
Genome-Wide Characterization of DGATs and Their Expression Diversity Analysis in Response to Abiotic Stresses in Brassica napus

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Identification and Annotation of DGAT Family Members in B. napus
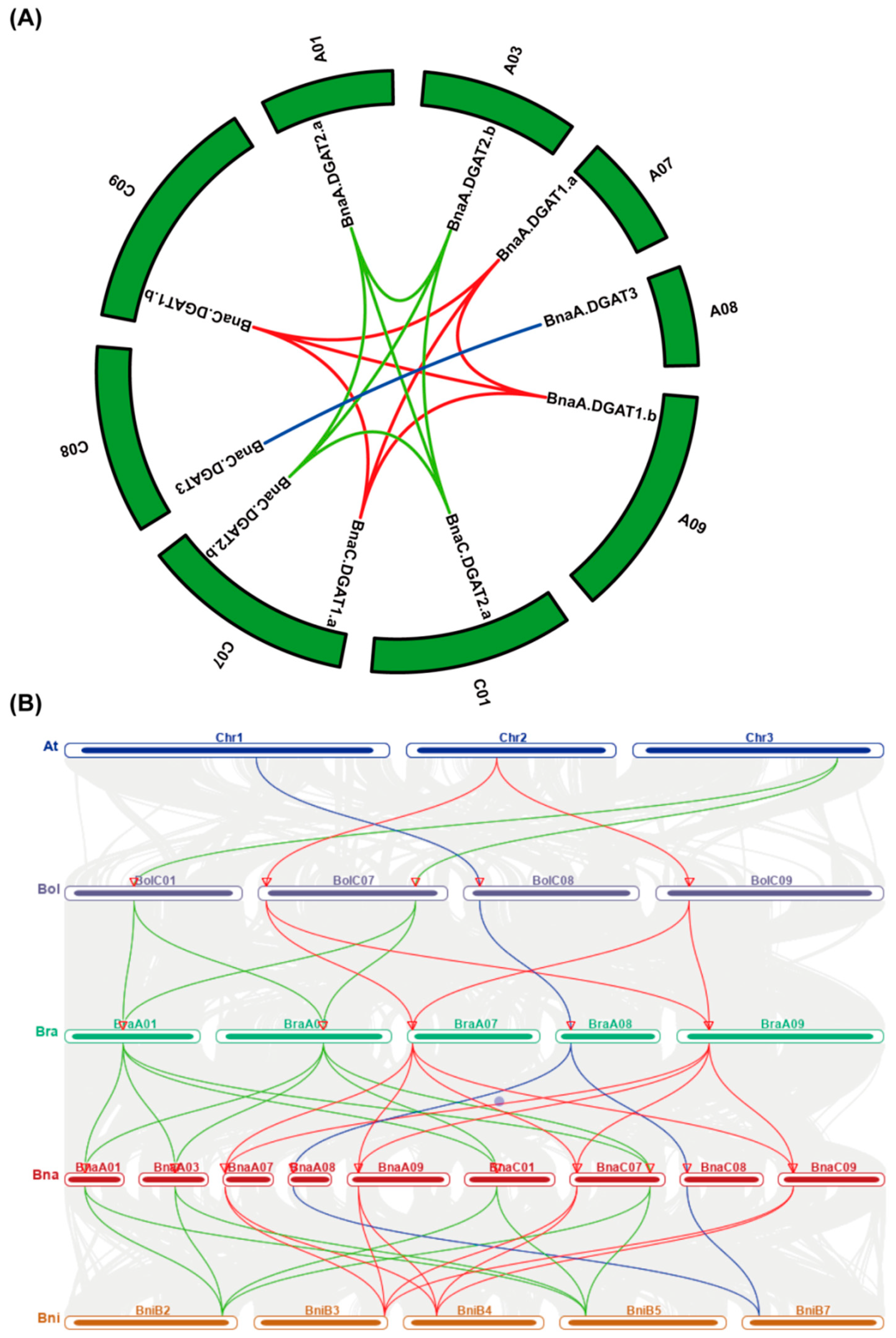
2.2. Chromosomal Location and Collinearity Analysis
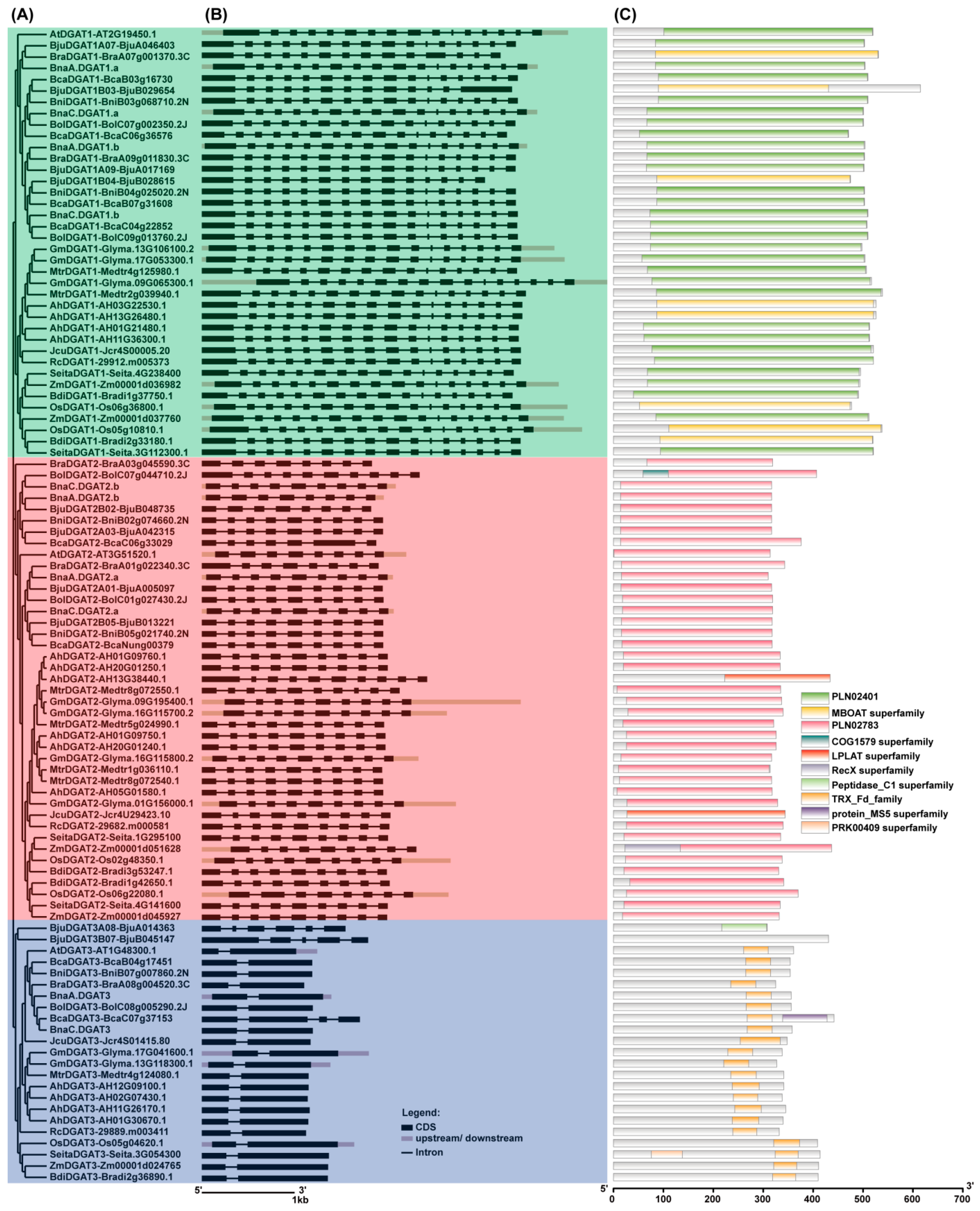
2.3. Evolutionary Relationship and Exon/Intron Gene Structure Analysis of BnaDGATs
2.4. The Conserved Domains and Motif Analyses
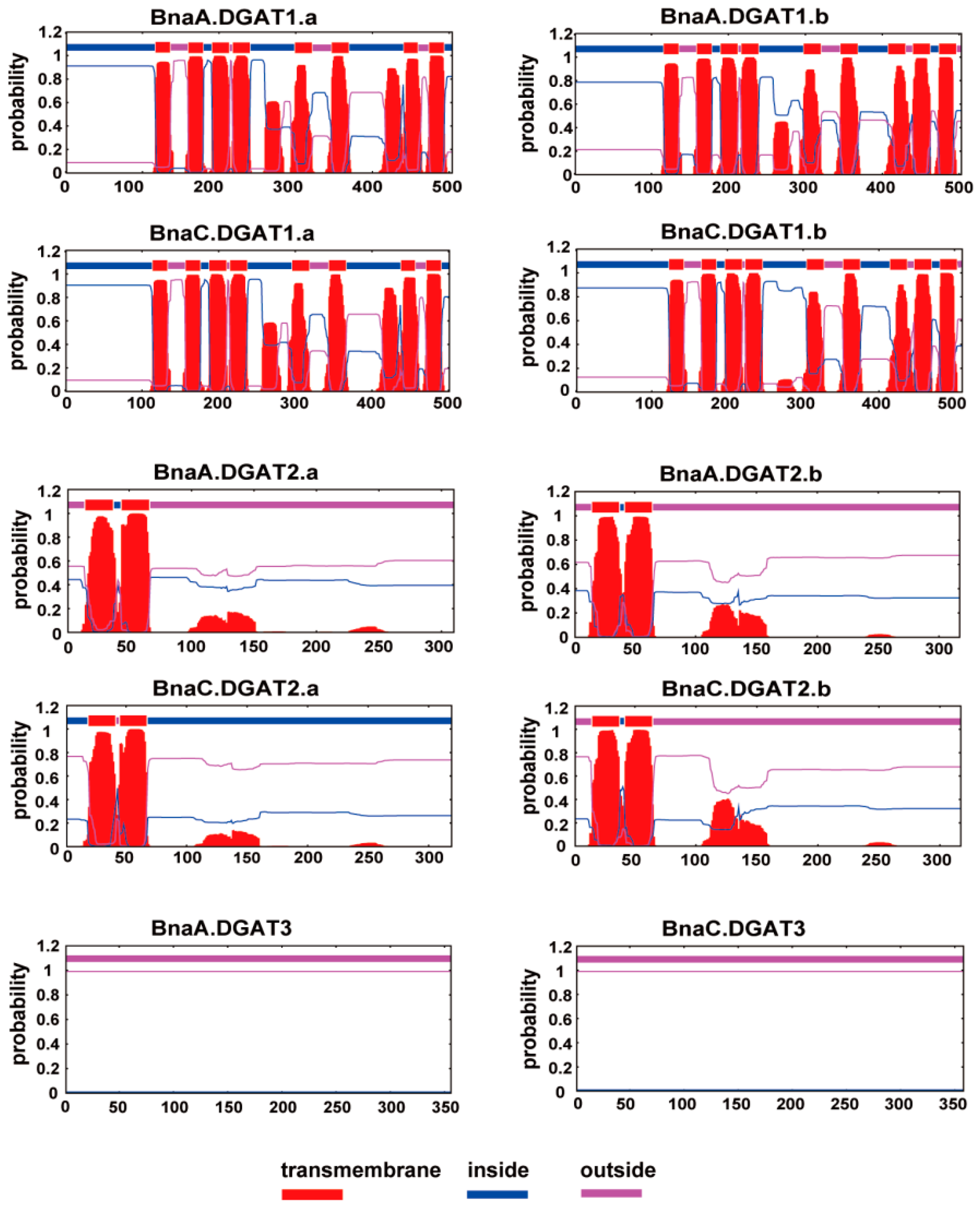
2.5. Putative Transmembrane Domains of DGAT Proteins
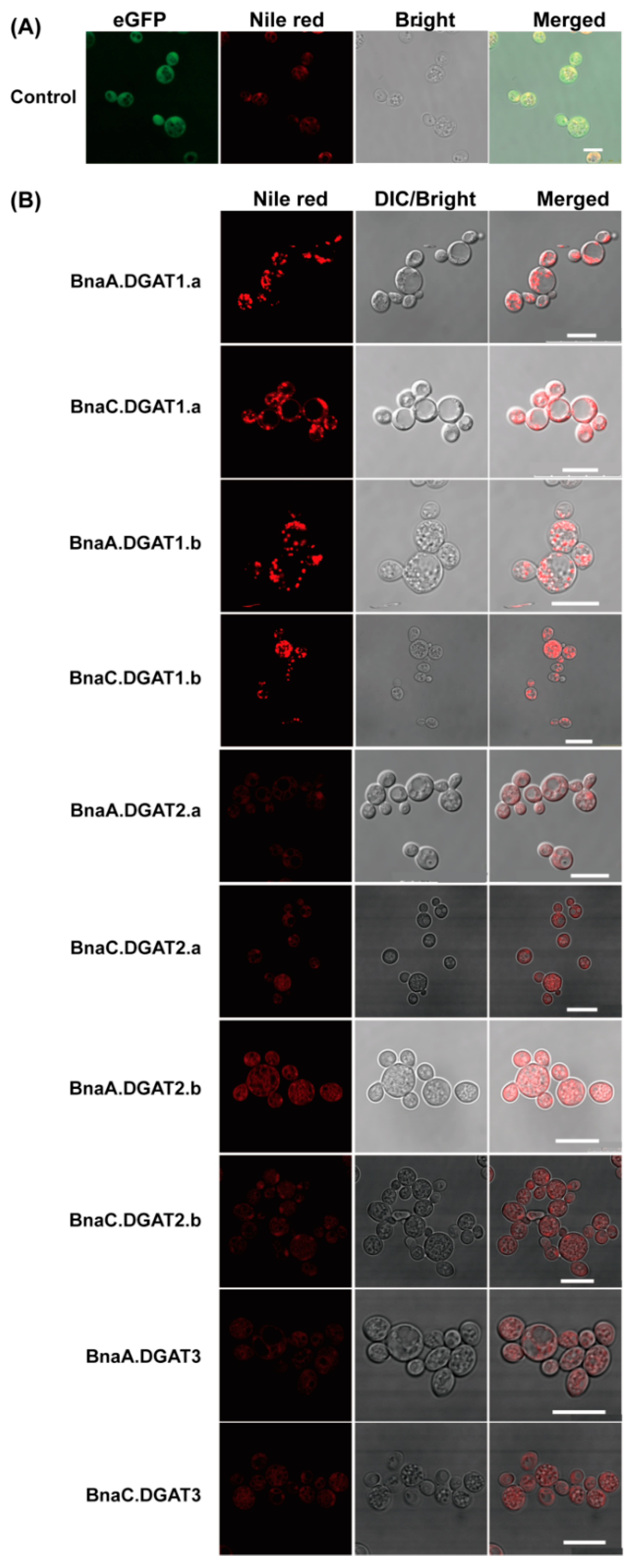
2.6. Oil Droplets in S. cerevisiae H1246 Overexpressing BnaDGATs
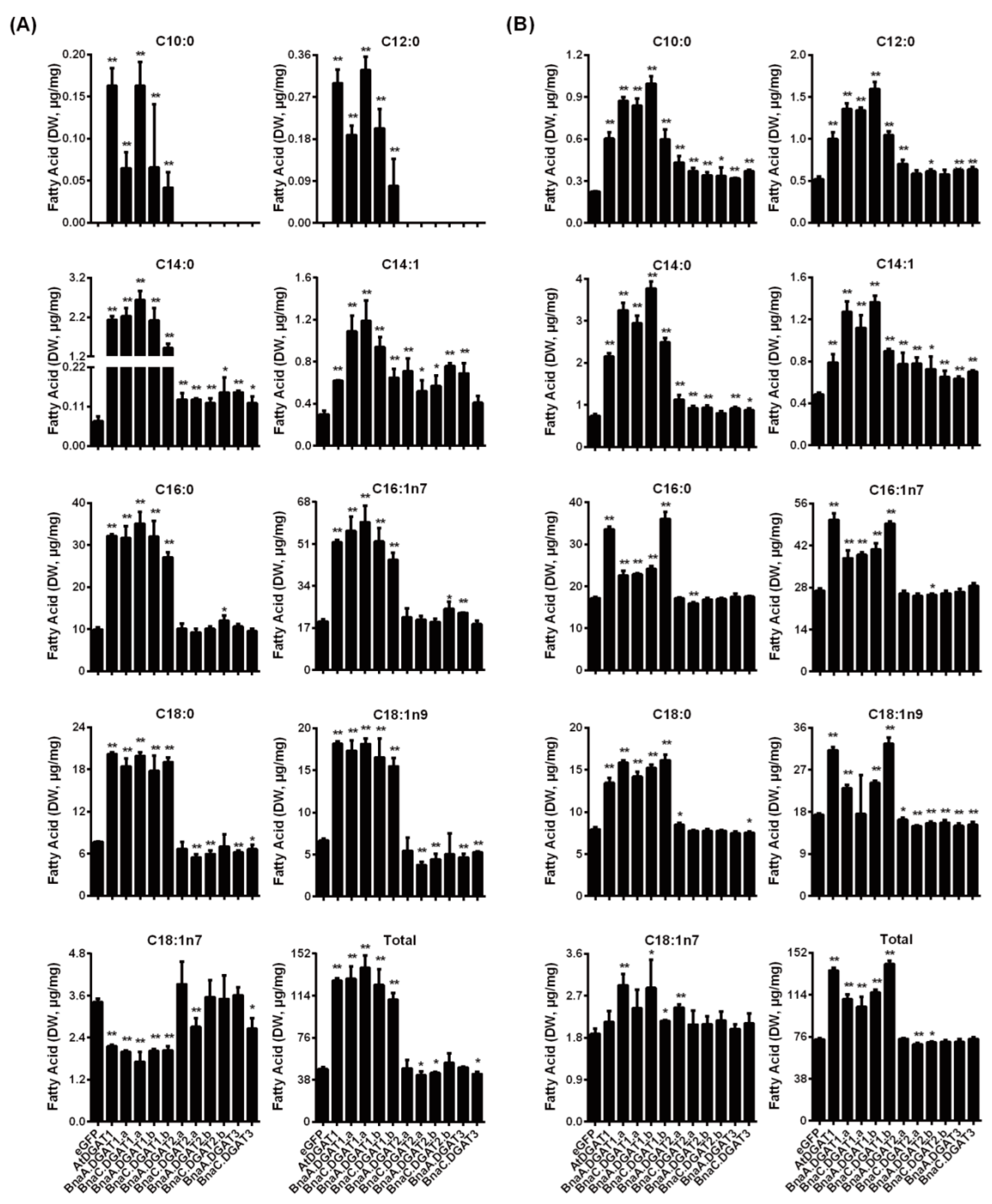
2.7. Fatty Acid Profiles in Yeast H1246 and INVSc1 Expressing BnaDGATs
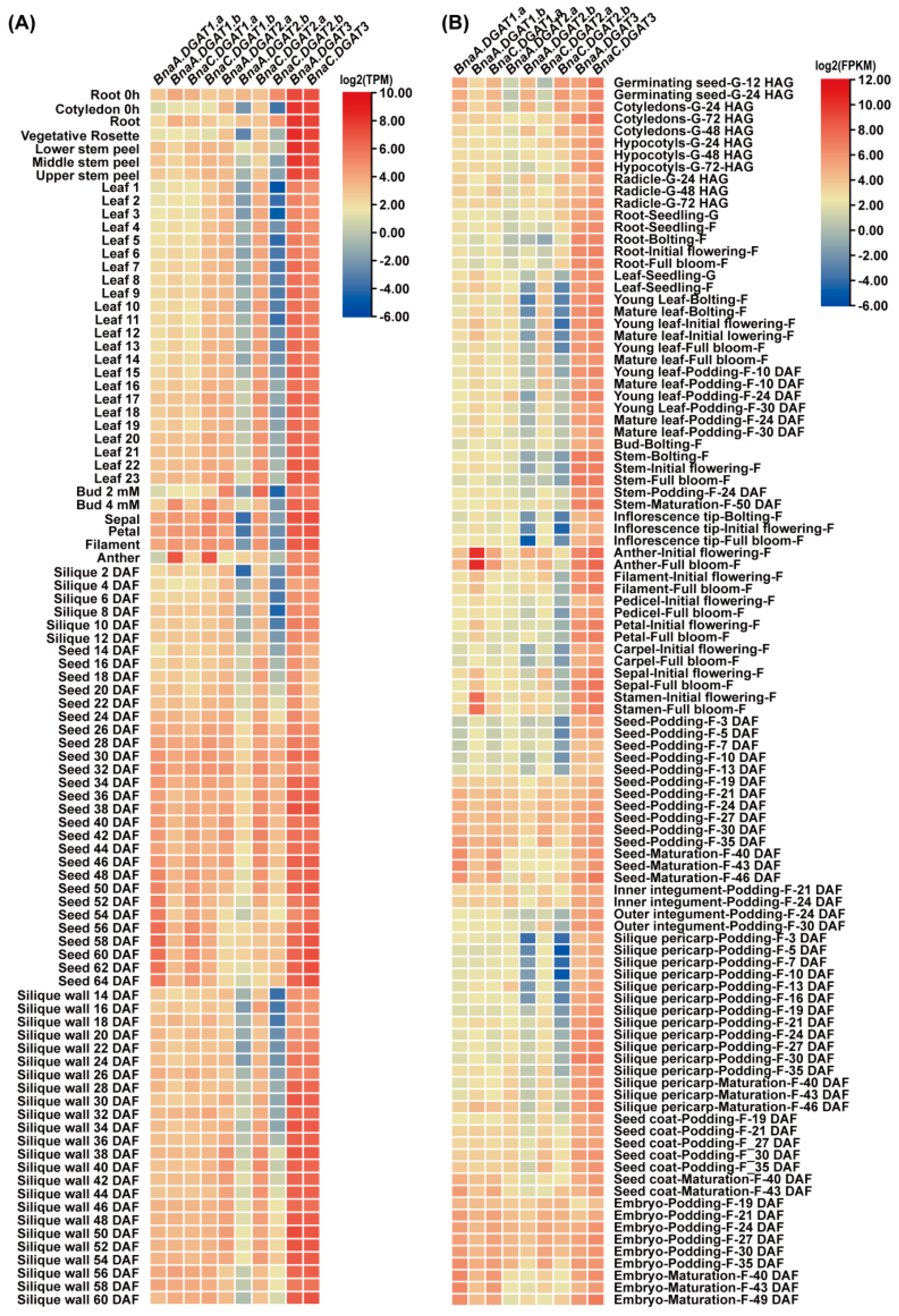
2.8. Transcriptomic and qRT-PCR Analysis of BnaDGATs in Different Tissues
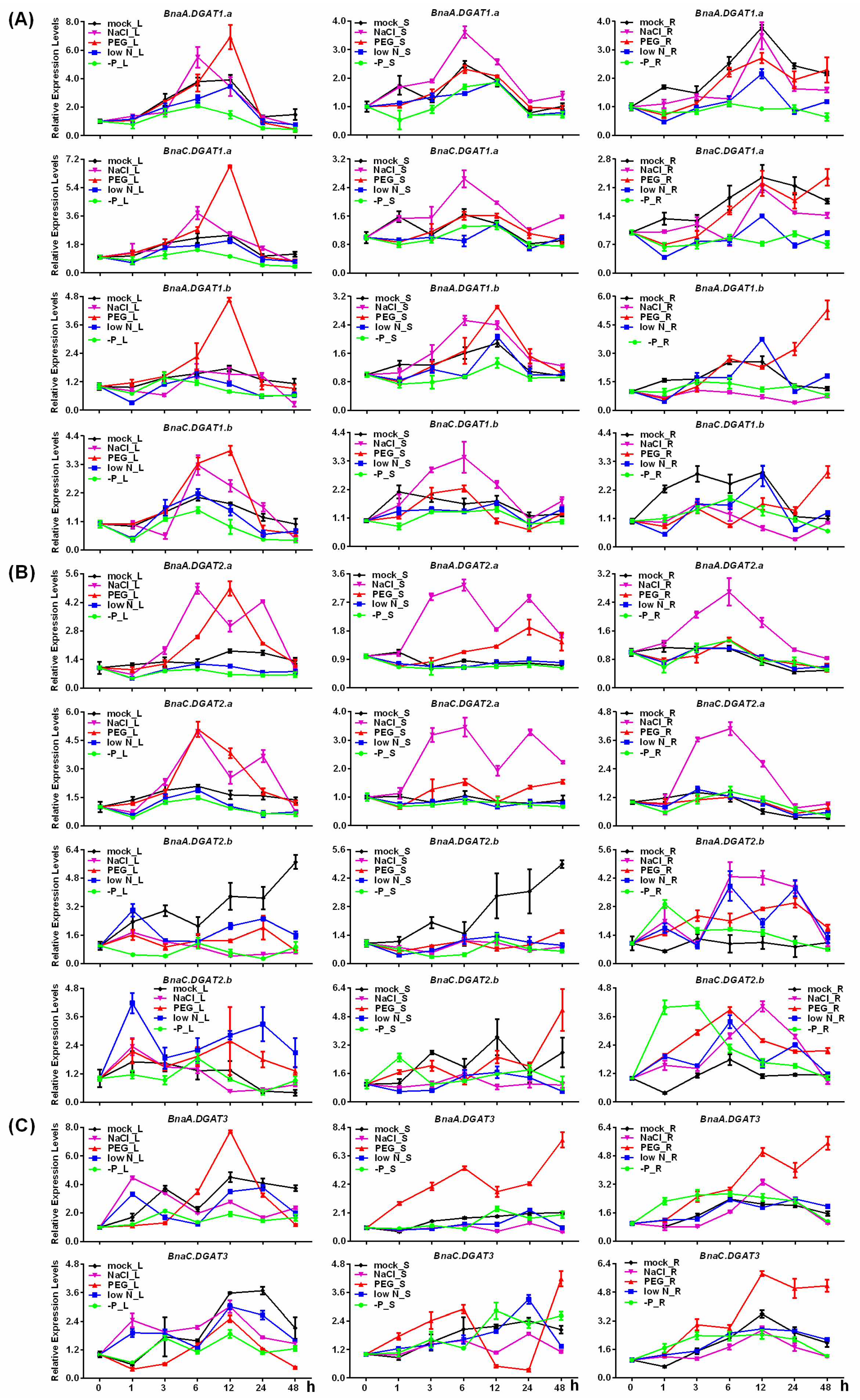
2.9. Expression Analysis of BnaDGATs under Different Stresses
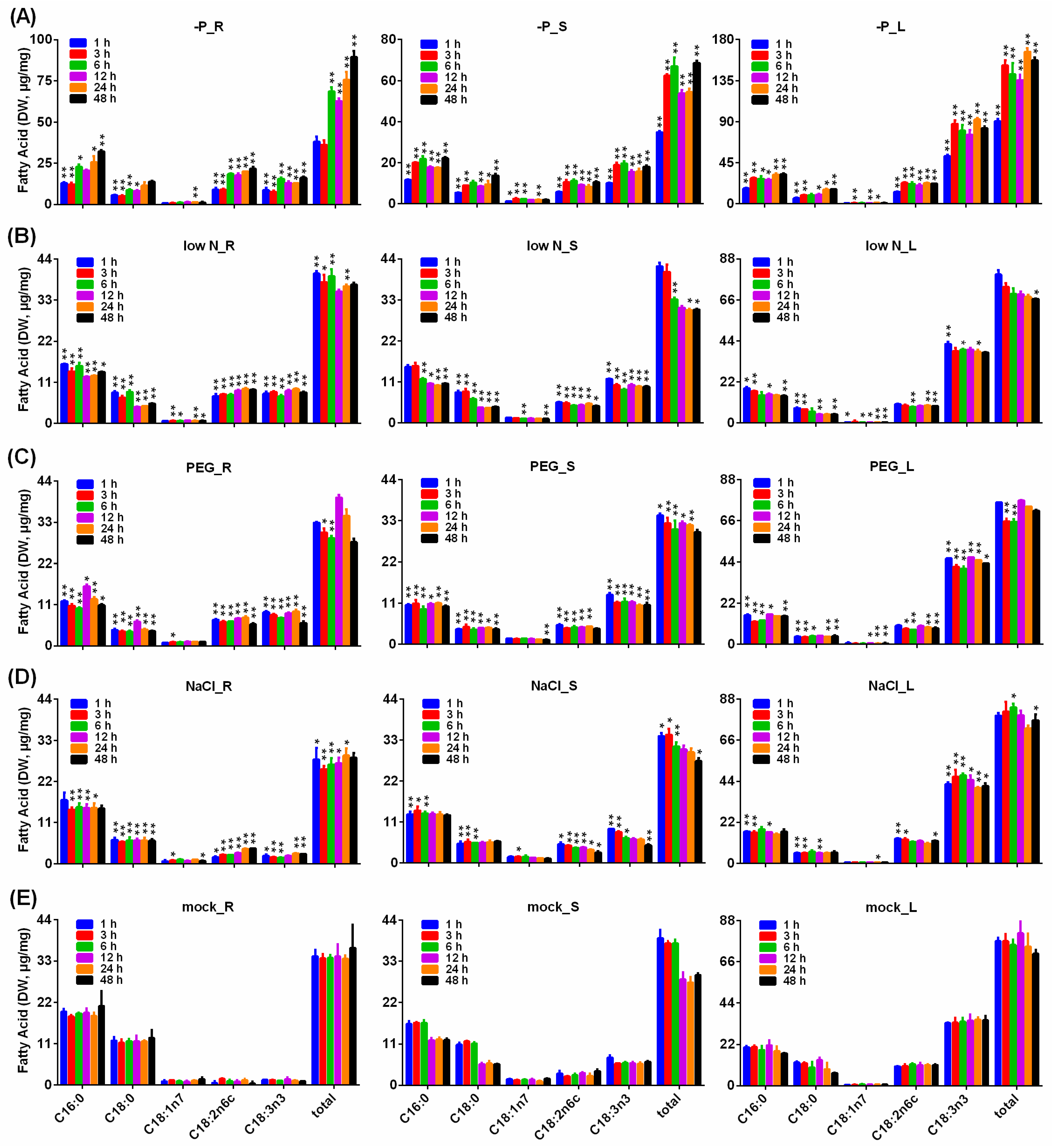
2.10. Analysis of Fatty Acids in B. napus Seedlings under P Starvation, Low N, Drought and Salinity Stresses
2.11. Cis-Elements in BnaDGAT Promoters and Transcription Factors and miRNA Regulating BnaDGATs
3. Discussion
3.1. Gene Duplication and Functional Diversification of DGAT Family Members
3.2. Role of BnaDGATs in Oil Biosynthesis
3.3. The Response of BnaDGATs to Abiotic Stresses
4. Materials and Methods
4.1. Identification of DGAT Family Members in B. napus and in Other Plants
4.2. Chromosomal Location and Gene Synteny Analysis
4.3. Phylogenetic, Gene Structure, Conserved Domain and Motif Analyses
4.4. Yeast Expression Vector Construction and Transformation
4.5. Nile Red Staining and Microscopy
4.6. Analysis of Fatty Acids in Yeast Transformants and B. napus Seedlings by GC–MS
4.7. B. napus Seedling Treatments and Sampling
4.8. Genomic DNA Extraction, Total RNA Isolation, Primary cDNA Synthesis and qRT–PCR
4.9. Expression Pattern Analysis Based on RNA-Seq Datasets
4.10. Analyses of Transcription Factors and miRNAs Targeting BnaDGATs and Cis-Acting Elements in BnaDGAT Promoters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.H.; Beisson, F.; Pollard, M.; Ohlrogge, J. Oil content of Arabidopsis seeds: The influence of seed anatomy, light and plant-to-plant variation. Phytochemistry 2006, 67, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Graham, I.A. Seed storage oil mobilization. Annu. Rev. Plant Biol. 2008, 59, 115–142. [Google Scholar] [CrossRef]
- Xu, Y.; Caldo, K.M.P.; Pal-Nath, D.; Ozga, J.; Lemieux, M.J.; Weselake, R.J.; Chen, G.Q. Properties and biotechnological applications of acyl-CoA:diacylglycerol acyltransferase and phospholipid:diacylglycerol acyltransferase from terrestrial plants and microalgae. Lipids 2018, 53, 663–688. [Google Scholar] [CrossRef]
- Liu, Q.; Siloto, R.M.P.; Lehner, R.; Stone, S.J.; Weselake, R.J. Acyl-CoA:diacylglycerol acyltransferase: Molecular biology, biochemistry and biotechnology. Prog. Lipid Res. 2012, 51, 350–377. [Google Scholar] [CrossRef]
- Kennedy, E.P.; Weiss, S.B. Function of cytidine coenzymes in the biosynthesis of phospholipides. J. Biol. Chem. 1956, 222, 193–214. [Google Scholar] [CrossRef]
- Marchive, C.; Nikovics, K.; To, A.; Lepiniec, L.; Baud, S. Transcriptional regulation of fatty acid production in higher plants: Molecular bases and biotechnological outcomes. Eur. J. Lipid Sci. Technol. 2014, 116, 1332–1343. [Google Scholar] [CrossRef]
- Hobbs, D.H.; Lu, C.F.; Hills, M.J. Cloning of a cDNA encoding diacylglycerol acyltransferase from Arabidopsis thaliana and its functional expression. FEBS Lett. 1999, 452, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Kroon, J.T.M.; Wei, W.; Simon, W.J.; Slabas, A.R. Identification and functional expression of a type 2 acyl-CoA:diacylglycerol acyltransferase (DGAT2) in developing castor bean seeds which has high homology to the major triglyceride biosynthetic enzyme of fungi and animals. Phytochemistry 2006, 67, 2541–2549. [Google Scholar] [CrossRef] [PubMed]
- Shockey, J.M.; Gidda, S.K.; Chapital, D.C.; Kuan, J.-C.; Dhanoa, P.K.; Bland, J.M.; Rothstein, S.J.; Mullen, R.T.; Dyer, J.M. Tung tree DGAT1 and DGAT2 have nonredundant functions in triacylglycerol biosynthesis and are localized to different subdomains of the endoplasmic reticulum. Plant Cell 2006, 18, 2294–2313. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Enugutti, B.; Rajakumari, S.; Rajasekharan, R. Cytosolic triacylglycerol biosynthetic pathway in oilseeds. Molecular cloning and expression of peanut cytosolic diacylglycerol acyltransferase. Plant Physiol. 2006, 141, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
