
First Survey of the Vascular and Cryptogam Flora on Bulgaria’s Ancient Mounds

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
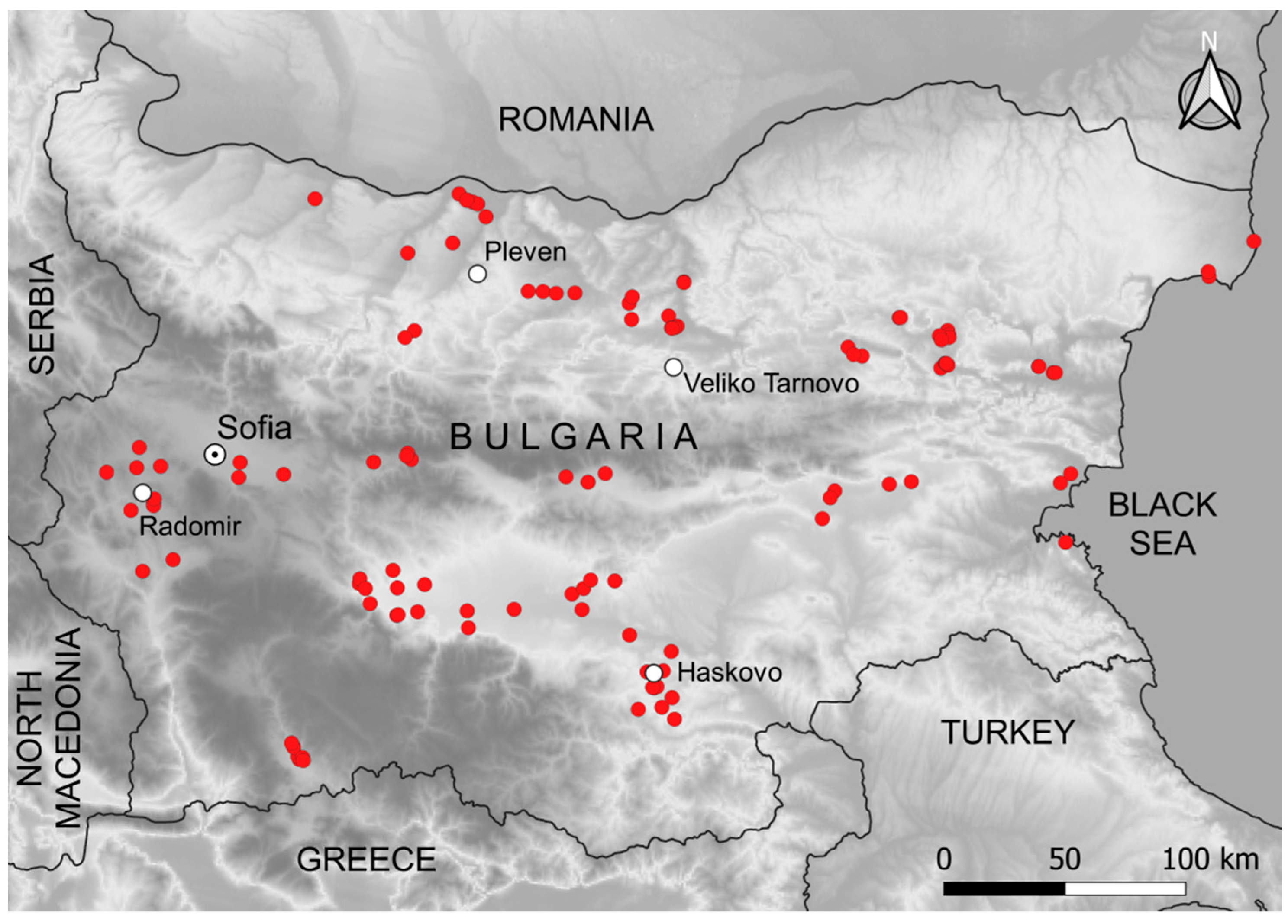
2.1. Study Objects and Study Area
2.2. Sampling Design, Data Collection and Data Analysis
3. Results
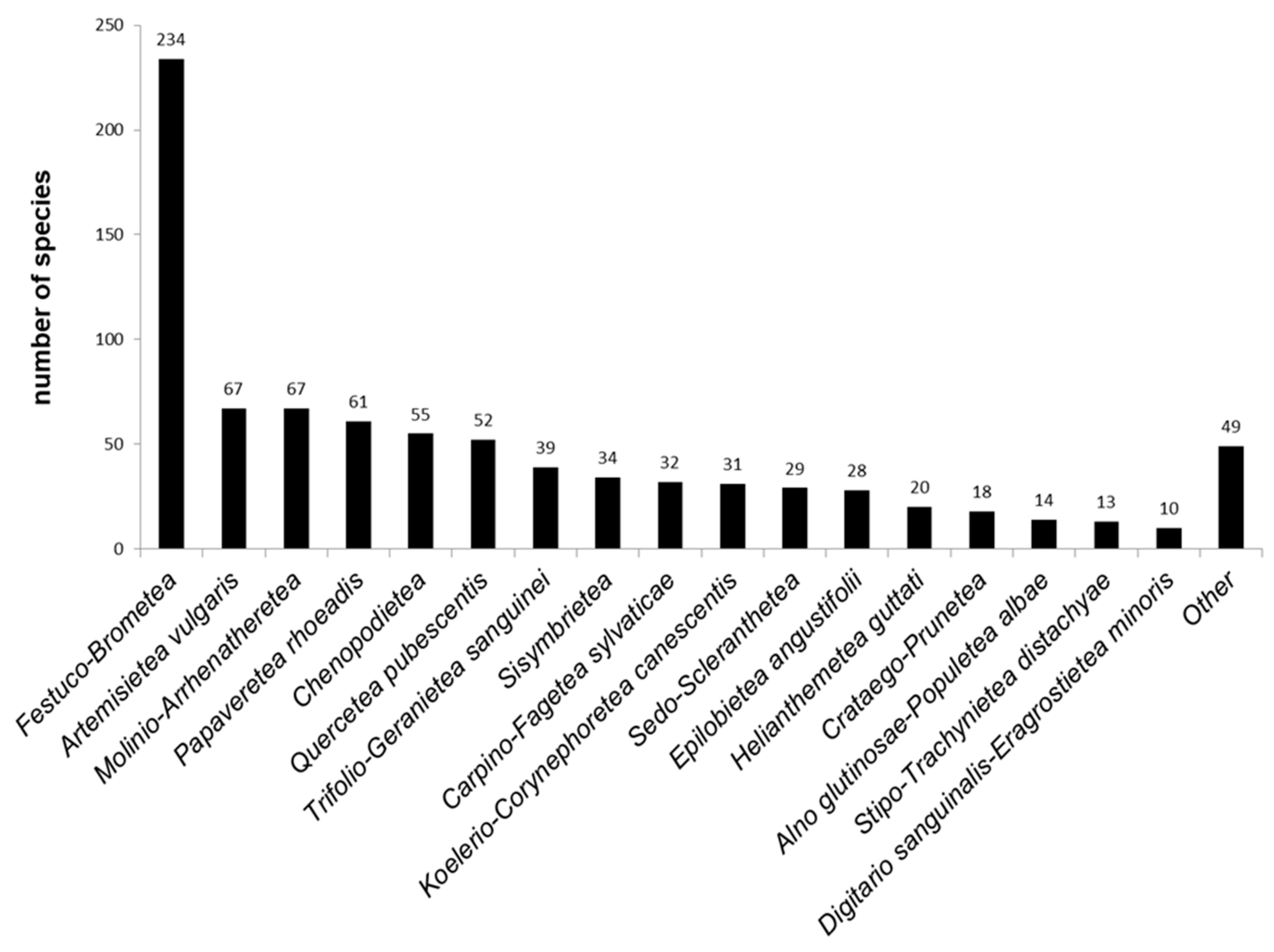
3.1. Diversity and Species Characteristics
3.1.1. Vascular Plants
3.1.2. Bryophytes
3.1.3. Lichens
4. Discussion
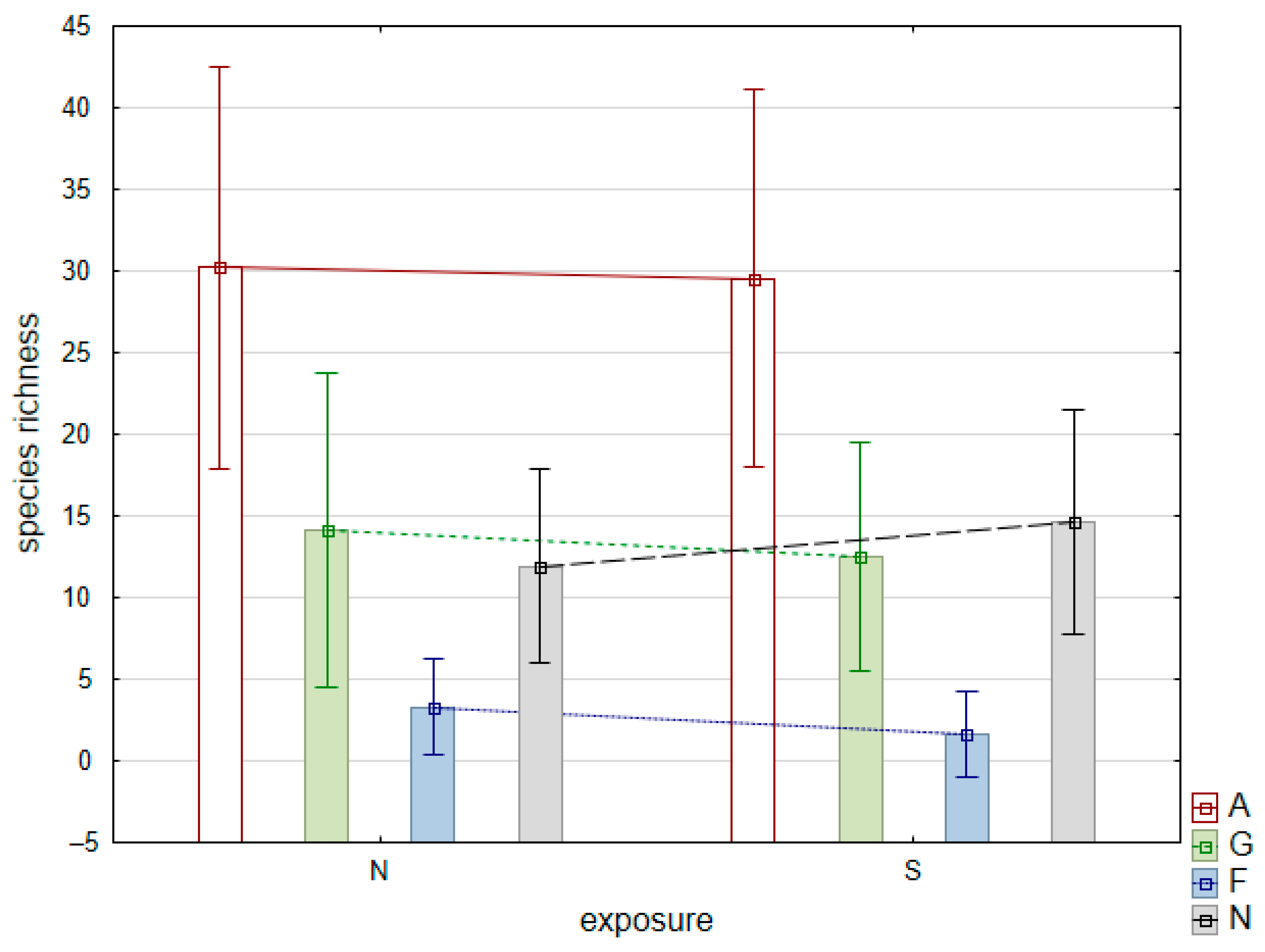
4.1. Species Diversity
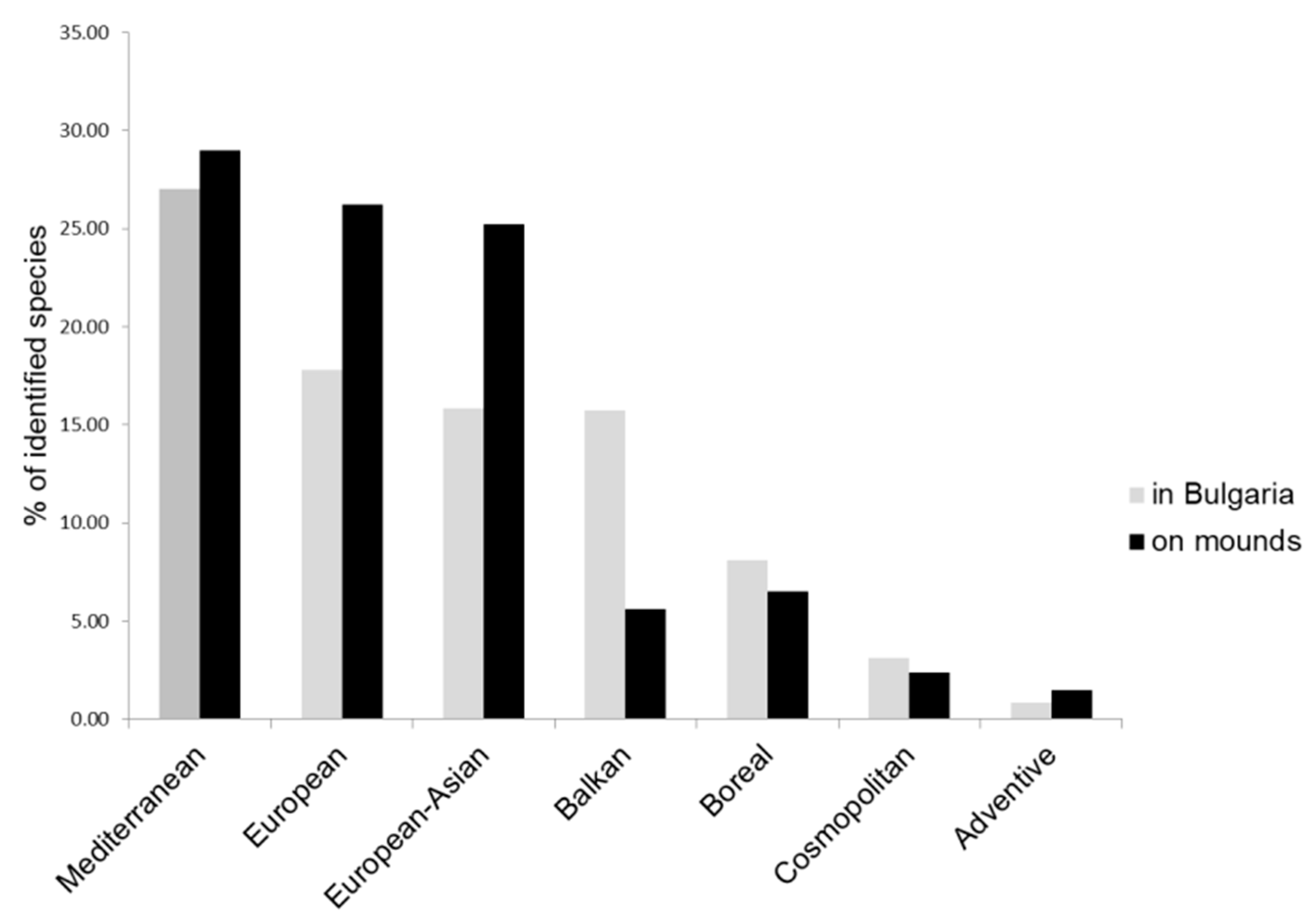
4.2. Species Characteristics
4.3. Significance for Nature Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sudnik-Wójcikowska, B.; Moysiyenko, I.I. Kurhany na “Dzikich Polach”—Dziedzictwo Kultury i Ostoja Ukraińskiego Stepu [Kurgans in the “Wild Field”—A Cultural