
Vascular Plant Species Inventory of Mexico’s Revillagigedo National Park: Awareness of Alien Invaders as a Sine Qua Non Prerequisite for Island Conservation
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
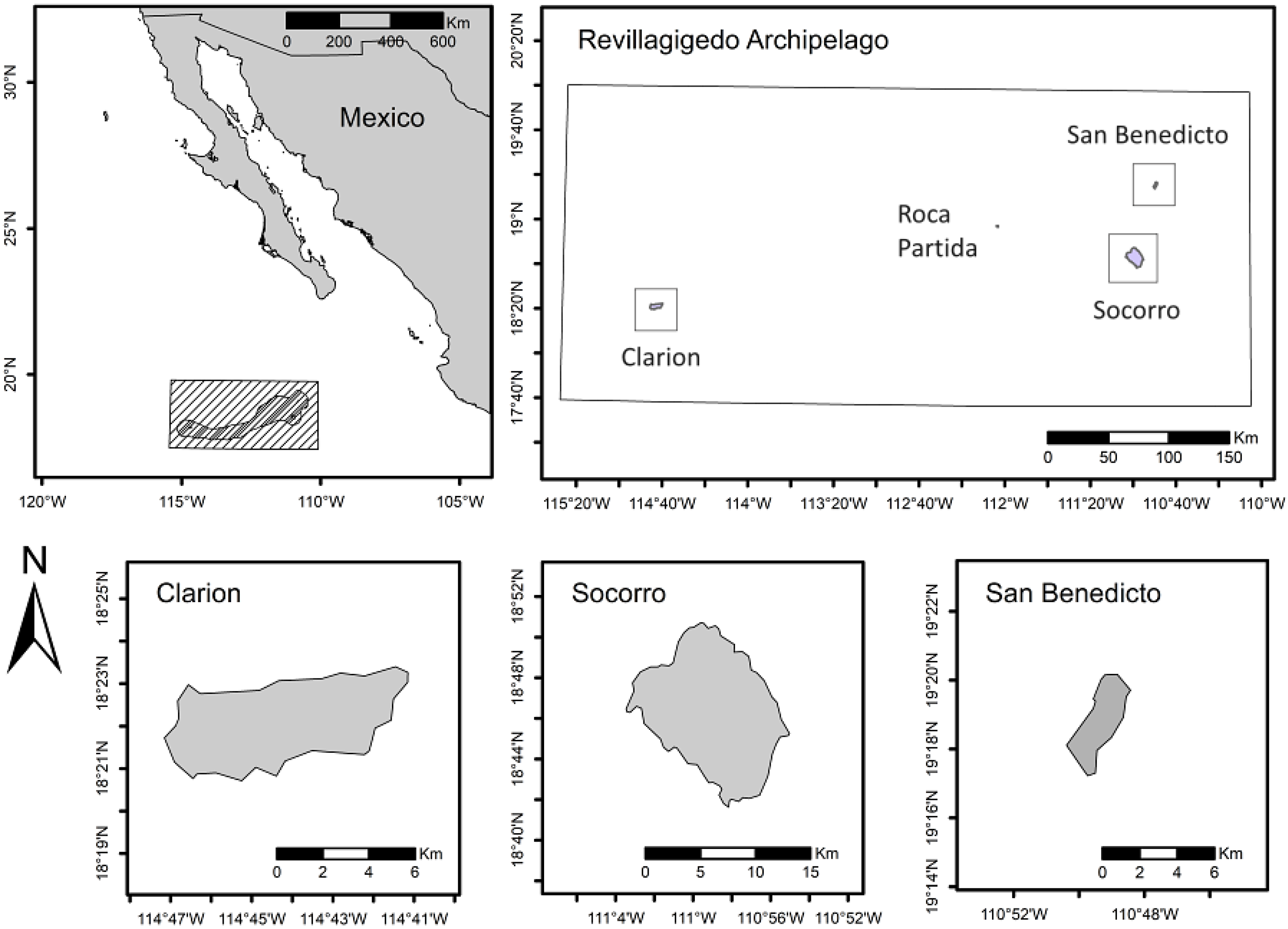
2.1. Study Area
2.2. Inventory of Plant Species Present on the Revillagigedo Archipelago
2.3. Species Traits: Origin, Life History, and Date of Introduction
2.4. Invasiveness Risk Assessment for Alien Plant Species
2.5. Comparison of the Status of Native and Alien Plant Species of Different Insular Regions
3. Results
3.1. Inventory Overview
3.2. Origin and Life History
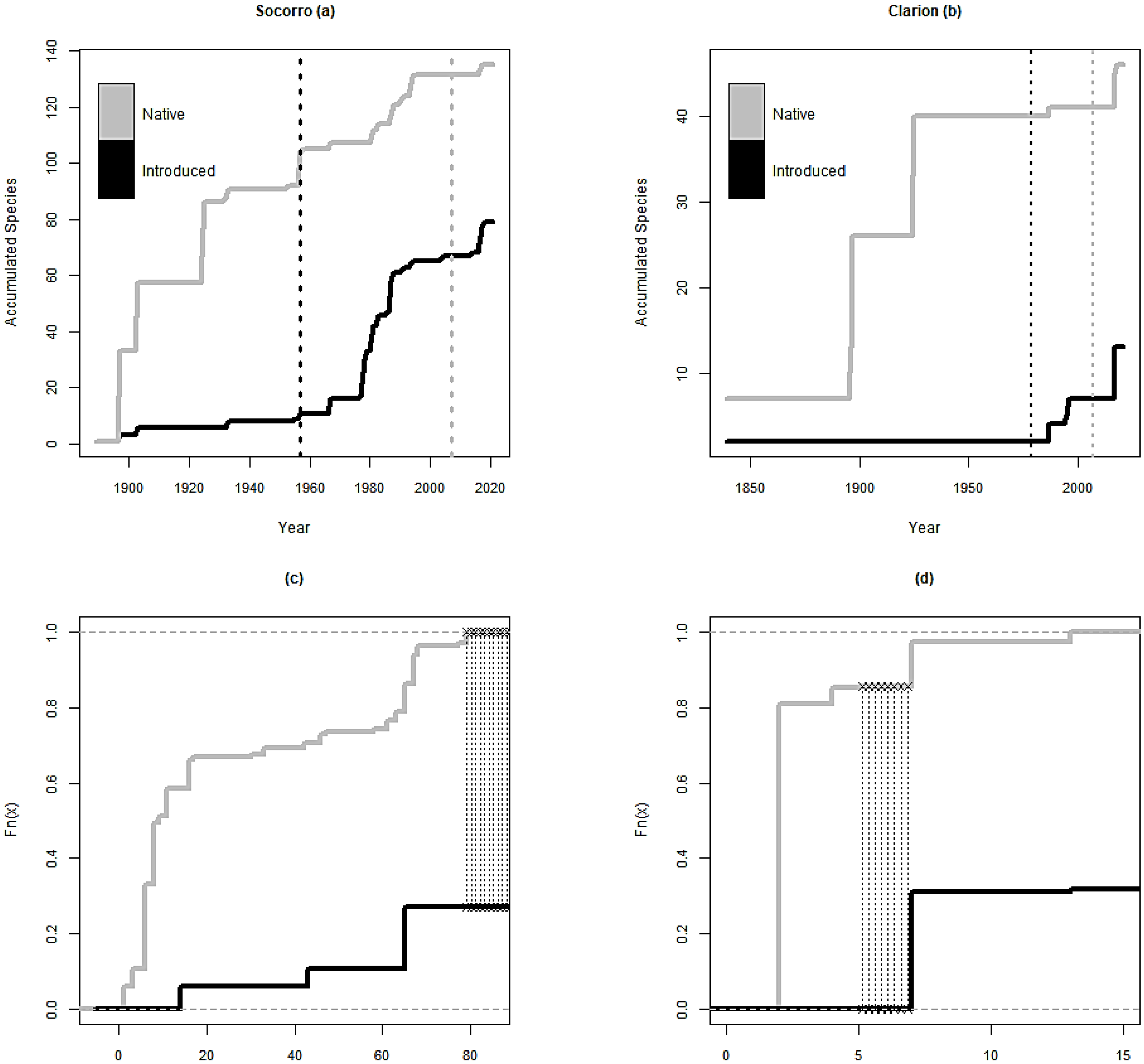
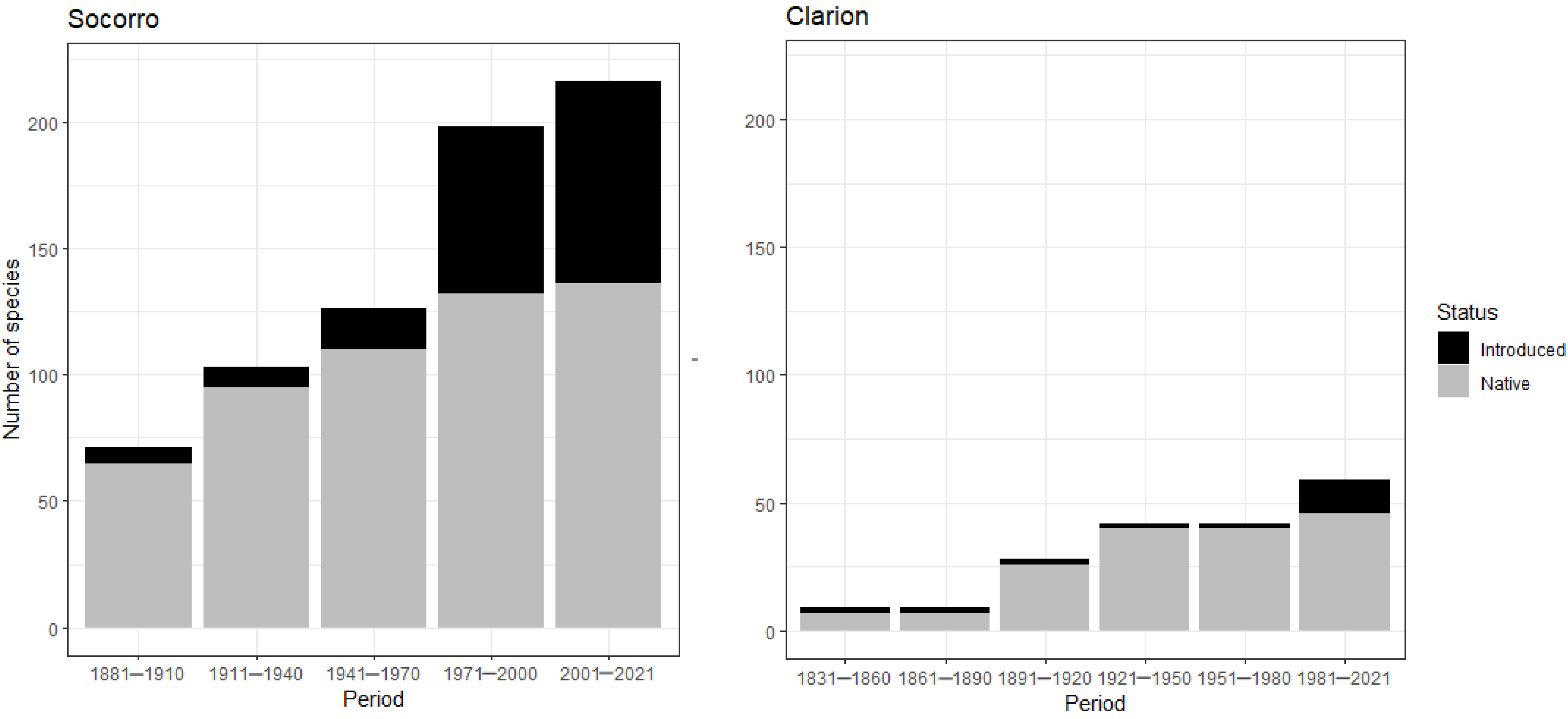
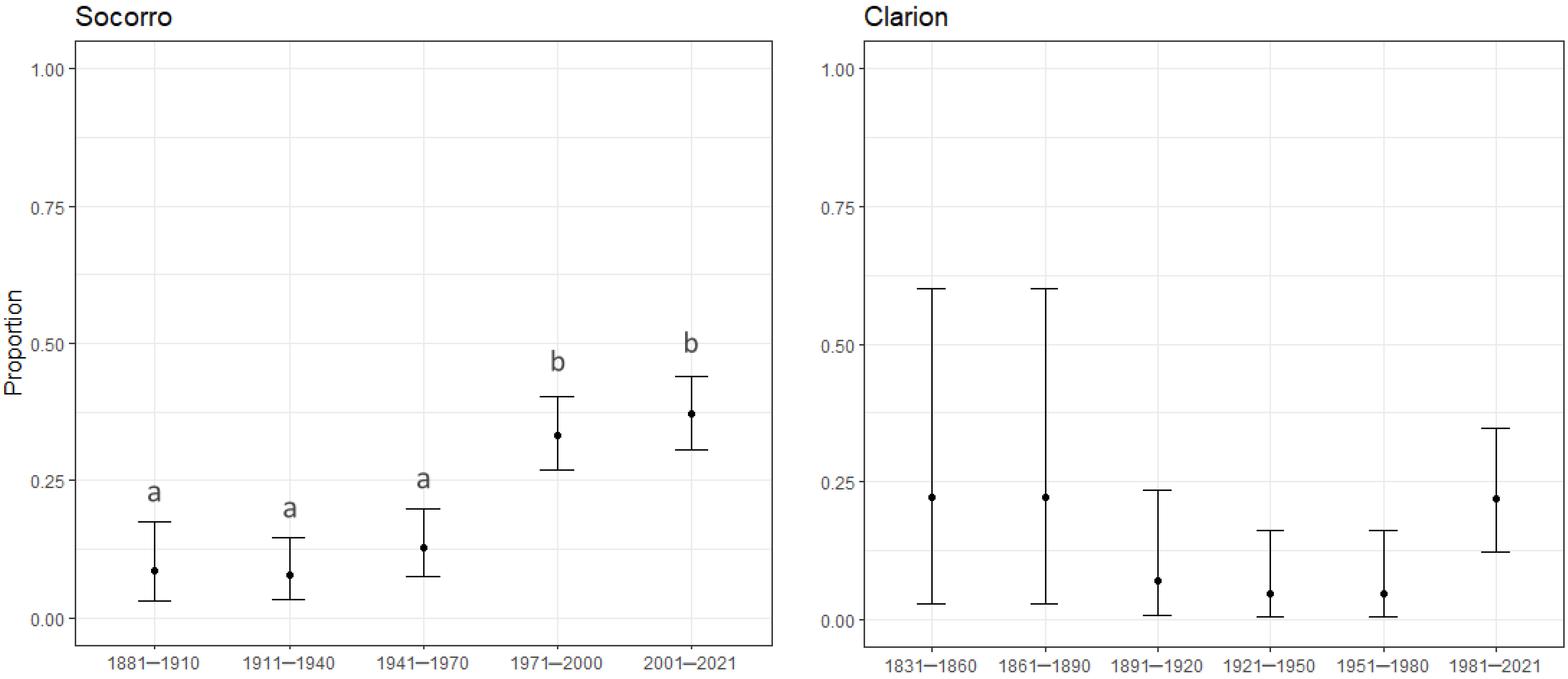
3.3. Year of Introduction
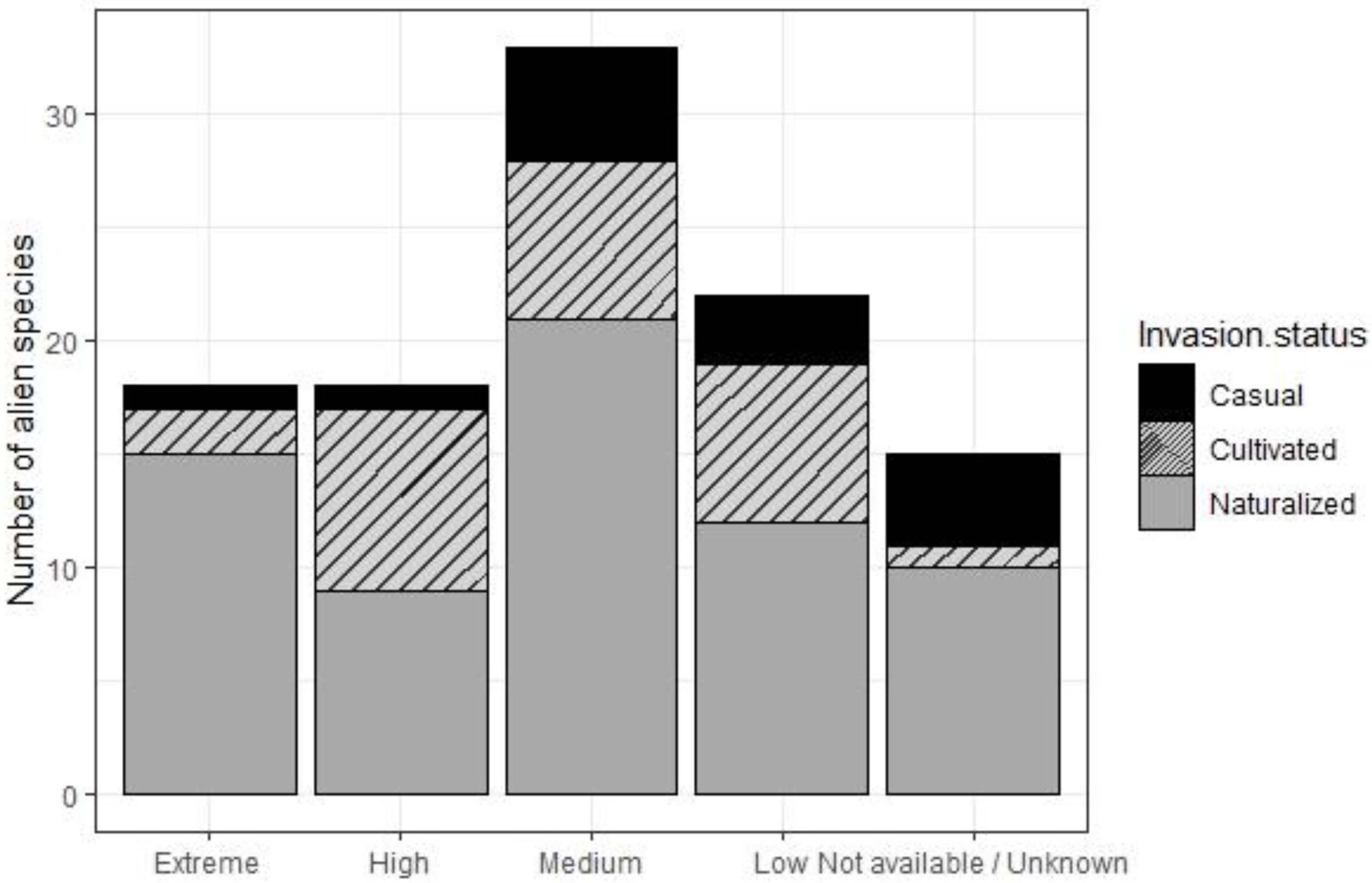
3.4. Invasiveness Risk Assessment for Alien Plant Species
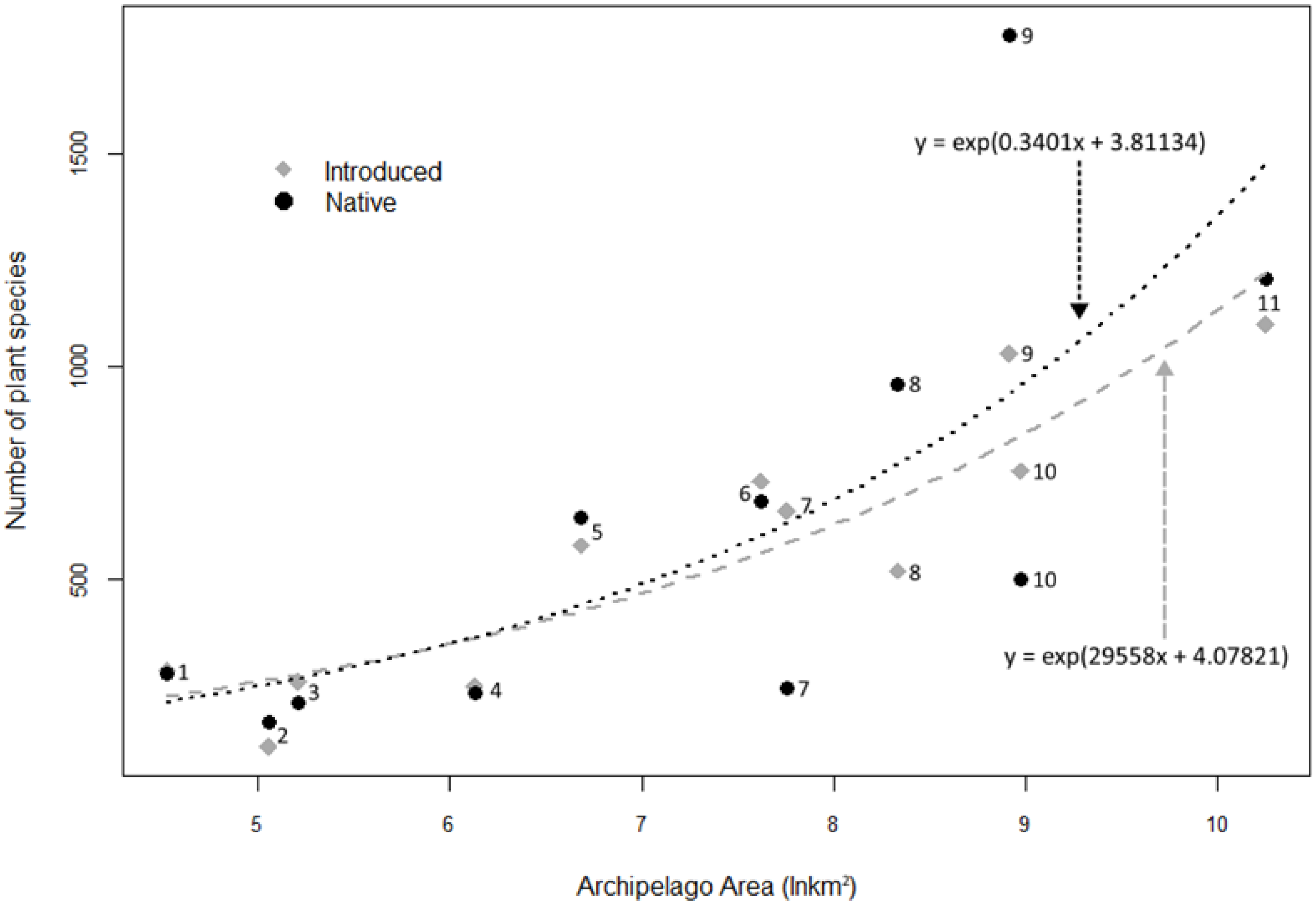
3.5. Comparison of the Status of Native and Alien Plant Species of Different Insular Regions
4. Discussion
4.1. Inventory Overview
4.2. Origin and Life History
4.3. Year of Introduction
4.4. Invasiveness Risk Assessment for Alien Plant Species
4.5. Comparison of the Status of Native and Alien Plant Species of Different Insular Regions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levin, G.; Moran, R. The Vascular Flora of Isla Socorro, México. Memoir; San Diego Society of Natural History: San Diego, CA, USA, 1989; pp. 1–71. [Google Scholar]
- Brattstrom, B.H. Biogeography of the Islas Revillagigedo, Mexico. J. Biogeogr. 1990, 17, 177–183. [Google Scholar] [CrossRef]
- Martínez-Gómez, J.E. La avifauna del archipiélago de Revillagigedo. Amenazas, estrategias e instrumentos para la conservación. In Conservación de Aves: Experiencias en México; Gómez Silva, H., Oliveras de Ita, A., Eds.; CIPAMEX: Mexico City, Mexico, 2002. [Google Scholar]
