
Effect of the Soil Matric Potential on the Germination Capacity of Prosopis chilensis, Quillaja saponaria and Cryptocarya alba from Contrasting Geographical Origins
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
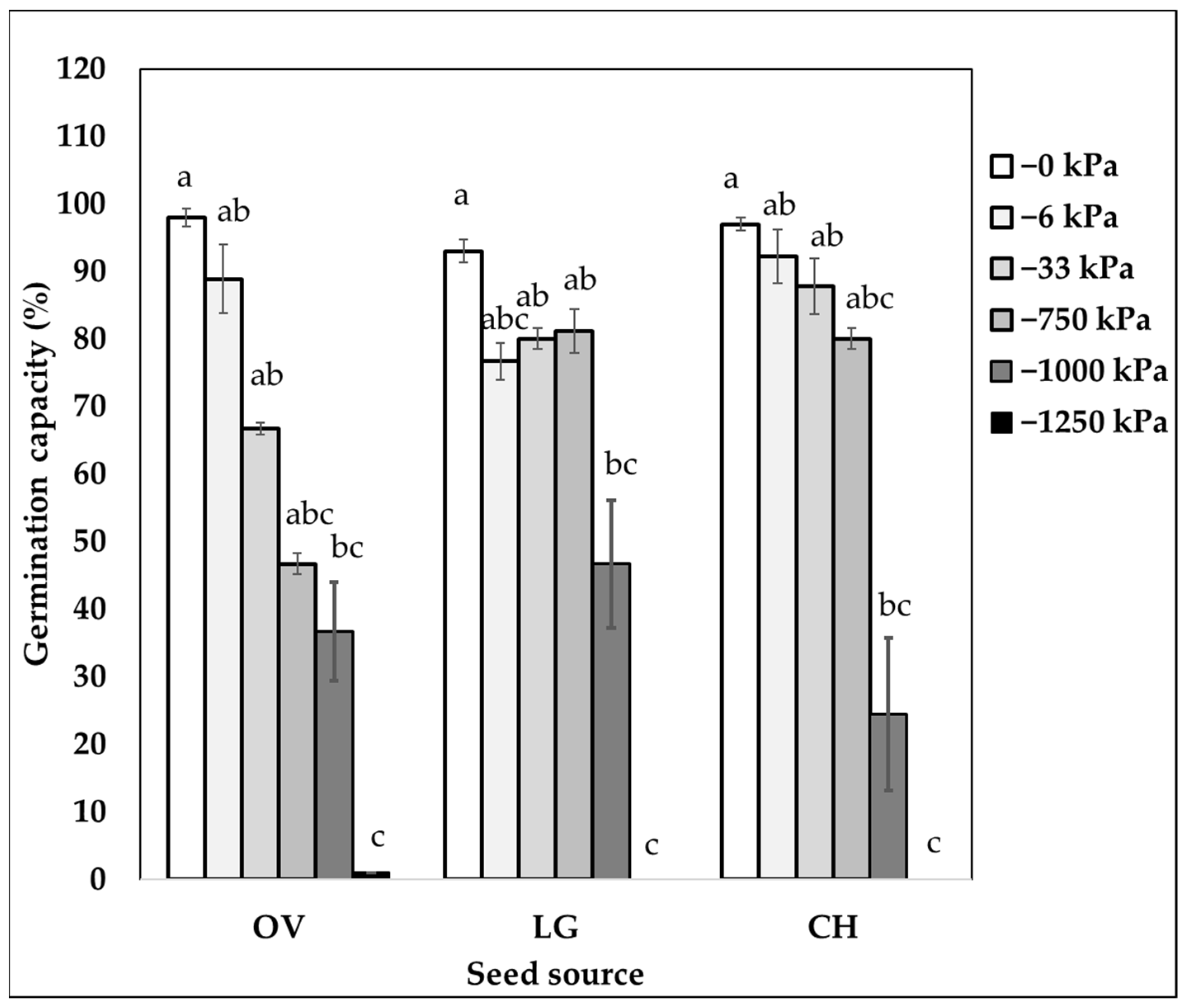
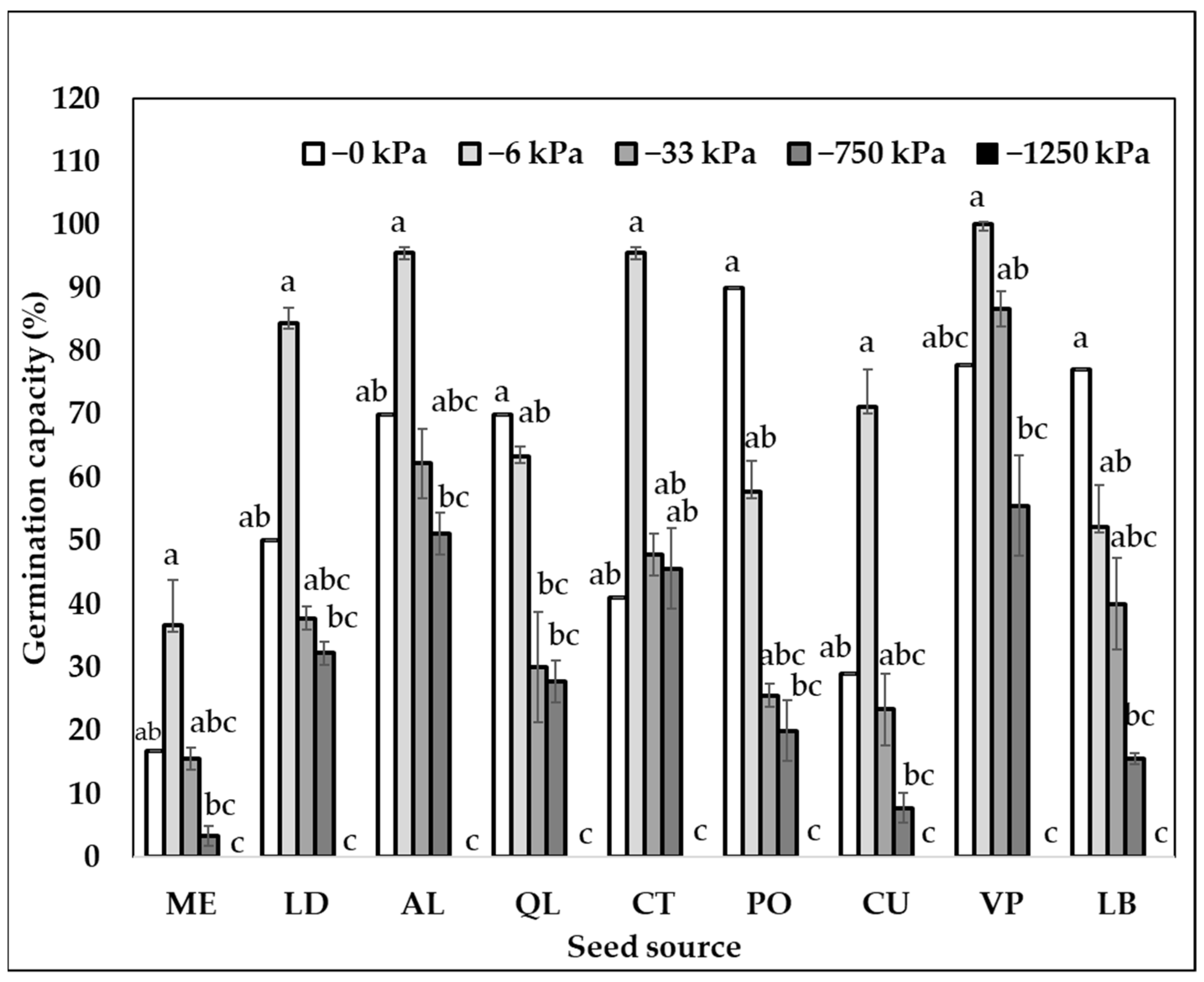
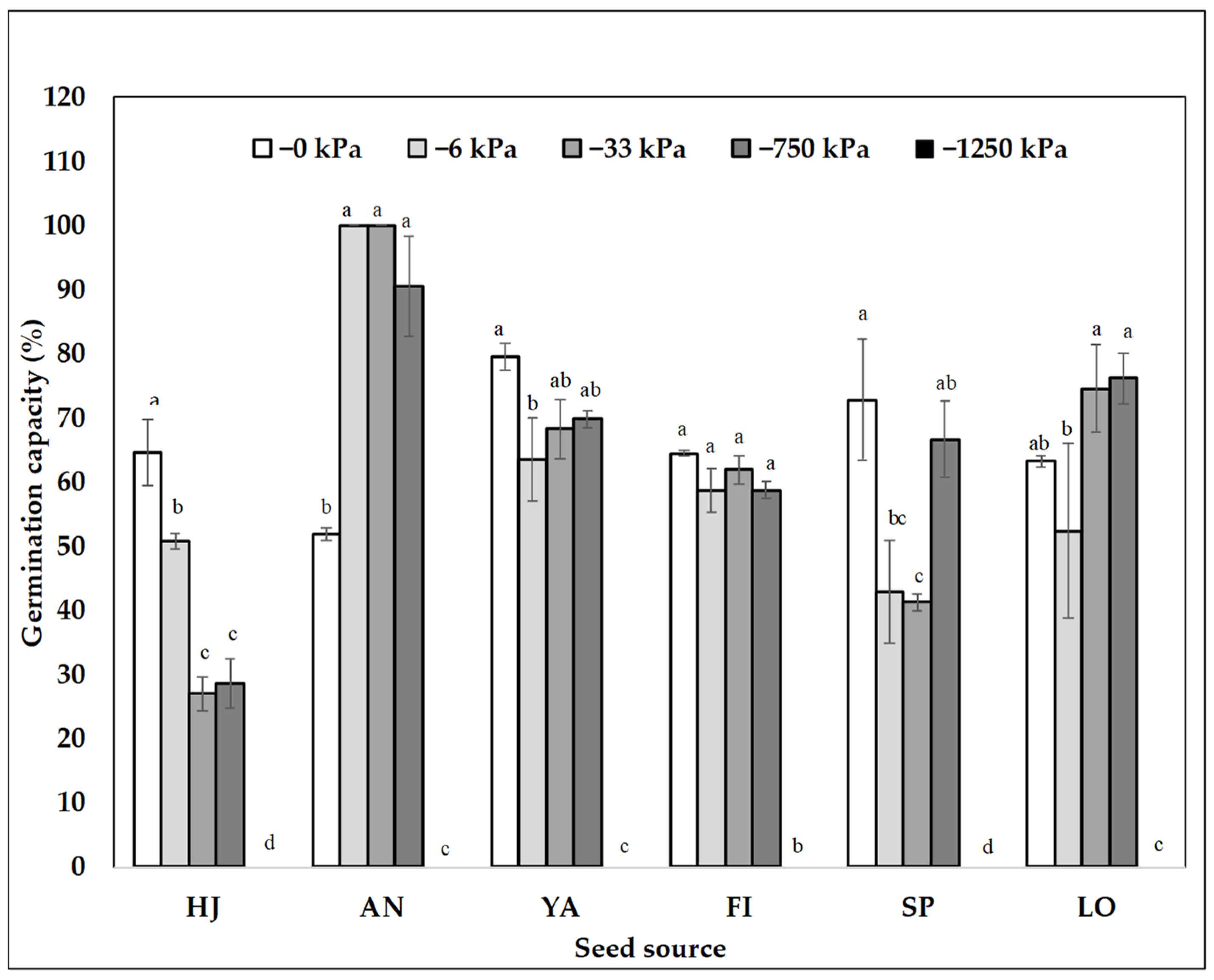
2.1. Germination Capacity at Different Seed Sources and Matric Potentials
2.2. Critical Soil Matric Potential for Germination
2.3. Environmental Factors Influencing the Germination Capacity
3. Discussion
4. Materials and Methods
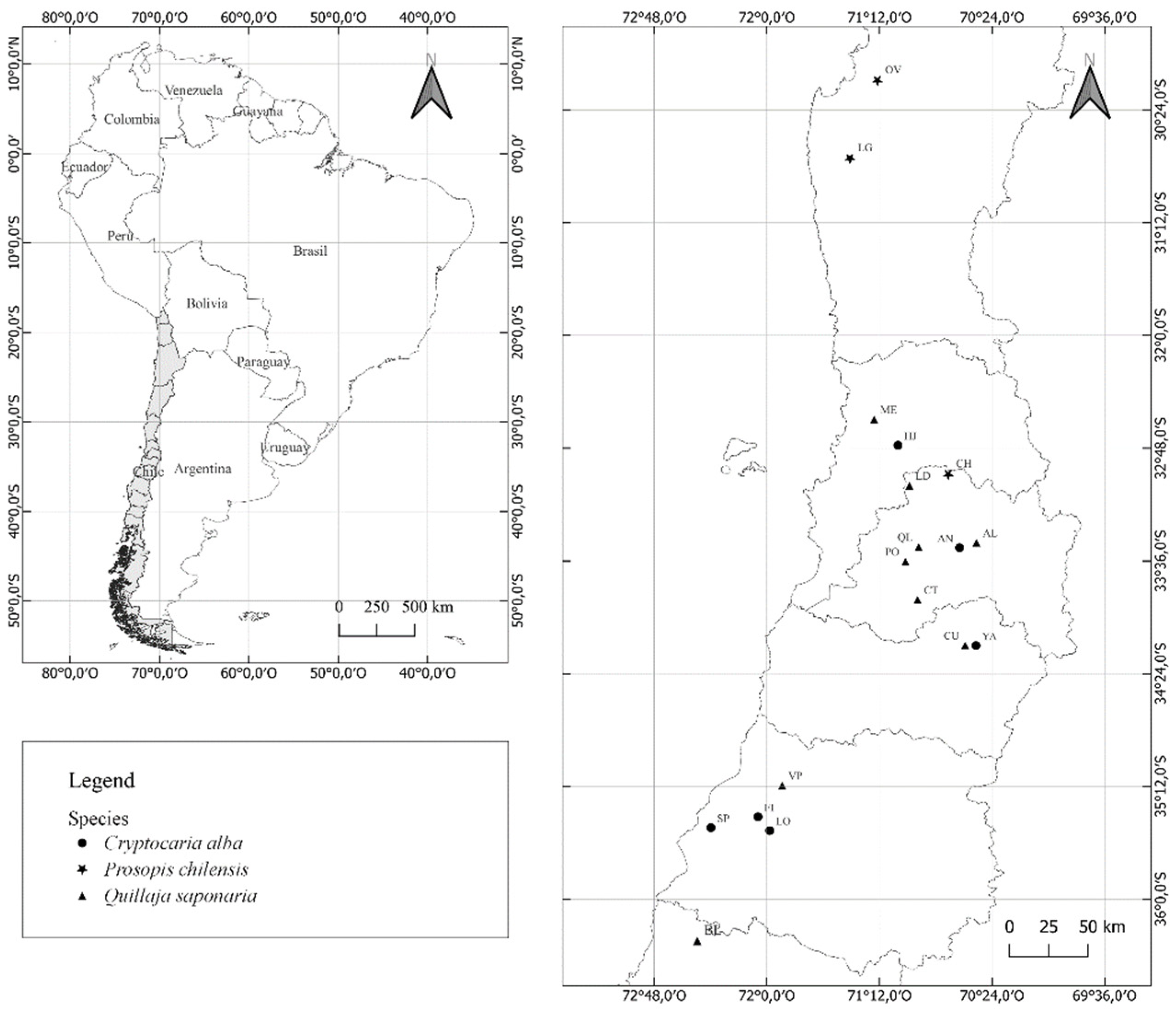
4.1. Seed Collection and Processing
4.1.1. Substrate Preparation, Water Retention Curves, and Matric Potential Treatments
4.1.2. Experimental Design
4.2. Measurements
4.3. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lloret, F.; Escudero, A.; Iriondo, J.M.; Martínez-Vilalta, J.; Valladares, F. Extreme climatic events and vegetation: The role of stabilizing processes. Glob. Chang. Biol. 2011, 18, 797–805. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J. Global Change and Forest Disturbances in the Mediterranean Basin: Breakthroughs, Knowledge Gaps, and Recommendations. Forests 2021, 12, 603. [Google Scholar] [CrossRef]
