
Altered Developmental and Metabolic Gene Expression in Basil Interspecific Hybrids

 ,
,
Abstract
:1. Introduction
2. Results
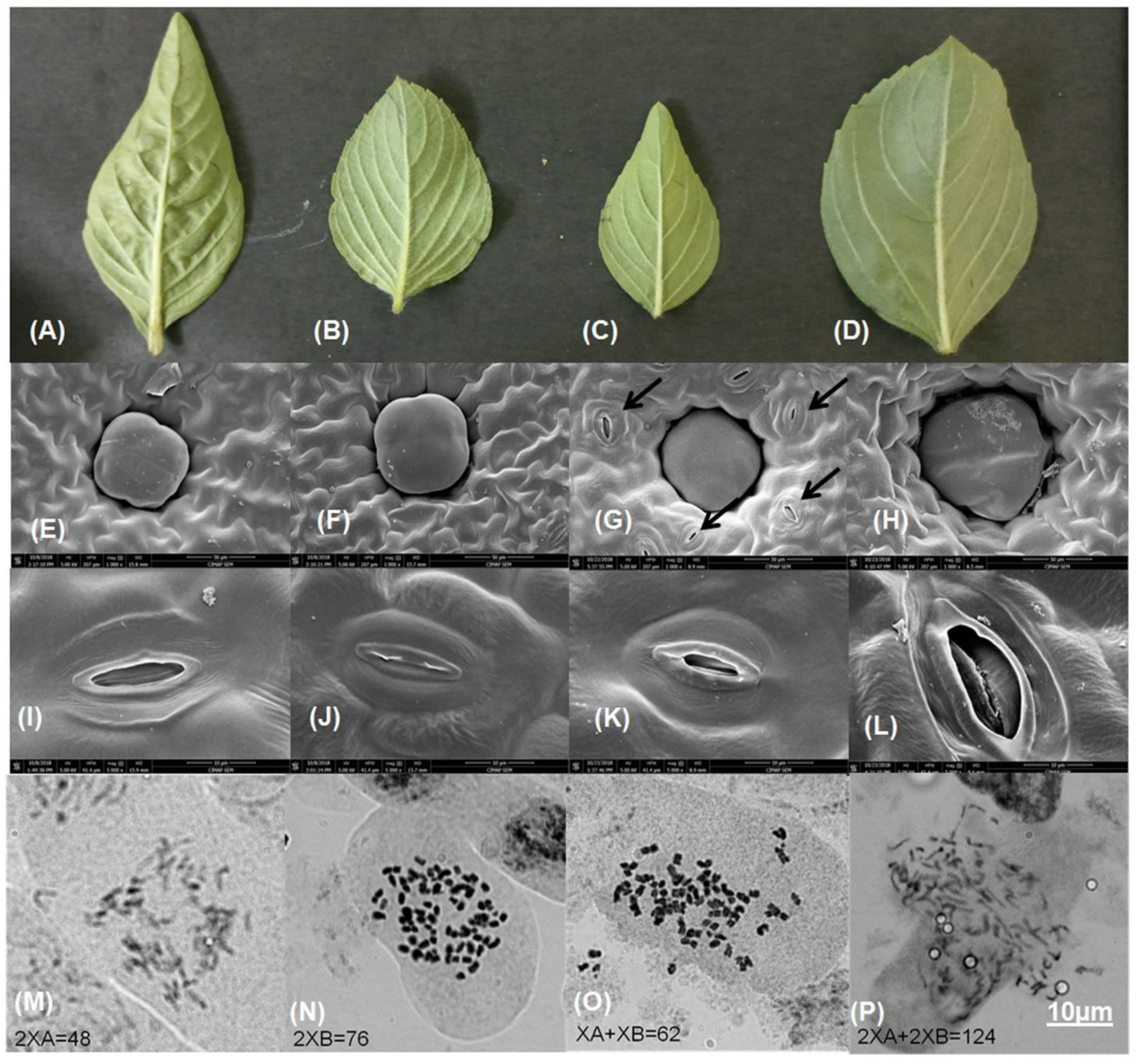
2.1. Comparative Phenotype
2.2. Chromosome Number
2.3. Transcriptome Sequencing, Assembly and Annotation
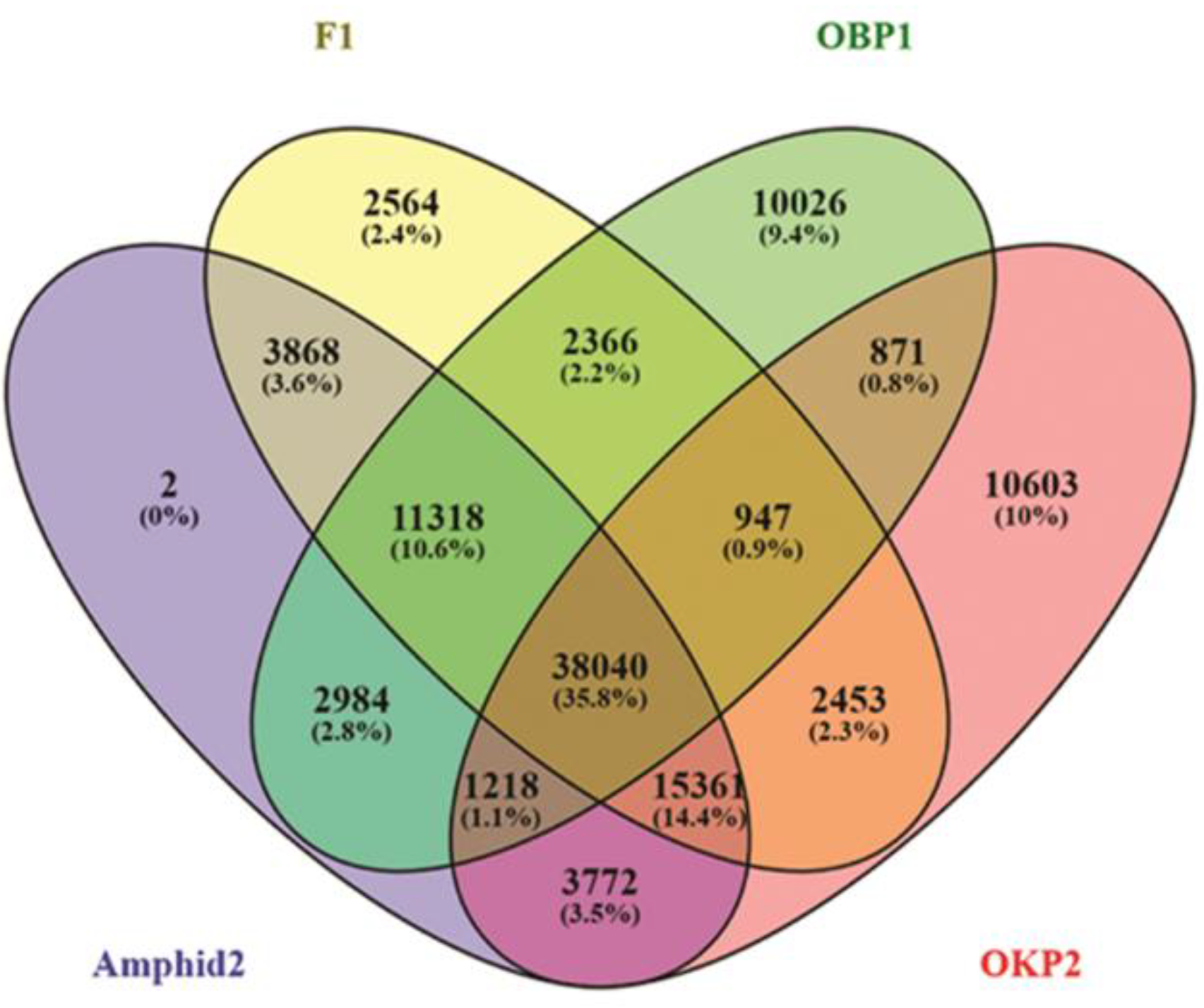
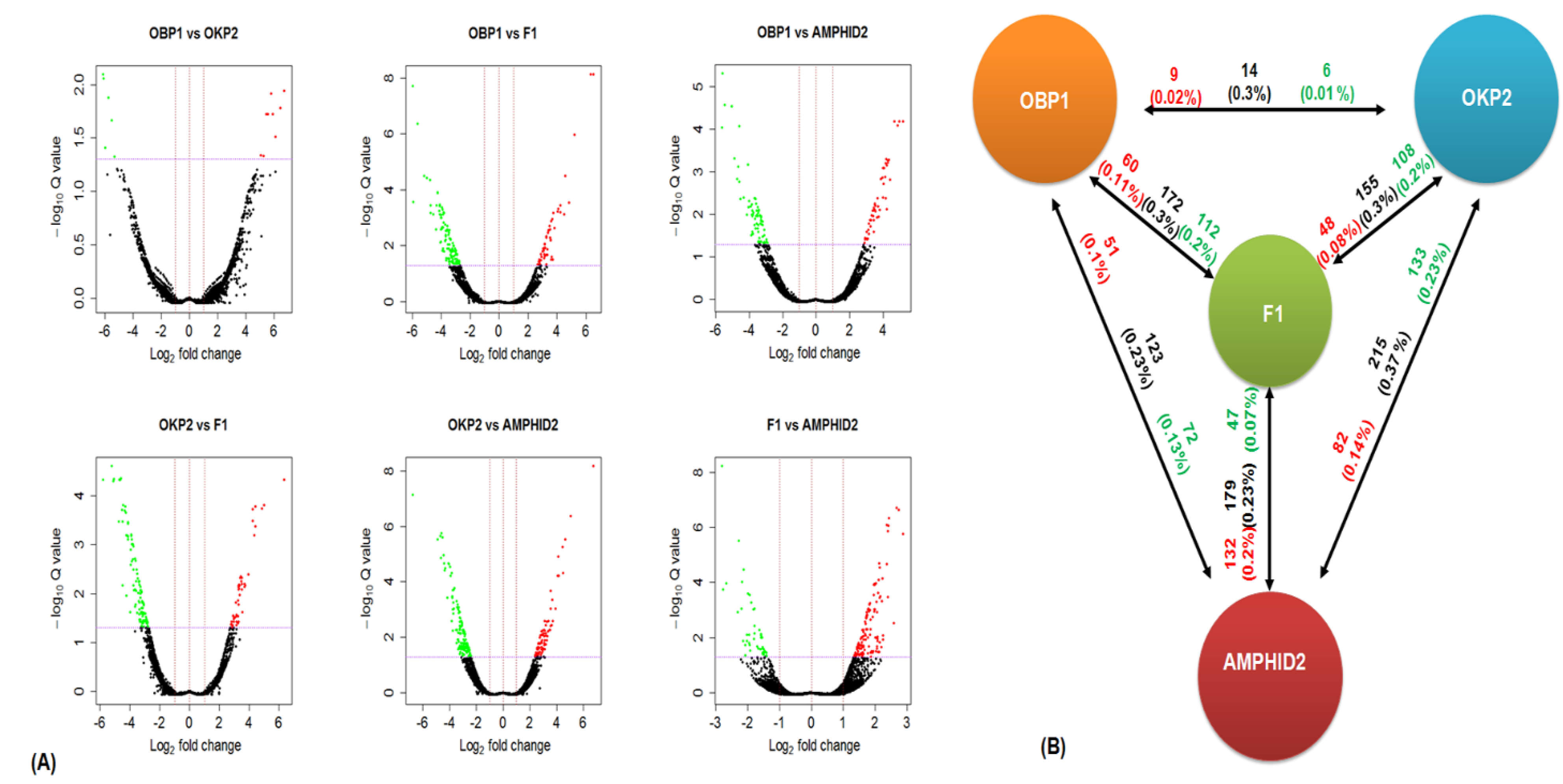
2.4. Detection of Gene Expression Alterations in Interspecific Hybrid F1 and Amphidiploid
2.5. Comparative Gene Expression Patterns
2.6. DEGs Co-Regulated
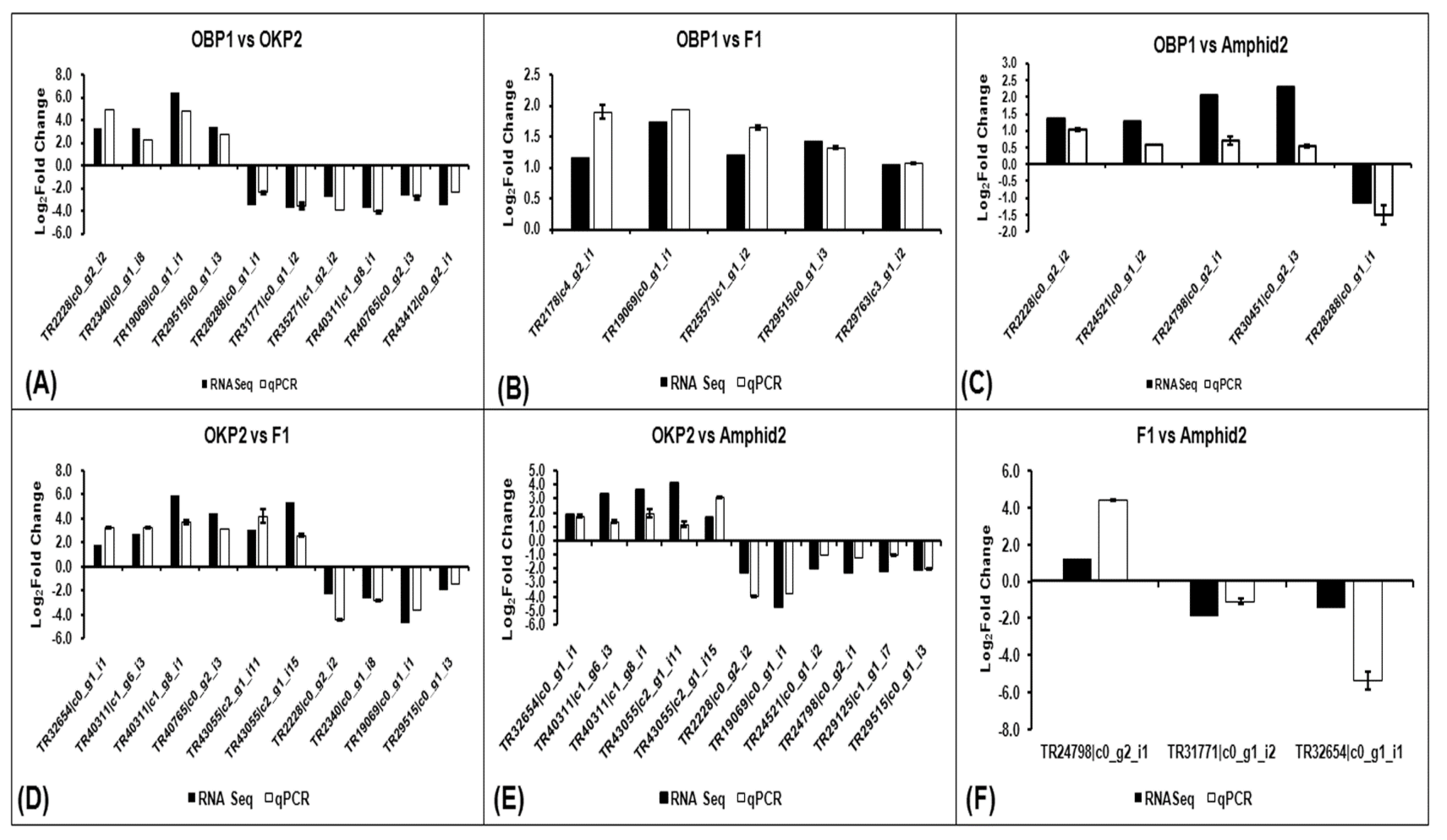
2.7. Verification of RNA-Seq Gene Expression Data
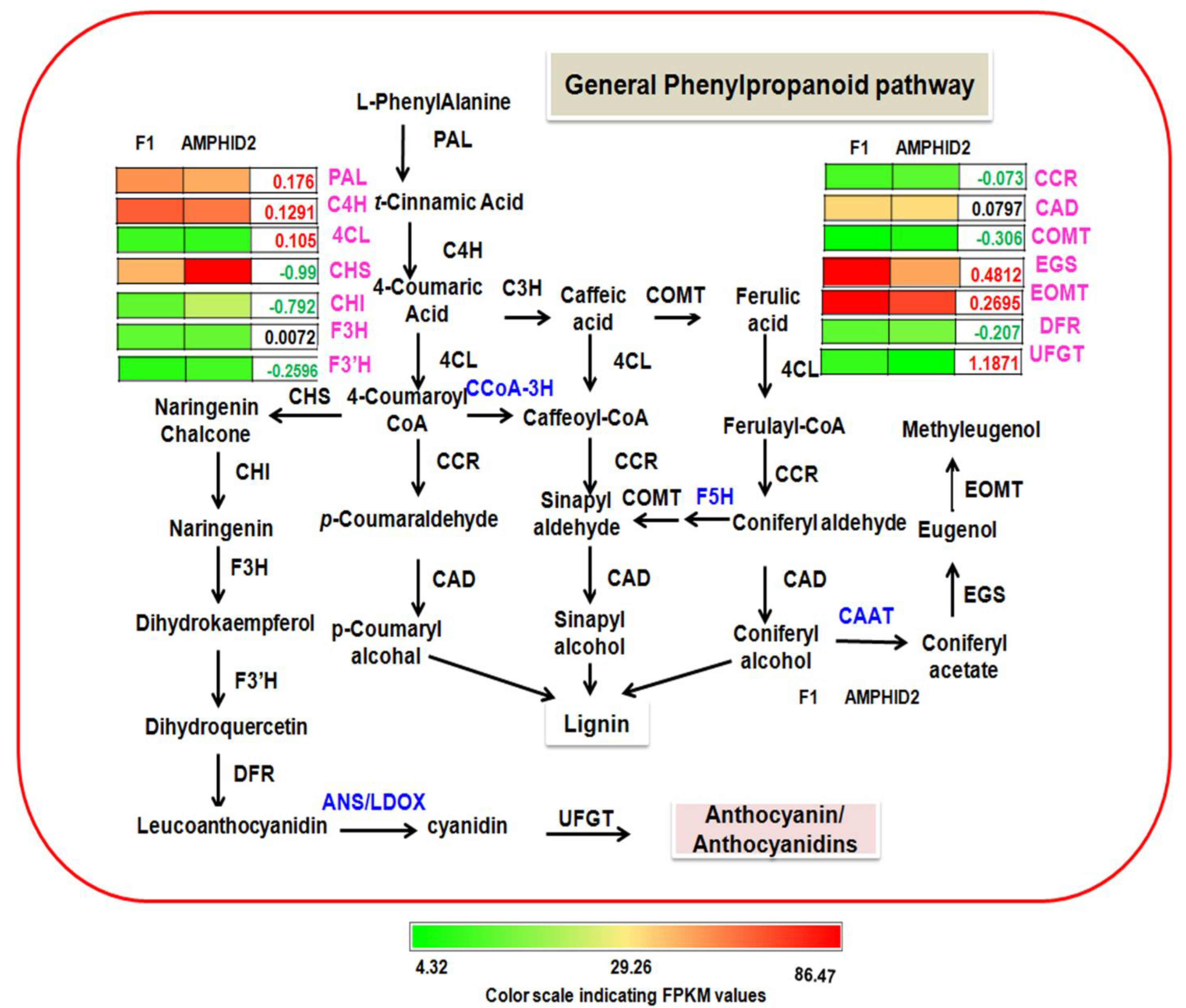
2.8. General Phenylpropanoid Pathway in F1 and Amphidiploid
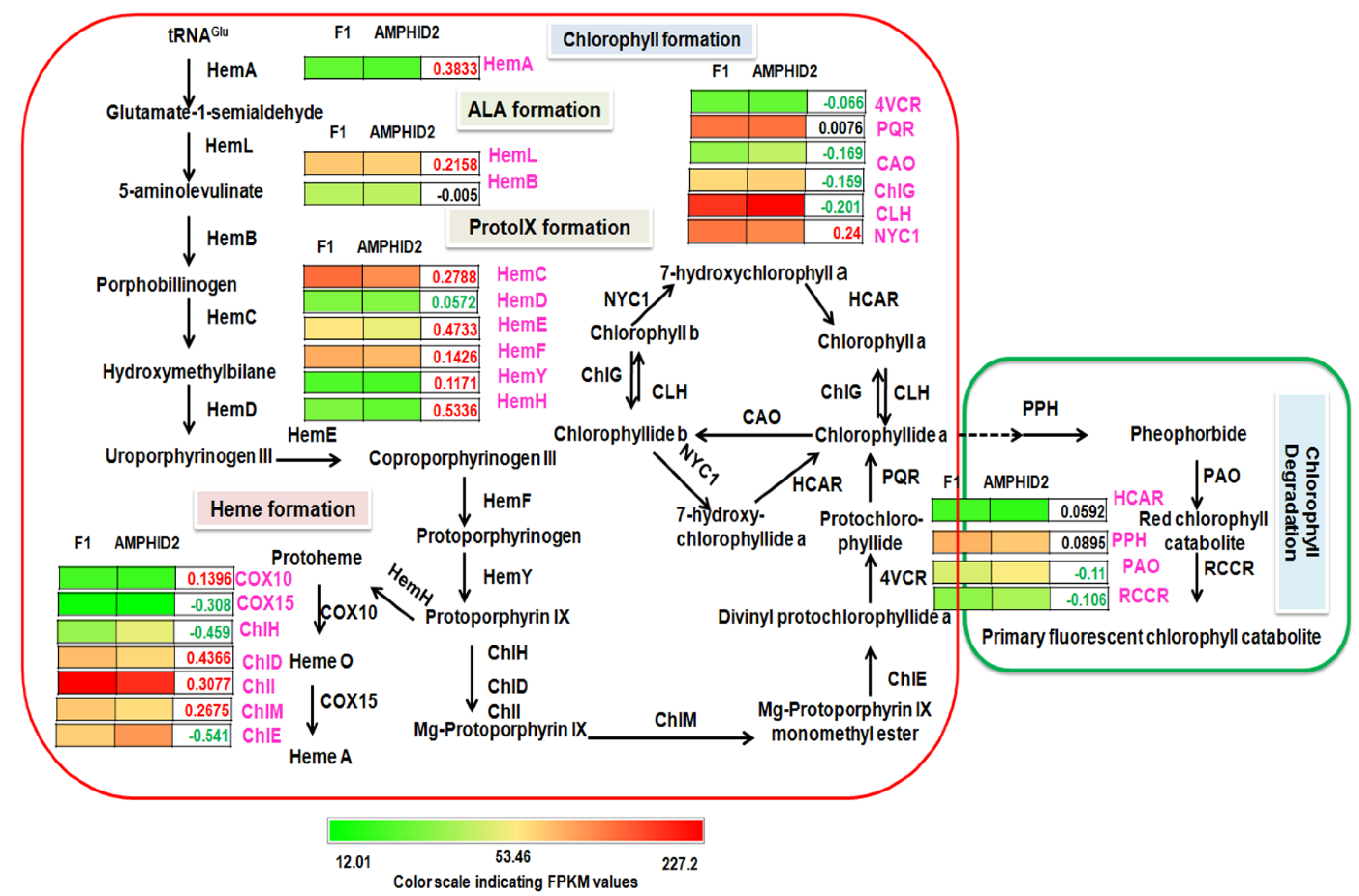
2.9. Chlorophyll Metabolism in F1 Hybrid and Amphidiploid
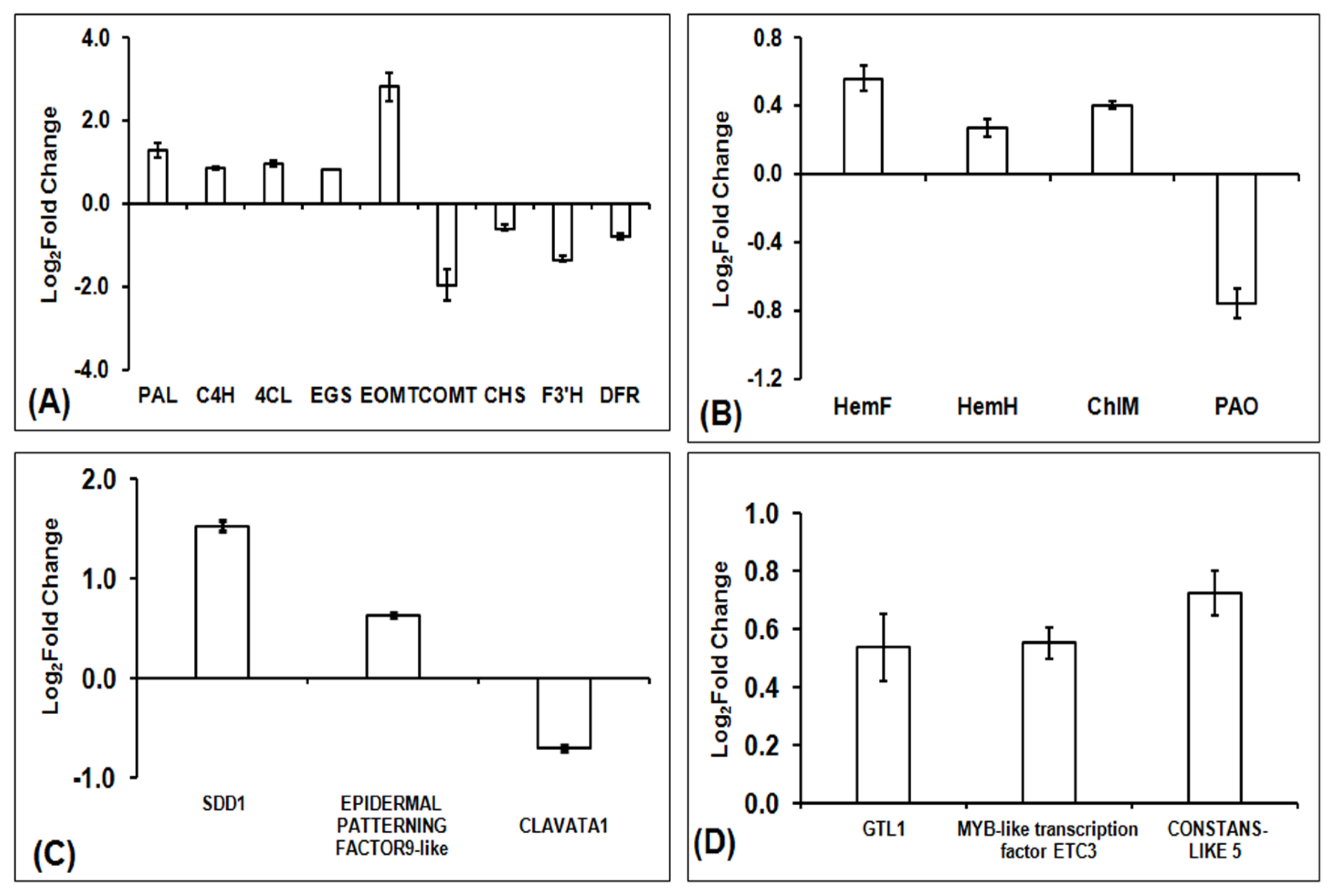
2.10. Differentially Expressed Stomatal Patterning and Development Genes
2.11. Transcription Factors
3. Discussion
3.1. Gene Silencing and Activation in Interspecific Hybrid F1 and Amphidiploids
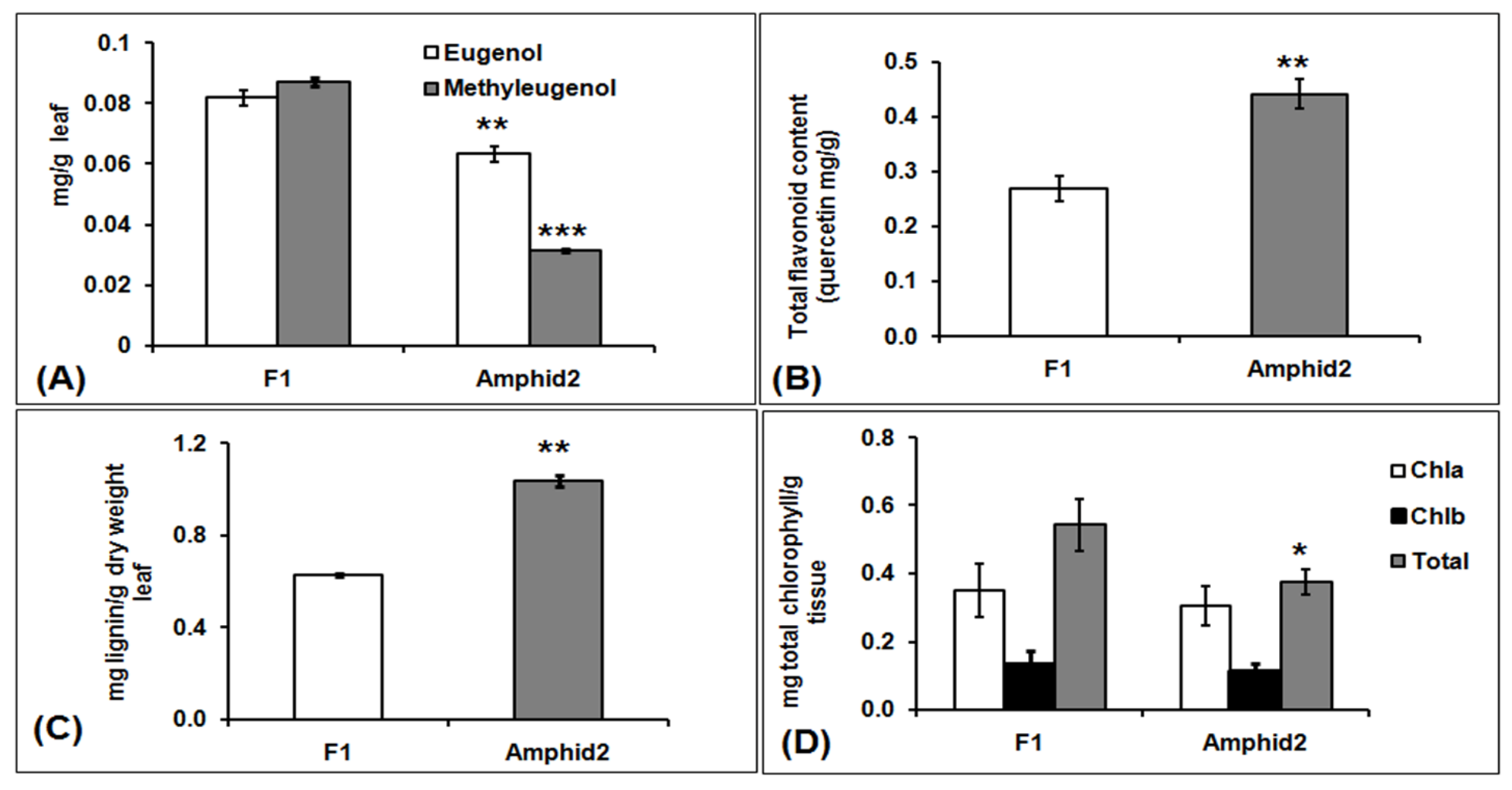
3.2. Higher Amount of Eugenol and Methyl Eugenol in Interspecific Hybrid F1 Is Possibly Associated with Reduced Lignin and Flavonoid Biosynthesis
3.3. Enhanced Chlorophyll Biosynthesis in Interspecific Hybrid F1
3.4. Effect of Hybridization on Stomatal Patterning and Development Associated Genes
3.5. Differentially Expressed Transcription Factors
4. Materials and Methods
4.1. Plant Materials
4.2. Phenotypic Changes and Their Statistical Analysis
4.3. Scanning Electron Microscopy
4.4. Chromosome Count
4.5. Total RNA Isolation and Library Preparation
4.6. De Novo Assembly and Functional Annotation
4.7. Differential Gene Expression (DGE) Analysis
4.8. Quantitative RT-PCR for Gene Expression Analysis
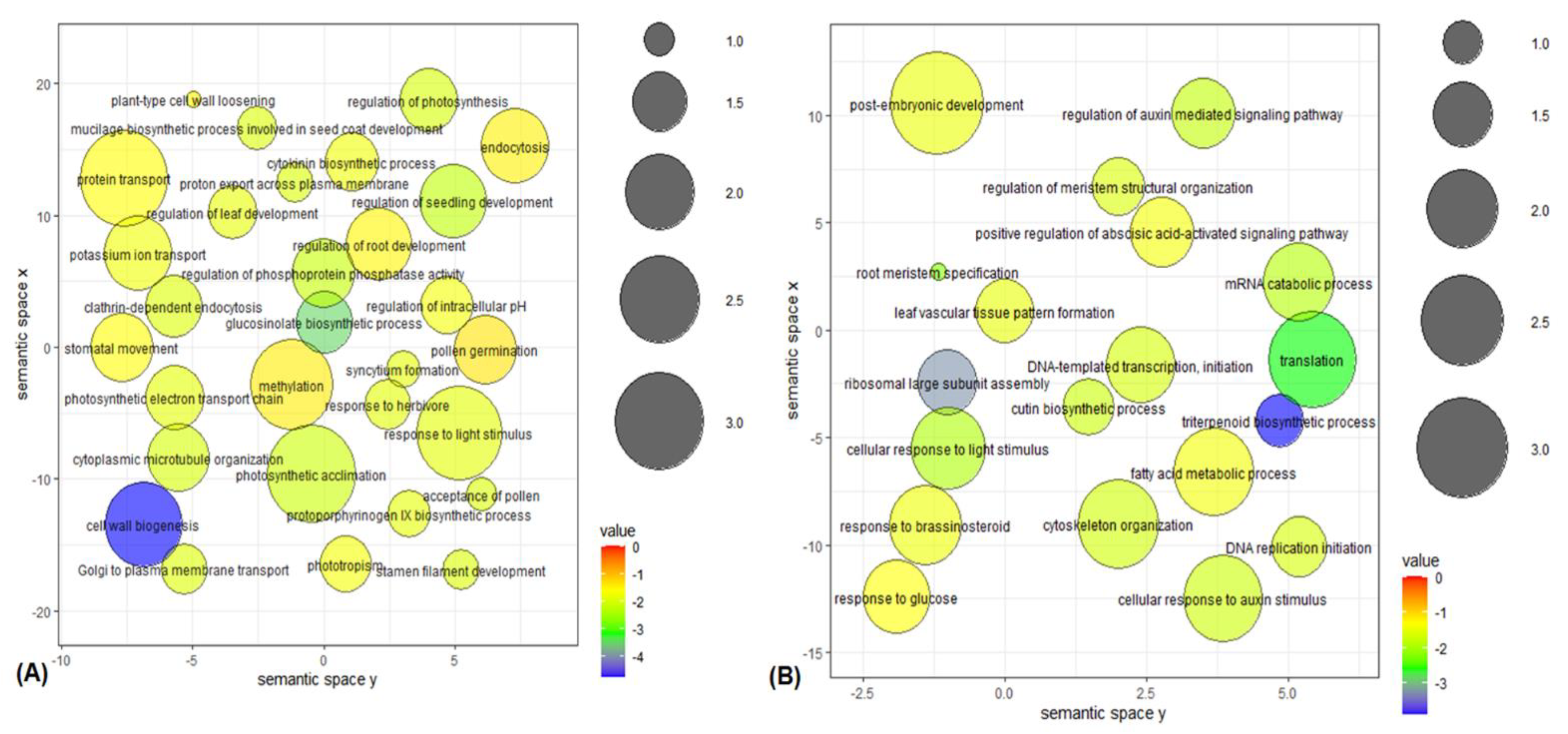
4.9. Gene Ontology Enrichment Analysis
4.10. Oil Analysis
4.11. Biochemical Assays for Estimation of Total Chlorophyll, Total Lignin and Total Flavonoid Contents
