1. Introduction
In addition to insects and weeds, pathogenic microorganisms cause structural and/or functional damage to plants, resulting in biotic stress. Plant-pathogen interactions can be viewed as a two-way communication process in which not only is the plant able to recognise a foreign organism and defend itself against it, but the pathogen must also be able to manipulate the plant’s biology to create an optimal environment for its own growth and development while avoiding the plant’s response [
1]. The first line of plant defence can be triggered through the pathogen-associated molecular pattern (PAMP) system, which recognizes the pathogen. This is followed by a series of signal changes that finally impart a defence strategy, also known as the “zigzag” model [
2]. Photosynthesis-related alterations, pathogenesis-related protein synthesis, restructuring of the cytoskeleton, generation of reactive oxygen species, and activation of programmed cell death constitute the fundamental level of plant pathogen recognition modifications [
3]. Due to the plasticity of the plants in response to the pathogen and the establishment of either a compatible or incompatible interaction, the modulation of several classes of primary and secondary metabolites alters. During the initial phases of the infection, plant reactions to oomycetes are identical, albeit less effective and slower in susceptible plants [
4].
Unlike primary metabolites, such as carbohydrates, lipids, and proteins, which are directly involved in plant development and growth, secondary metabolites are multifunctional compounds that are typically involved in the plant’s defence system or they act indirectly, mediating the signals between different parts of the same plant, from plant to plant and between plants and other organisms. Due to their sessile nature, plants synthesize these compounds to repel herbivores, build barriers against pathogen invasion and mitigate oxidative stress [
5]. On the basis of the accumulation and concentration of secondary metabolites in grapevine berries, such as polyphenolic and volatile organic compounds, it has been discovered that it is possible to classify
V. vinifera varieties into genetic-geographic groups [
6,
7].
Terpenes, alkanes, alkenes, alcohols, esters, and acids belong to the class of volatile organic compounds. Terpenes are the largest and most researched class of these compounds. Their building block is a five-carbon isoprene unit. Through the condensation of two or more isoprene units, mono-(C
10), sesqui-(C
15), and diterpene (C
20) precursors are formed [
8]. VOCs perform essential functions in how plants interact with other organisms and how they respond to biotic stress. They constitute about 1% of the secondary metabolites found in plants. Due to their low molecular weight and high vapour pressure, these lipophilic molecules freely diffuse into the environment and pass through biological membranes. Typically, pathogen-induced volatile organic compounds (VOCs) are composed of methyl salicylate (MeSA), mono- and sesquiterpenes, heterocyclic compounds, green leaf volatiles (GLVs), and ketones [
9,
10,
11]. Plants continuously produce GLVs, such as C
6 aldehydes, alcohols, and esters, and do so to a greater extent under stress conditions [
12].
In grapevine, both the defensive role and accumulation of volatile organic compounds, following an attack by
Plasmopara viticola [(Berk. et Curt.) Berl. et de Toni], have been demonstrated. Some volatile organic compounds, such as 2-phenylethanol, 2-ethylfuran, (
E)-2-pentenal,
ß-cyclocitral,
ß-caryophyllene, and
ß-selinene, inhibited
P. viticola infection in leaf tissues. The abundance of the studied VOCs was greater in resistant (BC4, Kober 5BB, SO4, and Solaris) genotypes than in susceptible genotypes (Pinot noir) [
11]. Monoterpenes and sesquiterpenes are found in higher quantities in SO4 and Kober 5BB plants compared to Pinot noir plants after inoculation [
13]. The direct antibacterial action of four selected compounds (farnesene, nerolidol, ocimene, and valencene) as well as the role of leaf terpenes in the resistance mechanism of two resistant cultivars (Mgaloblishvili, a pure
V. vinifera cultivar, and Bianca, an interspecific hybrid) were determined [
8]. Benzaldehyde has been suggested as a putative biomarker of resistance because it acts as a stimulator of salicylic acid (SA)-mediated defence [
14]. It was also discovered in greater amounts in mono-locus (Solaris) and pyramided (F12P60) resistant genotypes. Farnesene was abundantly expressed in three mono-locus resistant genotypes (BC4, Bianca, F12P160). Linalool was substantially more abundant in Bianca, whereas (
E)-nerolidol and neral distinguished the pyramidal genotype F12P60 [
15].
P. viticola, the causal agent of grapevine downy mildew, is an obligate biotrophic oomycete native to North America. Therefore, European grapevine (
V. vinifera) varieties are generally susceptible to this pathogen since it was introduced in this area in the late 19th century [
16]. Biotrophic microorganisms have an important function in the ecosystem because they decompose organic matter [
17]. However, when
P. viticola sporangia develop in large quantities under favourable weather conditions (temperatures between 20 and 25 °C and leaf wetness) [
18], they cause disastrous consequences such as defoliation, reduced and/or complete loss of grape quality and quantity [
16,
19]. In conventional vineyards, downy mildew epidemics cause severe economic losses when fungicides are not administered. Due to the detrimental effects of chemical pesticides on the environment, and human and animal health, as well as the emergence of pathogens resistant to these treatments [
20,
21,
22], the European Union has restricted their usage through the Farm to Fork Strategy [
23]. Today, the focus lies on the development of alternative tools, such as the breeding of resistant cultivars, the development of new active substances, and the search for natural compounds and biocontrol agents that can be applied individually or in combination to eradicate the pathogen or mitigate its effects [
4].
The genetic diversity of traditional grapevine varieties is an inexhaustible source of traits potentially useful in the upcoming challenging environmental conditions. Therefore, they should not be neglected since each variety has a unique characteristic that differentiates it from all other varieties. There is a possibility that these characteristics will become desirable or valuable in the future, despite the fact that they may seem mundane at present. Their preservation and continuous research are of utmost importance for maintaining biodiversity and expanding the scope of future breeding programs [
24]. Quantitative trait loci (QTL) named
Rpv, an acronym for resistance to
P. viticola, are responsible for grapevine’s resistance response. The Table of Loci for Traits in Grapevine Relevant for Breeding and Genetics (
https://www.vivc.de/, the access date: 15 November 2022) lists and describes the 31 QTLs discovered in
Vitis species to date. Most of them have been found in the
Muscadinia subgenus, along with several wild North American and Asian
Vitis species. However, the last three QTLs named
Rpv29,
Rpv30, and
Rpv31, have been identified in the Georgian
V. vinifera variety of Mgaloblishvili [
25] which confirms the importance of preserving and studying rare varieties that are cultivated in limited areas.
