
Phenotypic Acclimation of Maize Plants Grown under S Deprivation and Implications to Sulfur and Iron Allocation Dynamics


Abstract
:1. Introduction
2. Results
2.1. Plasticity of Phenotypic Traits
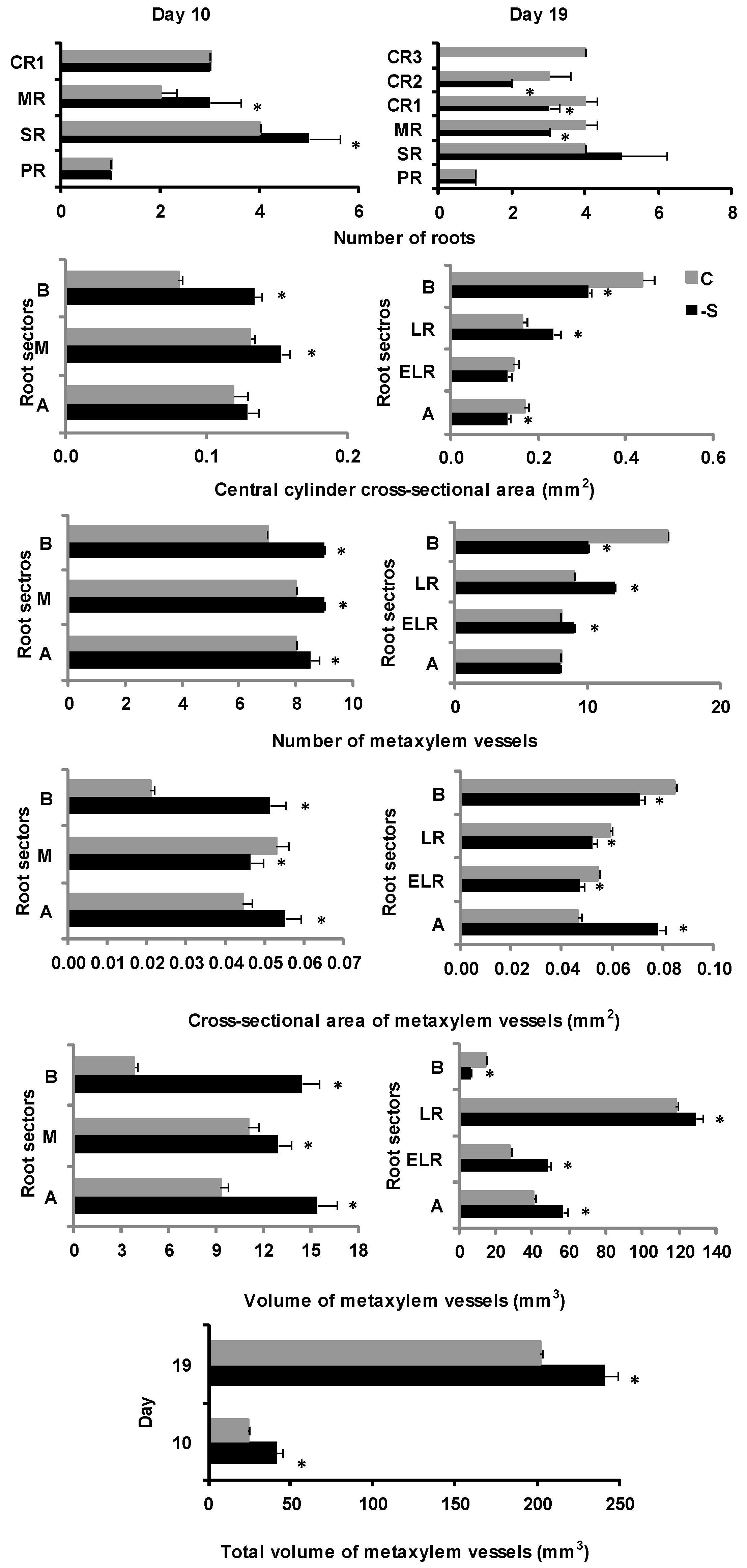
2.1.1. Root Morphological Features Related to Nutrient Exploration and Uptake
2.1.2. Root Anatomical Features Related to the Transport of Nutrients to the Aerial Plant Parts
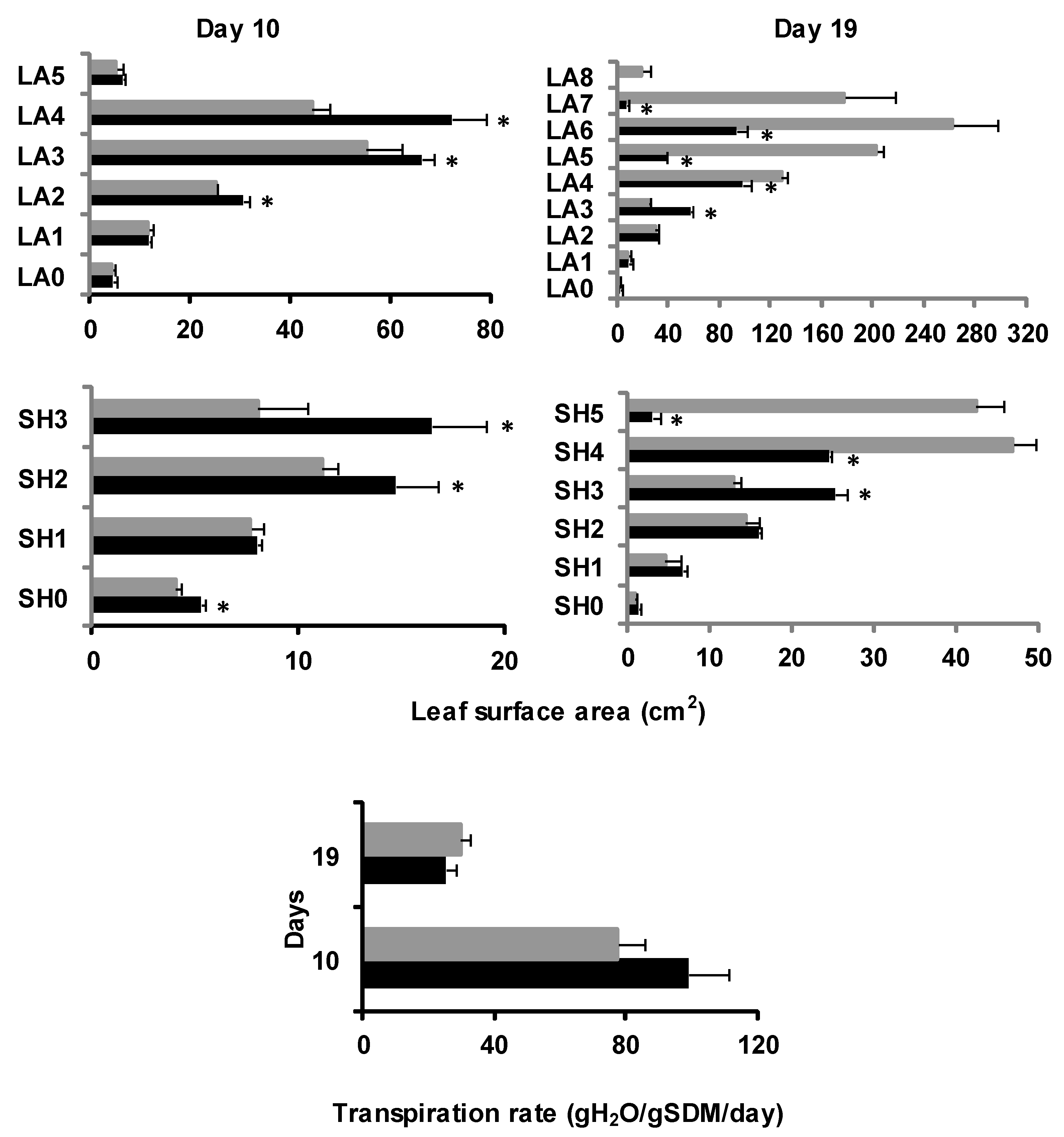