- SEDESOL. Decreto por el que se Declara Como Área Natural Protegida con el Carácter de Reserva de la Biosfera, la Región Conocida Como Archipiélago de Revillagigedo, Integrada por Cuatro Áreas: Isla San Benedicto, Isla Clarión o Santa Rosa, Isla Socorro o Santo Tomás e Isla Roca Partida. Diario Oficial de la Federación. 1994. Available online: https://faolex.fao.org/docs/pdf/mex179968.pdf (accessed on 16 November 2022).
- UNESCO. Decision: 40 COM 8B.14. In Proceedings of the World Heritage Committee: 40th Session. WHC/16/40.COM/19, Istanbul, Turkey, 10–20 July 2016; pp. 196–199. [Google Scholar]
- SEMARNAT. Decreto por el que se Declara Como Área Natural Protegida, con el Carácter de Parque Nacional, la Región Conocida Como Revillagigedo, Localizada en el Pacífico Mexicano. Diario Oficial de la Federación. 2017. Available online: https://www.dof.gob.mx/nota_detalle_popup.php?codigo=5505736 (accessed on 16 November 2022).
- March Mifsut, I.J.; González Moreno, J. Revillagigedo: Refugio de Gigantes; MAPorrúa: Mexico City, Mexico, 2018; p. 282. [Google Scholar]
- Jehl, J.R., Jr.; Parkes, K.C. “Replacements” of Landbird Species on Socorro Island, Mexico. Auk 1983, 100, 551–559. [Google Scholar] [CrossRef]
- Walter, H.S.; Levin, G.A. Feral sheep on Socorro Island: Facilitators of alien plant colonization and ecosystem decay. Divers. Distrib. 2008, 14, 422–431. [Google Scholar] [CrossRef]
- Jehl, J.; Parkes, K. The status of the avifauna of the Revillagigedo Islands, Mexico. Wilson Bull. 1982, 94, 1–19. [Google Scholar]
- Ortíz-Alcaraz, A.; Maya-Delgado, Y.; Cortés-Calva, P.; Aguirre-Muñoz, A.; Rojas-Mayoral, E.; Cordoba-Matson, M.V.; Ortega-Rubio, A. Recovery of Vegetation Cover and Soil after the Removal of Sheep in Socorro Island, Mexico. Forests 2016, 7, 91. [Google Scholar] [CrossRef]