- Bambach, N.; Meza, F.J.; Gilabert, H.; Miranda, M. Impacts of climate change on the distribution of species and communities in the Chilean Mediterranean ecosystem. Reg. Environ. Chang. 2013, 13, 1245–1257. [Google Scholar] [CrossRef]
- Valladares, F.E.; Benavides, R.A.; Rabasa, S.G.; Díaz, M.A.; Pausas, J.G.; Paula, S.U.; Simonson, W.D. Global change and Mediterranean forests: Current impacts and potential responses. In Forests and Global Change; Cambridge University Press: Cambridge, UK, 2014; pp. 47–75. [Google Scholar]
- Penuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.; Llusià, J.; Ogaya, R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O.; et al. Impacts of Global Change on Mediterranean Forests and Their Services. Forests 2017, 8, 463. [Google Scholar] [CrossRef] [Green Version]
- Miranda, A.; Lara, A.; Altamirano, A.; Di Bella, C.; González, M.E.; Camarero, J.J. Forest browning trends in response to drought in a highly threatened mediterranean landscape of South America. Ecol. Indic. 2020, 115, 106401. [Google Scholar] [CrossRef]
- Togenetti, R.; Smith, M.; Panzacchi, P. Climate-Smart Forestry in Mountain Regions. Managing Forest Ecosystems; Springer: Berlin/Heidelberg, Germany, 2022; p. 587. [Google Scholar] [CrossRef]