Heritage and Refugium of the Ukrainian Steppe]; Wydawnictwa Uniwersytetu Warszawskiego: Warsaw, Poland, 2012. [Google Scholar]
- Deák, B. Nature and Culture: The Role of Ancient Burial Mounds in the Conservation of Eurasian Steppe Vegetation; Centre for Ecological Research: Tihany, Hungary, 2020; p. 172. [Google Scholar]
- Deák, B.; Tóthmérész, B.; Valkó, O.; Sudnik-Wójcikowska, B.; Moysiyenko, I.; Bragina, T.; Apostolova, I.; Dembicz, I.; Bykov, N.; Török, P. Cultural monuments and nature conservation: A review of the role of kurgans in the conservation and restoration of steppe vegetation. Biodivers. Conserv. 2016, 25, 2473–2490. [Google Scholar] [CrossRef]
- Apostolova, I.; Palpurina, S.; Sopotlieva, D.; Terziyska, T.; Velev, N.; Vassilev, K.; Nekhrizov, G.; Tsvetkova, N. Ancient Burial Mounds-Stepping Stones for Semi-Natural Habitats in Agricultural Landscape. Ecol. Balk. 2020, 12, 43–52. [Google Scholar]
- Kitov, G. The Thracian tumuli. Thracia 1993, 10, 39–80. [Google Scholar]
- Sudnik-Wójcikowska, B.; Moysiyenko, I.I. The Flora of Kurgans in the West Pontic Grass Steppe Zone of Southern Ukraine. Chornomorski Bot. J. 2006, 2, 14–44. Available online: http://eKhSUIR.kspu.edu/handle/123456789/2868 (accessed on 5 February 2022). [CrossRef]
- Dembicz, I.; Moysiyenko, I.; Shaposhnikova, A.; Vynokurov, D.; Kozub, L.; Sudnik-Wójcikowska, B. Isolation and patch size drive specialist plant species density within steppe islands: A case study of kurgans in southern Ukraine. Biodivers. Conserv. 2016, 25, 2289–2307. [Google Scholar] [CrossRef] [Green Version]
- Sudnik-Wójcikowska, B.; Moysiyenko, I.I. The floristic differentiation of microhabitats within kurgans in the desert steppe zone of southern Ukraine. Acta Soc. Bot. Pol. 2008, 77, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Deák, B.; Kovács, B.; Rádai, Z.; Apostolova, I.; Kelemen, A.; Kiss, R.; Lukács, K.; Palpurina, S.; Sopotlieva, D.; Báthori, F.; et al. Linking environmental heterogeneity and plant diversity: The ecological role of small natural features in homogeneous landscapes. Sci. Total Environ. 2021, 763, 144199. [Google Scholar] [CrossRef]
- European Commission. Green Infrastructure (GI)—Enhancing Europe’s Natural Capital, Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of Regions. 2013. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:52013DC0249 (accessed on 19 July 2021).