The 4th century BCE saw the beginning of viticulture production in Croatia, both in the continental Pannonia region with the arrival of Celts and along the eastern Adriatic coast where Greeks founded cities [
26]. Due to turbulent historical events and the introduction of mildews (
P. viticola and
Erysiphe necator) and phylloxera (
Daktulosphaera vitifoliae) from the North American continent during the 19th century, the number of grapevine varieties and vineyard areas have changed significantly since then. Consequently, today’s Croatian grapevine biodiversity consists of about one hundred varieties [
27]. According to previous research conducted on some of these varieties, their susceptibility to the downy mildew disease under field conditions varies. Moreover, this was confirmed in controlled laboratory conditions using the leaf disc bioassay according to the OIV 452-1 descriptor [Leaf: degree of resistance to
Plasmopara (leaf disc test)]. After measuring chlorophyll fluorescence and multispectral imaging traits, it is possible to distinguish non-infected and infected leaf discs 24 h after inoculation, whereas, on the fourth day upon inoculation, the differences between varieties belonging to various OIV classes are observed [
28]. As far as secondary metabolites are considered, it was found that the constitutive polyphenolic profile of leaves is responsible for genotype differentiation among the OIV classes of resistance to
P. viticola [
29].
This study’s primary objective was to expand the analysis of secondary metabolites in leaves before and during the early stage of inoculation, with a particular emphasis on volatile organic compounds. The research was performed using Solid Phase Microextraction (SPME)–Arrow Gas Chromatography–Mass Spectrometry (GC–MS). Similar methods have been used previously [
8,
11,
14,
15], but this one has proven to be particularly effective for the detection of VOCs [
30]. Defining the differences between the metabolomic profiles of susceptible and resistant grapevines and detecting the resistance–related metabolites could broaden and introduce new concepts in plant protection strategies. Moreover, the detection of VOC emission patterns could be used to screen hybrids with varying resistance levels or to diagnose diseased plants [
31].
2. Results
Eighty-six VOCs were identified through SPME-
Arrow-GC/MS analysis of 17 genotype leaf samples collected at 0, 24, 48, and 94 h after
P. viticola inoculation. There were 19 alcohols, 18 carbonyls (aldehydes and ketones), 17 monoterpenes, 10 sesquiterpenes, 10 esters, 9 acids, and 3 compounds that belong to some other groups of compounds, such as C
13-norisoprenoids and lactones, were detected (
Tables S2 and S3). As far as individual compounds are concerned, the highest absolute peak areas were detected for 2-hexenal, benzyl alcohol, 3-hexen-1-ol, nonanal, and 4-hexen-1-ol acetate that were calculated as average values of all analysed samples. The interactions of all three factors (sampling term, treatment, and OIV resistance class), including T
0, were significant for 17 identified VOCs, the majority of which were carbonyls (5), alcohols (4) and acids (3). The mean values of the interactions are presented in
Table S2.
2.1. Changes throughout the Terms of Sampling
Of the 86 identified VOCs, 49 compounds significantly contributed to the ability to distinguish between the terms of sampling upon inoculation regardless of treatment and OIV class of resistance (
Table S3). A slightly higher number of compounds increased significantly upon inoculation in at least one term (28) compared to the number of compounds that decreased (18). The most numerous increasing compounds were those that belong to monoterpenes (7), alcohols (6), and sesquiterpenes (5).
The alcohols with the highest APA in T1 were: 1-nonanol, 1-butoxy-2-propanol, 1-methoxy-2-propanol, and α,α-dimethylbenzyl alcohol, whereas the alcohols with the lowest APA in T3 were 3,7-dimethyl-1,7-octanediol and benzyl alcohol. The ascending significant change between all three terms of sampling was obtained for 1-hexanol, (E)-2-hexen-1-ol, 2-ethyl-2-hexen-1-ol, and 3-hexen-1-ol. However, the 2-ethyl-1-hexanol APA plunged significantly from T1 to T3. Significant differences between T1 and T3 were observed for 1-octen-3-ol with higher APA in T1, whereas phenylethyl alcohol was more abundant in T3.
In T1, the 2-hexenal APA was significantly the lowest, whereas the (E)-2-nonenal, acetophenone, nonanal, and octanal APAs were the highest. The APA of benzaldehyde decreased significantly throughout the terms following the inoculation, whereas a significant increment was observed for 6-methyl-5-hepten-2-one from T1 to T3. The APA of (E,Z)-2,6-nonadienal and (E,E)-3,5-octadien-2-one distinguished between T1 and T3 with the highest APA being observed in T3 and T1, respectively. The APA of hexanal did not change significantly in the first two terms following the inoculation, although it increased significantly in the third term.
Markedly, the APA of (E)-3-hexenyl butanoate and (Z)-2-hexenyl acetate was lowest in T3. The lowest APA of hexyl acetate, phenylmethyl formate, and methyl salicylate was observed in T1. A higher APA of ethyl octanoate in T1 distinguished it significantly from T3.
In terms of monoterpenes, citronellol, neral, and α-terpineol had significantly lower APAs in T1, whereas p-cymene had a significantly lower APA in T3.
Significantly, the highest APAs in T1 and T2 were observed for geranyl vinyl ether and (E)-linalool oxide, respectively. The APAs of (Z)-linalool oxide and ß-ocimene increased significantly from T1 to T2, whereas geranylacetone increased significantly from T1 to T3.