2.1.3. Plant Foliage Development
2.2. Dry Mass and Water Allocation Dynamics
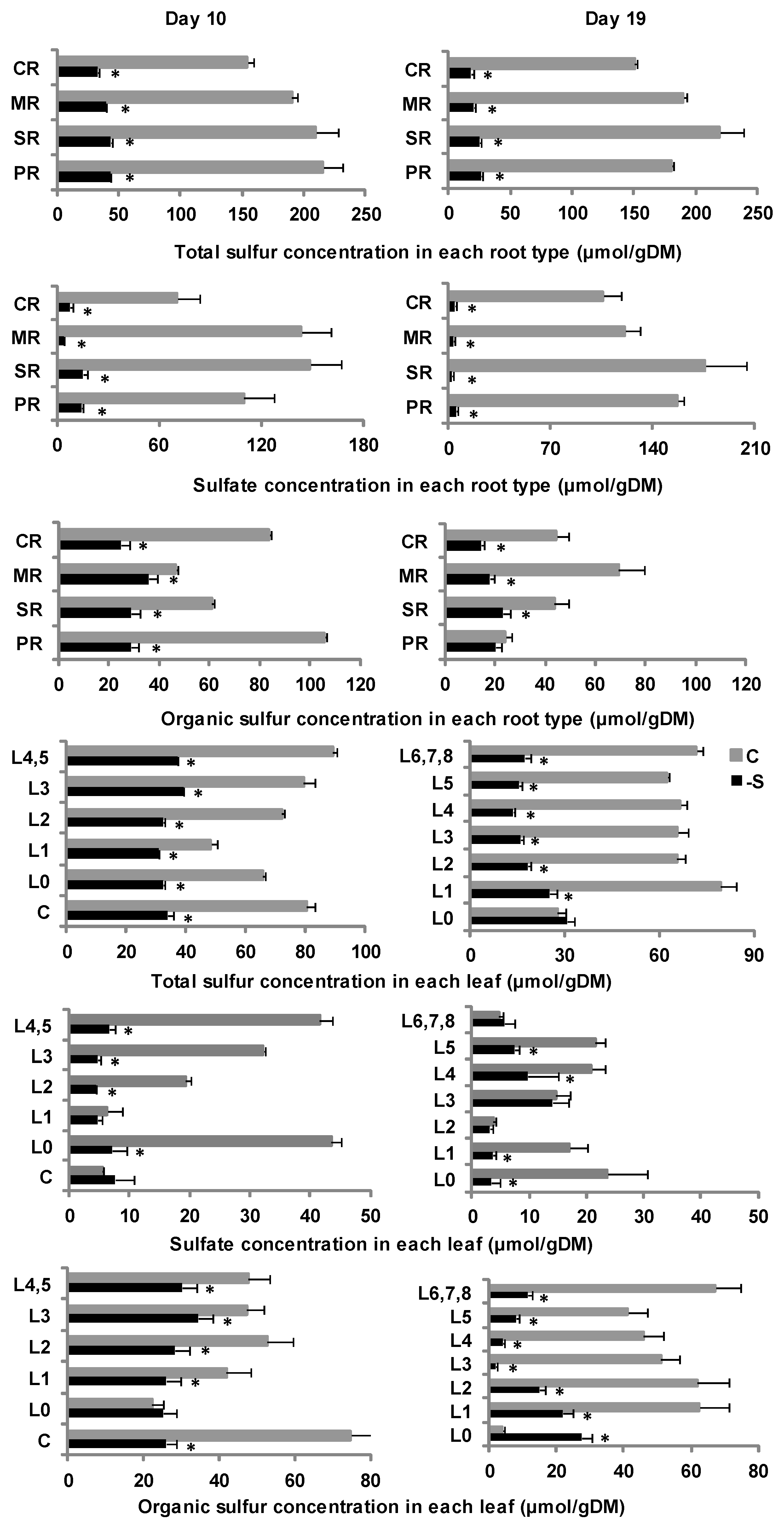
2.3. Sulfur Allocation Dynamics
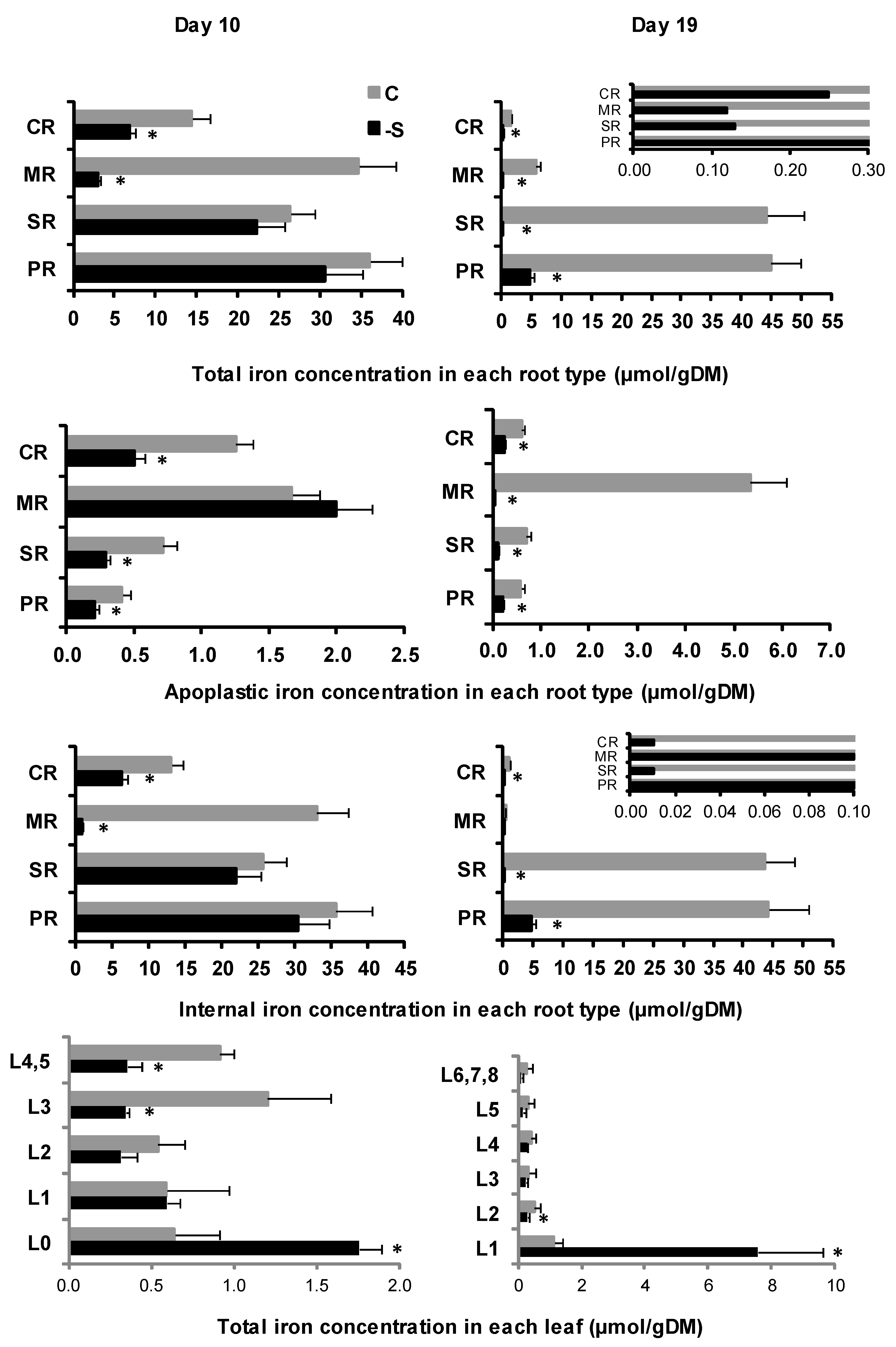
2.4. Iron Allocation Dynamics
3. Discussion
3.1. Root Morphology—Traits Involved in Exploration and Nutrient Uptake
3.2. Root Anatomy—Traits Involved in Transport/Translocation of Nutrients
3.3. Root Nutrients
3.4. Aerial Plant Parts and Nutrients’ Translocation
4. Materials and Methods
4.1. Plant Material and Hydroponics Set Up
4.2. Treatments and Samplings
4.3. Morphometric Analysis
4.4. Histological Study
4.5. Dry Mass Determination
4.6. Transpiration Rate Determination
4.7. Sulfate Determination
4.8. Total Sulfur Determination
4.9. Organic Sulfur Calculation
4.10. Total Iron Determination
4.11. Apoplastic Iron Determination
4.12. Internal Iron Calculation
4.13. Statistical Analysis
5. Conclusions
- They did not invest in new root axes.