4.12. Statistical Measurement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiao, Y.; Wickett, N.J.; Ayyampalayam, S.; Chanderbali, A.S.; Landherr, L.; Ralph, P.E.; Tomsho, L.P.; Hu, Y.; Liang, H.; Soltis, P.S.; et al. Ancestral polyploidy in seed plants and angiosperms. Nature 2011, 473, 97–100. [Google Scholar] [CrossRef]
- Wendel, J.F. The wondrous cycles of polyploidy in plants. Am. J. Bot. 2015, 102, 1753–1756. [Google Scholar] [CrossRef] [Green Version]
- Wendel, J.F.; Lisch, D.; Hu, G.; Mason, A.S. The long and short of doubling down: Polyploidy, epigenetics, and the temporal dynamics of genome fractionation. Curr. Opin. Genet. Dev. 2018, 49, 1–7. [Google Scholar] [CrossRef]
- Wood, T.E.; Takebayashi, N.; Barker, M.S.; Mayrose, I.; Greenspoon, P.B.; Rieseberg, L.H. The frequency of polyploid speciation in vascular plants. Proc. Natl. Acad. Sci. USA 2009, 106, 13875–13879. [Google Scholar] [CrossRef] [Green Version]
- Scarpino, S.V.; Levin, D.A.; Meyers, L.A. Polyploid formation shapes flowering plant diversity. Am. Nat. 2014, 184, 456–465. [Google Scholar] [CrossRef] [Green Version]
- Baker, R.L.; Yarkhunova, Y.; Vidal, K.; Ewers, B.E.; Weinig, C. Polyploidy and the relationship between leaf structure and function: Implications for correlated evolution of anatomy, morphology, and physiology in Brassica. BMC Plant Biol. 2017, 17, 3. [Google Scholar] [CrossRef] [Green Version]
- Van de Peer, Y.; Mizrachi, E.; Marchal, K. The evolutionary significance of polyploidy. Nat. Rev. Genet. 2017, 18, 411. [Google Scholar] [CrossRef]
- Levin, D.A. Plant speciation in the age of climate change. Ann. Bot. 2019, 124, 769–775. [Google Scholar] [CrossRef] [Green Version]
- Van de Peer, Y.; Ashman, T.-L.; Soltis, P.S.; Soltis, D.E. Polyploidy: An evolutionary and ecological force in stressful times. Plant Cell 2021, 33, 11–26. [Google Scholar] [CrossRef]
- Fasano, C.; Diretto, G.; Aversano, R.; D’Agostino, N.; Di Matteo, A.; Frusciante, L.; Giuliano, G.; Carput, D. Transcriptome and metabolome of synthetic Solanum autotetraploids reveal key genomic stress events following polyploidization. New Phytol. 2016, 210, 1382–1394. [Google Scholar] [CrossRef] [Green Version]