The α-farnesene sesquiterpene increased significantly from T1 to T3, whereas from T1 to T2 the same is true for humulene and ß-guaiene. Finally, compounds belonging to another group of VOCs, such as (E)-ß-ionone, dihydroactinidiolide, and 5-ethyl-2(5H)-furanone, also increased from T1 to T2.
2.2. Differences between Non-Inoculated and Inoculated Leaves
Compounds that belong to sesquiterpenes and alcohols contributed the most to the discrimination between non-inoculated and inoculated leaves, regardless of the sampling term following the inoculation and the OIV resistance class. More precisely, the infected samples contained significantly elevated levels of humulene, ylangene, and α-farnesene, as well as 1-hexanol, (E)-2-hexen-1-ol, and 2,4-dimethyl-3-pentanol. In contrast, non-infected samples contained significantly higher APAs of 2-ethyl-1-hexanol, 1-nonanol, and 1-butoxy-2-propanol alcohols.
Among the remaining compounds, inoculated leaves measured significantly higher monoterpene geranylacetone, aldehyde octanal, ketone 6-methyl-5-hepten-2-one, and 2-hexonoic acid APAs (
Table S3).
2.3. Differences between the OIV Resistance Classes
As previously explained, each genotype included in this research was assigned to the appropriate OIV class of resistance based on the severity of P. viticola sporulation and the OIV 452-1 descriptor. Although only seven out of eighty-six identified compounds failed to significantly differentiate the OIV resistance classes, no clear separation of genotypes into the OIV resistance classes was obtained. Thus, the contribution of individual compounds is described below.
The lowest APA of 2-hexenoic acid distinguished the completely resistant OIV class 9 from all other OIV classes, whereas the low APA of pentanoic acid distinguished OIV classes 1 and 9 from classes 3, 5, and 7. The lowest APAs of (E)-3-hexenoic acid and decanoic acid were observed in OIV class 3 compared to other OIV classes. Benzoic acid was significantly the most abundant in OIV class 5, whereas heptanoic acid was the most abundant in OIV classes 1 and 5. Decanoic acid distinguished OIV class 3 by its low APA compared to other classes.
The highest APA of 2-ethyl-1-hexanol distinguished the most susceptible OIV class 1 from the highly resistant OIV class 7, and the resistant OIV class 9. Similarly, the APA of 1-nonanol distinguished OIV class 1 and OIV class 9, in which the lowest APA of this alcohol was observed. OIV class 3 had the lowest APA of 1-octanol. Alcohols 2-ethyl-2-hexen-1-ol and (E)-2-hexen-1-ol were found in the highest APA in OIV classes 7 and 5, respectively. OIV classes 3 and 9 were specified by the lowest APA of 3-hexen-1-ol. The highest APAs of 3,7-dimethyl-3-octanol, 2,4-dimethyl-3-pentanol, benzyl alcohol, and phenylethyl alcohol distinguished OIV class 9 from all other classes.
As far as carbonyls are concerned, a higher APA of (E,E)-2,4-heptadienal differentiated OIV classes 1, 3, and 5 (pure V. vinifera varieties) from OIV classes 7 (interspecific hybrid Solaris) and 9 (V. riparia). Similarly, (E,Z)-2,6-nonadienal was observed in significantly higher APA in pure V. vinifera varieties compared to V. riparia. Aldehydes 2-hexenal and (E)-2-nonenal and their higher APAs discriminated OIV classes 1 and 3 from other classes, whereas a higher APA of (E,E)-3,5-octadien-2-one distinguished the susceptible OIV classes 1 and 3 from the partially resistant and resistant OIV classes 5, 7 and 9. OIV class 9 was specified by the highest APA of 4-pentenal compared to all other classes. The APA of 6-methyl-5-hepten-2-one was the highest in OIV class 7 followed by OIV class 9 and then OIV classes 5 and 1. Finally, the lowest APA of this ketone separated OIV class 3 from others. The highest APA of 2,5-dimethyl-benzaldehyde and heptanal distinguished OIV classes 9 and 5 from others, respectively. A higher APA of hexanal separated OIV class 5 from classes 1 and 3. Nonanal and octanal were highest in OIV classes 5 and 7 and lowest in OIV class 3.
The APA of phenylmethyl acetate was the highest in OIV class 9, followed by OIV classes 1 and 5, and the lowest in OIV classes 3 and 7. Significantly the highest APA of ethyl octanoate discriminated OIV class 1 from others. Similarly, 3-hexenyl butanoate distinguished OIV classes 1 and 5 from others. Significant differences were observed for (E)-2-hexenyl benzoate and its higher APA in OIV classes 3 and 5 compared to OIV class 7. In contrast, phenylmethyl formate had a higher APA in OIV class 7 compared to classes 1, 3, and 5. The highest APA of ethyl octanoate distinguished OIV class 1 from others, whereas the lowest APA of (Z)-2-hexenyl acetate was obtained in OIV class 9.
Among monoterpenes, the APA of ß-cyclocitral did not distinguish OIV classes 7 and 9, although it separated OIV class 9 from classes 1, 3, and 5 by being less abundant in OIV class 9. The highest APA of citronellol and nerol differentiated OIV class 7 from others. OIV class 9 was specific by the highest APA of (Z)-linalool oxide, eucalyptol, and p-cymene. A higher APA of geranylacetone differentiated OIV classes 7 and 9 from others. The highest APA of limonene was obtained in the most susceptible OIV class 1. In the same way, menthol distinguished OIV class 1 from classes 7 and 9, whereas linalool separated OIV class 1 from classes 3, 5, and 9. Low APA of α-terpineol distinguished OIV classes 3 and 5 from others. Higher APA of ß-ocimene differentiated OIV classes 1 and 3 from class 5. Geranylacetone was the most abundant in OIV class 9 followed by OIV class 7, whereas it did not distinguish OIV classes 1 and 5.
Higher APAs of (
Z)
-ß-farnesene and
α-farnesene differentiated OIV classes 7 and 9 from others. In comparison, the APAs of humulene and
ß-guaiene were the highest in OIV classes 1 and 5. OIV class 9 was specific due to the lowest APA of 5-ethyl-2(5H)-furanone, whereas a higher APA of dihydroactinidiolide was detected in OIV class 5 compared to OIV classes 1 and 3 (
Table S3).