- Banilas, G.; Karampelias, M.; Makariti, I.; Kourti, A.; Hatzopoulos, P. The olive DGAT2 gene is developmentally regulated and shares overlapping but distinct expression patterns with DGAT1. J. Exp. Bot. 2011, 62, 521–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannoulia, K.; Haralampidis, K.; Poghosyan, Z.; Murphy, D.J.; Hatzopoulos, P. Differential expression of diacylglycerol acyltransferase (DGAT) genes in olive tissues. Biochem. Soc. Trans. 2000, 28, 695–697. [Google Scholar] [CrossRef] [PubMed]
- Bouvier-Nave, P.; Benveniste, P.; Oelkers, P.; Sturley, S.L.; Schaller, H. Expression in yeast and tobacco of plant cDNAs encoding acyl CoA:diacylglycerol acyltransferase. Eur. J. Biochem. 2000, 267, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.B.; Wang, J.J.; Zhang, G.Y.; Liu, J.Q.; Manan, S.; Hu, H.H.; Zhao, J. Two types of soybean diacylglycerol acyltransferases are differentially involved in triacylglycerol biosynthesis and response to environmental stresses and hormones. Sci. Rep. 2016, 6, 28541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.Z.; Hatanaka, T.; Yu, K.S.; Wu, Y.M.; Fukushige, H.; Hildebrand, D. Soybean oil biosynthesis: Role of diacylglycerol acyltransferases. Funct. Integr. Genom. 2013, 13, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Zhang, J.S.; Gai, J.Y.; Chen, S.Y. Cloning and comparative analysis of the gene encoding diacylglycerol acyltransferase from wild type and cultivated soybean. Theor. Appl. Genet. 2006, 112, 1086–1097. [Google Scholar] [CrossRef]
- Chi, X.Y.; Hu, R.B.; Zhang, X.W.; Chen, M.N.; Chen, N.; Pan, L.J.; Wang, T.; Wang, M.A.; Yang, Z.; Wang, Q.F.; et al. Cloning and functional analysis of three diacylglycerol acyltransferase genes from peanut (Arachis hypogaea L.). PLoS ONE 2014, 9, e105834. [Google Scholar] [CrossRef]
- Peng, Z.Y.; Li, L.; Yang, L.Q.; Zhang, B.; Chen, G.; Bi, Y.P. Overexpression of peanut diacylglycerol acyltransferase 2 in Escherichia coli. PLoS ONE 2013, 8, e61363. [Google Scholar] [CrossRef]
- Nykiforuk, C.L.; Furukawa-Stoffer, T.L.; Huff, P.W.; Sarna, M.; Laroche, A.; Moloney, M.M.; Weselake, R.J. Characterization of cDNAs encoding diacylglycerol acyltransferase from cultures of Brassica napus and sucrose-mediated induction of enzyme biosynthesis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2002, 1580, 95–109. [Google Scholar] [CrossRef]
- Aznar-Moreno, J.; Denolf, P.; Van Audenhove, K.; De Bodt, S.; Engelen, S.; Fahy, D.; Wallis, J.G.; Browse, J. Type 1 diacylglycerol acyltransferases of Brassica napus preferentially incorporate oleic acid into triacylglycerol. J. Exp. Bot. 2015, 66, 6497–6506. [Google Scholar] [CrossRef] [Green Version]
- Greer, M.S.; Truksa, M.; Deng, W.; Lung, S.C.; Chen, G.Q.; Weselake, R.J. Engineering increased triacylglycerol accumulation in Saccharomyces cerevisiae using a modified type 1 plant diacylglycerol acyltransferase. Appl. Microbiol. Biotechnol. 2015, 99, 2243–2253. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Ouyang, C.; Kou, S.L.; Wang, S.H.; Yao, Y.Y.; Peng, T.; Xu, Y.; Tang, L.; Chen, F. Cloning and characterization of a cDNA encoding type 1 diacylglycerol acyltransferase from sunflower (Helianthus annuus L.). Z. Naturforsch. C J. Biosci. 2011, 66, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Y.; Francis, T.; Mietkiewska, E.; Giblin, E.M.; Barton, D.L.; Zhang, Y.; Zhang, M.; Taylor, D.C. Cloning and characterization of an acyl-CoA-dependent diacylglycerol acyltransferase 1 (DGAT1) gene from Tropaeolum majus, and a study of the functional motifs of the DGAT protein using site-directed mutagenesis to modify enzyme activity and oil content. Plant Biotechnol. J. 2008, 6, 799–818. [Google Scholar] [CrossRef]
- Zheng, P.; Allen, W.B.; Roesler, K.; Williams, M.E.; Zhang, S.; Li, J.; Glassman, K.; Ranch, J.; Nubel, D.; Solawetz, W.; et al. A phenylalanine in DGAT is a key determinant of oil content and composition in maize. Nat. Genet. 2008, 40, 367–372. [Google Scholar] [CrossRef]
- Zhao, Y.P.; Wu, N.; Li, W.J.; Shen, J.L.; Chen, C.; Li, F.G.; Hou, Y.X. Evolution and characterization of acetyl coenzyme A: Diacylglycerol acyltransferase genes in cotton identify the roles of GhDGAT3D in oil biosynthesis and fatty acid composition. Genes 2021, 12, 1045. [Google Scholar] [CrossRef]
- Zhang, F.Y.; Yang, M.F.; Xu, Y.N. Silencing of DGAT1 in tobacco causes a reduction in seed oil content. Plant Sci. 2005, 169, 689–694. [Google Scholar] [CrossRef]
