
Different Infectivity of Mediterranean and Southern Asian Tomato Leaf Curl New Delhi Virus Isolates in Cucurbit Crops

 , , and
, , and
Abstract
:
1. Introduction
2. Results
2.1. Viral Isolation and Phylogenetic Analysis of Two ToLCNDV Isolates
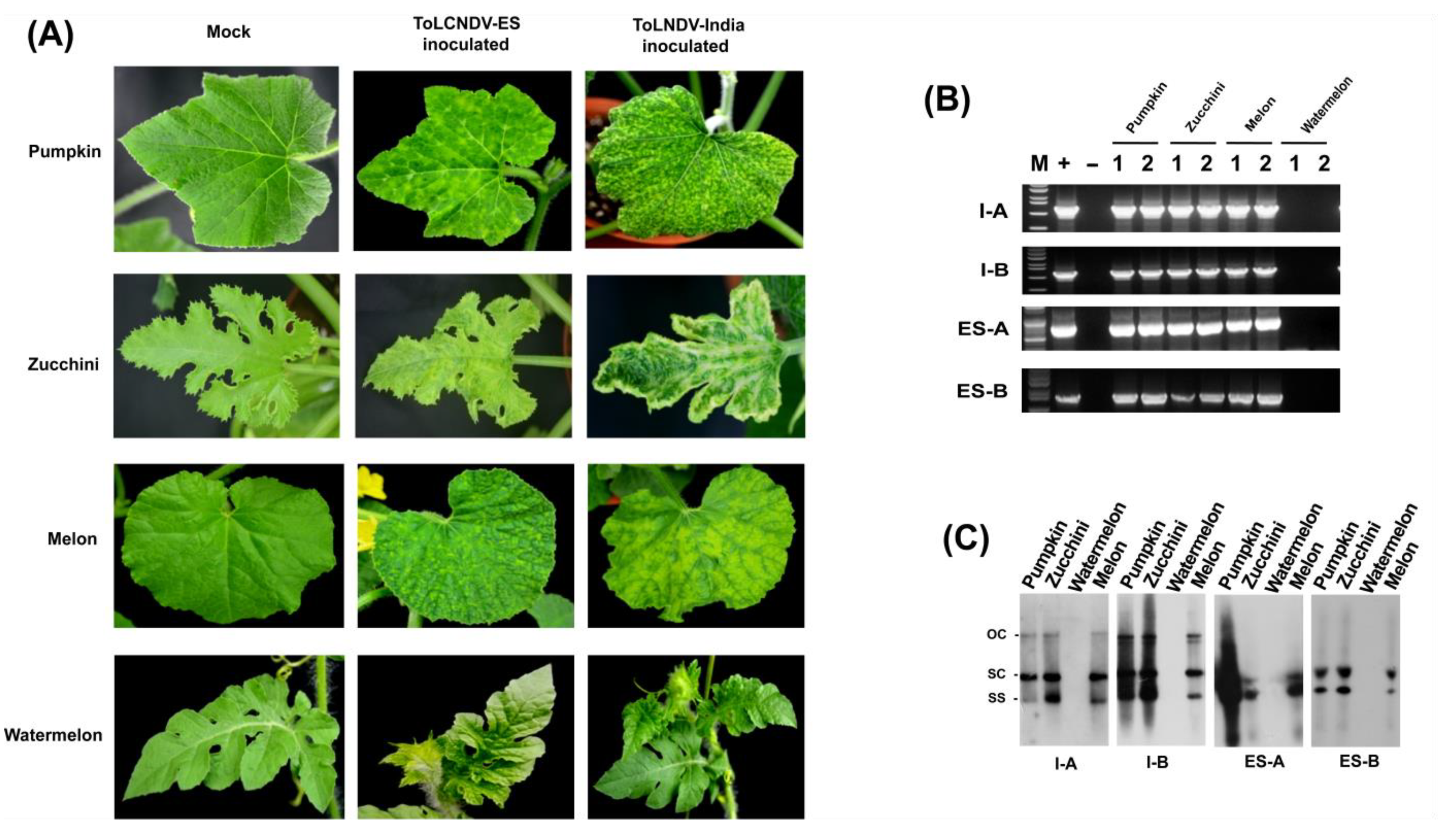
2.2. Infectivity of Two ToLCNDV Isolates’ Infectious Clone
2.3. Inoculation of Two ToLCNDV Isolates on Different Watermelon Cultivars
3. Discussion
4. Materials and Methods
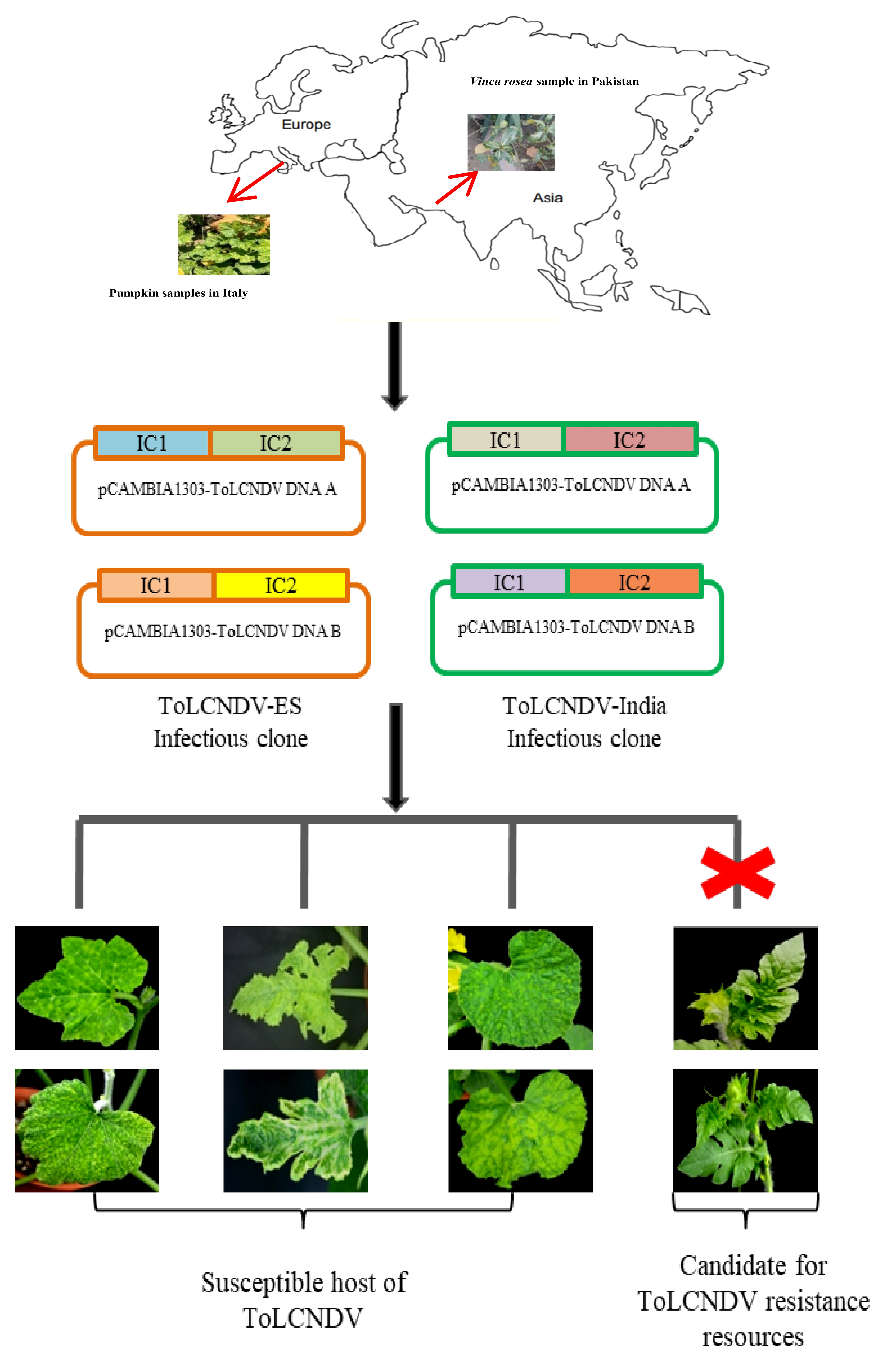
4.1. Virus Sources
4.2. Phylogenetic Analysis
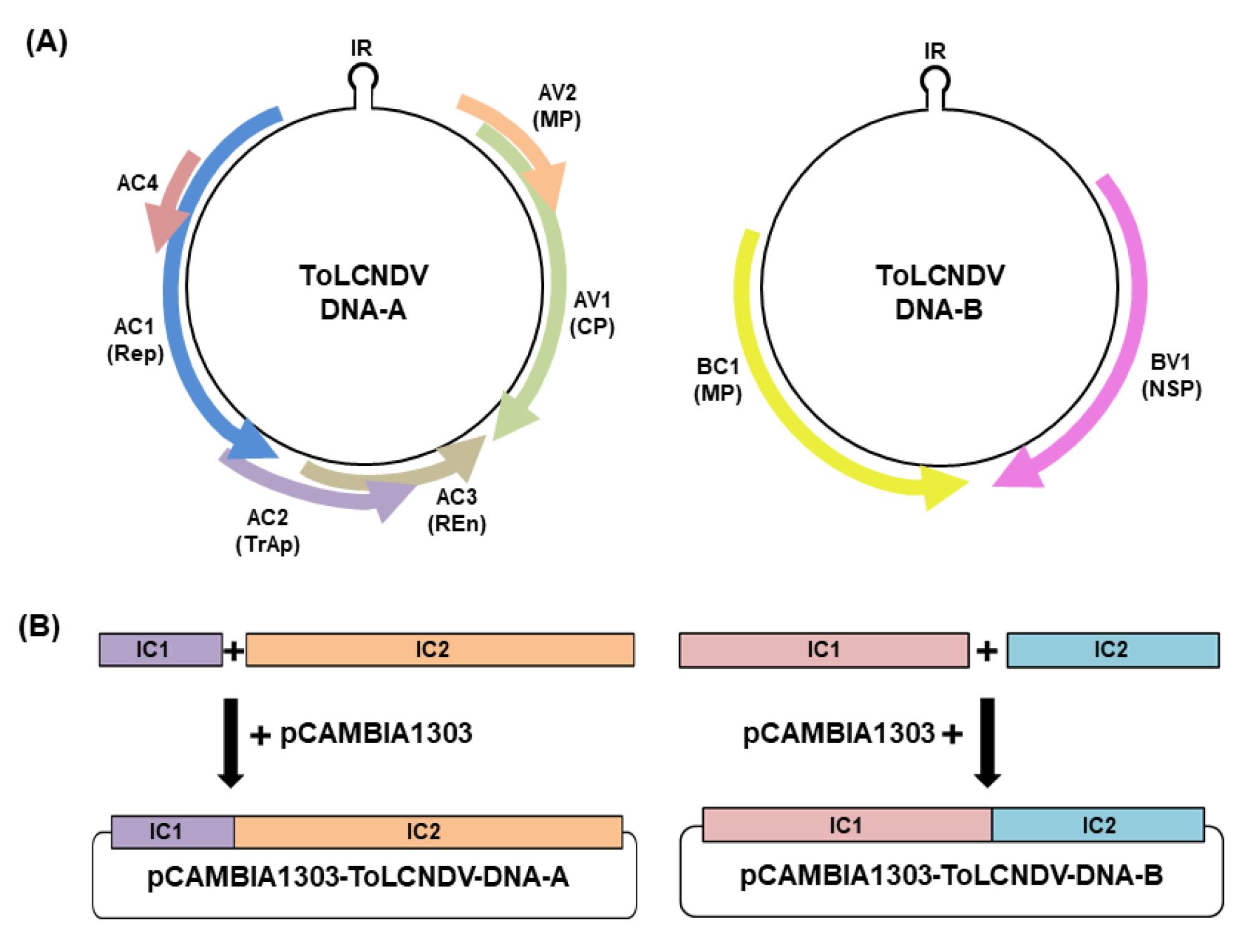
4.3. ToLCNDV Infectious Clone Construction
4.4. Inoculation Tests
4.5. Viral DNA Detection
4.6. Southern Hybridization Blot
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Srivastava, K.M.; Hallan, V.; Raizada, R.K.; Chandra, G.; Singh, B.P.; Sane, P.V. Molecular cloning of Indian tomato leaf curl virus genome following a simple method of concentrating the supercoiled replicative form of viral DNA. J. Virol. Methods 1995, 51, 297–304. [Google Scholar] [CrossRef]
- Hussain, M.; Mansoor, S.; Iram, S.; Zafar, Y.; Briddon, R. First report of tomato leaf curl New Delhi virus affecting chilli pepper in Pakistan. Plant Pathol. 2004, 53, 794. [Google Scholar] [CrossRef]
- Chang, H.-H.; Ku, H.-M.; Tsai, W.-S.; Chien, R.-C.; Jan, F.-J. Identification and characterization of a mechanical transmissible begomovirus causing leaf curl on oriental melon. Eur. J. Plant Pathol. 2010, 127, 219–228. [Google Scholar] [CrossRef]
- Maruthi, M.N.; Rekha, A.R.; Cork, A.; Colvin, J.; Alam, S.N.; Kader, K.A. First Report of Tomato leaf curl New Delhi virus Infecting Tomato in Bangladesh. Plant Dis. 2005, 89, 1011. [Google Scholar] [CrossRef]