- Mendez-Guardado, P. Analysis of the Environmental Impact Caused by Introduced Animals in the Clarion Island, Archipelago of Revillagigedo, Colima, Mexico. In Global Change and Protected Areas; Visconti, G., Beniston, M., Iannorelli, E.D., Barba, D., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 323–329. [Google Scholar]
- Brattstrom, B.H. Food Webs and Feeding Habits on the Revillagigedo Islands, Mexico. Pac. Sci. 2015, 69, 181–195. [Google Scholar] [CrossRef]
- Wanless, R.M.; Aguirre-Muñoz, A.; Angel, A.; Jacobsen, J.K.; Keitt, B.S.; McCann, J. Birds of Clarion Island, Revillagigedo Archipelago, Mexico. Wilson J. Ornithol. 2009, 121, 745–751. [Google Scholar] [CrossRef]
- Mack, R. Motivations and consequences of the human dispersal of plants. In The Great Reshuffling: Human Dimensions in Invasive Alien Species, IUCN; McNeely, J.A., Ed.; World Conservation Union: Gland, Switzerland, 2001; pp. 23–34. [Google Scholar]
- SEMARNAT. Aviso por el que se Informa al Público en General que la Secretaría de Medio Ambiente y Recursos Naturales ha Concluido la Elaboración del Programa de Manejo del Área Natural Protegida con el Carácter de Reserva de la Biosfera Archipiélago de Revillagigedo, Ubicada en el Océano Pacífico, Establecida Mediante Decreto Presidencial Publicado el 6 de Junio de 1994. Diario Oficial de la Federación. 2007. Available online: https://www.dof.gob.mx/nota_detalle_popup.php?codigo=5008088 (accessed on 16 November 2022).
- SEMARNAT. Acuerdo por el que se da a Conocer el Resumen del Programa de Manejo del Área Natural Protegida con la Categoría de Parque Nacional Revillagigedo. Diario Oficial de la Federación. 2018. Available online: https://dof.gob.mx/nota_detalle_popup.php?codigo=5545530 (accessed on 16 November 2022).