- Tan, F.Q.; Tu, H.; Wang, R.; Wu, X.M.; Xie, K.D.; Chen, J.J.; Zhang, H.Y.; Xu, J.; Guo, W.W. Metabolic adaptation following genome doubling in citrus doubled diploids revealed by non-targeted metabolomics. Metabolomics 2017, 13, 143. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. Ancient WGD events as drivers of key innovations in angiosperms. Curr. Opin. Plant Biol. 2016, 30, 159–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.J.; Coate, J.E. Polyploidy, the nucleotype, and novelty: The impact of genome doubling on the biology of the cell. Int. J. Plant Sci. 2019, 180, 1–52. [Google Scholar] [CrossRef]
- Levin, D.A. The role of chromosomal change in plant evolution. In Oxford Series in Ecology and Evolution; Oxford University Press: New York, NY, USA, 2002; Volume 79, pp. 460–461. ISBN 0-19-513859-7. [Google Scholar]
- Schoenfelder, K.P.; Fox, D.T. The expanding implications of polyploidy. J. Cell Biol. 2015, 209, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Vimala, Y.; Lavania, U.C.; Singh, M.; Lavania, S.; Srivastava, S.; Basu, S. Realization of lodging tolerance in the aromatic grass, Cymbopogon khasianus through ploidy intervention. Front. Plant Sci. 2022, 13, 908659. [Google Scholar] [CrossRef]
- Lavania, U.C. Genomic and ploidy manipulation for enhanced production of phyto-pharmaceuticals. Plant Genet. Resour. 2005, 3, 170–177. [Google Scholar] [CrossRef]
- Mishra, M.K. Stomatal Characteristics at Different Ploidy Levels in Coffea L. Ann. Bot. 1997, 80, 689–692. [Google Scholar] [CrossRef] [Green Version]
- Lavania, U.C.; Srivastava, S.; Lavania, S.; Basu, S.; Misra, N.K.; Mukai, Y. Autopolyploidy differentially influences body size in plants, but facilitates enhanced accumulation of secondary metabolites, causing increased cytosine methylation. Plant J. 2012, 71, 539–549. [Google Scholar] [CrossRef]
- Riddle, N.C.; Kato, A.; Birchler, J.A. Genetic variation for the response to ploidy change in Zea mays L. Theor. Appl. Genet. 2007, 114, 101–111. [Google Scholar] [CrossRef]
- Pandit, M.K.; Pocock, M.J.O.; Kunin, W.E. Ploidy influences rarity and invasiveness in plants. J. Ecol. 2011, 99, 1108–1115. [Google Scholar] [CrossRef]