- Juárez, M.; Tovar, R.; Fiallo-Olivé, E.; Aranda, M.A.; Gosálvez, B.; Castillo, P.; Moriones, E.; Navas-Castillo, J. First Detection of Tomato leaf curl New Delhi virus Infecting Zucchini in Spain. Plant Dis. 2014, 98, 857. [Google Scholar] [CrossRef]
- Radouane, N.; Tahiri, A.; Ghadraoui, L.E.; Figuigui, J.A.; Lahlali, R. First report of Tomato Leaf Curl New Delhi virus in Morocco. Plant Dis. 2018, 37, 2. [Google Scholar] [CrossRef] [Green Version]
- Orfanidou, C.G.; Malandraki, I.; Beris, D.; Kektsidou, O.; Vassilakos, N.; Varveri, C.; Katis, N.I.; Maliogka, V.I. First report of tomato leaf curl New Delhi virus in zucchini crops in Greece. J. Plant Pathol. 2019, 101, 799. [Google Scholar] [CrossRef] [Green Version]
- Panno, S.; Iacono, G.; Davino, M.; Marchione, S.; Zappardo, V.; Bella, P.; Tomassoli, L.; Accotto, G.P.; Davino, S. First report of Tomato leaf curl New Delhi virus affecting zucchini squash in an important horticultural area of southern Italy. New Dis. Rep. 2016, 33, 6. [Google Scholar] [CrossRef] [Green Version]
- Mnari-Hattab, M.; Zammouri, S.; Belkadhi, M.S.; Doña, D.B.; Nahia, E.b.; Hajlaoui, M.R. First report of Tomato leaf curl New Delhi virus infecting cucurbits in Tunisia. New Dis. Rep. 2015, 31, 21. [Google Scholar] [CrossRef] [Green Version]
- Parrella, G.; Troiano, E.; Formisano, G.; Accotto, G.P.; Giorgini, M. First Report of Tomato leaf curl New Delhi virus Associated with Severe Mosaic of Pumpkin in Italy. Plant Dis. 2018, 102, 459. [Google Scholar] [CrossRef]
- Ruiz, M.L.; Simón, A.; Velasco, L.; García, M.C.; Janssen, D. First Report of Tomato leaf curl New Delhi virus Infecting Tomato in Spain. Plant Dis. 2015, 99, 894. [Google Scholar] [CrossRef]