- McLellan, M.E. Expedition of the California Academy of Sciences to the Revillagigedo Islands. Science 1925, 62, 171–173. [Google Scholar] [CrossRef]
- Johnston, I. The flora of the Revillagigedo Islands. In Proceedings of the California Academy of Sciences; 4th Series; The Academy: San Francisco, CA, USA, 1931; Volume 20, pp. 9–104. [Google Scholar]
- Rebman, J.; Vanderplank, S.E.; Ezcurra, E. Vascular plants of the Revillagigedo Islands expedition in February 2017. In Report on the Terrestrial Expedition to the Revillagigedo Archipelago, 2020; Vanderplank, S.E., Wall, M.A., Ezcurra, E., Eds.; San Diego Society of Natural History: San Diego, CA, USA, 2020; Volume 48, pp. 53–76. [Google Scholar]
- Mickel, J.; Smith, A.R. The Pteridophytes of Mexico; The New York Botanical Garden Press: New York, NY, USA, 2004; Volume 88. [Google Scholar]
- Dávila, P.; Cabrera, L.I.; Lira, R. A new species of Sorghastrum (Poaceae) from Isla Socorro, Colima, Mexico. Brittonia 1998, 50, 301–308. [Google Scholar] [CrossRef]
- Benavides, E.; Kuethe, J.R.; Ortiz-Alcaraz, A.; De la Luz, J.L. Oenothera resicum (Onagraceae), a new species and the first record of the family from the Revillagigedo Archipelago, Mexico. Phytotaxa 2019, 416, 59–66. [Google Scholar] [CrossRef]
- Kiehn, M. Invasive alien species and islands. In The Biology of Island Floras; Bramwell, D., Caujapé-Castells, J., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 365–384. [Google Scholar]
- Magee, J.F.; McMullen, C.K.; Reaser, J.K.; Spitzer, E.; Struve, S.; Tufts, C.; Tye, A.; Woodruff, G. Green invaders of the Galapagos Islands. Science 2001, 294, 1279. [Google Scholar] [CrossRef]
- Rejmánek, M. Invasive plants: Approaches and predictions. Austral. Ecol. 2000, 25, 497–506. [Google Scholar] [CrossRef]
- Richardson, D.; Pyšek, P. Plant invasions: Merging the concepts of species invasiveness and community invasibility. Prog. Phys. Geogr. Earth Environ. 2006, 30, 409–431. [Google Scholar] [CrossRef]
- Flores-Palacios, A.; Martínez-Gómez, J.E.; Curry, R.L. La Vegetación de Isla Socorro, Archipiélago de Revillagigedo, México. Bol. Soc. Bot. Mex. 2009, 84, 13–23. [Google Scholar] [CrossRef]
- INEGI. Catálogo del Territorio Insular Mexicano; Instituto Nacional de Estadística y Geografía: Aguascalientes, Mexico, 2015; p. 243.
- Valero, A.; Schipper, J.; Allnutt, T. Islands of Socorro, Clarion, San Benedicto, and Roca Partida in the Pacific Ocean off the Coast of Mexico. Available online: https://www.worldwildlife.org/ecoregions/nt0216 (accessed on 2 September 2022).
- Martínez-Gómez, J. Informe Final: Programa de Manejo de Áreas Naturales Protegidas (PROMANP): Coordinación del Programa Nacional para la Conservación de la Paloma de Socorro. Convenio de concertación CONANP/PROMANP/MB/037/2017. 2017. Available upon request.
- Rodriguez-Estrella, R.; Leon de la Luz, J.L.; Breceda, A.; Castellanos, A.; Cancino, J.; Llinas, J. Status, density and habitat relationships of the endemic terrestrial birds of Socorro Island, Revillagigedo Islands, Mexico. Biol. Conserv. 1996, 76, 195–202. [Google Scholar] [CrossRef]
- Mulcahy, D.G.; Martínez-Gómez, J.E.; Aguirre-León, G.; Cervantes-Pasqualli, J.A.; Zug, G.R. Rediscovery of an Endemic Vertebrate from the Remote Islas Revillagigedo in the Eastern Pacific Ocean: The Clarión Nightsnake Lost and Found. PLoS ONE 2014, 9, e97682. [Google Scholar] [CrossRef]
- González-Sánchez, V.H.; Johnson, J.D.; González-Solís, D.; Fucsko, L.A.; Wilson, L.D. A review of the introduced herpetofauna of Mexico and Central America, with comments on the effects of invasive species and biosecurity methodology. Zookeys 2021, 1022, 79–154. [Google Scholar] [CrossRef]
- Brattstrom, B.H. Bárcena Volcano, 1952: A 60-year report on the repopulation of San Benedicto Island, Mexico, with a review of the ecological impacts of disastrous events. Pac. Conserv. Biol. 2015, 21, 38–59. [Google Scholar] [CrossRef]
- SEMARNAT-CONANP. Programa de Conservación y Manejo de la Reserva de la Biosfera Archipiélago de Revillagigedo, México. 2004. Available online: https://www.conanp.gob.mx/que_hacemos/pdf/programas_manejo/revillagigedo.pdf (accessed on 22 December 2021).
- SEMARNAT-CONANP. Programa de Manejo del Parque Nacional Archipiélago de Revillagigedo, México. 2018. Available online: https://simec.conanp.gob.mx/pdf_libro_pm/82_libro_pm.pdf (accessed on 22 December 2021).
- CONABIO. 2021. Available online: https://enciclovida.mx/ (accessed on 28 September 2023).
- The Plant List. Version 1.1. 2013. Available online: http://www.theplantlist.org/ (accessed on 10 August 2021).
- World Flora Online (WFO). 2021. Available online: https://www.worldfloraonline.org/ (accessed on 28 September 2023).
- International Plant Names Index (IPNI). 2020. Available online: https://www.ipni.org/ (accessed on 28 September 2023).
- Plants of the World Online (POWO). 2019. Available online: https://powo.science.kew.org/ (accessed on 28 September 2023).
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M.; Rejmánek, M.; Webster, G.L.; Williamson, M.; Kirschner, J. Alien plants in checklists and floras: Towards better communication between taxonomists and ecologists. TAXON 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Dorjee, J.; Barry, S.; Buckmaster, A.J.; Downey, P.O. Weeds in the land of Gross National Happiness: Knowing what to manage by creating a baseline alien plant inventory for Bhutan. Biol. Invasions 2020, 22, 2899–2914. [Google Scholar] [CrossRef]
- Ansong, M.; Pergl, J.; Essl, F.; Hejda, M.; van Kleunen, M.; Randall, R.; Pyšek, P. Naturalized and invasive alien flora of Ghana. Biol. Invasions 2019, 21, 669–683. [Google Scholar] [CrossRef]
- Brummitt, R.K.; Pando, F.; Hollis, S.; Brummitt, N. World Geographical Scheme for Recording Plant Distributions, 2nd ed.; International Working Group on Taxonomic Databases for Plant Sciences (TDWG): Geneva, Switzerland, 2001. [Google Scholar]
- Omer, A.; Kordofani, M.; Gibreel, H.H.; Pyšek, P.; van Kleunen, M. The alien flora of Sudan and South Sudan: Taxonomic and biogeographical composition. Biol. Invasions 2021, 23, 2033–2045. [Google Scholar] [CrossRef]
- Kueffer, C.; Daehler, C.C.; Torres-Santana, C.W.; Lavergne, C.; Meyer, J.-Y.; Otto, R.; Silva, L. A global comparison of plant invasions on oceanic islands. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 145–161. [Google Scholar] [CrossRef]
- Tokeshi, M. Species Abundance Patterns and Community Structure. In Advances in Ecological Research; Begon, M., Fitter, A.H., Eds.; Academic Press: Cambridge, MA, USA, 1993; Volume 24, pp. 111–186. [Google Scholar]
- Stürmer, S.L.; Stürmer, R.; Pasqualini, D. Taxonomic diversity and community structure of arbuscular mycorrhizal fungi (Phylum Glomeromycota) in three maritime sand dunes in Santa Catarina state, south Brazil. Fungal Ecol. 2013, 6, 27–36. [Google Scholar] [CrossRef]
- Wuertz, D.; Setz, T.; Chalabi, Y.; Maechler, M.; Setz, M. fBasics: Rmetrics-Markets and Basic Statistics, R Package Version 3042.89.1. 2020. Available online: https://packages.debian.org/bullseye/r-cran-fbasics (accessed on 30 April 2022).
- Crawley, M. The R Book, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- R_Core_Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Pheloung, P. Determining the Weed Potential of New Plant Introductions to Australia. 1995. Available online: https://nre.tas.gov.au/Documents/WA-Weed-Risk-Assessment-REPORT.pdf (accessed on 15 May 2022).