- Jako, C.; Kumar, A.; Wei, Y.; Zou, J.; Barton, D.L.; Giblin, E.M.; Covello, P.S.; Taylor, D.C. Seed-specific over-expression of an Arabidopsis cDNA encoding a diacylglycerol acyltransferase enhances seed oil content and seed weight. Plant Physiol. 2001, 126, 861–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, D.C.; Zhang, Y.; Kumar, A.; Francis, T.; Giblin, E.M.; Barton, D.L.; Ferrie, J.R.; Laroche, A.; Shah, S.; Zhu, W.; et al. Molecular modification of triacylglycerol accumulation by over-expression of DGAT1 to produce canola with increased seed oil content under field conditions. Botany 2009, 87, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Maravi, D.K.; Kumar, S.; Sharma, P.K.; Kobayashi, Y.; Goud, V.V.; Sakurai, N.; Koyama, H.; Sahoo, L. Ectopic expression of AtDGAT1, encoding diacylglycerol O-acyltransferase exclusively committed to tag biosynthesis, enhances oil accumulation in seeds and leaves of jatropha. Biotechnol. Biofuels 2016, 9, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lock, Y.Y.; Snyder, C.L.; Zhu, W.M.; Siloto, R.M.P.; Weselake, R.J.; Shah, S. Antisense suppression of type 1 diacylglycerol acyltransferase adversely affects plant development in Brassica napus. Physiol. Plant. 2009, 137, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Routaboul, J.M.; Benning, C.; Bechtold, N.; Caboche, M.; Lepiniec, L. The TAG1 locus of Arabidopsis encodes for a diacylglycerol acyltransferase. Plant Physiol. Biochem. 1999, 37, 831–840. [Google Scholar] [CrossRef]
- Kaup, M.T.; Froese, C.D.; Thompson, J.E. A role for diacylglycerol acyltransferase during leaf senescence. Plant Physiol. 2002, 129, 1616–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Fan, J.; Taylor, D.C.; Ohlrogge, J.B. DGAT1 and PDAT1 acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development. Plant Cell 2009, 21, 3885–3901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Hills, M.J. Arabidopsis mutants deficient in diacylglycerol acyltransferase display increased sensitivity to abscisic acid, sugars, and osmotic stress during germination and seedling development. Plant Physiol. 2002, 129, 1352–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weselake, R.J.; Shah, S.; Tang, M.; Quant, P.A.; Snyder, C.L.; Furukawastoffer, T.L.; Zhu, W.; Taylor, D.C.; Zou, J.; Kumar, A. Metabolic control analysis is helpful for informed genetic manipulation of oilseed rape (Brassica napus) to increase seed oil content. J. Exp. Bot. 2008, 59, 3543–3549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Yu, X.C.; Song, L.F.; An, C.C. ABI4 activates DGAT1 expression in Arabidopsis seedlings during nitrogen deficiency. Plant Physiol. 2011, 156, 873–883. [Google Scholar] [CrossRef] [Green Version]
- Kelly, A.A.; van Erp, H.; Quettier, A.L.; Shaw, E.; Menard, G.; Kurup, S.; Eastmond, P.J. The SUGAR-DEPENDENT1 lipase limits triacylglycerol accumulation in vegetative tissues of arabidopsis. Plant Physiol. 2013, 162, 1282–1289. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Chen, S.; Yang, Y.; An, C. ABA-insensitive (ABI) 4 and ABI5 synergistically regulate DGAT1 expression in Arabidopsis seedlings under Stress. FEBS Lett. 2013, 587, 3076–3082. [Google Scholar] [CrossRef] [Green Version]
- Tjellstrom, H.; Strawsine, M.; Ohlrogge, J.B. Tracking synthesis and turnover of triacylglycerol in leaves. J. Exp. Bot. 2015, 66, 1453–1461. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.L.; Yu, L.H.; Xu, C.C. A central role for triacylglycerol in membrane lipid breakdown, fatty acid beta-oxidation, and plant survival under extended darkness. Plant Physiol. 2017, 174, 1517–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arisz, S.A.; Heo, J.Y.; Koevoets, I.T.; Zhao, T.; van Egmond, P.; Meyer, A.J.; Zeng, W.Q.; Niu, X.M.; Wang, B.S.; Mitchell-Olds, T.; et al. Diacylglycerol acyltransferase1 contributes to freezing tolerance. Plant Physiol. 2018, 177, 1410–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.J.; Yang, Y.C.; Zhou, Y.; Huang, L.P.; Xu, L.; Chen, Q.F.; Yu, L.J.; Xiao, S. Diacylglycerol acyltransferase and diacylglycerol kinase modulate triacylglycerol and phosphatidic acid production in the plant response to freezing stress. Plant Physiol. 2018, 177, 1303–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgal, J.; Shockey, J.; Lu, C.F.; Dyer, J.; Larson, T.; Graham, I.; Browse, J. Metabolic engineering of hydroxy fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil. Plant Biotechnol. J. 2008, 6, 819–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.Z.; Yu, K.S.; Hatanaka, T.; Hildebrand, D.F. Vernonia DGATs increase accumulation of epoxy fatty acids in oil. Plant Biotechnol. J. 2010, 8, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Demski, K.; Jeppson, S.; Lager, I.; Misztak, A.; Jasieniecka-Gazarkiewicz, K.; Waleron, M.; Stymne, S.; Banas, A. Isoforms of acyl-CoA:diacylglycerol acyltransferase2 differ Substantially in their specificities towards erucic acid. Plant Physiol. 2019, 181, 1468–1479. [Google Scholar] [CrossRef] [PubMed]
- Ayme, L.; Arragain, S.; Canonge, M.; Baud, S.; Touati, N.; Bimai, O.; Jagic, F.; Louis-Mondesir, C.; Briozzo, P.; Fontecave, M.; et al. Arabidopsis thaliana DGAT3 is a [2Fe-2S] protein involved in TAG biosynthesis. Sci. Rep. 2018, 8, 17254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, M.L.; Whitehead, L.; He, Z.S.; Gazda, V.; Gilday, A.; Kozhevnikova, E.; Vaistij, F.E.; Larson, T.R.; Graham, I.A. A cytosolic acyltransferase contributes to triacylglycerol synthesis in sucrose-rescued Arabidopsis seed oil catabolism mutants. Plant Physiol. 2012, 160, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.H.; Turner, C.; Chen, G.Q.; Lin, J.T.; McKeon, T.A. Cloning and characterization of a cDNA encoding diacylglycerol acyltransferase from castor bean. Lipids 2004, 39, 311–318. [Google Scholar] [CrossRef]