- They developed more and larger xylem vessels in roots.
- Organic sulfur predominated over sulfates into their roots.
- Sulfates predominated over organic sulfur into their leaves.
- Τhere were differences in the subcellular localization of iron in their roots.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bouranis, D.L.; Chorianopoulou, S.N. Advances in understanding plant root anatomy and nutrient acquisition. In Understanding and Improving Crop Root Function; Gregory, P., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; ISBN 978178676 3600. Available online: www.bdspublishing.com (accessed on 23 January 2022). [CrossRef]
- Zhao, F.J.; Hawkesford, M.J.; McGrath, S.P. Sulphur assimilation and effects on yield and quality of wheat. J. Cereal Sci. 1999, 30, 1–17. [Google Scholar] [CrossRef]
- Hawkesford, M. Uptake, distribution and subcellular transport of sulfate. In Sulfur Metabolism in Phototrophic Organisms, 1st ed.; Hell, R., Dahl, C., Knaff, D., Leustek, T., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 27, pp. 15–30. [Google Scholar]
- Bell, C.I.; Clarkson, D.T.; Cram, W.J. Partitioning and redistribution of sulfur during S-stress in Macroptilium atropurpureum cv. Siratro. J. Exp. Bot. 1995, 46, 73–81. [Google Scholar] [CrossRef]
- Rennenberg, H.; Schmitz, K.; Bergmann, L. Long distance transport of sulfur in Nicotiana tabacum. Planta 1979, 147, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Sunarpi; Anderson, J.W. Distribution and redistribution of sulfur supplied as [35S]sulfate to roots during vegetative growth of soybean. Plant Physiol. 1996, 110, 1151–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman-Salit, A.; Veith, N.; Wirtz, M.; Hell, R.; Kummer, U. Distribution of control in the sulfur assimilation in Arabidopsis thaliana depends on environmental conditions. New Phytol. 2019, 222, 1392–1404. [Google Scholar] [CrossRef] [PubMed]
- Cram, W.J. Uptake and transport of sulfate. In Sulfur Nutrition and Sulfur Assimilation in Higher Plants; Rennenberg, H., Brunold, C.H., De Kok, L.J., Stulen, I., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1990; pp. 3–11. [Google Scholar]
- Bouma, D. The uptake and transport of sulphur in plants. In Sulphur in Australasian Agriculture; McLachlan, K.D., Ed.; Sydney University Press: Sydney, Australia, 1975; pp. 79–86. [Google Scholar]
- Clarkson, D.T.; Saker, L.R.; Purves, J.V.; Lee, R.B. Depression of nitrate and ammonium transport in barley plants with diminished sulphate status. Evidence of Co-regulation of nitrogen and sulphate intake. J. Exp. Bot. 1989, 40, 953–963. [Google Scholar] [CrossRef]
- Clarkson, D.T.; Smith, F.W.; Van den Berg, P.J. Regulation of sulphate transport in a tropical legume, Macroptillum atropurpureum, cv Sitatro. J. Exp. Bot. 1983, 34, 1463–1483. [Google Scholar] [CrossRef]
- Droux, M.; Ruffet, M.L.; Douce, R.; Job, D. Interactions between serine acetyltransferase and O-acetylserine (thiol) lyase in higher plants—Structural and kinetic properties of the free and bound enzymes. Eur. J. Biochem. 1998, 255, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, M.; Berkowitz, O.; Droux, M.; Hell, R. The cysteine synthase complex from plants. Mitochondrial serine acetyltransferase from Arabidopsis thaliana carries a bifunctional domain for catalysis and protein-protein interaction. Eur. J. Biochem. 2001, 268, 686–693. [Google Scholar] [CrossRef]
- De Kok, L.J.; Kuiper, P.J.C. Effect of short-term dark incubation with sulfate, cbloride and selenate on the glutathione content of spinach leaf discs. Physiol. Plant. 1986, 68, 477–482. [Google Scholar] [CrossRef]
- De Kok, L.J. Sulfur metabolism in plants exposed to atmospheric sulfur. In Sulfur Nutrition and Sulfur Assimilation in Higher Plants; Rennenberg, H., Brunold, C., De Kok, L.J., Stulen, I., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1990; pp. 111–130. [Google Scholar]
- Brunold, C. Reduction of sulfate to sulfide. In Sulfur Nutrition and Sulfur Assimilation in Higher Plants; Rennenberg, H., Brunold, C., De Kok, L.J., Stuten, I., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1990; pp. 13–33. [Google Scholar]
- Farago, S.; Brunold, B. Regulation of assimilatory sulfate reduction by herbicide safeners in Zea mays L. Plant Physiol. 1990, 94, 1808–1812. [Google Scholar] [CrossRef] [Green Version]
- Romheld, V.; Marschner, H. Evidence for a specific uptake system for iron phytosiderophores in roots of grasses. Plant Physiol. 1986, 80, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, K.; Suzuki, K.; Nakanishi, H.; Yamaguchi, H.; Nishizawa, N.K.; Mori, S. Cloning of nicotianamine synthase genes, novel genes involved in the biosynthesis of phytosiderophores. Plant Physiol. 1999, 119, 471–480. [Google Scholar] [CrossRef] [Green Version]