2.4. Compounds Related to Higher Resistance
Aiming to identify VOCs that could be responsible for higher resistance to P. viticola, highly positive correlations between the infected samples and genotypes that belong to OIV classes 5, 7, and 9 were sought throughout the experiment. At the same time, negative or low correlations were sought for the non-infected (control) samples to verify that the ascending APA of a VOC in the infected sample is associated with a defence mechanism against infection. Moreover, when the same trend was observed for VOCs in varieties that are most susceptible to P. viticola (OIV class 1), these compounds were excluded from consideration.
Throughout the experiment, highly positive correlations were obtained for 2-hexenoic acid in samples from all three OIV class 5 (Malvazija istarska, Ranfol, and Teran) infected varieties, whereas negative or low correlations were obtained for non-infected (control) samples of the same resistance class. The same is true for the following VOCs in Malvazija istarska specifically: 2-ethyl-2-hexen-1-ol,
α,α-dimethylbenzyl alcohol, phenylmethyl acetate, phenylmethyl formate, limonene,
α-terpineol,
ß-myrcene,
ß-ocimene, geranylactone, copaene,
α-farnesene, and
γ-muurolene. The infected samples of Teran exhibited highly positive correlation for (
E)-2-hexen 1-ol, (
E)-2-hexenyl benzoate,
ß-cyclocitral, citronellol, linalool, and
p-cymene, whereas Malvazija istarska and Ranfol exhibited the same for benzeneacetaldehyde, ethyl benzoate, citronellol, menthol, caryophyllene, humulene, and
α-muurolene. However, the same trend was observed for the majority of the above-mentioned compounds in varieties that are the most susceptible to
P. viticola (OIV class 1) (
Table S4). Therefore, they cannot be considered the cause of resistance reactions.
Nevertheless, a few compounds detected in OIV 5 were also detected in OIV 7 or 9 but not in OIV 1, having a high positive correlation in infected samples and a low or negative correlation in non-infected ones. In particular, VOCs detected in OIV 9 were ocimene, (
E)-2-hexen-1-ol, whereas geranylacetone was found in OIV 7.
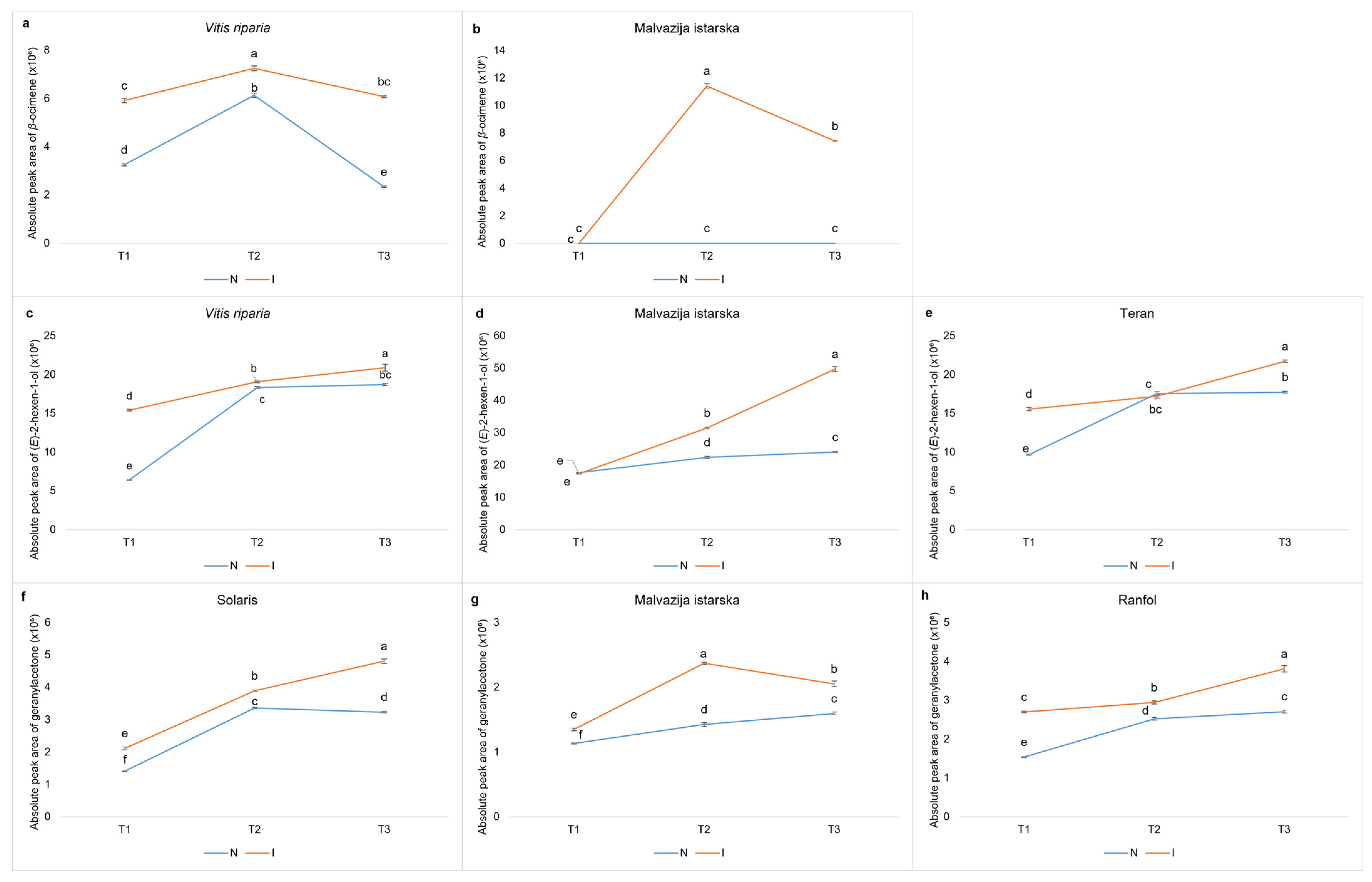
Figure 1 depicts the APA changes of specific VOCs in response to inoculation, which may have contributed to the increased resistance of genotypes belonging to OIV classes 5, 7, or 9.