- Moysiyenko, I.; Zachwatowicz, M.; Sudnik-Wójcikowska, B.; Jabłońska, E. Kurgans help to protect endangered steppe species in the Pontic grass steppe zone, Ukraine. Wulfenia 2014, 21, 83–94. [Google Scholar]
- Sudnik-Wójcikowska, B.; Moysiyenko, I.I.; Zachwatowicz, M.; Jabłońska, E. The value and need for protection of kurgan flora in the anthropogenic landscape of steppe zone in Ukraine. Plant Biosyst. 2011, 145, 638–653. [Google Scholar] [CrossRef]
- Deák, B.; Tölgyesi, C.; Kelemen, A.; Bátori, Z.; Gallé, R.; Bragina, T.M.; Yerkin, A.I.; Valkó, O. The effects of micro-habitats and grazing intensity on the vegetation of burial mounds in the Kazakh steppes. Plant Ecol. Divers. 2017, 10, 509–520. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Nagy, D.D.; Török, P.; Torma, A.; Lőrinczi, G.; Kelemen, A.; Nagy, A.; Bede, Á.; Mizser, S.; et al. Habitat islands outside nature reserves—Threatened biodiversity hotspots of grassland specialist plant and arthropod species. Biol. Conserv. 2020, 241, 108254. [Google Scholar] [CrossRef]
- Dembicz, I.; Moysiyenko, I.I.; Kozub, Ł.; Dengler, J.; Zakharova, M.; Sudnik-Wójcikowska, B. Steppe islands in a sea of fields: Where island biogeography meets the reality of a severely transformed landscape. J. Veg. Sci. 2021, 32, e12930. [Google Scholar] [CrossRef]
- Duelli, P.; Obrist, M.K. In search of the best correlates for local organismal biodiversity in cultivated areas. Biodivers. Conserv. 1998, 7, 297–309. [Google Scholar] [CrossRef]
- Sudnik-Wójcikowska, B.; Moysiyenko, I.I. Ukrainian kurgans as refugia of steppe flora and their role in steppe restoration. In Steppenlebensräume Europas—Gefährdung, Erhaltungsmaßnahmen und Schutz; Baumbach, H., Pfützenreuter, S., Eds.; Thüringer Ministerium für Landwirtschaft, Forsten, Umwelt und Naturschutz: Erfurt, Germany, 2013; pp. 201–210. [Google Scholar]
- Deák, B.; Valkó, O.; Török, P.; Kelemen, A.; Bede, Á.; Csathó, A.I.; Tóthmérész, B. Landscape and habitat filters jointly drive richness and abundance of specialist plants in terrestrial habitat islands. Landsc. Ecol. 2018, 33, 1117–1132. [Google Scholar] [CrossRef]
- Czerepko, J.; Gawryś, R.; Szymczyk, R.; Pisarek, W.; Janek, M.; Haidt, A.; Kowalewska, A.; Piegdoń, A.; Stebel, A.; Kukwa, M.; et al. How sensitive are epiphytic and epixylic cryptogams as indicators of forest naturalness? Testing bryophyte and lichen predictive power in stands under different management regimes in the Białowieża forest. Ecol. Indic. 2021, 125, 107532. [Google Scholar] [CrossRef]
- Velev, S. Climatic regioning. In Geography of Bulgaria; Kopralev, I., Ed.; Publishing House ForCom: Sofia, Bulgaria, 2002; pp. 155–156. [Google Scholar]
- Bohn, U.; Golub, G.; Hettwer, C. (Eds.) Karte der Naturlichen Vegetation Europas. Legende/Map of the Natural Vegetation of Europe, Part 2: Legend (German/English); Bundesamt für Naturschutz: Bonn, Germany, 2003. [Google Scholar]
- Meshinev, T. Vegetation and phytogeography: A brief characteristic. In Biogeography and Ecology of Bulgaria; Monographie Biologicae; Fet, V., Popov, A., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 82, pp. 581–588. [Google Scholar]
- Tzonev, R.; Roussakova, V.; Dimitrov, M. The Western-Pontic steppe vegetation in Bulgaria. Hacquetia 2006, 5, 5–23. [Google Scholar]
- Euro+Med. Euro+Med—The Information Resource for Euro-Mediterranean Plant Diversity. 2006–2021. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 20 September 2021).