- Moriones, E.; Shelly, P.; Chakraborty, S. Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops. Viruses 2017, 9, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.S.-E.-A.; Martin, D.P.; Amin, I.; Farooq, M.; Mansoor, S. Tomato leaf curl New Delhi virus: A widespread bipartite begomovirus in the territory of monopartite begomoviruses. Mol. Plant Pathol. 2017, 18, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Kil, E.-J.; Vo, T.T.B.; Fadhila, C.; Ho, P.T.; Lal, A.; Troiano, E.; Parrella, G.; Lee, S. Seed Transmission of Tomato Leaf Curl New Delhi Virus from Zucchini Squash in Italy. Plants 2020, 9, 563. [Google Scholar] [CrossRef] [PubMed]
- López, C.; Ferriol, M.; Picó, M.B. Mechanical transmission of Tomato leaf curl New Delhi virus to cucurbit germplasm: Selection of tolerance sources in Cucumis melo. Euphytica 2015, 204, 679–691. [Google Scholar] [CrossRef]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Fiallo-Olivé, E.; Briddon, R.W.; Hernández-Zepeda, C.; Idris, A.; et al. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef]
- Fortes, I.M.; Sánchez-Campos, S.; Fiallo-Olivé, E.; Díaz-Pendón, J.A.; Navas-Castillo, J.; Moriones, E. A Novel Strain of Tomato Leaf Curl New Delhi Virus Has Spread to the Mediterranean Basin. Viruses 2016, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Snehi, S.K.; Raj, S.K.; Prasad, V.; Singh, V. Recent Research Findings Related to Management Strategies of Begomoviruses. J. Plant Pathol. Microbiol. 2015, 6, 1–12. [Google Scholar]