- Shasany, A.K.; Kole, C. Genomic Resources of Ocimum. In The Ocimum Genome; Springer: Cham, Switzerland, 2018; pp. 99–110. ISBN 978-3-319-97430-9. [Google Scholar]
- Vina, A.; Murillo, E. Essential oil composition from twelve varieties of basil (Ocimum spp.) grown in Colombia. J. Braz. Chem. Soc. 2003, 14, 744–749. [Google Scholar] [CrossRef]
- Jin, J.P.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.C.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, 1040–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaked, H.; Kashkush, K.; Ozkan, H.; Feldman, M.; Levy, A.A. Sequence elimination and cytosine methylation are rapid and reproducible responses of the genome to wide hybridization and allopolyploidy in wheat. Plant Cell 2001, 13, 1749–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozkan, H.; Levy, A.A.; Feldman, M. Allopolyploidy-induced rapid genome evolution in the wheat (Aegilops-Triticum) group. Plant Cell 2001, 13, 1735–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comai, L.; Tyagi, A.P.; Winter, K.; Holmes-Davis, R.; Reynolds, H. Phenotypic instability and rapid gene silencing in newly formed Arabidopsis allotetraploids. Plant Cell 2000, 12, 1551–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashkush, K.; Feldman, M.; Levy, A.A. Gene loss, silencing, and activation in a newly synthesized wheat allotetraploid. Genetics 2002, 160, 1651–1659. [Google Scholar] [CrossRef]
- Galitski, T.; Saldanha, A.J.; Styles, C.A.; Lander, E.S.; Fink, G.R. Ploidy regulation of gene expression. Science 1999, 285, 251–254. [Google Scholar] [CrossRef]
- Yao, T.; Feng, K.; Xie, M.; Barros, J.; Tschaplinski, T.J.; Tuskan, G.A.; Muchero, W.; Chen, J.G. Phylogenetic occurrence of the phenylpropanoid pathway and lignin biosynthesis in plants. Front. Plant Sci. 2021, 12, 704697. [Google Scholar] [CrossRef]
- Fraser, C.M.; Chapple, C. The Phenylpropanoid Pathway in Arabidopsis. Arab. Book 2011, 9, e0152. [Google Scholar] [CrossRef] [Green Version]