- The Plant List. Version 1.1. 2013. Available online: http://www.theplantlist.org/ (accessed on 1 September 2021).
- Hill, M.O.; Bell, N.; Bruggeman-Nannenga, M.A.; Brugués, M.; Cano, M.J.; Enroth, J.; Flatberg, K.I.; Frahm, J.-P.; Gallego, M.T.; Garilleti, R.; et al. An annotated checklist of the mosses of Europe and Macaronesia. J. Bryol. 2006, 28, 198–267. [Google Scholar] [CrossRef]
- Nimis, P.L.; Hafellner, J.; Roux, C.; Clerc, P.; Mayrhofer, H.; Martellos, S.; Bilovitz, P.O. The lichens of the Alps − an annotated checklist. MycoKeys 2018, 31, 1–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flora of Republic of Bulgaria; Academic Publishing “Prof. Marin Drinov”: Sofia, Bulgaria, 2012.
- Delipavlov, D.; Cheshmedzhiev, I.; Popova, M.; Terziyski, D.; Kovachev, I. Guide to Vascular Plants in Bulgaria; Academic Publishing of the Agricultural University: Plovdiv, Bulgaria, 2003. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Assyov, B.; Petrova, A. (Eds.) Conspectus of the Bulgarian Vascular Flora. Distribution Maps and Floristic Elements; Bulgarian Biodiversity Foundantion: Sofia, Bulgaria, 2012. [Google Scholar]
- Ganeva, A.; Düll, R. A contribution to the Bulgarian bryoflora. Checklist of Bulgarian bryophytes. In Contributions to the Bryoflora of Former Yugoslavia and Bulgaria; Düll, R., Ganeva, A., Martincic, A., Pavletic, Z., Eds.; IDH: Bad Münstereifel, Germany, 1999; pp. 111–199. [Google Scholar]
- Wirth, V. Die Flechten Baden-Württemmbergs; E. Ulmer GMBH Co: Stuttgart, Germany, 1995. [Google Scholar]
- Nimis, P.L. ITALIC—The Information System on Italian Lichens; Version 6.0; University of Trieste, Department of Biology: Berlin, Germany, 2016; Available online: http://dryades.units.it/italic (accessed on 1 August 2021).
- Petrova, A.; Vladimirov, V.; Georgiev, V. Invasive Alien Plant Species in Bulgaria; Institute of Biodiversity and Ecosystem Research, BAS: Sofia, Bulgaria, 2012. [Google Scholar]
- Petrova, A.; Vladimirov, V. Balkan endemics in the Bulgarian flora. Phytol. Balc. 2010, 16, 293–311. [Google Scholar]
- Petrova, A.; Vladimirov, V. Red list of Bulgarian vascular plants. Phytol. Balc. 2009, 15, 63–94. [Google Scholar]
- Biological Diversity Act. State Gazette, Sofia, Issue 77, August 2002, Last Change and Addition Issue 58, July 2016. Available online: http://eea.government.bg/bg/legislation/biodiversity/ZBR_en_26_07_2016.pdf (accessed on 27 October 2021).
- Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. OJ L 206. 22 July 1992, pp. 7–50. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:31992L0043 (accessed on 22 February 2021).
- Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES), 1973. Signed at Washington, D.C.; on 3 March 1973, Amended at Bonn, on 22 June 1979, Amended at Gaborone, on 30 April 1983. Available online: https://checklist.cites.org/#/en (accessed on 1 September 2021).
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- PRIMER-E Ltd. PRIMER (Plymouth Routines In Multivariate Ecological Research), Version 7. 2017. Available online: http://www.primer-e.com (accessed on 19 December 2021).
- Hammer, Ø.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4. Available online: https://www.nhm.uio.no/english/research/infrastructure/past/ (accessed on 18 January 2022).
- Dell Inc. Dell Statistica (Data Analysis Software System), Version 13. 2016. Available online: http://www.dell.com (accessed on 18 January 2022).