- Pheloung, P.C.; Williams, P.A.; Halloy, S.R. A weed risk assessment model for use as a biosecurity tool evaluating plant introductions. J. Environ. Manag. 1999, 57, 239–251. [Google Scholar] [CrossRef]
- Daehler, C.C.; Denslow, J.S.; Ansari, S.; Kuo, H.-C. A Risk-Assessment System for Screening Out Invasive Pest Plants from Hawaii and Other Pacific Islands. Conserv. Biol. 2004, 18, 360–368. [Google Scholar] [CrossRef]
- Randall, R.P. A Global Compendium of Weeds, 3rd ed.; RP Randall: Perth, WA, Australia, 2017. [Google Scholar]
- Randall, R.P. Can a plant’s cultural status and weed history provide a generalised weed risk score? In Proceedings of the 20th Australasian Weeds Conference, Perth, WA, Australia, 11–15 September 2016; pp. 5–12. [Google Scholar]
- Martínez-Gómez, J.E.; Matías-Ferrer, N. Island Biogeography of Avian Haemosporidians in the Neotropical Region. In Avian Malaria and Related Parasites in the Tropics: Ecology, Evolution and Systematics; Santiago-Alarcon, D., Marzal, A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 281–329. [Google Scholar]
- Beierkuhnlein, C.; Walentowitz, A.; Welss, W. FloCan—A Revised Checklist for the Flora of the Canary Islands. Diversity 2021, 13, 480. [Google Scholar] [CrossRef]
- Guézou, A.; Trueman, M.; Buddenhagen, C.E.; Chamorro, S.; Guerrero, A.M.; Pozo, P.; Atkinson, R. An Extensive Alien Plant Inventory from the Inhabited Areas of Galapagos. PLoS ONE 2010, 5, e10276. [Google Scholar] [CrossRef]
- Castro, S.A.; Daehler, C.C.; Silva, L.; Torres-Santana, C.W.; Reyes-Betancort, J.A.; Atkinson, R.; Jaramillo, P.; Guezou, A.; Jaksic, F.M. Floristic homogenization as a teleconnected trend in oceanic islands. Divers. Distrib. 2010, 16, 902–910. [Google Scholar] [CrossRef]
- Gustafson, R.J.; Herbst, D.R.; Rundel, P.W. Hawaiian Plant Life: Vegetation and Flora; University of Hawaii Press: Honolulu, HI, USA, 2014. [Google Scholar]
- Blackburn, T.M.; Delean, S.; Pyšek, P.; Cassey, P. On the island biogeography of aliens: A global analysis of the richness of plant and bird species on oceanic islands. Glob. Ecol. Biogeogr. 2016, 25, 859–868. [Google Scholar] [CrossRef]
- Cuevas, J.G.; Marticorena, A.; Cavieres, L.A. New additions to the introduced flora of the Juan Fernández Islands: Origin, distribution, life history traits, and potential of invasion. Rev. Chil. Hist. Nat. 2004, 77, 523–538. [Google Scholar] [CrossRef]
- Meyer, J.-Y.; Lavergne, C. Beautés fatales: Acanthaceae species as invasive alien plants on tropical Indo-Pacific Islands. Divers. Distrib. 2004, 10, 333–347. [Google Scholar] [CrossRef]
- Meyer, J.-Y. Threat of invasive alien plants to native flora and forest vegetation of eastern Polynesia (1). Pac. Sci. 2004, 58, 357–375. [Google Scholar] [CrossRef]
- Lavergne, C.; Rameau, J.-C.; Figier, J. The Invasive Woody Weed Ligustrum robustum subsp. walkeri Threatens Native Forests on La Réunion. Biol. Invasions 1999, 1, 377–392. [Google Scholar] [CrossRef]
- González-Gallegos, J.G.; Bedolla-García, B.Y.; Cornejo-Tenorio, G.; Fernández-Alonso, J.L.; Fragoso-Martínez, I.; García-Peña, M.d.R.; Harley, R.M.; Klitgaard, B.; Martínez-Gordillo, M.J.; Wood, J.R.I.; et al. Richness and Distribution of Salvia Subg. Calosphace (Lamiaceae). Int. J. Plant Sci. 2020, 181, 831–856. [Google Scholar] [CrossRef]
- Wood, J.R.I.; Muñoz-Rodríguez, P.; Williams, B.R.M.; Scotland, R.W. A foundation monograph of Ipomoea (Convolvulaceae) in the New World. PhytoKeys 2020, 143, 1–823. [Google Scholar] [CrossRef] [PubMed]
- Tye, A.; Soria, M.; Gardener, M. A strategy for Galapagos weeds. In Turning the Tide: The Eradication of Invasive Species; Veitch, C.R., Clout, M.N., Eds.; Invasive Species Specialist Group, IUCN: Gland, Switzerland; Cambridge, UK, 2002; pp. 336–341. [Google Scholar]
- Vorsino, A.E.; Fortini, L.B.; Amidon, F.A.; Miller, S.E.; Jacobi, J.D.; Price, J.P.; Gon, S.O., III; Koob, G.A. Modeling Hawaiian Ecosystem Degradation due to Invasive Plants under Current and Future Climates. PLoS ONE 2014, 9, e95427. [Google Scholar] [CrossRef] [PubMed]
- Caujapé-Castells, J.; Tye, A.; Crawford, D.J.; Santos-Guerra, A.; Sakai, A.; Beaver, K.; Lobin, W.; Vincent Florens, F.B.; Moura, M.; Jardim, R.; et al. Conservation of oceanic island floras: Present and future global challenges. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 107–129. [Google Scholar] [CrossRef]
- Rentería, J.; Atkinson, R.; Buddenhage, C. Estrategias para la Erradicación de 21 Especies de Plantas Potencialmente Invasoras en Galápagos; Fundación Charles Darwin, Departamento de Botánica, Programa de Especies Invasoras en Galápagos: Puerto Ayora, Ecuador, 2007; p. 108. [Google Scholar]
- Mueller-Dombois, D.; Fosberg, F.R. Vegetation of the Tropical Pacific Islands; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1998; p. 733. [Google Scholar]
- Pyšek, P.; Pergl, J.; Essl, F.; Lenzner, B.; Dawson, W.; Kreft, H.; Weigelt, P.; Winter, M.; Kartesz, J.; Nishino, M.; et al. Naturalized alien flora of the world. Preslia 2017, 89, 203–274. [Google Scholar] [CrossRef]
- Dunlop, E.A.; Wilson, J.C.; Mackey, A.P. The potential geographic distribution of the invasive weed Senna obtusifolia in Australia. Weed Res. 2006, 46, 404–413. [Google Scholar] [CrossRef]
- David, A.S.; Menges, E.S. Microhabitat preference constrains invasive spread of non-native natal grass (Melinis repens). Biol. Invasions 2011, 13, 2309–2322. [Google Scholar] [CrossRef]
- Stokes, C.A.; MacDonald, G.E.; Adams, C.R.; Langeland, K.A.; Miller, D.L. Seed Biology and Ecology of Natalgrass (Melinis repens). Weed Sci. 2011, 59, 527–532. [Google Scholar] [CrossRef]
- García-Cancel, J.G.; Swinnerton, K.; Albarracín, R.; Feliciano, A.; Figuerola-Hernández, C.E.; Silander, S. Update on the Desecheo Island Flora. Caribb. J. Sci. 2019, 49, 295–304. [Google Scholar] [CrossRef]
- Daehler, C.C.; Goergen, E.M. Experimental Restoration of an Indigenous Hawaiian Grassland after Invasion by Buffel Grass (Cenchrus ciliaris). Restor. Ecol. 2005, 13, 380–389. [Google Scholar] [CrossRef]
- Jackson, J. Is there a relationship between herbaceous species richness and buffel grass (Cenchrus ciliaris)? Austral Ecol. 2005, 30, 505–517. [Google Scholar] [CrossRef]
- Barbosa, F.G. The future of invasive African grasses in South America under climate change. Ecol. Inform. 2016, 36, 114–117. [Google Scholar] [CrossRef]
- Visser, V.; Wilson, J.R.U.; Fish, L.; Brown, C.; Cook, G.D.; Richardson, D.M. Much more give than take: South Africa as a major donor but infrequent recipient of invasive non-native grasses. Glob. Ecol. Biogeogr. 2016, 25, 679–692. [Google Scholar] [CrossRef]
- Martínez-Gómez, J.E. Proyecto para remover a las especies de árboles exóticos introducidos en Isla Socorro. Oficio de Autorización SGPA/DGVS/0792/16. Solicitud Complementaria: Oficio de Autorización SGPA/DGVS/1152/19. Dirección General de Vida Silvestre, Secretaría de Medio Ambiente y Recursos Naturales. 2015. Available upon request.