2.5. Sesquiterpenes and the OIV Resistance Classes
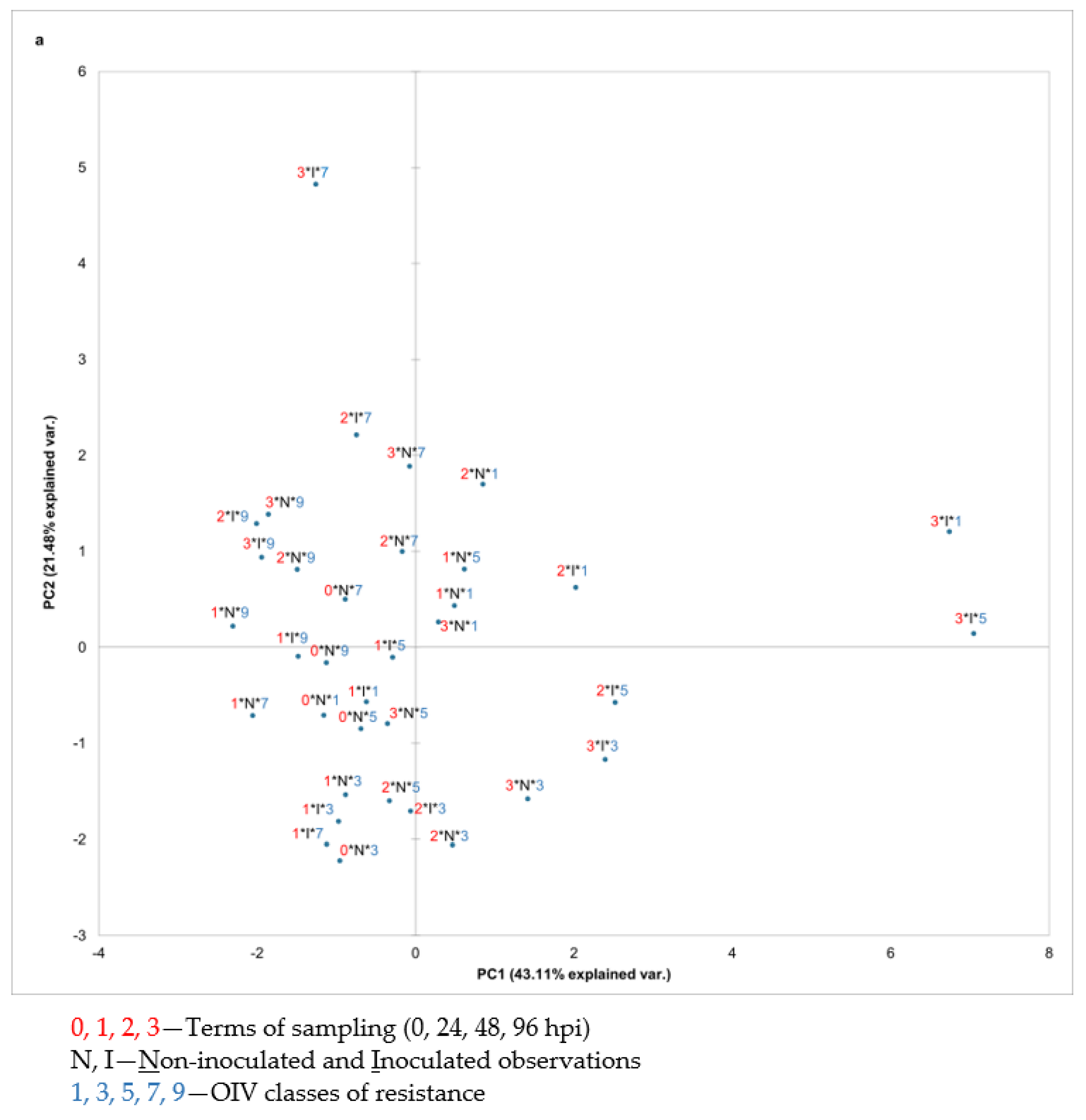
To analyse the total variability of the volatile compounds’ absolute peak area related to the division of the OIV resistance classes (1, 3, 5, 7, and 9), terms of sampling (0, 24, 48, and 96 hpi), and treatments (considering non-inoculated and inoculated samples), a principal component analysis (PCA) was performed using all detected VOCs and using each group of detected volatile compounds separately. Most groups of compounds did not contribute to a distinct separation of the samples by any of the aforementioned factors (data not shown). However, the PCA based on the APA of individual sesquiterpenes contributed the most to distinguishing the OIV classes of resistance. In particular, the PCA scatter plot of the first two components explained 64.59% of the variability (
Figure 2) with the first principal component (PC1) accounting for 43.11% and the second (PC2) for 21.48%. The projection on these two axes distinguished the two highly resistant genotypes (OIV classes 7 and 9) from
V. vinifera genotypes (OIV classes 1, 3, and 5). However, in OIV classes 1, 3, and 5, terms and treatments were not clearly separated (
Figure 2a).