- Islam, S.; Munshi, A.D.; Mandal, B.; Kumar, R.; Behera, T.K. Genetics of resistance in Luffa cylindrica Roem. against Tomato leaf curl New Delhi virus. Euphytica 2010, 174, 83–89. [Google Scholar] [CrossRef]
- Sáez, C.; Martínez, C.; Ferriol, M.; Manzano, S.; Velasco, L.; Jamilena, M.; López, C.; Picó, B. Resistance to Tomato leaf curl New Delhi virus in Cucurbita spp. Ann. Appl. Biol. 2016, 169, 91–105. [Google Scholar] [CrossRef]
- Sáez, C.; Esteras, C.; Martínez, C.; Ferriol, M.; Dhillon, N.P.S.; López, C.; Picó, B. Resistance to tomato leaf curl New Delhi virus in melon is controlled by a major QTL located in chromosome 11. Plant Cell Rep. 2017, 36, 1571–1584. [Google Scholar] [CrossRef] [PubMed]
- Sáez, C.; Ambrosio, L.G.M.; Miguel, S.M.; Valcárcel, J.V.; Díez, M.J.; Picó, B.; López, C. Resistant Sources and Genetic Control of Resistance to ToLCNDV in Cucumber. Microorganisms 2021, 9, 913. [Google Scholar] [CrossRef]
- Seal, S.E.; Jeger, M.J.; Van den Bosch, F. Begomovirus Evolution and Disease Management. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2006; Volume 67, pp. 297–316. [Google Scholar]
- Crespo, O.; Robles, C.; Ruiz, L.; Janssen, D. Antagonism of Cucumber green mottle mosaic virus against Tomato leaf curl New Delhi virus in zucchini and cucumber. Ann. Appl. Biol. 2020, 176, 147–157. [Google Scholar] [CrossRef]
- Panno, S.; Caruso, A.G.; Troiano, E.; Luigi, M.; Manglli, A.; Vatrano, T.; Iacono, G.; Marchione, S.; Bertin, S.; Tomassoli, L.; et al. Emergence of tomato leaf curl New Delhi virus in Italy: Estimation of incidence and genetic diversity. Plant Pathol. 2019, 68, 601–608. [Google Scholar] [CrossRef]