- Lu, C.F.L.; de Noyer, S.B.; Hobbs, D.H.; Kang, J.L.; Wen, Y.C.; Krachtus, D.; Hills, M.J. Expression pattern of diacylglycerol acyltransferase-1, an enzyme involved in triacylglycerol biosynthesis, in Arabidopsis thaliana. Plant Mol. Biol. 2003, 52, 31–41. [Google Scholar] [CrossRef]
- Zhao, C.Z.; Li, H.; Zhang, W.X.; Wang, H.L.; Xu, A.X.; Tian, J.H.; Zou, J.T.; Taylor, D.C.; Zhang, M. BnDGAT1s function similarly in oil deposition and are expressed with uniform patterns in tissues of Brassica napus. Front. Plant Sci. 2017, 8, 2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uagaharu, N. Genomic analysis of Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Jpn. J. Bot. 1935, 7, 389–452. [Google Scholar]
- Snowdon, R.J. Cytogenetics and genome analysis in Brassica crops. Chromosome Res. 2007, 15, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, B.; Denoeud, F.; Liu, S.Y.; Parkin, I.A.P.; Tang, H.B.; Wang, X.Y.; Chiquet, J.; Belcram, H.; Tong, C.B.; Samans, B.; et al. Early allopolyploid evolution in the post-neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Ostergaard, L.; King, G.J. Standardized gene nomenclature for the Brassica genus. Plant Methods 2008, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Wu, J.; Wang, X.W. Genome triplication drove the diversification of Brassica plants. Hortic. Res. 2014, 1, 14024. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.N.; Wang, J.Y.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Sandager, L.; Gustavsson, M.H.; Stahl, U.; Dahlqvist, A.; Wiberg, E.; Banas, A.; Lenman, M.; Ronne, H.; Stymne, S. Storage lipid synthesis is non-essential in yeast. J. Biol. Chem. 2002, 277, 6478–6482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Yu, L.; Wei, L.; Yu, P.; Wang, J.; Zhao, H.; Zhang, Y.; Zhang, S.; Yang, Z.; Chen, G.; et al. BnTIR: An online transcriptome platform for exploring RNA-seq libraries for oil crop Brassica napus. Plant Biotechnol. J. 2021, 19, 1895–1897. [Google Scholar] [CrossRef]
- Chao, H.; Li, T.; Luo, C.; Huang, H.; Ruan, Y.; Li, X.; Niu, Y.; Fan, Y.; Sun, W.; Zhang, K.; et al. BrassicaEDB: A gene expression database for Brassica crops. Int. J. Mol. Sci. 2020, 21, 5831. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Noman, A.; Fahad, S.; Aqeel, M.; Ali, U.; Amanullah; Anwar, S.; Baloch, S.K.; Zainab, M. miRNAs: Major modulators for crop growth and development under abiotic stresses. Biotechnol. Lett. 2017, 39, 685–700. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J.H. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 1819, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Turchetto-Zolet, A.C.; Christoff, A.P.; Kulcheski, F.R.; Loss-Morais, G.; Margis, R.; Margis-Pinheiro, M. Diversity and evolution of plant diacylglycerol acyltransferase (DGATs) unveiled by phylogenetic, gene structure and expression analyses. Genet. Mol. Biol. 2016, 39, 524–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weselake, R.J.; Madhavji, M.; Szarka, S.J.; Patterson, N.A.; Wiehler, W.B.; Nykiforuk, C.L.; Burton, T.L.; Boora, P.S.; Mosimann, S.C.; Foroud, N.A.; et al. Acyl-CoA-binding and self-associating properties of a recombinant 13.3 kDa N-terminal fragment of diacylglycerol acyltransferase-1 from oilseed rape. BMC Biochem. 2006, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldo, K.M.P.; Greer, M.S.; Chen, G.Q.; Lemieux, M.J.; Weselake, R.J. Purification and properties of recombinant Brassica napus diacylglycerol acyltransferase 1. FEBS Lett. 2015, 589, 773–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Ji, H.; Yang, Z. Functional characterization of three novel genes encoding diacylglycerol acyltransferase (DGAT) from oil-rich tubers of Cyperus esculentus. Plant Cell Physiol. 2020, 61, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.R.; Shrestha, P.; Yin, F.; Petrie, J.R.; Singh, S.P. AtDGAT2 is a functional acyl-CoA:diacylglycerol acyltransferase and displays different acyl-CoA substrate preferences than AtDGAT1. FEBS Lett. 2013, 587, 2371–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Cui, Q.; Xu, Y.; Yang, S.; Gao, M.; Wang, Y. Effects of tung oilseed FAD2 and DGAT2 genes on unsaturated fatty acid accumulation in Rhodotorula glutinis and Arabidopsis thaliana. Mol. Genet. Genom. 2015, 1605–1613. [Google Scholar] [CrossRef] [PubMed]