- Bannister, J.R. Recuperar bosques no es solo plantar árboles: Lecciones aprendidas luego de 7 años restaurando bosques de Pilgerodendron uviferum (D. Don) florin en chiloé. In Anales del Instituto de la Patagonia; Universidad de Magallanes: Punta Arenas, Chile, 2015; Volume 43, pp. 35–51. [Google Scholar]
- Fuentes-Castillo, T.; Hernández, H.J.; Pliscoff, P. Hotspots and ecoregion vulnerability driven by climate change velocity in Southern South America. Reg. Environ. Chang. 2020, 20, 27. [Google Scholar] [CrossRef]
- Pulido, F.; Díaz, M. Regeneration of a Mediterranean oak: A whole-cycle approach. Écoscience 2005, 12, 92–102. [Google Scholar] [CrossRef]
- Rodrigo, A.; Retana, J.; Picó, F.X. Direct regeneration is not the only response of Mediterranean forests to large fires. Ecology 2004, 85, 716–729. [Google Scholar] [CrossRef]
- Buhk, C.; Meyn, A.; Jentsch, A. The challenge of plant regeneration after fire in the Mediterranean Basin: Scientific gaps in our knowledge on plant strategies and evolution of traits. Plant Ecol. 2006, 192, 1–19. [Google Scholar] [CrossRef]