- Janssen, D.; Simón, A.; Boulares, M.; Ruiz, L. Host Species-Dependent Transmission of Tomato Leaf Curl New Delhi Virus-ES by Bemisia tabaci. Plants 2022, 11, 390. [Google Scholar] [CrossRef]
- Juárez, M.; Rabadán, M.P.; Martínez, L.D.; Tayahi, M.; Grande-Pérez, A.; Gómez, P. Natural Hosts and Genetic Diversity of the Emerging Tomato Leaf Curl New Delhi Virus in Spain. Front. Microbiol. 2019, 10, 140. [Google Scholar] [CrossRef]
- Yamamoto, H.; Wakita, Y.; Kitaoka, T.; Fujishiro, T.; Kesumawati, E.; Koeda, S. Southeast Asian Isolate of the Tomato Leaf Curl New Delhi Virus Shows Higher Pathogenicity Against Tomato and Cucurbit Crops Compared to that of the Mediterranean Isolate. Hort. J. 2021, 90, 314–325. [Google Scholar] [CrossRef]
- Venkataravanappa, V.; Ashwathappa, K.V.; Reddy, C.N.L.; Shankarappa, K.S.; Reddy, M.K. Characterization of Tomato leaf curl New Delhi virus associated with leaf curl and yellowing disease of Watermelon and development of LAMP assay for its detection. 3 Biotech 2020, 10, 282. [Google Scholar] [CrossRef]
- Tian, G.; Yang, Y.; Zhang, S.; Miao, H.; Lu, H.; Wang, Y.; Xie, B.; Gu, X. Genetic analysis and gene mapping of papaya ring spot virus resistance in cucumber. Mol. Breed. 2015, 35, 110. [Google Scholar] [CrossRef]
- Tian, G.; Miao, H.; Yang, Y.; Zhou, J.; Lu, H.; Wang, Y.; Xie, B.; Zhang, S.; Gu, X. Genetic analysis and fine mapping of Watermelon mosaic virus resistance gene in cucumber. Mol. Breed. 2016, 36, 131. [Google Scholar] [CrossRef]
- Sáez, C.; Martínez, C.; Montero-Pau, J.; Esteras, C.; Sifres, A.; Blanca, J.; Ferriol, M.; López, C.; Picó, B. A Major QTL Located in Chromosome 8 of Cucurbita moschata Is Responsible for Resistance to Tomato Leaf Curl New Delhi Virus. Front. Plant Sci. 2020, 11, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, M.R. Use of degenerate primers in the polymerase chain reaction to detect whitefly-transmitted geminiviruses. Plant Dis. 1993, 77, 340–347. [Google Scholar] [CrossRef]