- Baxter, H.L.; Stewart, C.N., Jr. Effects of altered lignin biosynthesis on phenylpropanoid metabolism and plant stress. Biofuels 2013, 4, 635–650. [Google Scholar] [CrossRef]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Muhlemann, J.K.; Woodworth, B.D.; Morgan, J.A.; Dudareva, N. The monolignol pathway contributes to the biosynthesis of volatile phenylpropenes in flowers. New Phytol. 2014, 204, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, S.; Kumar, R.; Chanotiya, C.S.; Shanker, K.; Gupta, M.M.; Nagegowda, D.A.; Shasany, A.K. 4-Coumarate: CoA ligase partitions metabolites for eugenol biosynthesis. Plant Cell Physiol. 2013, 54, 1238–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuk, M.; Działo, M.; Richter, D.; Dymińska, L.; Matuła, J.; Kotecki, A.; Hanuza, J.; Szopa, J. Chalcone synthase (CHS) gene suppression in flax leads to changes in wall synthesis and sensing genes, cell wall chemistry and stem morphology parameters. Front. Plant Sci. 2016, 7, 894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molin, W.T.; Meyers, S.P.; Baer, G.R.; Schrader, L.E. Ploidy effects in isogenic populations of alfalfa: II. Photosynthesis, chloroplast number, ribulose-1, 5-bisphosphate carboxylase, chlorophyll, and DNA in protoplasts. Plant Physiol. 1982, 70, 1710–1714. [Google Scholar] [CrossRef] [Green Version]
- Meyers, S.P.; Nichols, S.L.; Baer, G.R.; Molin, W.T.; Schrader, L.E. Ploidy effects in isogenic populations of alfalfa. I. Ribulose-1, 5-biphosphate-carboxylase, soluble protein, chlorophyll, and DNA in leaves. Plant Physiol. 1982, 70, 1704–1709. [Google Scholar] [CrossRef] [Green Version]
- Leech, R.M.; Leese, B.M.; Jellings, A.J. Variation in cellular ribulose-1, 5-biphosphate-carboxylase content in leaves of Triticum genotypes at three levels of ploidy. Planta 1985, 166, 259–263. [Google Scholar] [CrossRef]
- Warner, D.A.; Edwards, G.E. Effects of polyploidy on photosynthetic rates, photosynthetic enzymes, contents of DNA, chlorophyll, and sizes and numbers of photosynthetic cells in the C4 dicot Atriplex confertifolia. Plant Physiol. 2007, 91, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Timko, M.P.; Vasconcelos, A.C. Euploidy in Ricinus. Plant Physiol. 1981, 67, 1084–1089. [Google Scholar] [CrossRef] [Green Version]