- Tye, A. Invasive plant problems and requirements for weed risk assessment in the Galápagos islands. In Weed Risk Assessment; Groves, R.H., Panetta, F.D., Virtue, J.G., Eds.; CSIRO Publishing: Melbourne, VIC, Australia, 2001; Volume 153, pp. 154–175. [Google Scholar]
- Sax, D.F.; Gaines, S.D. Species invasions and extinction: The future of native biodiversity on islands. Proc. Natl. Acad. Sci. USA 2008, 105, 11490–11497. [Google Scholar] [CrossRef]
- Denslow, J.S.; Space, J.C.; Thomas, P.A. Invasive Exotic Plants in the Tropical Pacific Islands: Patterns of Diversity. Biotropica 2009, 41, 162–170. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Scientific Name | Island | Endemic | Introduced | Probably Not Present |
|---|---|---|---|---|---|
| Acanthaceae | Elytraria imbricata (Vahl.) Pers. | S | |||
| Aizoaceae | Sesuvium portulacastrum (L.) L. | C | |||
| Amaranthaceae | Amaranthus palmeri S. Watson | S | * | ||
| Amaranthaceae | Gomphrena nitida Rothr. | S | * | ||
| Amaranthaceae | Gomphrena serrata L. | S | * | ||
| Amaranthaceae | Iresine diffusa Humb. & Bonpl. ex Willd. | C | |||
| Anacardiaceae | Mangifera indica L. | S | * | ||
| Annonaceae | Annona muricata L. | S | * | ||
| Apiaceae | Daucus montanus Humb. & Bonpl. ex Schult. | S | |||
| Apocynaceae | Cascabela thevetia (L.) Lippold | S | * | ||
| Apocynaceae | Metastelma californicum Benth. | S | |||
| Apocynaceae | Metastelma cuneatum Brandegee | S | |||
| Apocynaceae | Metastelma minutiflorum Wiggins | S | |||
| Aquifoliaceae | Ilex socorroensis Brandegee | S | * | ||
| Araliaceae | Oreopanax xalapensis (Kunth) Decne. & Planch. | S | |||
| Arecaceae | Cocos nucifera L. | S, C | * | ||
| Arecaceae | Phoenix dactylifera L. | S | * | ||
| Aristolochiaceae | Aristolochia islandica Pfeiffer | SB | * | * | |
| Aristolochiaceae | Aristolochia socorroensis Pfeifer | S | * | ||
| Aristolochiaceae | Aristolochia sp. | C | * | ||
| Asparagaceae | Agave americana L. | S | * | ||
| Asphodelaceae | Aloe vera (L.) Burm.f. | S | * | ||
| Aspleniaceae | Asplenium formosum Willd. | S | |||
| Aspleniaceae | Asplenium sessilifolium Desv. | S | |||
| Asteraceae | Ageratina pacifica (B.L. Rob. ex B.L. Rob.) R.M. King & H. Rob. | S | |||
| Asteraceae | Ageratina pichinchensis (Kunth) R.M. King & H. Rob. | S | |||
| Asteraceae | Ambrosia confertiflora DC. | S | * | ||
| Asteraceae | Bahiopsis chenopodina (E. Greene) E. E. Schill. & Panero | S | |||
| Asteraceae | Bidens socorrensis Moran & G. A. Levin | S | * | ||
| Asteraceae | Brickellia peninsularis Brandegee | S | * | ||
| Asteraceae | Brickellia peninsularis Brandegee var. amphithalassa I. M. Johnst. | C | * | ||
| Asteraceae | Coreocarpus insularis (Brandegee) E. B. Smith | S | * | ||
| Asteraceae | Eclipta prostrata (L.) L. | S | |||
| Asteraceae | Erigeron crenatus Eastwood ex I. M. Johnst. | SB | * | * | |
| Asteraceae | Erigeron socorrensis Brandegee | S | * | ||
| Asteraceae | Gamochaeta sphacelata (Kunth) Cabrera | S | |||
| Asteraceae | Gnaphalium attenuatum DC. | S | |||
| Asteraceae | Laennecia confusa (Cronq.) G. L. Nesom | S | |||
| Asteraceae | Perityle socorrosensis Rose | S, C, SB | * | ||
| Asteraceae | Blumea viscosa (Miller) D’Arcy | S | * | ||
| Asteraceae | Sonchus asper (L.) Hill | S | * | ||
| Asteraceae | Sonchus tenerrimus L. | S | * | ||
| Asteraceae | Tridax procumbens L. | S | * | ||
| Asteraceae | Eremosis littoralis Gleason | S | * | ||
| Boraginaceae | Cordia curassavica (Jacq.) Roemer & Schultes | S | |||
| Boraginaceae | Cordia cylindrostachya (Ruiz & Pav.) Roem. & Schult. | S | * | ||
| Boraginaceae | Heliotropium curassavicum L. | S | |||
| Boraginaceae | Heliotropium curassavicum L. var. oculatum (Heller) Thorne | C | |||
| Boraginaceae | Heliotropium procumbens Miller | S | * | ||
| Boraginaceae | Tournefortia capitata M. Martens & Galeotti | S | |||
| Boraginaceae | Tournefortia hartwegiana Steud. | S | |||
| Brassicaceae | Lepidium lasiocarpum Torr. & A. Gray var. latifolium C. L. Hitchc. | C | |||
| Brassicaceae | Lepidium virginicum L. ssp. pubescens (Greene) C. L. Hitchc. | S | |||
| Burseraceae | Bursera epinnata (Rose) Engl. | S | |||
| Cactaceae | Opuntia sp. 1 | C | |||
| Cactaceae | Opuntia sp. 2 | S | |||
| Campanulaceae | Calcaratolobelia cordifolia (Hook. & Arn.) Wilbur | S | |||
| Campanulaceae | Triodanis perfoliata subsp. biflora (Ruiz & Pav.) Lammers | S | |||
| Caricaceae | Carica papaya L. | S | * | * | |
| Caryophyllaceae | Arenaria lanuginosa (Michx.) Rohrb. | S | |||
| Chenopodiaceae | Dysphania ambrosioides (L.) Mosyakin & Clemants | S | * | ||
| Cleomaceae | Arivela viscosa (L.) Raf. | S, C | * | ||
| Combretaceae | Conocarpus erectus L. | S | |||
| Combretaceae | Terminalia catappa L. | S | * | ||
| Commelinaceae | Commelina erecta L. | C | |||
| Convolvulaceae | Cressa truxillensis Kunth | S, C | |||
| Convolvulaceae | Ipomoea carnea subsp. fistulosa (Mart. ex Choisy) D.F. Austin | S | * | ||
| Convolvulaceae | Ipomoea halierca I. M. Johnst. | C | * | ||
| Convolvulaceae | Ipomoea imperati (Vahl) Griseb. | S | |||
| Convolvulaceae | Ipomoea indica (Burm. f.) Merrill | C | |||
| Convolvulaceae | Ipomoea pes-caprae (L.) R. Br. | S, C, SB | |||
| Convolvulaceae | Ipomoea purpurea (L.) Roth | S | * | ||
| Convolvulaceae | Ipomoea triloba L. | S | |||
| Convolvulaceae | Distimake quinquefolius (L.) A.R Simões & Staples | S | * | ||
| Cucurbitaceae | Citrullus lanatus (Thunb.) Matsum. & Nakai. | S | * | * | |
| Cucurbitaceae | Lagenaria siceraria (Mol.) Standl. | S | * | * | |
| Cucurbitaceae | Luffa cylindrica (L.) M. Roem. | S, C | * | * | |
| Cycadaceae | Cycas revoluta Thunb. | S | * | ||
| Cyperaceae | Bolboschoenus maritimus (L.) Palla ssp. paludosus (A. Nelson) T. Koyama | C | |||
| Cyperaceae | Bulbostylis nesiotica (I. M. Johnst.) Fern. | S, C, SB | * | SB * | |
| Cyperaceae | Cyperus duripes I. M. Johnst. | S, C, SB | * | ||
| Cyperaceae | Cyperus ligularis Hemsl. | S | |||
| Cyperaceae | Cyperus rotundus L. | S | * | ||
| Cyperaceae | Cyperus sordidus J. Presl & C. Presl | S | |||
| Cyperaceae | Eleocharis mutata (L.) Roem. & Schult. | C | |||
| Dennstaedtiaceae | Pteridium caudatum (L.) Maxon | S | |||
| Dryopteridaceae | Ctenitis equestris (Kunze) Ching | S | |||
| Dryopteridaceae | Dryopteris knoblochii A. R. Smith | S | |||
| Dryopteridaceae | Polystichum hartwegii (Klotzsch) Hieron | S | |||
| Euphorbiaceae | Acalypha umbrosa Brandegee | S | * | ||
| Euphorbiaceae | Croton masonii I. M. Johnst. | S | * | ||
| Euphorbiaceae | Euphorbia anthonyi (Brandegee) G. A. Levin var. anthonyi | S | * | ||
| Euphorbiaceae | Euphorbia anthonyi (T.S. Brandegee) G. Levin | SB | * | ||
| Euphorbiaceae | Euphorbia anthonyi Brandegee var. clarionensis (Brandegee) I. M. Johnst. | C | * | ||
| Euphorbiaceae | Euphorbia californica Benth. | C | |||
| Euphorbiaceae | Euphorbia heterophylla L. | S | * | ||
| Euphorbiaceae | Euphorbia hirta L. var. hirta | S | * | ||
| Euphorbiaceae | Euphorbia hyssopifolia L. | S | * | ||
| Euphorbiaceae | Euphorbia incerta Brandegee | S | |||
| Euphorbiaceae | Euphorbia thymifolia L. | S, C | * | ||
| Euphorbiaceae | Hippomane mancinella L. | S | |||
| Euphorbiaceae | Ricinus communis L. | S | * | ||
| Fabaceae | Guilandina bonduc L. | S, C | |||