- Kainou, K.; Kamisaka, Y.; Kimura, K.; Uemura, H. Isolation of Δ12 and ω3-fatty acid desaturase genes from the yeast Kluyveromyces lactis and their heterologous expression to produce linoleic and α-linolenic acids in Saccharomyces cerevisiae. Yeast 2006, 23, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Gao, Y.; Zhang, F.; Liu, B.; Ji, C.; Xue, J.; Yuan, L.; Li, R. Functional characterization of an novel acyl-CoA:diacylglycerol acyltransferase 3-3 (CsDGAT3-3) gene from Camelina sativa. Plant Sci. 2021, 303, 110752. [Google Scholar] [CrossRef]
- Yan, B.W.; Xu, X.X.; Gu, Y.N.; Zhao, Y.; Zhao, X.C.; He, L.; Zhao, C.J.; Li, Z.T.; Xu, J.Y. Genome-wide characterization and expression profiling of diacylglycerol acyltransferase genes from maize. Genome 2018, 61, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.P.; Wang, Y.M.; Huang, Y.; Cui, Y.P.; Hua, J.P. Gene network of oil accumulation reveals expression profiles in developing embryos and fatty acid composition in upland cotton. J. Plant Physiol. 2018, 228, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Sajjadi, B.; Chen, W.Y.; Raman, A.A.A.; Ibrahim, S. Microalgae lipid and biomass for biofuel production: A comprehensive review on lipid enhancement strategies and their effects on fatty acid composition. Renew. Sustain. Energy Rev. 2018, 97, 200–232. [Google Scholar] [CrossRef]
- Liang, K.H.; Zhang, Q.H.; Gu, M.; Cong, W. Effect of phosphorus on lipid accumulation in freshwater microalga Chlorella sp. J. Appl. Phycol. 2013, 25, 311–318. [Google Scholar] [CrossRef]
- Li, X.; Hu, H.Y.; Gan, K.; Sun, Y.X. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494–5500. [Google Scholar] [CrossRef]
- Khozin-Goldberg, I.; Cohen, Z. The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water eustigmatophyte Monodus subterraneus. Phytochemistry 2006, 67, 696–701. [Google Scholar] [CrossRef]
- Guschina, I.A.; Dobson, G.; Harwood, J.L. Lipid metabolism in cultured lichen photobionts with different phosphorus status. Phytochemistry 2003, 64, 209–217. [Google Scholar] [CrossRef]
- Markou, G.; Chatzipavlidis, I.; Georgakakis, D. Carbohydrates production and bio-flocculation characteristics in cultures of Arthrospira (Spirulina) platensis: Improvements through phosphorus limitation process. Bioenergy Res. 2012, 5, 915–925. [Google Scholar] [CrossRef]
- Shimojima, M.; Madoka, Y.; Fujiwara, R.; Murakawa, M.; Yoshitake, Y.; Ikeda, K.; Koizumi, R.; Endo, K.; Ozaki, K.; Ohta, H. An engineered lipid remodeling system using a galactolipid synthase promoter during phosphate starvation enhances oil accumulation in plants. Front. Plant Sci. 2015, 6, 664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pant, B.D.; Burgos, A.; Pant, P.; Cuadros-Inostroza, A.; Willmitzer, L.; Scheible, W.R. The transcription factor PHR1 regulates lipid remodeling and triacylglycerol accumulation in Arabidopsis thaliana during phosphorus starvation. J. Exp. Bot. 2015, 66, 1907–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.Q.; Xu, Y.; Siloto, R.M.P.; Caldo, K.M.P.; Vanhercke, T.; El Tahchy, A.; Niesner, N.; Chen, Y.Y.; Mietkiewska, E.; Weselake, R.J. High-performance variants of plant diacylglycerol acyltransferase 1 generated by directed evolution provide insights into structure function. Plant J. 2017, 92, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Caldo, K.M.P.; Shen, W.; Xu, Y.; Hanley-Bowdoin, L.; Chen, G.Q.; Weselake, R.J.; Lemieux, M.J. Diacylglycerol acyltransferase 1 is activated by phosphatidate and inhibited by SnRK1-catalyzed phosphorylation. Plant J. 2018, 96, 287–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Ramirez, A.; Oropeza-Aburto, A.; Razo-Hernandez, F.; Ramirez-Chavez, E.; Herrera-Estrella, L. Phospholipase DZ2 plays an important role in extraplastidic galactolipid biosynthesis and phosphate recycling in Arabidopsis roots. Proc. Natl. Acad. Sci. USA 2006, 103, 6765–6770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oropeza-Aburto, A.; Cruz-Ramirez, A.; Acevedo-Hernandez, G.J.; Perez-Torres, C.A.; Caballero-Perez, J.; Herrera-Estrella, L. Functional analysis of the Arabidopsis PLDZ2 promoter reveals an evolutionarily conserved low-pi-responsive transcriptional enhancer element. J. Exp. Bot. 2012, 63, 2189–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fragoso, S.; Espindola, L.; Paez-Valencia, J.; Gamboa, A.; Camacho, Y.; Martinez-Barajas, E.; Coello, P. SnRK1 Isoforms AKIN10 and AKIN11 are differentially regulated in Arabidopsis plants under phosphate starvation. Plant Physiol. 2009, 149, 1906–1916. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.D.; Li, Y.J.; Fei, X.W. The mRNA abundance of pepc2 gene is negatively correlated with oil content in Chlamydomonas reinhardtii. Biomass Bioenergy 2011, 35, 1811–1817. [Google Scholar] [CrossRef]
- Illman, A.M.; Scragg, A.H.; Shales, S.W. Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enzym. Microb. Technol. 2000, 27, 631–635. [Google Scholar] [CrossRef]
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef]