- Becerra, P.; Smith-Ramírez, C.Y.; Arellano, E. Evaluación de Técnicas Pasivas y Activas para la Recuperación del Bosque Esclerófilo de Chile Central; Corporación Nacional Forestal: Santiago, Chile, 2018; p. 44. ISBN 9789561423374. [Google Scholar]
- Eriksson, O.; Ehrlén, J. Seedling recruitment and population ecology. In Seedling Ecology and Evolution; Alessio, M., Parker, V., Simpson, R., Eds.; Cambridge University Press: New York, NY, USA, 2008; pp. 239–254. [Google Scholar]
- Padilla, F. Factores limitantes y estrategias de establecimiento de plantas leñosas en ambientes semiáridos. Implicaciones para la restauración. Ecosistemas 2008, 17, 155–159. [Google Scholar]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.; Aukema, B.H.; Bond-Lamberty, B.; Chini, L.; Clark, J.S.; Dietze, M.; Grossiord, C.; Hanbury-Brown, A.; et al. Pervasive shifts in forest dynamics in a changing world. Science 2020, 368, eaaz9463. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Centro de Ciencia del Clima y la Resiliencia (CR2). La Megasequía 2010–2015: Una Lección para el Futuro; Centro de Ciencia del Clima y la Resiliencia: Santiago, Chile, 2015; p. 27. [Google Scholar]