| Fabaceae | Canavalia rosea (Sw.) DC. | S, C | |||
| Fabaceae | Cassia fistula L. | S | * | ||
| Fabaceae | Crotalaria incana L. | S | * | ||
| Fabaceae | Delonix regia (Hook.) Raf. | S | * | ||
| Fabaceae | Desmanthus bicornutus S. Watson | S | * | ||
| Fabaceae | Desmodium procumbens (Mill.) Hitchc. | S | * | ||
| Fabaceae | Desmodium scorpiurus (Sw.) Desf. | S | * | ||
| Fabaceae | Galactia striata (Jacq.) Urban | C | |||
| Fabaceae | Indigofera suffruticosa Miller | S | * | ||
| Fabaceae | Leucaena leucocephala (Lam.) de Wit | S | * | ||
| Fabaceae | Macroptilium atropurpureum (DC.) Urban | S, C | * | ||
| Fabaceae | Neptunia plena (L.) Benth. | S, C | * | ||
| Fabaceae | Phaseolus lunatus L. | S | |||
| Fabaceae | Pithecellobium dulce (Roxb.) Benth. | S | * | ||
| Fabaceae | Prosopis juliflora (Sw.) DC. | S | * | ||
| Fabaceae | Rhynchosia minima (L.) DC. | S, C | * | ||
| Fabaceae | Senna alata (L.) Roxb. | S | * | ||
| Fabaceae | Senna obtusifolia (L.) Irwin & Barneby | S | * | ||
| Fabaceae | Sophora tomentosa L. | C | |||
| Fabaceae | Tamarindus indica L. | S | * | ||
| Fabaceae | Vachellia campechiana (Mill.) Seigler & Ebinger | S | * | ||
| Fabaceae | Vachellia farnesiana (L.) Wight & Arn. var. farnesiana | S | * | ||
| Fabaceae | Zapoteca formosa subsp. rosei (Wiggins) H.M. Hern. | S, C | * | ||
| Gentianaceae | Centaurium capense C. R. Broome | S | |||
| Gentianaceae | Zeltnera wigginsii (Broome) Mansion | S | |||
| Goodeniaceae | Scaevola plumieri (L.) Vahl | S | |||
| Hypericaceae | Hypericum eastwoodianum I. M. Johnst. | S | * | ||
| Lamiaceae | Lepechinia hastata (A. Gray) Epling ssp. socorrensis Moran | S | * | ||
| Lamiaceae | Hyptis pectinata (L.) Poit. | S | * | ||
| Lamiaceae | Salvia pseudomisella Moran & G. A. Levin | S | * | ||
| Lamiaceae | Salvia misella Kunth | S | * | ||
| Lamiaceae | Teucrium townsendii Vasey & Rose var. affine (Brandegee) Moran | S | * | ||
| Lamiaceae | Teucrium townsendii Vasey & Rose var. townsendii | C | * | ||
| Lamiaceae | Teucrium affine Vasey & Rose var. dentosum I.M. Johnst. | SB | * | * | |
| Lauraceae | Persea americana Mill. | S | * | ||
| Lycopodiaceae | Huperzia dichotoma (Jacq.) Trevis | S | |||
| Malvaceae | Abutilon californicum Benth. | S | |||
| Malvaceae | Corchorus aestuans L. | S | * | ||
| Malvaceae | Corchorus orinocensis Kunth | S | * | ||
| Malvaceae | Gossypium hirsutum L. | S | * | ||
| Malvaceae | Hibiscus diversifolius Jacq. | S | * | ||
| Malvaceae | Hibiscus furcellatus Desr. | S | |||
| Malvaceae | Talipariti tiliaceum var. pernambucense (Arruda) Fryxell | S | * | ||
| Malvaceae | Malvastrum americanum (L.) Torrey | S | * | ||
| Malvaceae | Malvastrum coromandelianum (L.) Garcke | S | * | ||
| Malvaceae | Malvella leprosa (Ortega) Krapov. | C | |||
| Malvaceae | Melochia pyramidata L. | S, C | S * | ||
| Malvaceae | Pavonia hastata Cav. | S | |||
| Malvaceae | Sida acuta Burm.f. | S | * | ||
| Malvaceae | Sida barclayi Baker f. | S | * | ||
| Malvaceae | Sida ciliaris L. | S | * | ||
| Malvaceae | Sida nesogena I. M. Johnst. | S | * | ||
| Malvaceae | Sida rhombifolia L. | S | * | ||
| Malvaceae | Sida salviifolia C. Presl | S | * | ||
| Malvaceae | Triumfetta socorrensis Brandegee | S | * | ||
| Malvaceae | Waltheria indica L. | S, C | |||
| Meliaceae | Cedrela odorata L. | S | * | ||
| Meliaceae | Melia azedarach L. | S | * | ||
| Meliaceae | Swietenia humilis Zucc. | S | * | ||
| Meliaceae | Swietenia macrophylla King | S | * | ||
| Molluginaceae | Glinus radiatus (Ruíz López & Pavón) Rohrb. | S | * | ||
| Molluginaceae | Mollugo verticillata L. | S | * | ||
| Moraceae | Ficus benjamina L. | S | * | ||
| Moraceae | Ficus cotinifolia Kunth | S | |||
| Myrtaceae | Psidium guajava L. | S | * | ||
| Myrtaceae | Psidium socorrense I. M. Johnst. | S | * | ||
| Nyctaginaceae | Boerhavia coccinea Miller | S, C | |||
| Nyctaginaceae | Boerhavia erecta L. | S | * | ||
| Nyctaginaceae | Boerhavia scandens L. | C | |||
| Nyctaginaceae | Bougainvillea glabra Choisy | S | * | ||
| Oleaceae | Forestiera rhamnifolia Griseb. | S | |||
| Onagraceae | Oenothera resicum Benavides, Kuethe, Ortiz-Alcaráz & León de la Luz | C | * | ||
| Ophioglossaceae | Botrychium socorrense W.H. Wagner | S | * | ||
| Ophioglossaceae | Ophioglossum reticulatum L. | S | |||
| Orchidaceae | Acianthera unguicallosa (Ames & C. Schweinf.) Solano | S | * | ||
| Orchidaceae | Dendrophylax porrectus (Rchb. f.) Carlsward & Whitten | S | |||
| Orchidaceae | Epidendrum nitens Rchb. f. | S | |||
| Orchidaceae | Epidendrum rigidum Jacq. | S | |||
| Orchidaceae | Guarianthe aurantiaca (Bateman ex Lindl.) Dressler & W.E. Higgins | S | |||
| Orchidaceae | Sarcoglottis schaffneri (Rchb. f.) Ames | S | |||
| Orobanchaceae | Castilleja bryantii Brandegee var. socorrensis (Moran) J. M. Egger | S | * | ||
| Papaveraceae | Argemone ochroleuca Sweet | S | * | ||
| Passifloraceae | Passiflora suberosa L. | S | |||
| Piperaceae | Peperomia socorronis Trel. | S | * | ||
| Piperaceae | Peperomia tetraphylla (G. Forster) Hook. & Arn. | S | |||
| Plantaginaceae | Nuttallanthus texanus (Scheele) D. A. Sutton | S | |||
| Plantaginaceae | Scoparia dulcis L. | S | * | ||
| Poaceae | Aristida adscensionis L. | S | |||
| Poaceae | Aristida tenuifolia Hitchc. | C | * | ||
| Poaceae | Aristida vaginata A. Hitchc. | S | * | ||
| Poaceae | Bothriochloa pertusa (L.) A. Camus | S | * | ||
| Poaceae | Cenchrus ciliaris L. | S, C | * | ||
| Poaceae | Cenchrus echinatus L. | S, C | * | ||
| Poaceae | Cenchrus myosuroides Kunth | S, SB | SB * | ||
| Poaceae | Chloris barbata Swartz | S, C | * | ||
| Poaceae | Cynodon dactylon (L.) Pers. | S | * | ||
| Poaceae | Dactyloctenium aegyptium (L.) Willd. | S, C | * | ||
| Poaceae | Digitaria bicornis (Lam.) Roem. & Schult. | S | * | ||
| Poaceae | Echinochloa colona (L.) Link | S | * | ||
| Poaceae | Eragrostis tenella (L.) P. Beauv. ex Roem. & Schult. | S | * | ||
| Poaceae | Eragrostis ciliaris (L.) Link. | S | * | ||
| Poaceae | Eragrostis prolifera Vasey | SB | |||
| Poaceae | Eragrostis pectinacea (Michx.) Nees | S | * | ||
| Poaceae | Eriochloa acuminata (Presl). Kunth. | C | |||
| Poaceae | Heteropogon contortus (L.) P. Beauv. ex Roem. & Schult. | S, C | |||
| Poaceae | Jouvea pilosa (Presl) Scribn. | S | |||
| Poaceae | Leptochloa panicea (Retz.) Ohwi | S | * | ||
| Poaceae | Melinis repens (Willdenow) Zizka | S, C | * | ||
| Poaceae | Muhlenbergia solisii (G. A. Levin) P. M. Peterson | S | * | ||
| Poaceae | Oplismenus hirtellus (L.) Beauv. | S | |||
| Poaceae | Paspalum longum Chase | S | * | ||
| Poaceae | Schizachyrium mexicanum (Hitchc.) A. Camus | S | |||
| Poaceae | Schizachyrium sanguineum (Retz.) Alston | S | |||
| Poaceae | Setaria geminata (Forssk) Veldkamp | S | |||
| Poaceae | Setaria parviflora (Poir.) Kerguelen | S | |||
| Poaceae | Sorghastrum pohlianum Dávila, L.I. Cabrera & R. Lira | S | * | ||
| Poaceae | Sporobolus purpurascens (Swartz) Hamilton | S | |||
| Poaceae | Sporobolus pyramidatus (Lam.) Hitchc. | C | |||
| Poaceae | Urochloa maxima (Jacq.) R.D. Webster | S | * | ||
| Poaceae | Urochloa reptans (L.) Stapf | C | * | ||
| Polygonaceae | Antigonon leptopus Hook. & Arn. | S | * | ||