- Curie, C.; Panaviene, Z.; Loulergue, C.; Dellaporta, S.L.; Briat, J.F.; Walker, E.L. Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III) uptake. Nature 2001, 409, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Guerinot, M.L. Mining iron: Iron uptake and transport in plants. FEBS Lett. 2007, 581, 2273–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curie, C.; Cassin, G.; Couch, D.; Divol, F.; Higuchi, K.; Le Jean, M.; Misson, J.; Schikora, A.; Czernic, P.; Mari, S. Metal movement within the plant: Contribution of nicotianamine and yellow stripe 1-like transporters. Ann. Bot. 2009, 103, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydon, M.J.; Cobbett, C.S. Transporters of ligands for essential metal ions in plants. New Phytol. 2007, 174, 499–506. [Google Scholar] [CrossRef]
- Grotz, N.; Guerinot, M.L. Molecular aspects of Cu, Fe and Zn homeostasis in plants. Biochim. Biophys. Acta 2006, 1763, 595–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briat, J.F.; Curie, C.; Gaymard, F. Iron utilization and metabolism in plants. Curr. Opin. Plant Biol. 2007, 10, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Hintze, K.J.; Theil, E.C. Cellular regulation and molecular interactions of the ferritins. Cell. Mol. Life Sci. 2006, 63, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Forieri, I.; Wirtz, M.; Hell, R. Toward new perspectives on the interaction of iron and sulfur metabolism in plants. Front. Plant Sci. 2013, 4, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astolfi, S.; Zuchi, S.; Hubberten, H.M.; Pinton, R.; Hoefgen, R. Supply of sulphur to S-deficient young barley seedlings restores their capability to cope with iron shortage. J. Exp. Bot. 2010, 61, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, S.; Zuchi, S.; Neumann, G.; Cesco, S.; Di Toppi, L.S.; Pinton, R. Response of barley plants to Fe deficiency and Cd contamination as affected by S starvation. J. Exp. Bot. 2011, 63, 1241–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciaffi, M.; Paolacci, A.R.; Celletti, S.; Catarcione, G.; Kopriva, S.; Astolfi, S. Transcriptional and physiological changes in the S assimilation pathway due to single or combined S and Fe deprivation in durum wheat (Triticum durum L.) seedlings. J. Exp. Bot. 2013, 64, 1663–1675. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Zhang, C.; Dai, C.; Ge, Y. Sufficient sulfur supply promotes seedling growth, alleviates oxidation stress, and regulates iron uptake and translocation in rice. Biol. Plant 2015, 59, 788–792. [Google Scholar] [CrossRef]
- Wawrzyñska, A.; Sirko, A. The role of selective protein degradation in the regulation of iron and sulfur homeostasis in plants. Int. J. Mol. Sci. 2020, 21, 2771. [Google Scholar] [CrossRef] [Green Version]
- Gojon, A.; Nacry, P.; Davidian, J.C. Root uptake regulation: A central process for NPS homeostasis in plants. Curr. Opin. Plant Biol. 2009, 12, 328–338. [Google Scholar] [CrossRef]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Gruber, B.D.; Giehl, R.F.H.; Friedel, S.; von Wirén, N. Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Giehl, R.F.H.; von Wirén, N. Root Nutrient Foraging. Plant Physiol. 2014, 166, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Wan, F.; Chang, X.; Zhang, J.; Sun, M.; Liu, Y. Effects of nutrient deficiency on root morphology and nutrient allocation in Pistacia chinensis bunge seedlings. Forests 2019, 10, 1035. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Chen, F.; Yuan, L.; Zhang, F.; Mi, G. A comprehensive analysis of root morphological changes and nitrogen allocation in maize in response to low nitrogen stress. Plant Cell Environ. 2015, 38, 740–750. [Google Scholar] [CrossRef]
- Wang, Y.; Mi, G.H.; Chen, F.J.; Zhang, J.H.; Zhang, F.S. Response of root morphology to nitrate supply and its contribution to nitrogen accumulation in maize. J. Plant Nutr. 2004, 27, 2189–2202. [Google Scholar] [CrossRef]
- Gaudin, A.C.M.; Mc Clymont, S.A.; Holmes, B.M.; Lyons, E.; Raizada, M.N. Novel temporal, fine-scale and growth variation phenotypes in roots of adult-stage maize (Zea mays L.) in response to low nitrogen stress. Plant Cell Environ. 2011, 34, 2122–2137. [Google Scholar] [CrossRef] [PubMed]