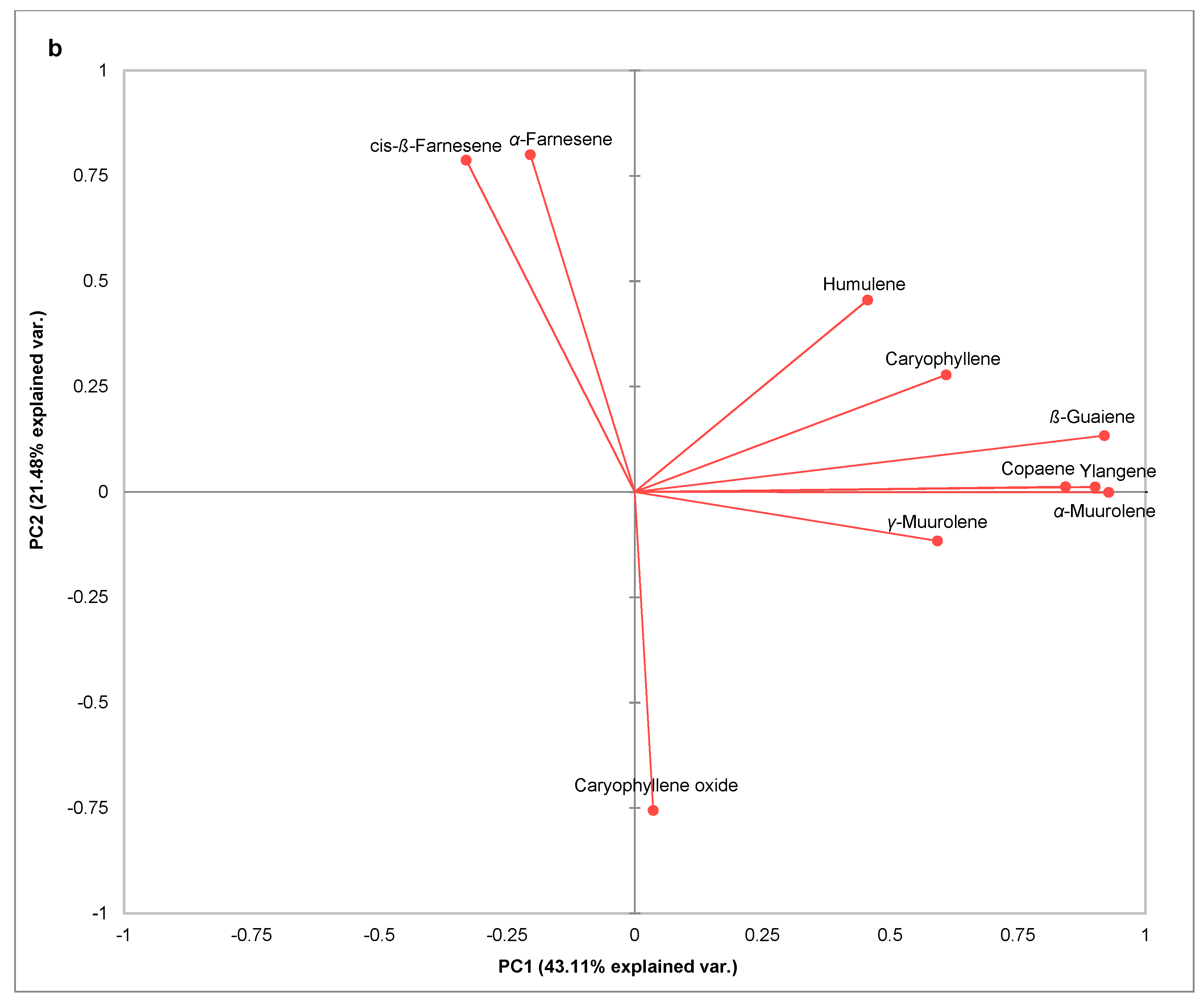
Based on the related vector diagram (
Figure 2b), it is possible to define the sesquiterpenes that contributed to such distribution and grouping of samples that belong to either OIV classes 7 and 9 or OIV classes 1, 3, and 5 in the space defined by the first two principal components. One group containing all the samples belonging to OIV classes 7 and 9 regardless of the treatment and sampling term was separated from the other group due to the higher APA of
α-farnesene and (
Z)-ß-farnesene. Most of these observations are located in the second quadrant and a few of them are in the third quadrant.
3. Discussion
P. viticola maintains its life cycle in living tissue as its tubular mycelium grows intercellularly and obtains nutrients by parasitizing the host cells through haustoria [
32]. Leaves are typically the first to show symptoms of downy mildew, especially young leaves that have not yet developed ontogenic resistance to the disease [
33]. Biosynthesis of VOCs occurs in the leaf mesophyll tissues, specifically in the palisade mesophyll cells [
34]. Consequently, since phytopathogens alter plant VOCs emission, decreasing or increasing the amount of some pre-existing VOCs, and inducing the appearance of newly synthesized VOCs, this research was conducted on young leaves and their volatile organic compounds (VOCs) [
35]. It is known that VOCs can directly inhibit pathogen growth, induce plant resistance mechanisms in neighbouring plants, and mediate associational resistance by adsorption to the cuticle of receiver tissues [
36]. During the early stages of infection, the susceptible cultivar undergoes the following changes: at 24 hpi, the zoospores germinate and the germ tube penetrates the substomatal cavity; at 48 hpi, the
P. viticola hyphae are observed in the intercellular spaces; at 96 hpi, the sporangiophores begin to develop from the stomata [
37]. A novel SPME-Arrow GC/MS technique proved to be efficient for this kind of analysis by processing a large number of samples and providing a whole range of VOCs [
30].
As mentioned previously, phenotypic differences among Croatian native varieties have been investigated, and some specificities corresponding to OIV resistance classes have been observed. Based on the content and composition of polyphenolic compounds, it was found that their constitutive profiles in leaves are responsible for diverse levels of resistance to
P. viticola [
29]. The analysis of 86 volatile organic compounds (VOCs) in the leaves of 14 native Croatian varieties, including Chardonnay, Solaris, and
V. riparia, was performed with the intention of expanding the current findings based on secondary metabolites. To the best of our knowledge, this is the first time such extensive research on this topic has been conducted. Although a clear separation of differently resistant genotypes was not accomplished, as the inoculation time progressed, some specificities were defined among the OIV classes of resistance and the APA of sesquiterpenes. Moreover, a few compounds, such as geranylacetone,
ß-ocimene, and (
E)-2-hexen-1-ol could be responsible for a higher resistance of OIV classes 5, 7, and 9.