- Padoan, D.; Mossad, A.; Chiancone, B.; Germana, M.A.; Khan, P.S.S.V. Ploidy levels in Citrus clementine affects leaf morphology, stomatal density and water content. Theor. Exp. Plant Physiol. 2013, 25, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Casson, S.; Gray, J.E. Influence of environmental factors on stomatal development. New Phytol. 2008, 178, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, D.C.; Lukowitz, W.; Somerville, C.R. Stomatal development and pattern controlled by a MAPKK kinase. Science 2004, 304, 1494–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Ngwenyama, N.; Liu, Y.; Walker, J.C.; Zhang, S. Stomatal development and patterning are regulated by environmentally responsive mitogen-activated protein kinases in Arabidopsis. Plant Cell 2007, 19, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coupe, S.A.; Palmer, B.G.; Lake, J.A.; Overy, S.A.; Oxborough, K.; Woodward, F.I.; Gray, J.E.; Quick, W.P. Systemic signaling of environmental cues in Arabidopsis leaves. J. Exp. Bot. 2006, 57, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, S.; Hiratsuka, K.; Chua, N.H. Transcription factors in plant growth and development. Curr. Opin. Genet. Dev. 1994, 4, 642–646. [Google Scholar] [CrossRef]
- Rounsley, S.D.; Ditta, G.S.; Yanofsky, M.F. Diverse roles for MADS box genes in Arabidopsis development. Plant Cell 1995, 7, 1259–1269. [Google Scholar] [CrossRef] [Green Version]
- Mouhu, K.; Kurokura, T.; Koskela, E.A.; Albert, V.A.; Elomaa, P.; Hytönen, T. The Fragaria vesca homolog of suppressor of overexpression of CONSTANS1 represses flowering and promotes vegetative growth. Plant Cell 2013, 25, 3296–3310. [Google Scholar] [CrossRef] [Green Version]
- Putterill, J.; Robson, F.; Lee, K.; Simon, R.; Coupland, G. The CONSTANS gene of Arabidopsis promotes flowering and encodes a protein showing similarities to zinc finger transcription factors. Cell 1995, 80, 847–857. [Google Scholar] [CrossRef] [Green Version]
- Hassidim, M.; Harir, Y.; Yakir, E.; Kron, I.; Green, R.M. Over-expression of CONSTANS-LIKE 5 can induce flowering in short-day grown Arabidopsis. Planta 2009, 230, 481–491. [Google Scholar] [CrossRef]