- Peev, D.; Delcheva, M. Florogeographical elements and evolutionary trends in the Bulgarian Flora. In Biogeography and Ecology of Bulgaria; Monographie Biologicae; Fet, V., Popov, A., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 82, pp. 571–579. [Google Scholar]
- Arnaudov, N. The Moss Flora of Vitosha Mt; Ann. de l’Universite de Sofia: Sofia, Bulgaria, 1908; Volume 3–4, pp. 1–37. [Google Scholar]
- Petrova, A.; Vladimirov, V. Recent progress in floristic and taxonomic studies in Bulgaria. Bot. Serb. 2018, 42, 35–69. [Google Scholar] [CrossRef]
- Bartha, D.; Király, G.; Schmidt, D.; Tiborcz, V.; Barinza, Z.; Csiky, J.; Jakab, G.; Lesku, B.; Schmotzer, A.; Vidéki, R.; et al. (Eds.) Magyarország Edényes Növényfajainak Elterjedési Atlasza [Distribution Atlas of Vascular Plants of Hungary; Nyugat-magyarországi Egyetem Kiadó/University of West Hungary: Sopron, Hungary, 2015. [Google Scholar]
- Mosyakin, S.L.; Fedoronchuk, N.M. Vascular Plants of Ukraine: A Nomenclatural Checklist; National Academy of Sciences of Ukraine-MG Kholodny Institute of Botany: Kyiv, Ukraine, 1999. [Google Scholar]
- Sudnik-Wójcikowska, B.; Moysiyenko, I. Zonal character of the flora of kurgans in central and southern Ukraine. Biodiv. Res. Conserv. 2010, 17, 47–52. [Google Scholar] [CrossRef]
- Barczi, A. Data for the botanical and pedological surveys of the Hungarian kurgans (Great Hungarian Plain, Hortobágy). Thaiszia J. Bot. 2003, 13, 113–126. [Google Scholar]
- Cwener, A. Ros´liny naczyniowe kurhano´w w dorzeczu dolnej Szreniawy i Nidzicy (Wy_zyna Małopolska, południowa Polska) [The vascular plants of the mounds in the lower Szreniawa and Nidzica river-basin (Małopolska Upland, South Poland)]. Fragm. Florist. Geobot. Pol. 2004, 11, 27–40. [Google Scholar]
- Deák, B.; Valkó, O.; Török, P.; Tóthmérész, B. Factors threatening grassland specialist plants—A multi-proxy study on the vegetation of isolated grasslands. Biol. Conserv. 2016, 204, 255–262. [Google Scholar] [CrossRef]
- Erdős, L.; Méri, A.; Bátori, Z.; Gallé, R.; Kőrmőczi, L. North–south facing vegetation gradients in the Villány Mountains: A case study on the population and the community level. Pak. J. Bot. 2012, 44, 927–932. [Google Scholar]
- Batič, F.; Primožič, K.; Surina, B.; Trošt, T.; Mayrhofer, H. Contributions to the lichen flora of Slovenia X. Lichens from the Slovenian Julian Alps. Herzogia 2003, 16, 143–154. [Google Scholar]
- Coppins, B.J.; Orange, A. Arthopyrenia A. Massal. In The lichens of Great Britain and Ireland, 2nd ed.; Smith, C.W., Aptroot, A., Coppins, B.J., Fletcher, A., Gilbert, O.L., James, P.W., Wolseley, P.A., Eds.; The British Lichen Society: London, UK, 2009; pp. 136–138. [Google Scholar]
- Richardson, D.H.S. Pollution Monitoring with Lichens. Naturalists’ Handbooks 19; The Richmond Publishing: Slough, UK, 1992. [Google Scholar]
- van Haluwyn, C.; van Herk, C.M. Bioindication: The community approach. In Monitoring with Lichens—Monitoring Lichens; Nimis, P.L., Scheidegger, C., Wolseley, P., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 2002; pp. 39–64. [Google Scholar]
- Valkó, O.; Zmihorski, M.; Biurrun, I.; Loos, J.; Labadessa, R.; Venn, S. Ecology and conservation of steppes and semi-natural grasslands. Hacquetia 2016, 15, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Teleki, B.; Sonkoly, J.; Erdős, L.; Tóthmérész, B.; Prommer, M.; Török, P. High resistance of plant biodiversity to moderate native woody encroachment in loess steppe grassland fragments. Appl. Veg. Sci. 2020, 23, 175–184. [Google Scholar] [CrossRef]
- Boch, S.; Müller, J.; Prati, D.; Fischer, M. Low-intensity management promotes bryophyte diversity in grasslands. Tuexenia 2018, 38, 311–328. [Google Scholar] [CrossRef]
- Chytrý, M.; Pyšek, P.; Tichý, L.; Knollová, I.; Danihelka, J. Invasions by alien plants in the Czech Republic: A quantitative assessment across habitats. Preslia 2005, 77, 339–354. [Google Scholar]
- Celesti-Grapow, L.; Blasi, C. The Role of Alien and Native Weeds in the Deterioration of Archaeological Remains in Italy. Weed Technol. 2004, 18, 1508–1513. [Google Scholar]
- Sudnik-Wójcikowska, B.; Moysiyenko, I. The synanthropic flora of kurgans within three steppe zones in southern Ukraine. Biodiv. Res. Conserv. 2008, 11–12, 41–48. [Google Scholar]
- CICES—The Common International Classification of Ecosystem Services. Available online: https://cices.eu (accessed on 18 January 2022).