Considering the sampling terms upon
P. viticola inoculation, a slightly higher number of VOCs was detected in increased APAs over time, based on the average values of all 17 analysed genotypes. This has already been observed for the resistant genotypes of BC4, Kober 5BB, SO4, and Solaris whose leaf VOCs were analysed at 6 dpi and compared to 0 dpi, whereas the APA of VOCs in Pinot noir leaves decreased [
11]. Although benzaldehyde was not an indicator of
P. viticola inoculation, its APA decreased throughout the experiment which is in accordance with Ricciardi et al., (2021) [
8] whose samples were frozen likewise. At the same time, benzaldehyde content increased in the Bianca cultivar when fresh leaves were analysed [
14], meaning that VOC emission patterns could be related to the sample preparation and the term of sampling. The most numerous increasing compounds were alcohols (e.g., 3-hexen-1-ol and 2-ethyl-2-hexen-1-ol), monoterpenes (e.g., citronellol and neral), and carbonyls (e.g., hexanal and (
E,Z)-2,6-nonadienal). These groups of VOCs increased in the leaves of Bianca and Mgaloblishvili [
8].
Terpenes were found to be the most discriminative among the OIV classes of resistance. The high APA of sesquiterpenes
α-farnesene and (
Z)-ß-farnesene distinguished OIV classes 7 and 9 from the more susceptible OIV classes 1, 3, and 5 regardless of the treatment and the sampling term. Moreover,
α-farnesene was suitable for distinguishing treatments throughout the experiment due to its higher APA in inoculated leaves compared to non-inoculated ones. Similarly, increased emission of sesquiterpenes were detected in in vitro plantlets of the downy mildew–resistant genotypes of SO4 and Kober 5BB, whereas the increment of these VOCs was lower in the Pinot noir susceptible variety [
13]. Likewise, an increased amount of farnesene was detected in the resistant genotypes of Mgaloblishvili and Bianca upon
P. viticola inoculation together with the up-regulation terpene synthase genes, suggesting a pathogen-dependent transcriptional regulation of terpene biosynthesis [
8]. In a study conducted by Ciubotaru et al., (2021), farnesene was expressed in high concentrations in the genotypes with mono-locus resistance, namely BC4 (
Rpv1), Bianca (
Rpv3-1) and F12P160 (
Rpv12), and in the pyramided resistant genotype F12P127 (
Rpv3-1,
Rpv3-3,
Rpv10) [
15]. These findings indicate that VOCs, especially sesquiterpenes, produced by downy mildew-resistant genotypes contribute to grapevine defence against
P. viticola.
Algarra Alarcon et al., (2015) [
13] detected a higher content of monoterpenes in SO4 which contradicts our findings as far as total monoterpenes are concerned since their APA was the highest in the most susceptible OIV class 1. Considering individual monoterpenes, a higher APA of
ß-cyclocitral differentiated pure
V. vinifera varieties (OIV classes 1, 3, and 5) from
V. riparia, whereas linalool was significantly higher in OIV class 1 compared to OIV classes 3, 5, and 9. In contrast, these monoterpenes were detected in higher amounts in the leaves of resistant genotypes (i.e., BC4, Kober 5BB, SO4, and Solaris) compared to the susceptible Pinot noir [
11]. Higher contents of linalool and neral were detected in Bianca and F12P60 suggesting their antimicrobial activity [
15]. In addition to linalool, our research identified its volatile oxides, namely (
Z)- and (
E)-linalool oxides. Defence–related activity could be ascribed to (
Z)-linalool oxide since its highest APA was detected in
V. riparia (OIV 9) distinguishing this genotype from the others evaluated. Neral was detected in Solaris (OIV 7) with a high APA, but it was also found in the most susceptible varieties (OIV 1).
Aldehyde 4-pentenal distinguished
V. riparia from other genotypes by its low APA, whereas Lazazzara et al., (2018) [
11] detected a higher APA of (
E)-2-pentenal in the resistant genotypes of BC4 and Kober 5BB compared to Pinot noir. As far as (
E,E)-2,4-heptadienal and benzeneacetaldehyde are concerned, lower APAs were detected in resistant genotypes in both of these studies. The same authors found benzaldehyde to be more abundant in resistant varieties, whereas, in our study, it was most abundant in OIV classes 5 and 9, although a high APA was also detected in OIV class 1. Nevertheless, Chitarrini et al., (2017) [
14] suggested benzaldehyde as a putative biomarker of resistance to
P. viticola infection since it was detected in higher concentrations in infected samples of the resistant cultivar Bianca at 48 and 96 hpi. Bianca, Solaris, and F12P60, which possess at least one locus of resistance in their genomes, had a higher level of benzaldehyde [
15]. Moreover, it was found that benzaldehyde acts as a promoter of salicylic acid (SA)–mediated defence, as it accumulated early and in high concentration in the plant metabolome with
Rpv12–mediated resistance [
38]. Salicylic acid acts as a phytohormone precursor of the volatile compound methyl salicylate, which is known for activating induced resistance upon attack by biotrophic microorganisms, such as
P. viticola [
12]. Aldehyde 2-hexenal was found in increasing APAs throughout the sampling terms and its high APAs were detected in OIV classes 1 and 5. In our experiment, however, the APA of nonanal decreased. Furthermore, the high APA of nonanal in OIV class 5, the same was detected in Solaris (OIV 7), which could be one of the possible commonalities related to lower susceptibility among these two OIV classes. Previously, a higher content of 2-hexenal was associated with the Solaris cultivar resistance, whereas a higher content of nonanal was detected in the leaves of the F12P127 genotype, and based on that, these VOCs were proposed as biomarkers of resistance [
15].