- Spoehr, H.A.; Milner, H.W. The chemical composition of chlorella; effect of environmental conditions. Plant Physiol. 1949, 24, 120–149. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.J.; Fan, C.M.; Chen, Y.H.; Wang, J.Q.; Yin, W.B.; Wang, R.R.C.; Hu, Z.M. Identification and characterization of an efficient acyl-CoA: Diacylglycerol acyltransferase 1 (DGAT1) gene from the microalga Chlorella ellipsoidea. BMC Plant Biol. 2017, 17, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, C.; Godfrey, V.; Wahlen, B.; Seefeldt, L.; Bugbee, B. Understanding precision nitrogen stress to optimize the growth and lipid cross mark content tradeoff in oleaginous green microalgae. Bioresour. Technol. 2013, 131, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Biswas, K.; Singh, P.K.; Singh, P.K.; Elumalai, S.; Shukla, P.; Pabbi, S. Lipid production and molecular dynamics simulation for regulation of accD gene in cyanobacteria under different N and P regimes. Biotechnol. Biofuels 2017, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.; Oswald, O.; Graham, I.A. Arabidopsis seedling growth, storage lipid mobilization, and photosynthetic gene expression are regulated by carbon; nitrogen availability. Plant Physiol. 2002, 128, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Gaude, N.; Brehelin, C.; Tischendorf, G.; Kessler, F.; Dormann, P. Nitrogen deficiency in Arabidopsis affects galactolipid composition and gene expression and results in accumulation of fatty acid phytyl esters. Plant J. 2007, 49, 729–739. [Google Scholar] [CrossRef] [Green Version]
- Eastmond, P.J.; Germain, V.; Lange, P.R.; Bryce, J.H.; Smith, S.M.; Graham, I.A. Postgerminative growth and lipid catabolism in oilseeds lacking the glyoxylate cycle. Proc. Natl. Acad. Sci. USA 2000, 97, 5669–5674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svenningsson, H.; Liljenberg, C. Membrane lipid changes in root cells of rape (Brassica napus) as a function of water-deficit stress. Physiol. Plant. 1986, 68, 53–58. [Google Scholar] [CrossRef]
- Dakhma, W.S.; Zarrouk, M.; Cherif, A. Effects of drought-stress on lipids in rape leaves. Phytochemistry 1995, 40, 1383–1386. [Google Scholar] [CrossRef]
- Aziz, A.; Larher, F. Osmotic stress induced changes in lipid composition and peroxidation in leaf discs of Brassica napus L. J. Plant Physiol. 1998, 153, 754–762. [Google Scholar] [CrossRef]
- Aslam, M.N.; Nelson, M.N.; Kailis, S.G.; Bayliss, K.L.; Speijers, J.; Cowling, W.A. Canola oil increases in polyunsaturated fatty acids and decreases in oleic acid in drought-stressed Mediterranean-type environments. Plant Breed. 2009, 128, 348–355. [Google Scholar] [CrossRef]
- Douglas, T.J.; Paleg, L.G. Lipid composition of Zea mays seedlings and water stress-induced changes. J. Exp. Bot. 1981, 32, 499–508. [Google Scholar] [CrossRef]
- Martin, B.A.; Schoper, J.B.; Rinne, R.W. Changes in soybean (Glycine max [L.] Merr) glycerolipids in response to water-stress. Plant Physiol. 1986, 81, 798–801. [Google Scholar] [CrossRef] [Green Version]
- Thi, A.T.P.; Borrelflood, C.; Dasilva, J.V.; Justin, A.M.; Mazliak, P. Effects of water stress on lipid metabolism in cotton leaves. Phytochemistry 1985, 24, 723–727. [Google Scholar] [CrossRef]
- Urao, T.; Yamaguchishinozaki, K.; Urao, S.; Shinozaki, K. An Arabidopsis myb homolog is induced by dehydration stress and its gene product binds to the conserved MYB recognition sequence. Plant Cell 1993, 5, 1529–1539. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.W.; Wang, M.P.; Guo, Z.T.; Zhu, X.F.; Xia, Z.L. Identification of a 119-bp promoter of the maize sulfite oxidase gene (ZmSO) that confers high-level gene expression and ABA or drought inducibility in transgenic plants. Int. J. Mol. Sci. 2019, 20, 3326. [Google Scholar] [CrossRef] [Green Version]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Zhu, Y.; Liu, R.; Zhang, A.; Zhu, M.; Xu, W.; Lin, A.; Lu, K.; Li, J. Genome wide identification and comparative analysis of glutathione transferases (GST) family genes in Brassica napus. Sci. Rep. 2019, 9, 9196. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soil. Calif. Agric. Exp. Stn. Bull. 1938, 347, 36–39. [Google Scholar]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Yang, D.C.; Meng, Y.Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2020, 48, D1104–D1113. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.J.; Chen, H.; Yang, M.; Wang, J.P.; Pandey, M.K.; Zhang, C.; Chang, W.C.; Zhang, L.S.; Zhang, X.T.; Tang, R.H.; et al. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication. Nat. Genet. 2019, 51, 865–876. [Google Scholar] [CrossRef]
- Kaul, S.; Koo, H.L.; Jenkins, J.; Rizzo, M.; Rooney, T.; Tallon, L.J.; Feldblyum, T.; Nierman, W.; Benito, M.I.; Lin, X.Y.; et al. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [Green Version]