- Dan, H.; Yang, G.; Zheng, Z.L. A negative regulatory role for auxin in sulphate deficiency response in Arabidopsis thaliana. Plant Mol. Biol. 2007, 63, 221–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Q.Y.; Chen, F.J.; Liu, J.X.; Zhang, F.S.; Mi, G.H. Inhibition of maize root growth by high nitrate supply is correlated with reduced IAA levels in roots. Plant Physiol. 2008, 165, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Kutz, A.; Muller, A.; Hennig, P.; Kaiser, W.M.; Piotrowski, M.; Weiler, E.W. A role for nitrilase 3 in the regulation of root morphology in sulphur-starving Arabidopsis thaliana. Plant J. 2002, 30, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Torres, C.A.; López-Bucio, J.; Cruz-Ramírez, A.; Ibarra-Laclette, E.; Dharmasiri, S.; Estelle, M.; Herrera-Estrella, L. Phosphate availability alters lateral root development in Arabidopsis by modulating auxin sensitivity via a mechanism involving the TIR1 auxin receptor. Plant Cell 2008, 20, 3258–3272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Donner, E.; Lombi, E.; Li, R.Y.; Wu, Z.C.; Zhao, F.J.; Wu, P. Assessing the contributions of lateral roots to element uptake in rice using an auxin-related lateral root mutant. Plant Soil 2013, 372, 125–136. [Google Scholar] [CrossRef]
- Postma, J.A.; Dathe, A.; Lynch, J.P. The optimal lateral root branching density for maize depends on nitrogen and phosphorus availability. Plant Physiol. 2014, 166, 590–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouranis, D.L.; Chorianopoulou, S.N.; Kollias, C.; Maniou, P.; Protonotarios, V.E.; Siyiannis, V.F.; Hawkesford, M.J. Dynamics of aerenchyma distribution in the cortex of sulfate-deprived adventitious roots of maize. Ann. Bot. 2006, 97, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Konings, H.; Verschuren, G. Formation of aerenchyma in roots of Zea mays in aerated solutions, and its relation to nutrient supply. Physiol. Plant. 1980, 49, 265–270. [Google Scholar] [CrossRef]
- Drew, M.C.; He, C.J.; Morgan, P.W. Decreased ethylene biosynthesis, and induction of aerenchyma, by nitrogen-or phosphate starvation in adventitious roots of Zea mays L. Plant Physiol. 1989, 91, 266–271. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Zhu, J.; Richards, C.; Brown, K.M.; Lynch, J.P. Physiological roles for aerenchyma in phosphorus-stressed roots. Funct. Plant Biol. 2003, 30, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Bouranis, D.L.; Chorianopoulou, S.N.; Siyiannis, V.F.; Protonotarios, V.E.; Hawkesford, M.J. Aerenchyma formation in roots of maize during sulphate starvation. Planta 2003, 217, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Root phenotypes for improved nutrient capture: An underexploited opportunity for global agriculture. New Phytol. 2019, 223, 548–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saengwilai, P.; Nord, E.A.; Chimungu, J.; Brown, K.M.; Lynch, J.P. Root cortical aerenchyma enhances nitrogen acquisition from low nitrogen soils in maize. Plant Physiol. 2014, 166, 726–735. [Google Scholar] [CrossRef] [Green Version]
- Schneider, H.M.; Postma, J.A.; Wojciechowski, T.; Kuppe, C.; Lynch, J.P. Root cortical senescence improves growth under suboptimal availability of N, P, and K. Plant Physiol. 2017, 174, 2333–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X. Enlarged Cortical Cells and Reduced Cortical Cell File Number Improve Maize Growth under Suboptimal Nitrogen, Phosphorus, and Potassium Availability. Master’s Thesis, The Pennsylvania State University, University Park, State College, PA, USA, 2017. [Google Scholar] [CrossRef]
- Chimungu, J.G.; Loades, K.W.; Lynch, J.P. Root anatomical phenes predict root penetration ability and biomechanical properties in maize (Zea mays). J. Exp. Bot. 2015, 66, 3151–3162. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P.; Wojciechowski, T. Opportunities and challenges in the subsoil: Pathways to deeper rooted crops. J. Exp. Bot. 2015, 66, 2199–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postma, J.A.; Lynch, J.P. Theoretical evidence for the functional benefit of root cortical aerenchyma in soils with low phosphorus availability. Ann. Bot. 2010, 107, 829–841. [Google Scholar] [CrossRef]
- Postma, J.A.; Lynch, J.P. Root cortical aerenchyma enhances the growth of maize on soils with suboptimal availability of nitrogen, phosphorus, and potassium. Plant Physiol. 2011, 156, 1190–1201. [Google Scholar] [CrossRef] [Green Version]
- Maniou, F.; Chorianopoulou, S.N.; Bouranis, D.L. New insights into trophic aerenchyma formation strategy in maize (Zea mays L.) organs during sulphate deprivation. Front. Plant Sci. 2014, 5, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astolfi, S.; Zuchi, S.; Cesco, S.; Varanini, Z.; Pinton, R. Influence of iron nutrition on sulphur uptake and metabolism in maize (Zea mays L.) roots. Soil Sci. Plant Nutr. 2004, 50, 1079–1083. [Google Scholar] [CrossRef]
- Pate, J.S. Roots as organs of assimilation of sulfate. Science 1965, 149, 547–548. [Google Scholar] [CrossRef]