- Garreaud, R.D.; Alvarez-Garreton, C.; Barichivich, J.; Boisier, J.P.; Christie, D.; Galleguillos, M.; LeQuesne, C.; McPhee, J.; Zambrano-Bigiarini, M. The 2010–2015 megadrought in central Chile: Impacts on regional hydroclimate and vegetation. Hydrol. Earth Syst. Sci. 2017, 21, 6307–6327. [Google Scholar] [CrossRef] [Green Version]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010–2018): A climate dynamics perspective. Int. J. Clim. 2020, 40, 421–439. [Google Scholar] [CrossRef]
- Fischlin, A.; Midgley, G.F.; Price, J.T. Ecosystems, their properties, goods, and services. In Climate Change 2007: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., Van Der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 211–272. [Google Scholar]
- Kolström, M.; Lindner, M.; Vilén, T.; Maroschek, M.; Seidl, R.; Lexer, M.J.; Netherer, S.; Kremer, A.; Delzon, S.; Barbati, A.; et al. Reviewing the science and implementation of climate change adaptation measures in European forestry. Forests 2011, 2, 961–982. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.; Silva, H.R.; Garrido, M.; Acevedo, E. Manual de Studio y Ejercicios Relacionados con el Contenido de Agua en el Suelo y su uso por los Cultivos; Universidad de Chile: Santiago, Chile, 2015; p. 86. [Google Scholar]