- Huala, E.; Dickerman, A.W.; Garcia-Hernandez, M.; Weems, D.; Reiser, L.; LaFond, F.; Hanley, D.; Kiphart, D.; Zhuang, M.; Huang, W.; et al. The Arabidopsis Information Resource (TAIR): A comprehensive database and web-based information retrieval, analysis, and visualization system for a model plant. Nucleic Acids Res. 2001, 29, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Vogel, J.P.; Garvin, D.F.; Mockler, T.C.; Schmutz, J.; Rokhsar, D.; Bevan, M.W.; Barry, K.; Lucas, S.; Harmon-Smith, M.; Lail, K.; et al. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef]
- Yang, J.; Liu, D.; Wang, X.; Ji, C.; Cheng, F.; Liu, B.; Hu, Z.; Chen, S.; Pental, D.; Ju, Y.; et al. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection. Nat. Genet. 2016, 48, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Rousseau-Gueutin, M.; Belser, C.; Da Silva, C.; Richard, G.; Istace, B.; Cruaud, C.; Falentin, C.; Boideau, F.; Boutte, J.; Delourme, R.; et al. Long-read assembly of the Brassica napus reference genome Darmor-bzh. GigaScience 2020, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Perumal, S.; Koh, C.S.; Jin, L.; Buchwaldt, M.; Higgins, E.; Zheng, C.; Sankoff, D.; Robinson, S.J.; Kagale, S.; Navabi, Z.-K.; et al. High contiguity long read assembly of Brassica nigra allows localization of active centromeres and provides insights into the ancestral Brassica genome. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Cai, X.; Wu, J.; Liu, M.; Grob, S.; Cheng, F.; Liang, J.; Cai, C.; Liu, Z.; Liu, B.; et al. Improved Brassica rapa reference genome by single-molecule sequencing and chromosome conformation capture technologies. Hortic. Res. 2018, 5, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Wu, J.; Liang, J.; Lin, R.; Zhang, K.; Cheng, F.; Wang, X. Improved Brassica oleracea JZS assembly reveals significant changing of LTR-RT dynamics in different morphotypes. Theor. Appl. Genet. 2020, 133, 3187–3199. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.X.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.J.; Thelen, J.J.; Cheng, J.L.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valliyodan, B.; Cannon, S.B.; Bayer, P.E.; Shu, S.Q.; Brown, A.V.; Ren, L.H.; Jenkins, J.; Chung, C.Y.L.; Chan, T.F.; Daum, C.G.; et al. Construction and comparison of three reference-quality genome assemblies for soybean. Plant J. 2019, 100, 1066–1082. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Hirakawa, H.; Isobe, S.; Fukai, E.; Watanabe, A.; Kato, M.; Kawashima, K.; Minami, C.; Muraki, A.; Nakazaki, N.; et al. Sequence analysis of the genome of an oil-bearing tree, Jatropha curcas L. DNA Res. 2011, 18, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.B.; Krishnakumar, V.; Bidwell, S.; Rosen, B.; Chan, A.N.; Zhou, S.G.; Gentzbittel, L.; Childs, K.L.; Yandell, M.; Gundlach, H.; et al. An improved genome release (version Mt4.0) for the model legume Medicago truncatula. BMC Genom. 2014, 15, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, S.; Zhu, W.; Hamilton, J.; Lin, H.; Campbell, M.; Childs, K.; Thibaud-Nissen, F.; Malek, R.L.; Lee, Y.; Zheng, L.; et al. The TIGR Rice Genome Annotation Resource: Improvements and new features. Nucleic Acids Res. 2007, 35, D883–D887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft genome sequence of the oilseed species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Wu, D.; Yang, T.; Sun, C.; Wang, Z.; Han, B.; Wu, S.; Yu, A.; Chapman, M.A.; Muraguri, S.; et al. Genomic insights into the origin, domestication and genetic basis of agronomic traits of castor bean. Genome Biol. 2021, 22, 113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.Y.; Liu, X.; Quan, Z.W.; Cheng, S.F.; Xu, X.; Pan, S.K.; Xie, M.; Zeng, P.; Yue, Z.; Wang, W.L.; et al. Genome sequence of foxtail millet (Setaria italica) provides insights into grass evolution and biofuel potential. Nat. Biotechnol. 2012, 30, 549–554. [Google Scholar] [CrossRef] [Green Version]
- Bennetzen, J.L.; Schmutz, J.; Wang, H.; Percifield, R.; Hawkins, J.; Pontaroli, A.C.; Estep, M.; Feng, L.; Vaughn, J.N.; Grimwood, J.; et al. Reference genome sequence of the model plant Setaria. Nat. Biotechnol. 2012, 30, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornowski, N.; Michel, K.J.; Hamilton, J.P.; Ou, S.J.; Seetharam, A.S.; Jenkins, J.; Grimwood, J.; Plott, C.; Shu, S.Q.; Talag, J.; et al. Genomic variation within the maize stiff-stalk heterotic germplasm pool. Plant Genome 2021. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Arabidopsis thaliana | Brassica napus | Brassica rapa | Brassica oleracea | Brassica nigra | Brassica juncea | Brassica carinata | |
|---|---|---|---|---|---|---|---|
| Brassica_napus_v4.1 | ZS11 | Brara_Chiifu_V3.0 | Braol_JZS_V2.0 | Brani_Ni100_V2 | Braju_tum_V1.5 | ||
| AtDGAT1 At2G19450 | BnaA07g36000D | BnaA07G0011800ZS | BraA07g001370.3C | — | — | BjuA046403-A07 | — |
| BnaAnng30990D | BnaA09G0121200ZS | BraA09g011830.3C | — | — | BjuA017169-A09 | — | |
