Lipka, A.E.; Bradbury, P.J.; DellaPenna, D. Eleven biosynthetic genes explain the majority of natural variation in carotenoid levels in maize grain. Plant Cell 2021, 33, 882–900. [Google Scholar] [CrossRef] [PubMed]
- Muthusamy, V.; Hossain, F.; Thirunavukkarasu, N.; Saha, S.; Agrawal, P.K.; Guleria, S.K.; Gupta, H.S. Genetic variability and inter-relationship of kernel carotenoids among indigenous and exotic maize (Zea mays L.) inbreds. Cereal Res. Comm. 2015, 43, 567–578. [Google Scholar] [CrossRef]
- LaPorte, M.F.; Vachev, M.; Fenn, M.; Diepenbrock, C. Simultaneous dissection of grain carotenoid levels and kernel color in biparental maize populations with yellow-to-orange grain. G3 Genes Genom. Genet. 2022, 12, jkac006. [Google Scholar] [CrossRef]
- Owens, B.F.; Mathew, D.; Diepenbrock, C.H.; Tiede, T.; Wu, D.; Mateos-Hernandez, M.; Gore, M.A.; Rocheford, T. Genome-wide association study and pathway-level analysis of kernel color in maize. G3 Genes Genom. Genet. 2014, 9, 1945–1955. [Google Scholar] [CrossRef]
- Nuss, E.T.; Tanumihardjo, S.A. Maize: A paramount staple crop in the context of global nutrition. Compr. Rev. Food Sci. Saf. 2010, 9, 417–436. [Google Scholar] [CrossRef]
- Saenz, E.; Abdala, L.J.; Borrás, L.; Gerde, J.A. Maize kernel color depends on the interaction between hardness and carotenoid concentration. J. Cereal. Sci. 2020, 91, 102901. [Google Scholar] [CrossRef]
- Saenz, E.; Borrás, L.; Gerde, J.A. Carotenoid profiles in maize genotypes with contrasting kernel hardness. J. Cereal. Sci. 2021, 99, 103206. [Google Scholar] [CrossRef]
- Šimić, D.; Presterl, T.; Seitz, G.; Geiger, H.H. Comparing methods for integrating exotic germplasm into European forage maize breeding programs. Crop Sci. 2003, 43, 1952–1959. [Google Scholar] [CrossRef]
- Galić, V.; Anđelković, V.; Kravić, N.; Grčić, N.; Ledenčan, T.; Jambrović, A.; Zdunić, Z.; Nicolas, S.; Charcosset, A.; Šatović, Z.; et al. Genetic diversity and selection signatures in a gene bank panel of maize inbred lines from Southeast Europe compared with two West European panels. BMC Plant Biol. 2023, 23, 118. [Google Scholar] [CrossRef]
- Croatian Plant Genetic Resources Database. Available online: https://cpgrd.hapih.hr/gb/cm/main/accessions_list?has_document=ZEA (accessed on 2 April 2023).
- Leng, E.R.; Tavčar, A.; Trifunović, V. Maize of southeastern Europe and its potential value in breeding programs elsewhere. Euphytica 1962, 11, 263–272. [Google Scholar] [CrossRef]
- IBPGR. Descriptors for Maize; International Maize and Wheat Improvement Center, Mexico City—International Board for Plant Genetic Resources: Rome, Italy, 1991. [Google Scholar]
- Kurilich, A.C.; Juvik, J.A. Quantification of carotenoid and tocopherol antioxidants in Zea mays. J. Agric. Food Chem. 1999, 47, 1948–1955. [Google Scholar] [CrossRef] [PubMed]
- Utz, H. PLABSTAT: A Computer Program for the Statistical Analysis of Plant Breeding Experiments; Institute for Plant Breeding, Seed Science and Population Genetics, University of Hohenheim: Stuttgart, Germany, 1997. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.r-project.org (accessed on 10 June 2023).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Squares | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Source | df | LUT | ZEA | αCX | βCX | αC | βC | LUT/ZEA | TOT |
| Location(L) | 1 | 0.1 | 0.4 | 0.2 | 0.4 | 0.0 | 0.4 * | 0.6 | 7.2 |
| Genotype(G) | 87 | 58.3 ** | 31.6 ** | 3.4 ** | 2.0 ** | 0.2 ** | 1.4 ** | 9.2 ** | 103.9 ** |
| L × G | 87 | 6.0 ** | 1.4 | 0.4 + | 0.2 * | 0.03 ** | 0.1 | 0.7 | 10.9 |
| Error | 198 | 3.5 | 1.9 | 0.3 | 0.1 | 0.02 | 0.1 | 1.7 | 27.9 |
| Repeatability | 0.90 | 0.94 | 0.88 | 0.90 | 0.90 | 0.93 | 0.83 | 0.77 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šimić, D.; Galić, V.; Jambrović, A.; Ledenčan, T.; Kljak, K.; Buhiniček, I.; Šarčević, H. Genetic Variability in Carotenoid Contents in a Panel of Genebank Accessions of Temperate Maize from Southeast Europe. Plants 2023, 12, 3453. https://doi.org/10.3390/plants12193453
Šimić D, Galić V, Jambrović A, Ledenčan T, Kljak K, Buhiniček I, Šarčević H. Genetic Variability in Carotenoid Contents in a Panel of Genebank Accessions of Temperate Maize from Southeast Europe. Plants. 2023; 12(19):3453. https://doi.org/10.3390/plants12193453
Chicago/Turabian StyleŠimić, Domagoj, Vlatko Galić, Antun Jambrović, Tatjana Ledenčan, Kristina Kljak, Ivica Buhiniček, and Hrvoje Šarčević. 2023. "Genetic Variability in Carotenoid Contents in a Panel of Genebank Accessions of Temperate Maize from Southeast Europe" Plants 12, no. 19: 3453. https://doi.org/10.3390/plants12193453