- Lal, A.; Kil, E.-J.; Vo, T.T.B.; Fadhila, C.; Ho, P.T.; Shuja, M.N.; Ali, M.; Lee, S. First Report of Duranta leaf curl virus Infecting Ficus virens Showing Leaf Curl Symptoms in Pakistan. Plant Dis. 2020, 104, 2034. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Urbino, C.; Thébaud, G.; Granier, M.; Blanc, S.; Peterschmitt, M. A novel cloning strategy for isolating, genotyping and phenotyping genetic variants of geminiviruses. Virol. J. 2008, 5, 135. [Google Scholar] [CrossRef] [Green Version]
- Hosseinpour azad, N.; Nematzadeh, G. Introducing a new method of genomic DNA extraction in dicotyledonous medical plants. Sch. J. Agric. Sci. 2012, 8, 111–120. [Google Scholar]
- Southern, E. Southern blotting. Nat. Protoc. 2006, 1, 518–525. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Plant | Infectivity * | PCR | Symptom |
|---|---|---|---|---|
| ToLCNDV-India | Pumpkin (cv. Shin Asia Matdol) | 7/7 | + | Curling, yellow mosaic |
| Pumpkin (cv. Napoletana) | 7/7 | + | Curling, yellow mosaic | |
| Zucchini (cv. Saeroun) | 6/7 | + | Severe curling, yellow mosaic | |
| Zucchini (cv. Romanesco) | 5/7 | + | Severe curling, yellow mosaic | |
| Melon (cv. Earl’s Mountain PMR) | 6/7 | + | Curling, yellow mosaic | |
| Melon (cv. RetatoDegliOrtolani) | 5/7 | + | Curling, yellow mosaic | |
| Watermelon (cv. Yeoreumen) | 0/9 | - | -- | |
| Watermelon (cv. Sugar Baby) | 0/9 | - | -- | |