| Polypodiaceae | Pecluma alfredii (Rosenst.) M. G. Price var. occidentalis A. R. Smith | S | |||
| Polypodiaceae | Polypodium californicum Kaulfuss | S | |||
| Polypodiaceae | Polypodium polypodioides (L.) Watt. var. aciculare Weath. | S | |||
| Portulacaceae | Portulaca oleracea L. | S, C | * | ||
| Portulacaceae | Portulaca pilosa L | S, C | |||
| Potamogetonaceae | Potamogeton nodosus Poiret | S | |||
| Psilotaceae | Psilotum nudum (L.) P. Beauv. | S | |||
| Pteridaceae | Hemionitis radiata (L.) Christenh. | S | |||
| Pteridaceae | Cheilanthes peninsularis Maxon var. insularis Weath. | S, C | * | ||
| Pteridaceae | Pityrogramma ebenea (L.) Proctor | S | |||
| Rhamnaceae | Frangula discolor (Donn.Sm.) Grubov | S | |||
| Rhamnaceae | Rhamnus humboldtiana Willd. ex Schult. | C | |||
| Rosaceae | Prunus serotina Ehrh. | S | |||
| Rosaceae | Rubus sp. | S | |||
| Rubiaceae | Chiococca alba (L.) C. L. Hitchc. | S | |||
| Rubiaceae | Galium mexicanum Kunth | S | |||
| Rubiaceae | Guettarda insularis Brandegee | S | * | ||
| Rubiaceae | Houstonia mucronata (Benth.) B.L. Rob. | S | |||
| Rubiaceae | Mitracarpus hirtus (L.) DC. | S | * | ||
| Rubiaceae | Spermacoce nesiotica (Robinson) G. A. Levin | S, C | * | ||
| Rutaceae | Citrus × limon (L.) Osbeck | S | * | ||
| Rutaceae | Citrus x× aurantium L. | S | * | ||
| Rutaceae | Zanthoxylum fagara (L.) Sarg. | C | |||
| Rutaceae | Zanthoxylum insulare Rose | S | |||
| Sabiaceae | Meliosma nesites I. M. Johnst. | S | * | ||
| Santalaceae | Phoradendron quadrangulare (Kunth) Griseb. | S | |||
| Santalaceae | Phoradendron tonduzii Trel. | S | |||
| Sapindaceae | Cardiospermum halicacabum L. | S, C | |||
| Sapindaceae | Dodonaea viscosa (L.) Jacq. | S, C | |||
| Sapindaceae | Sapindus saponaria L. | C | |||
| Sapindaceae | Sapindus sp. | S | * | ||
| Sapotaceae | Manilkara zapota (L.) P. Royen | S | * | ||
| Sapotaceae | Sideroxylon socorrense (Brandegee) T. D. Penn. | S | * | ||
| Solanaceae | Cestrum pacificum Brandegee | S | * | ||
| Solanaceae | Datura aff. discolor Bernh. | C | |||
| Solanaceae | Nicotiana stocktonii T. S. Brandegee | S, C, SB | * | ||
| Solanaceae | Physalis clarionensis Waterf. | C | * | ||
| Solanaceae | Physalis mimulus Waterf. | S | * | ||
| Solanaceae | Physalis philadelphica Lam. | C | * | ||
| Solanaceae | Physalis pruinosa L. | S | * | ||
| Solanaceae | Solanum americanum Miller | S | * | ||
| Solanaceae | Solanum ferrugineum Jacq. | S | |||
| Thelypteridaceae | Thelypteris oligocarpa (Willd.) Ching | S | |||
| Verbenaceae | Citharexylum danirae León de la Luz & F. Chiang | S | * | ||
| Verbenaceae | Lantana involucrata L. var. socorrensis Moldenke | S | * | ||
| Verbenaceae | Lantana velutina M. Martens & Galeotti | S | |||
| Verbenaceae | Verbena sphaerocarpa L. M. Perry | S | * | ||
| Zygophyllaceae | Tribulus cistoides L. | S, C | * |
| Region | Cultivated | Casual | Naturalized | Total |
|---|---|---|---|---|
| Africa | 5 | 1 | 26 | 32 |
| Asia Tem. | 7 | - | 25 | 32 |
| Asia Trop. | 7 | - | 19 | 26 |
| Australasia | 4 | - | 9 | 13 |
| Europe | - | - | 5 | 5 |
| N. Am. | 11 | 12 | 43 | 66 |
| S. Am. | 9 | 11 | 38 | 58 |
| Family | Species | Island | Invasion Status | GCW Score | GCW Rating |
|---|---|---|---|---|---|
| Fabaceae | Leucaena leucocephala (Lam.) de Wit | S | N | 64 | Extreme |
| Fabaceae | Pithecellobium dulce (Roxb.) Benth. | S | Cas | 51.2 | Extreme |
| Fabaceae | Vachellia farnesiana (L.) Wight & Arn. var. farnesiana | S | N | 51.2 | Extreme |
| Cyperaceae | Cyperus rotundus L. | S | N | 44.8 | Extreme |
| Fabaceae | Macroptilium atropurpureum (DC.) Urban | S, C | N | 44.8 | Extreme |
| Fabaceae | Senna obtusifolia (L.) Irwin & Barneby | S | N | 44.8 | Extreme |
| Poaceae | Cenchrus echinatus L. | S, C | N | 44.8 | Extreme |
| Poaceae | Cynodon dactylon (L.) Pers. | S | N | 44.8 | Extreme |
| Portulacaceae | Portulaca oleracea L. | S, C | N | 44.8 | Extreme |
| Euphorbiaceae | Ricinus communis L. | S | N | 43.2 | Extreme |
| Arecaceae | Phoenix dactylifera L. | S | Cul | 38.4 | Extreme |
| Meliaceae | Swietenia macrophylla King | S | Cul | 38.4 | Extreme |
| Asteraceae | Sonchus asper (L.) Hill ssp. asper | S | N | 35.84 | Extreme |
| Fabaceae | Crotalaria incana L. | S | N | 35.84 | Extreme |
| Myrtaceae | Psidium guajava L. | S | N | 34.56 | Extreme |
| Poaceae | Cenchrus ciliaris L. | S, C | N | 33.6 | Extreme |
| Poaceae | Dactyloctenium aegyptium (L.) Willd. | S, C | N | 33.6 | Extreme |
| Poaceae | Echinochloa colona (L.) Link | S | N | 33.6 | Extreme |
| Arecaceae | Cocos nucifera L. | S, C | Cul | 28.8 | High |
| Asteraceae | Tridax procumbens L. | S | N | 26.88 | High |
| Malvaceae | Sida acuta Burm.f. | S | N | 26.88 | High |
| Chenopodiaceae | Dysphania ambrosioides (L.) Mosyakin & Clemants | S | N | 26.4 | High |
| Anacardiaceae | Mangifera indica L. | S | Cul | 25.92 | High |
| Fabaceae | Cassia fistula L. | S | Cul | 25.92 | High |
| Meliaceae | Cedrela odorata L. | S | N | 25.92 | High |
| Meliaceae | Melia azedarach L. | S | Cul | 25.92 | High |
| Convolvulaceae | Ipomoea purpurea (L.) Roth | S | Cul | 24 | High |
| Polygonaceae | Antigonon leptopus Hook. & Arn. | S | N | 24 | High |
| Solanaceae | Solanum americanum Miller | S | N | 24 | High |
| Poaceae | Melinis repens (Willdenow) Zizka | S, C | N | 20.16 | High |
| Caricaceae | Carica papaya L. | S | Cul | 19.44 | High |
| Combretaceae | Terminalia catappa L. | S | Cul | 19.44 | High |
| Euphorbiaceae | Euphorbia heterophylla L. | S | Cas | 19.2 | High |
| Zygophyllaceae | Tribulus cistoides L. | S, C | N | 19.2 | High |
| Poaceae | Eragrostis amabilis (L.) Wight & Arn. | S | N | 17.92 | High |
| Fabaceae | Tamarindus indica L. | S | Cul | 17.28 | High |
| Estimate | SE | t-Value | p | |
|---|---|---|---|---|
| Introduced | ||||
| Intercept | 4.07821 | 0.41238 | 9.889 | 3.92 × 10−6 |
| ln area (km2) | 0.29558 | 0.04989 | 5.925 | 0.000222 |
| Dispersion parameter: 42.16917 | ||||
| Null deviance: 1989.76 (10 df) | ||||
| Residual deviance: 400.98 (9 df) | ||||
| Native | ||||
| Intercept | 3.81134 | 0.81587 | 4.672 | 0.00117 |
| ln area (km2) | 0.3401 | 0.09741 | 3.491 | 0.00682 |
| Dispersion parameter: 169.7447 | ||||
| Null deviance: 3750.7 (10 df) | ||||
| Residual deviance: 1484.2 (9 df) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Meneses, A.; Martínez-Gómez, J.E.; Mejía-Saulés, T.; Acosta-Rosado, I.; Stadler, S. Vascular Plant Species Inventory of Mexico’s Revillagigedo National Park: Awareness of Alien Invaders as a Sine Qua Non Prerequisite for Island Conservation. Plants 2023, 12, 3455. https://doi.org/10.3390/plants12193455
Domínguez-Meneses A, Martínez-Gómez JE, Mejía-Saulés T, Acosta-Rosado I, Stadler S. Vascular Plant Species Inventory of Mexico’s Revillagigedo National Park: Awareness of Alien Invaders as a Sine Qua Non Prerequisite for Island Conservation. Plants. 2023; 12(19):3455. https://doi.org/10.3390/plants12193455
Chicago/Turabian StyleDomínguez-Meneses, Alejandra, Juan Esteban Martínez-Gómez, Teresa Mejía-Saulés, Israel Acosta-Rosado, and Stefan Stadler. 2023. "Vascular Plant Species Inventory of Mexico’s Revillagigedo National Park: Awareness of Alien Invaders as a Sine Qua Non Prerequisite for Island Conservation" Plants 12, no. 19: 3455. https://doi.org/10.3390/plants12193455