- Yadvinder-Singh; Surinder, S.K.; Magni, L.J.; Harminder, S.S. Improving Water Productivity of Wheat-Based Cropping Systems in South Asia for Sustained Productivity. Adv. Agron. 2014, 127, 157–258. [Google Scholar]
- Gao, C.; Liu, F.; Zhang, C.; Feng, D.; Li, K.; Cui, K. Germination responses to water potential and temperature variation among provenances of Pinus yunnanensis. Flora 2021, 276–277, 151786. [Google Scholar] [CrossRef]
- Rodríguez Rivera, M.F.; Sosa, L.R.; Fernández, E.A.; Reale, M.I.; Villarreal, V. Efecto del estrés hídrico a distintas temperaturas sobre la germinación de semillas de Bulnesia retama (Gill. ex. Hook.) Griseb. -Zigofiláceas-en San Luis, Argentina. Phyton 2007, 76, 5–17. [Google Scholar] [CrossRef]
- Santelices, R.; Espinoza, S.; Magni, C.; Cabrera, A.; Donoso, S.; Peña, K. Variability in seed germination and seedling growth at the intra- and inter-provenance levels of Nothofagus glauca (Lophozonia glauca), an endemic species of Central Chile. N. Z. J. For. Sci. 2017, 47, 10. [Google Scholar] [CrossRef] [Green Version]
- Corporación Nacional Forestal (CONAF). Catastro Vegetacional. 2017. Available online: http://www.conaf.cl/nuestros-bosques/bosques-en-chile/catastro-vegetacional/ (accessed on 17 October 2021).