- Liu, J.; Osbourn, A.; Ma, P. MYB transcription factors as regulators of phenylpropanoid metabolism in plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef] [Green Version]
- Xun, H.; Zhang, Z.; Zhou, Y.; Qian, X.; Dong, Y.; Feng, X.; Pang, J.; Wang, S.; Liu, B. Identification, and functional characterization of R3 MYB transcription factor genes in soybean. Plant J. 2018, 61, 85–96. [Google Scholar] [CrossRef]
- Tominaga, R.; Iwata, M.; Sano, R.; Inoue, K.; Okada, K.; Wada, T. Arabidopsis CAPRICE-LIKE MYB 3 (CPL3) controls endoreduplication and flowering development in addition to trichome and root hair formation. Development 2008, 135, 1335–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.; Zhou, Y.; Han, X.; Shen, C.; Wang, S.; Liu, C.; Yin, W.; Xia, X. The GATA transcription factor GNC plays an important role in photosynthesis and growth in poplar. J. Exp. Bot. 2020, 71, 1969–1984. [Google Scholar] [CrossRef]
- Yoo, C.Y.; Pence, H.E.; Jin, J.B.; Miura, K.; Gosney, M.J.; Hasegawa, P.M.; Mickelbart, M.V. The Arabidopsis GTL1 transcription factor regulates water use efficiency and drought tolerance by modulating stomatal density via transrepression of SDD1. Plant Cell 2010, 22, 4128–4141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breuer, C.; Kawamura, A.; Ichikawa, T.; Tominaga-Wada, R.; Wada, T.; Kondou, Y.; Muto, S.; Matsui, M.; Sugimoto, K. The trihelix transcription factor GTL1 regulates ploidy-dependent cell growth in the Arabidopsis trichome. Plant Cell 2009, 21, 2307–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, A.; Wolfgang, H. Differential Expression of RNA-Seq Data at the Gene Level—The DESeq Package; European Molecular Biology Laboratory (EMBL): Heidelberg, Germany, 2012. [Google Scholar]
- Akhtar, M.Q.; Qamar, N.; Yadav, P.; Kulkarni, P.; Kumar, A.; Shasany, A.K. Comparative Glandular trichome transcriptome-based gene characterization reveals reasons for differential (−)-menthol biosynthesis in Mentha species. Physiol. Plant. 2017, 160, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Vashisth, D.; Misra, A.; Akhtar, M.Q.; Jalil, S.U.; Shanker, K.; Gupta, M.M.; Rout, P.K.; Gupta, A.K.; Shasany, A.K. RNAi down-regulation of cinnamate-4-hydroxylase increases artemisinin biosynthesis in Artemisia annua. Sci. Rep. 2016, 6, 26458. [Google Scholar] [CrossRef] [Green Version]