Phenylethyl alcohol increased throughout the experiment and was detected in the highest APA in OIV class 9 corroborating the findings of Lazazzara et al., (2018) [
11]. Throughout the experiment, the alcohols 1-hexanol and (
E)-2-hexen-1-ol were found in ascending APAs, while their higher APAs in inoculated leaves distinguished them from non-inoculated leaves. OIV 5 varieties have the highest APA of (
E)-2-hexen-1-ol, which suggests that it may play a role in the defence mechanism of
V. vinifera varieties that are less susceptible to
P. viticola. Both alcohols, together with 2-ethyl-1-hexanol and 1-octen-3-ol, have been identified as potential biomarkers of resistance in Bianca, Solaris, and F12P60 genotypes due to their higher concentration upon inoculation compared to Pinot noir, which is susceptible [
15]. In our study, the APA of 2-ethyl-1-hexanol decreased with time and was higher in non-inoculated leaves and in varieties that are the most susceptible to
P. viticola (OIV 1), whereas 1-octen-3-ol was not significant for any of these parameters.
Among esters, (
Z)-3-hexenyl benzoate was proposed as a potential biomarker of resistance in previous research due to its higher up-regulation upon
P. viticola inoculation in the resistant genotype of F12P60 compared to Pinot noir [
15]. Similarly, we identified a relatively high APA of (
E)-2-hexenyl benzoate in
V. riparia. Methyl salicylate is synthesized from salicylic acid by salicylate methyl transferase and is widespread in plants as a volatile odorous compound associated with mint-like and green pepper aromas [
39]. It was demonstrated that a higher concentration of methyl salicylate in stems, grapes, and consequently in red and white wines was related to vine diseases (downy mildew, grape black rot, Esca) suggesting a host plant-induced defence mechanism against fungal infection. Thus, methyl salicylate can serve as a volatile indicator of the vineyard’s infection status [
40]. In our study, it was detected in the highest abundance in the resistant genotype of
V. riparia as well as the highly resistant Solaris cultivar confirming its potent antifungal activity in these genotypes. Salicylic acid and methyl salicylate induce systemic acquired resistance and hypersensitive response (cell death) as a reaction to a pathogen attack [
41]. Although methyl salicylate did not help distinguish the non-inoculated and inoculated samples, its higher APA was observed 48 h upon inoculation corroborating that the synthesis of this VOC is induced by a pathogen attack. Acting as a volatile form of a defence phytohormone, methyl salicylate systemically induces defence responses in plant parts and organs that are distant from the initial infection site. Moreover, these airborne signals can be perceived by uninfected neighbouring plants and induce a resistance reaction in them as well [
12]. Previously, methyl salicylate has been proposed as a biomarker of downy mildew infection [
42] and as a potential biomarker of resistance to
P. viticola since it was detected in higher concentrations in Bianca compared to Pinot noir [
15].
In previous studies that included field trials and in vitro leaf disc bioassay, three Croatian native grapevine varieties, namely Malvazija istarska, Ranfol, and Teran (OIV class 5), proved to be more resistant to
P. viticola compared to other evaluated
V. vinifera varieties [
28]. Moreover, their constitutive polyphenolic profile of leaves, i.e., higher content of flavonol glycosides mostly, distinguish them from the more susceptible varieties [
29]. Aiming to define VOCs that could be responsible for the defence mechanism of these three Croatian grapevine varieties, VOCs from OIV class 5 were compared with VOCs from OIV classes 7 and 9. Therefore, a few compounds with a higher APA upon inoculation in inoculated leaves compared to non-inoculated ones in OIV classes 5, 7, and 9 were observed, namely geranylacetone,
ß-ocimene, and (
E)-2-hexen-1-ol.
Geranylacetone belongs to the class of organic compounds known as acyclic monoterpenes. It is a component of essential oils in various plants including
Nelumbo nucifera whose leaf extract has strong antioxidant properties [
43]. Up to now, geranylacetone was not recognized as a biomarker of grapevine resistance to
P. viticola, although it was detected in higher concentrations in the leaves of the resistant pyramided genotype F12P60 compared to Pinot noir [
15]. Ricciardi et al., (2021) [
8] did not find significant changes in the quantity of this compound upon inoculation in either Bianca or Mgaloblishvili. In this study, geranylacetone was found to have an ascending APA throughout the experiment and a significantly higher APA in each sampling term in the inoculated leaves of the native varieties Malvazija istarska and Ranfol, as well as the highly resistant Solaris cultivar, compared to non-inoculated leaves, indicating that geranylacetone may be involved in the defence mechanism of these cultivars.
Another possible indicator of resistance is
ß-ocimene, a volatile organic compound that belongs to the class of monoterpenes. During T
2 (48 hpi), its induced accumulation was observed in the inoculated leaves of the native variety Malvazija istarska, whereas it was never detected in the control leaves.
ß-ocimene was detected in both inoculated and non-inoculated leaves of the
V. riparia resistant genotype, although its APAs were higher in inoculated leaves in each term following inoculation. Previously, terpenes were often recognized as compounds associated with the defence mechanism against downy mildew [
8,
11,
13]. Specifically, allo-ocimene was found to activate defence genes and induce resistance against
Botrytis cinerea in
Arabidopsis thaliana [
44]. Functional properties of terpenes, such as farnesene, nerolidol, valencene, and ocimene were examined and found efficient in counteracting
P. viticola. Not only were they synthesized in higher amounts in the resistant variety of Mgaloblishvili, but their antisporulant activity was also proved in ad hoc experimental inoculations in which disease severity and sporangia concentration were inhibited. Among these terpenes, ocimene was found to be the most effective [
8].
As previously mentioned, the volatile alcohol (
E)-2-hexen-1-ol was proposed as a biomarker of resistance in the study conducted by Ciubotaru et al., (2021) [
15]. In our research, the same compound was detected in inoculated leaves of Teran and
V. riparia in ascending APAs upon
P. viticola inoculation. (
Z)-3-hexenol, an alcohol with a similar structure, was detected in higher concentrations in the resistant
V. labrusca and
V. riparia genotypes compared to the susceptible
V. vinifera varieties [
45]. (
Z)-3-hexenol is known to induce up-regulation of defence genes during the
Botrytis cinerea infection of
Arabidopsis thaliana, acting in the same way as allo-ocimene [
44].

 ,
, 



{kind=link}
{kind=link}
{kind=link}