- Rodríguez, R.; Matthei, O.; Quezada, M. Flora arbórea de Chile; Editorial de la Universidad de Concepción: Concepción, Chile; Instituto Nacional Forestal: Los Lagos, Chile, 1983; p. 408. [Google Scholar]
- Varela, D.; Arana, V. Latencia y Germinación de Semillas. Tratamientos Pregerminativos; Serie técnica: “Sistemas Forestales Integrados” Área Forestal; INTA EEA Bariloche: San Carlos de Bariloche, Argentina, 2011; p. 10. [Google Scholar]
- Figueroa, J.A.; Jaksic, F.M. Latencia y banco de semillas en plantas de la región mediterránea de Chile central. Rev. Chil. Hist. Nat. 2004, 77, 201–215. [Google Scholar] [CrossRef]
- Matilla, A.J. Desarrollo y germinación de las semillas. In Fundamentos de Fisiología Vegetal, 2nd ed.; Azcón-Bieto, J., Talón, M., Eds.; McGraw Hill: New York, NY, USA, 2007; p. 23. [Google Scholar]
- Bonner, F.T.; Vozzo, J.A.; Elam, W.W.; Land, S.B., Jr. Tree Seed Technology Training Course: Instructor’s Manual; SO-106; USDA Forest Service, Southern Forest Experiment Station: Asheville, NC, USA, 1994; p. 160. [Google Scholar]
- Ibañez, C.; Jamett, F. Establecimiento de Protocolos de Micropropagación e Identificación de Bioproductos en Especies Nativas de alto valor Ecológico del Norte Chico Este Proyecto fue Financiado Por el Fondo de Investigación del Bosque Nativo; proyecto N° 037/2011; Fondo de Investigación del Bosque Nativo: Santiago, Chile, 2013; p. 93. [Google Scholar]
- Cabello, A. Propagación de especies nativas: Aspectos claves. In Avance en Silvicultura, Primera Reunión. Programa y Resúmenes; Depto. De Silvicultura, Facultad de Ciencias Agrarias y Forestales, Universidad de Chile: Santiago, Chile, 1993. [Google Scholar]
- Benedetti, S. Monografía de Peumo, Cryptocarya alba (Mol.) Looser. In Programa de Investigación de Productos Forestales No Madereros; Instituto Forestal (INFOR): Santiago, Chile, 2012. [Google Scholar]
- Acuña, M. Formulación de un Protocolo de Trabajo para el Análisis de Semillas de Especies Leñosas Nativas; Memory Professional Title of Forestry Engineer; Universidad de Chile, Facultad de Ciencias Forestales: Santiago, Chile, 2001; p. 87. [Google Scholar]
- Magni, C.R.; Saavedra, N.; Espinoza, S.E.; Yáñez, M.A.; Quiroz, I.; Faúndez, Á.; Grez, I.; Martinez-Herrera, E. The Recruitment of the Recalcitrant-Seeded Cryptocarya alba (Mol.) Looser, Established via Direct Seeding Is Mainly Affected by the Seed Source and Forest Cover. Plants 2022, 11, 2918. [Google Scholar] [CrossRef]
- Grime, J.P.; Mason, G.; Curtis, A.V.; Rodman, J.; Band, S.R. A Comparative Study of Germination Characteristics in a Local Flora. J. Ecol. 1981, 69, 1017. [Google Scholar] [CrossRef]
- Roach, D.A.; Wulff, R.D. Maternal effects in plants. Annu. Rev. Ecol. Syst. 1987, 18, 209–235. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic: San Diego, Chile, 2014; ISBN 978-0120802630. [Google Scholar]
- Meyer, S.E.; Kitchen, S.G.; Carlson, S.L. Seed Germination Timing Patterns in Intermountain Penstemon (Scrophulariaceae). Am. J. Bot. 1995, 82, 377–389. [Google Scholar] [CrossRef]
- Rodríguez Araujo, M.E.; Milano, C.; Pérez, D.R. Germination of Ephedra ochreata Miers: Contribution for productive restoration of arid environments in Argentina. Agrociencia 2019, 53, 617–629. [Google Scholar]
- Gutiérrez-Vázquez, B.N.; Flores-Montaño, A. Rodales semilleros: Opción para la conservación in situ de recursos genéticos forestales en Chiapas, México. For. Veracruzana 2017, 19, 41–48. [Google Scholar]