| ToLCNDV-ES | Pumpkin (cv. Shin Asia Matdol) | 7/7 | + | Mild curling, yellow mosaic |
| Pumpkin (cv. Napoletana) | 6/7 | + | Mild curling, yellow mosaic | |
| Zucchini (cv. Saeroun) | 5/7 | + | Curling, yellow mosaic | |
| Zucchini (cv. Romanesco) | 5/7 | + | Curling, yellow mosaic | |
| Melon (cv. Earl’s Mountain PMR) | 5/7 | + | Mild curling, yellow mosaic | |
| Melon (cv Retato Degli Ortolani) | 7/7 | + | Mild curling, yellow mosaic | |
| Watermelon (cv. Yeoreumen) | 0/10 | - | -- | |
| Watermelon (cv. Sugar Baby) | 0/10 | - | -- |
| Cultivars | Infectivity * | |
|---|---|---|
| ToLCNDV-India | ToLCNDV-ES | |
| Black honey | 0/7 | 0/7 |
| Honey gold | 0/7 | 0/7 |
| Sinseolgang102 | 0/7 | 0/7 |
| Plus Honey | 0/7 | 0/7 |
| Nevada-F1 | 0/7 | 0/7 |
| Brera-F1 | 0/7 | 0/7 |
| Talete-F1 | 0/7 | 0/7 |
| Crimson Sweet | 0/7 | 0/7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vo, T.T.B.; Lal, A.; Ho, P.T.; Troiano, E.; Parrella, G.; Kil, E.-J.; Lee, S. Different Infectivity of Mediterranean and Southern Asian Tomato Leaf Curl New Delhi Virus Isolates in Cucurbit Crops. Plants 2022, 11, 704. https://doi.org/10.3390/plants11050704
Vo TTB, Lal A, Ho PT, Troiano E, Parrella G, Kil E-J, Lee S. Different Infectivity of Mediterranean and Southern Asian Tomato Leaf Curl New Delhi Virus Isolates in Cucurbit Crops. Plants. 2022; 11(5):704. https://doi.org/10.3390/plants11050704
Chicago/Turabian StyleVo, Thuy T. B., Aamir Lal, Phuong T. Ho, Elisa Troiano, Giuseppe Parrella, Eui-Joon Kil, and Sukchan Lee. 2022. "Different Infectivity of Mediterranean and Southern Asian Tomato Leaf Curl New Delhi Virus Isolates in Cucurbit Crops" Plants 11, no. 5: 704. https://doi.org/10.3390/plants11050704