- Matsler, M.; Meerow, S.; Mell, I.; Pavao-Zuckerman, M. A ‘green’ chameleon: Exploring the many disciplinary definitions, goals, and forms of “green infrastructure”. Landsc. Urban Plan. 2021, 214, 104145. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Min | Max | Mean | SD |
|---|---|---|---|---|
| Altitude (m, a.s.l.) | 61.0 | 920.0 | 354.4 | 238.8 |
| Height (m) | 1.2 | 24.5 | 5.3 | 3.1 |
| Diameter (m) | 9.5 | 88.4 | 40.7 | 15.8 |
| Coverage of herbaceous vegetation (%) | 0 | 100 | 78.3 | 30.5 |
| Coverage of shrub/forest vegetation (%) | 0 | 100 | 21.7 | 30.5 |
| Surrounding of semi-natural vegetation in 200 m buffer (%) | 0 | 100 | 23.1 | 26.4 |
| Surrounding of agricultural and other anthropogenic lands in 200 m buffer (%) | 0 | 100 | 76.9 | 26.4 |
| On a Mound | |||||
|---|---|---|---|---|---|
| Parameter | Total | Min | Max | Mean | SD |
| Biological type | |||||
| Short-lived | 378 | 4 | 64 | 24.77 | 10.90 |
| Perennial | 495 | 10 | 88 | 36.13 | 13.57 |
| Dwarf-shrub | 17 | 0 | 7 | 0.80 | 1.09 |
| Shrub | 49 | 0 | 12 | 4.02 | 2.80 |
| Tree | 35 | 0 | 7 | 2.55 | 1.87 |
| N/A | 85 | 0 | 10 | 2.20 | 2.26 |
| Plants with conservation importance | |||||
| Critically Endangered (CR) | 2 | 0 | 1 | 0.04 | 0.19 |
| Endangered (EN) | 9 | 0 | 2 | 0.22 | 0.43 |
| Vulnerable (VU) | 7 | 0 | 2 | 0.15 | 0.39 |
| Near Threatened (NT) | 6 | 0 | 1 | 0.07 | 0.26 |
| Least concern (LC) | 2 | 0 | 2 | 0.21 | 0.45 |
| Bulgarian Biodiversity Act | 10 | ||||
| Habitat Directive | 1 | ||||
| CITES | 11 | ||||
| N/A | 85 | 0 | 10 | 2.20 | 2.26 |
| No conservation status | 947 | 27 | 137 | 67.50 | 20.87 |
| Native status | |||||
| Native (including archaeophytes) | 948 | 27 | 141 | 66.76 | 21.30 |
| Naturalized alien | 5 | 0 | 2 | 0.15 | 0.39 |
| Doubtfully native | 1 | ||||
| In large-scale cultivation | 2 | ||||
| Alien (status unknown) | 13 | 0 | 5 | 0.83 | 0.97 |
| N/A | 85 | 0 | 10 | 2.20 | 2.26 |
| No data | 5 | 0 | 2 | 0.45 | 0.63 |
| Invasive alien plants (for Bulgaria) | |||||
| Invasive alien plants (IAP) | 12 | 0 | 4 | 0.50 | 0.80 |
| Not classified as IAP | 962 | 27 | 142 | 67.76 | 21.38 |
| N/A | 85 | 0 | 10 | 2.20 | 2.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apostolova, I.; Sopotlieva, D.; Valcheva, M.; Ganeva, A.; Shivarov, V.; Velev, N.; Vassilev, K.; Terziyska, T.; Nekhrizov, G. First Survey of the Vascular and Cryptogam Flora on Bulgaria’s Ancient Mounds. Plants 2022, 11, 705. https://doi.org/10.3390/plants11050705
Apostolova I, Sopotlieva D, Valcheva M, Ganeva A, Shivarov V, Velev N, Vassilev K, Terziyska T, Nekhrizov G. First Survey of the Vascular and Cryptogam Flora on Bulgaria’s Ancient Mounds. Plants. 2022; 11(5):705. https://doi.org/10.3390/plants11050705
Chicago/Turabian StyleApostolova, Iva, Desislava Sopotlieva, Magdalena Valcheva, Anna Ganeva, Veselin Shivarov, Nikolay Velev, Kiril Vassilev, Tsvetelina Terziyska, and Georgi Nekhrizov. 2022. "First Survey of the Vascular and Cryptogam Flora on Bulgaria’s Ancient Mounds" Plants 11, no. 5: 705. https://doi.org/10.3390/plants11050705