- Bouranis, D.L.; Chorianopoulou, S.N.; Protonotarios, V.E.; Siyiannis, V.F.; Hopkins, L.; Hawkesford, M.J. Leaf responses of young iron−inefficient maize plants to sulfur deprivation. J. Plant Nutr. 2003, 26, 1189–1202. [Google Scholar] [CrossRef]
- Astolfi, S.; Cesco, S.; Zuchi, S.; Neumann, G.; Roemheld, V. Sulfur starvation reduces phytosiderophores release by iron-deficient barley plants. Soil Sci. Plant Nutr. 2006, 52, 43–48. [Google Scholar] [CrossRef]
- Zuchi, S.; Cesco, S.; Astolfi, S. High S supply improves Fe accumulation in durum wheat plants grown under Fe limitation. Environ. Exp. Bot. 2012, 77, 25–32. [Google Scholar] [CrossRef]
- Celletti, S.; Paolacci, A.R.; Mimmo, T.; Pii, Y.; Cesco, S.; Ciaffi, M. The effect of excess sulfate supply on iron accumulation in three graminaceous plants at the early vegetative phase. Environ. Exp. Bot. 2016, 128, 31–38. [Google Scholar] [CrossRef]
- Astolfi, S.; Zuchi, S.; Cesco, S.; Sanità di Toppi, L.; Pirazzi, D.; Badiani, M. Iron deficiency induces sulfate uptake and modulates redistribution of reduced sulfur pool in barley plants. Funct. Plant Biol. 2006, 33, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Bienfait, H.F.; van den Briel, W.; Mesland-Mul, N.T. Free space iron pools in roots: Generation and mobilization. Plant Physiol. 1985, 78, 596–600. [Google Scholar] [CrossRef] [Green Version]
- Longnecker, N.; Welch, R.M. Accumulation of apoplastic iron in plant roots. Plant Physiol. 1990, 92, 17–22. [Google Scholar] [CrossRef]
- Zhang, F.S.; Römheld, V.; Marschner, H. Role of the root apoplasm for iron acquisition by wheat plants. Plant Physiol. 1991, 97, 1302–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masalha, J.; Kosegarten, H.; Elmaci, O.; Mengel, K. The central role of microbial activity for iron acquisition in maize and sunflower. Biol. Fertil. Soils 2000, 30, 433–439. [Google Scholar] [CrossRef]
- Kosegarten, H.; Koyro, H.W. Apoplastic accumulation of iron in the epidermis of maize (Zea mays) roots grown in calcareous soil. Physiol. Plant. 2001, 113, 515–522. [Google Scholar] [CrossRef]
- Strasser, O.; Kohl, K.; Romheld, V. Overestimation of apoplastic Fe in roots of soil grown plants. Plant Soil 1999, 210, 179–187. [Google Scholar] [CrossRef]
- Mori, S.; Nishizawa, N. Methionine as a dominant precursor of phytosiderophores in Graminaea plants. Plant Cell Physiol. 1987, 28, 1081–1092. [Google Scholar] [CrossRef]
- Astolfi, S.; Zuchi, S.; Passera, C.; Cesco, S. Does the sulfur assimilation pathway play a role in the response to Fe deficiency in Maize (Zea mays L.) Plants? J. Plant Nutr. 2003, 26, 2111–2121. [Google Scholar] [CrossRef]
- Saito, K. Sulfur Assimilatory Metabolism. The Long and Smelling Road. Plant Physiol. 2004, 136, 2443–2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, F.W.; Hawkesford, M.J.; Ealing, P.M.; Clarkson, D.T.; Vanden Berg, P.J.; Belcher, A.R.; Warrilow, A.G. Regulation of expression of a cDNA from barley roots encoding a high affinity sulphate transporter. Plant J. 1997, 12, 875–884. [Google Scholar] [CrossRef]
- Koprivova, A.; Suter, M.; Op den Camp, R.; Brunold, C.; Kopriva, S. Regulation of sulfate assimilation by nitrogen in Arabidopsis. Plant Physiol. 2000, 122, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Bouranis, D.L.; Chorianopoulou, S.N.; Siyiannis, V.F.; Protonotarios, V.E.; Koufos, C.; Maniou, P. Changes in nutrient allocation between roots and shoots of young maize plants during sulfate deprivation. J. Plant Nutr. 2012, 175, 499–510. [Google Scholar] [CrossRef]
- Wu, Z.; Naveed, S.; Zhang, C.; Ge, Y. Adequate supply of sulfur simultaneously enhances iron uptake and reduces cadmium accumulation in rice grown in hydroponic culture. Environ. Pollut. 2020, 262, 114327. [Google Scholar] [CrossRef] [PubMed]
- Von Wirén, N.; Klair, S.; Bansal, S.; Briat, J.F.; Khodr, H.; Shioiri, T. Nicotianamine chelates both Fe(III) and Fe(II) implications for metal transport in plants. Plant Physiol. 1999, 119, 1107–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochholdinger, F. The Maize Root System: Morphology, Anatomy, and Genetics. In Handbook of Maize: Its Biology; Bennetzen, J.L., Hake, S.C., Eds.; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Mills, H.A.; Jones, J.B., Jr. Plant Analysis Handbook II. A Practical Sampling, Preparation, Analysis, and Interpretation Guide; Micro Macro Publishing Inc.: Athens, GA, USA, 1996. [Google Scholar]
- Sörbo, B. Sulfate: Turbidimetric and nephelometric methods. In Methods in Enzymology: Sulfur and Sulfur Amino Acids; Jakoby, W.B., Griffith, O.W., Eds.; Academic Press, Inc.: New York, NY, USA, 1987; Volume 143, pp. 3–6. [Google Scholar]