| — | — | — | — | BniB03g068710.2N | BjuB029654-B03 | BcaB03g16730 | |
| — | — | — | — | BniB04g025020.2N | BjuB028615-B04 | BcaB07g31608 | |
| BnaCnng52810D | BnaC07G0026200ZS | — | BolC07g002350.2J | — | — | BcaC04g22852 | |
| — | BnaC09G0126800ZS | — | BolC09g013760.2J | — | — | BcaC06g36576 | |
| AtDGAT2 At3G51520 | BnaA01g19390D | BnaA01G0206700ZS | BraA01g022340.3C | — | — | BjuA005097-A01 | — |
| BnaA03g41350D | BnaA03G0420700ZS | BraA03g045590.3C | — | — | BjuA042315-A03 | — | |
| — | — | — | — | BniB02g074660.2N | BjuB048735-B02 | BcaC06g33029 | |
| — | — | — | — | BniB05g021740.2N | BjuB013221-B05 | BcaNung00379 | |
| BnaC01g23350D | BnaC01G0259700ZS | — | BolC01g027430.2J | — | — | — | |
| BnaC07g32270D | BnaC07G0393500ZS | — | BolC07g044710.2J | — | — | — | |
| AtDGAT3 At1G48300 | BnaA08g03400D | BnaA08G0039500ZS | BraA08g004520.3C | — | — | BjuA014363-A08 | — |
| — | — | — | — | BniB07g007860.2N | BjuB045147-B07 | BcaB04g17451 | |
| BnaC08g46660D | BnaC08G0049200ZS | — | BolC08g005290.2J | — | — | BcaC07g37153 | |
| Scheme | Gene Name | Gene Locus ID | Chromosome Location | Protein | Putative Promoter | |||
|---|---|---|---|---|---|---|---|---|
| AA | pI | Mw (Da) | Subcellular Location | |||||
| DGAT1 | BnaA.DGAT1.a | BnaA07G0011800ZS | A07:935,937-939,473 | 504 | 8.75 | 57,743.80 | E.R. | 1759 bp |
| BnaA.DGAT1.b | BnaA09G0121200ZS | A09:7,271,130-7,274,477 | 503 | 8.09 | 57,241.36 | E.R. | 1488 bp | |
| BnaC.DGAT1.a | BnaC07G0026200ZS | C07:4,705,347-4,708,653 | 501 | 8.43 | 57,538.87 | E.R. | 1657 bp | |
| BnaC.DGAT1.b | BnaC09G0126800ZS | C09:9,270,137-9,273,644 | 510 | 8.20 | 57,958.08 | E.R. | 1500 bp | |
| DGAT2 | BnaA.DGAT2.a | BnaA01G0206700ZS | A01:12,907,146-12,908,853 | 368 | 8.77 | 41,813.30 | E.R. | 836 bp |
| BnaA.DGAT2.b | BnaA03G0420700ZS | A03:22,826,130-22,827,759 | 367 | 8.32 | 41,280.40 | E.R. | 775 bp | |
| BnaC.DGAT2.a | BnaC01G0259700ZS | C01:20,695,151-20,696,870 | 319 | 8.88 | 36,103.24 | E.R. | 2151 bp | |
| BnaC.DGAT2.b | BnaC07G0393500ZS | C07:51,188,778-51,190,546 | 317 | 7.75 | 35,636.69 | E.R. | 1973 bp | |
| DGAT3 | BnaA.DGAT3 | BnaA08G0039500ZS | A08:3,290,776-3,291,925 | 356 | 8.90 | 38,476.61 | None | 1660 bp |
| BnaA.DGAT3 | BnaC08G0049200ZS | C08:4,709,675-4,710,823 | 356 | 8.84 | 38,184.10 | None | 1459 bp | |
| Gene_1 | Gene_2 | Ka | Ks | Ka/Ks | Duplication Time (MYA) | Average (MYA) | |
|---|---|---|---|---|---|---|---|
| Orthologous gene pairs | AtDGAT1 | BnaA.DGAT1.a | 0.062 | 0.320 | 0.194 | 10.67 | 15.06 |
| AtDGAT1 | BnaA.DGAT1.b | 0.064 | 0.400 | 0.161 | 13.33 | ||
| AtDGAT1 | BnaC.DGAT1.a | 0.064 | 0.347 | 0.184 | 11.56 | ||
| AtDGAT1 | BnaC.DGAT1.b | 0.060 | 0.415 | 0.145 | 13.82 | ||
| AtDGAT2 | BnaA.DGAT2.a | 0.098 | 0.486 | 0.201 | 16.21 | ||
| AtDGAT2 | BnaA.DGAT2.b | 0.123 | 0.448 | 0.275 | 14.92 | ||
| AtDGAT2 | BnaC.DGAT2.a | 0.099 | 0.495 | 0.200 | 16.51 | ||
| AtDGAT2 | BnaC.DGAT2.b | 0.124 | 0.436 | 0.284 | 14.55 | ||
| AtDGAT3 | BnaA.DGAT3 | 0.081 | 0.568 | 0.143 | 18.92 | ||
| AtDGAT3 | BnaC.DGAT3 | 0.086 | 0.603 | 0.142 | 20.11 | ||
| Paralogous gene pairs | BnaA.DGAT1.a | BnaA.DGAT1.b | 0.056 | 0.346 | 0.161 | 11.52 | 8.36 |
| BnaA.DGAT1.a | BnaC.DGAT1.a | 0.003 | 0.041 | 0.064 | 1.37 | ||
| BnaA.DGAT1.a | BnaC.DGAT1.b | 0.054 | 0.329 | 0.163 | 10.97 | ||
| BnaA.DGAT1.b | BnaC.DGAT1.a | 0.058 | 0.358 | 0.162 | 11.92 | ||
| BnaA.DGAT1.b | BnaC.DGAT1.b | 0.007 | 0.097 | 0.073 | 3.22 | ||
| BnaC.DGAT1.a | BnaC.DGAT1.b | 0.056 | 0.328 | 0.170 | 10.95 | ||
| BnaA.DGAT2.a | BnaA.DGAT2.b | 0.111 | 0.347 | 0.321 | 11.58 | ||
| BnaA.DGAT2.a | BnaC.DGAT2.a | 0.009 | 0.085 | 0.107 | 2.83 | ||
| BnaA.DGAT2.a | BnaC.DGAT2.b | 0.112 | 0.372 | 0.301 | 12.41 | ||
| BnaA.DGAT2.b | BnaC.DGAT2.a | 0.111 | 0.376 | 0.296 | 12.52 | ||
| BnaA.DGAT2.b | BnaC.DGAT2.b | 0.010 | 0.092 | 0.107 | 3.07 | ||
| BnaC.DGAT2.a | BnaC.DGAT2.b | 0.111 | 0.383 | 0.290 | 12.78 | ||
| BnaA.DGAT3 | BnaC.DGAT3 | 0.019 | 0.107 | 0.179 | 3.57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, X.; Guo, X.; Hu, L.; Li, S.; Chen, Y.; Wang, J.; Wang, R.R.-C.; Fan, C.; Hu, Z. Genome-Wide Characterization of DGATs and Their Expression Diversity Analysis in Response to Abiotic Stresses in Brassica napus. Plants 2022, 11, 1156. https://doi.org/10.3390/plants11091156
Yin X, Guo X, Hu L, Li S, Chen Y, Wang J, Wang RR-C, Fan C, Hu Z. Genome-Wide Characterization of DGATs and Their Expression Diversity Analysis in Response to Abiotic Stresses in Brassica napus. Plants. 2022; 11(9):1156. https://doi.org/10.3390/plants11091156
Chicago/Turabian StyleYin, Xiangzhen, Xupeng Guo, Lizong Hu, Shuangshuang Li, Yuhong Chen, Jingqiao Wang, Richard R.-C. Wang, Chengming Fan, and Zanmin Hu. 2022. "Genome-Wide Characterization of DGATs and Their Expression Diversity Analysis in Response to Abiotic Stresses in Brassica napus" Plants 11, no. 9: 1156. https://doi.org/10.3390/plants11091156