- Sankhalkar, S.; Vernekar, V. Quantitative and Qualitative analysis of Phenolic and Flavonoid content in Moringa oleifera Lam and Ocimum tenuiflorum L. Pharmacognosy Res. 2016, 8, 16. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | OBP1 | OKP2 | F1 | Amphid2 |
|---|---|---|---|---|
| Plant height (cm) | 87.40 ± 7.80 | 110.00 ± 8.9 *** | 112.70 ± 6.24 *** | 101.50 ± 9.30 ** |
| Leaf area (cm2) | 7.61 ± 0.61 | 6.85 ± 0.45 * | 3.50 ± 0.18 *** | 9.63 ± 0.75 *** |
| Stem diameter (cm) | 3.40 ± 0.60 | 3.47 ± 0.50 | 2.92 ± 0.40 | 6.02 ± 0.70 *** |
| Length of inflorescence (cm) | 12.06 ± 1.70 | 13.30 ± 1.20 | 24.30 ± 1.90 *** | 17.60 ± 1.90 *** |
| Trichome density | 42.20 ± 2.87 | 29.80 ± 1.18 *** | 62.30 ± 1.87 *** | 26.20 ± 0.54 *** |
| Trichome length (µm) | 69.85 ± 2.75 | 72.54 ± 1.87 * | 70.92 ± 1.32 | 88.30 ± 2.71 *** |
| Stomata length (µm) | 12.44 ± 0.65 | 13.00 ± 1.08 | 7.90 ± 0.61 *** | 21.20 ± 1.08 *** |
| Oil yield (mg/100 g leaves) | 434.67 ± 25.98 | 307.33 ± 31.22 ** | 411.33 ± 43.57 | 488.67 ± 22.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, S.; Rastogi, S.; Vashisth, D.; Rout, P.K.; Lal, R.K.; Lavania, U.C.; Shasany, A.K. Altered Developmental and Metabolic Gene Expression in Basil Interspecific Hybrids. Plants 2022, 11, 1873. https://doi.org/10.3390/plants11141873
Shah S, Rastogi S, Vashisth D, Rout PK, Lal RK, Lavania UC, Shasany AK. Altered Developmental and Metabolic Gene Expression in Basil Interspecific Hybrids. Plants. 2022; 11(14):1873. https://doi.org/10.3390/plants11141873
Chicago/Turabian StyleShah, Saumya, Shubhra Rastogi, Divya Vashisth, Prashant Kumar Rout, Raj Kishori Lal, Umesh Chandra Lavania, and Ajit Kumar Shasany. 2022. "Altered Developmental and Metabolic Gene Expression in Basil Interspecific Hybrids" Plants 11, no. 14: 1873. https://doi.org/10.3390/plants11141873