- Miller, R.O. Extractable chloride, nitrate, orthophosphate, potassium, and sulfate-sulfur in plant tissue: 2% acetic acid extraction. In Handbook of Reference Methods for Plant Analysis; Kalra, Y.P., Ed.; CRC Press LLC.: Boca Raton, FL, USA, 1998; pp. 115–118. [Google Scholar]
- Taylor, G.J.; Crowder, A.A. Use of DCB technique for extraction of hydrous iron oxides from roots of wetland plants. Am. J. Bot. 1983, 70, 1254–1257. [Google Scholar] [CrossRef]
- Otte, M.L.; Dekkers, M.J.; Rozema, J.; Broekman, R.A. Uptake of arsenic by Aster tripolium in relation to rhizosphere oxidation. Can. J. Bot. 1991, 69, 2670–2677. [Google Scholar] [CrossRef]
- Liu, W.J.; Zhu, Y.G.; Smith, F.A.; Smith, S.E. Do phosphorus nutrition and iron plaque alter arsenate (As) uptake by rice seedlings in hydroponic culture? New Phytol. 2004, 162, 481–488. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf 5 * | Day 10 | Day 19 | ||||||||
| Leaf length LA5 | = | C | ||||||||
| Leaf surface area LA5 | = | C | ||||||||
| Dry mass LA5 | C | C | ||||||||
| Water content LA5 | -S | -S | ||||||||
| Total sulfur concentration | C | C | ||||||||
| Sulfate concentration | C | C | ||||||||
| Organic sulfur concentration | C | C | ||||||||
| Total iron concentration | C | C | ||||||||
| * the youngest lamina at day 10 | ||||||||||
| Leaf 3 ** | Day 10 | Day 19 | ||||||||
| Leaf length LA3 | = | = | ||||||||
| Leaf surface area LA3 | = | -S | ||||||||
| Dry mass LA3 | = | = | ||||||||
| Water content LA3 | = | -S | ||||||||
| Total sulfur concentration | C | C | ||||||||
| Sulfate concentration | C | = | ||||||||
| Organic sulfur concentration | C | C | ||||||||
| Total iron concentration | C | C | ||||||||
| ** constant lamina length between days 10 and 19 | ||||||||||
| (its development has been completed) | ||||||||||
| Shoot | Day 10 | Day 19 | ||||||||
| Transpiration rate | -S | = | ||||||||
| Total sulfur concentration | C | C | ||||||||
| Sulfate concentration | C | = | ||||||||
| Organic sulfur concentration | C | C | ||||||||
| Total iron concentration | C | (-S) | ||||||||
| C | C | C | CR1 | Day 10 | Day 19 | C | C | C | ||
| C | C | C | Length of sector | B | x | = | C | C | C | |
| Total sulfur concentration | Sulfate concentration | Organic sulfur concentration | LR | x | (C) | Total iron concentration | Apoplastic iron concentration | Internal iron concentration | ||
| ELR | x | -S | ||||||||
| A | = | (C) | ||||||||
| Root cross sectional area | B | -S | C | |||||||
| LR | -S | -S | ||||||||
| ELR | -S | = | ||||||||
| A | -S | C | ||||||||
| Epidermis and hypodermis cross-sectional area | B | -S | C | |||||||
| LR | -S | = | ||||||||
| ELR | -S | -S | ||||||||
| A | -S | C | ||||||||
| Cortical parenchyma cross-sectional area | B | -S | C | |||||||
| LR | -S | -S | ||||||||
| ELR | -S | C | ||||||||
| A | -S | C | ||||||||
| Aerenchymatous area | B | -S | -S | |||||||
| LR | -S | -S | ||||||||
| ELR | -S | C | ||||||||
| A | -S | -S | ||||||||
| = | C | (C) | = | Central cylinder cross-sectional area | B | -S | C | |||
| = | = | x | x | LR | -S | -S | ||||
| ELR | -S | = | ||||||||
| Root length | Number of roots | Number of lateral roots | Length of lateral roots | A | = | C | ||||
| Number of metaxylem vessels | B | -S | C | = | (-S) | |||||
| LR | -S | -S | ||||||||
| ELR | -S | -S | -S | C | ||||||
| A | -S | = | ||||||||
| Cross-sectional area of metaxylem vessels | B | -S | C | Dry mass | Water content | |||||
| LR | C | C | ||||||||
| ELR | C | C | ||||||||
| A | -S | -S | ||||||||
| Total volume of metaxylem vessels | -S | -S | ||||||||
| Day | PR | SR | MR | CR | |
|---|---|---|---|---|---|
| % FeAPO | |||||
| C | 0 | 43 | 78 | ||
| 10 | 1 | 3 | 5 | 9 | |
| 19 | 1 | 2 | 94 | 38 | |
| -S | 0 | 43 | 78 | ||
| 10 | 1 | 1 | 67 | 7 | |
| 19 | 4 | 95 | 14 | 97 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maniou, F.S.; Bouranis, D.L.; Ventouris, Y.E.; Chorianopoulou, S.N. Phenotypic Acclimation of Maize Plants Grown under S Deprivation and Implications to Sulfur and Iron Allocation Dynamics. Plants 2022, 11, 703. https://doi.org/10.3390/plants11050703
Maniou FS, Bouranis DL, Ventouris YE, Chorianopoulou SN. Phenotypic Acclimation of Maize Plants Grown under S Deprivation and Implications to Sulfur and Iron Allocation Dynamics. Plants. 2022; 11(5):703. https://doi.org/10.3390/plants11050703
Chicago/Turabian StyleManiou, Filippa S., Dimitris L. Bouranis, Yannis E. Ventouris, and Styliani N. Chorianopoulou. 2022. "Phenotypic Acclimation of Maize Plants Grown under S Deprivation and Implications to Sulfur and Iron Allocation Dynamics" Plants 11, no. 5: 703. https://doi.org/10.3390/plants11050703