- Donoso, C.; Cabello, A. Antecedentes fenológicos y de germinación de especies leñosas chilenas. Cienc. For. 1978, 1, 31–41. [Google Scholar]
- Magni, C.; Espinoza, S.; Garrido, E.; Santelices, R.; Cabrera, A. Differential adaptations in nursery seedlings from diverse Chilian provenances of Peumus boldus Mol. iFor.-Biogeosci. For. 2016, 9, 409–413. [Google Scholar] [CrossRef] [Green Version]
- SSF, Servicio de Sanidad Forestal y Equilibrios Biológicos. Red de Seguimiento a Gran Escala de Daños en los Montes (Red de Nivel I). Manual de Campo; Ministerio de Agricultura, Alimentación y Medio Ambiente: Barcelona, España, 2012. [Google Scholar]
- Kidson, R.; Westoby, M. Seed mass and seedling dimensions in relation to seedling establishment. Oecologia 2000, 125, 11–17. [Google Scholar] [CrossRef]
- Baloch, H.A.; Ditommaso, A.; Watson, A.K. Intrapopulation variation in Abutilon theophrasti seed mass and its relationship to seed germinability. Seed Sci. Res. 2001, 11, 333–345. [Google Scholar] [CrossRef]
- Ayala-Cordero, G.; Terraza, T.; López-Mata, L.; Trejo, C. Variación en el tamaño y peso de la semilla y su relación con la germinación en una población de Stenocereus beneckei. Interciencia 2004, 29, 692–697. [Google Scholar]
- Hall, L.S.; Krausman, P.R.; Morrison, M.L. The habitat concept and a plea for standard terminology. Wildl. Soc. Bull. 1997, 25, 173–182. [Google Scholar]
- Hechenleitner, P.; Gardner, M.; Thomas, P.; Echeverría, C.; Escobar, B.; Brownless, P.; Martínez, C. Plantas Amenazadas del Centro-sur de Chile. Distribución, Conservación y Propagación; Universidad Austral de Chile: Valdivia, Chile; Real Jardín de Edimbrugo: Edinburgh, UK, 2005; p. 188. [Google Scholar]
- Mccaugey-Espinoza, D.; Ayala, G.; Garcia, C.; Buitemea, C.; Ochoa-Meza, A. Germinación in vitro e inducción de callo y raíz en Bursera laxiflora S. Watson. Abanico Agrofor. 2021, 2, 1–14. [Google Scholar] [CrossRef]
- Montserrat-Martí, G.; Camarero, J.J.; Palacio, S.; Pérez-Rontomé, C.; Milla, R.; Albuixech, J.; Maestro, M. Summer-drought constrains the phenology and growth of two coexisting Mediterranean oaks with contrasting leaf habit: Implications for their persistence and reproduction. Trees 2009, 23, 787–799. [Google Scholar] [CrossRef] [Green Version]
- Morin, X.; Roy, J.; Sonié, L.; Chuine, I. Changes in leaf phenology of three European oak species in response to experimental climate change. New Phytol. 2010, 186, 900–910. [Google Scholar] [CrossRef]
- Magni, C.; Espinoza, S.; Poch, P.; Abarca, B.; Grez, I.; Martínez, E.; Yáñez, M.; Santelices, R.; Cabrera, A. Growth and biomass partitioning of nine provenances of Quillaja saponaria seedlings to water stress. South. For. J. For. Sci. 2018, 81, 103–109. [Google Scholar] [CrossRef]
- Del Río, M.; Pretzsch, H.; Bončina, A.; Avdagić, A.; Bielak, K.; Binder, F.; Coll, L.; Hilmers, T.; Höhn, M.; Kašanin-Grubin, M.; et al. Carper 3. Assessment of Indicators for Climate Smart Management in Mountain Forests, pp. 59–107. In Climate-Smart Forestry in Mountain Regions. Managing Forest Ecosystems; Togenetti, R., Smith, M., Panzacchi, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2022; p. 587. [Google Scholar]
- Kairis, O.; Karavitis, C.; Salvati, L.; Kounalaki, A.; Kosmas, K. Exploring the impact of overgrazing on soil erosion and land degradation in a dry Mediterranean agro-forest landscape. Arid Land Res. Manag. 2015, 29, 360–374. [Google Scholar] [CrossRef]
- Atlas Agroclimático de Chile, Tomo I, II and III; FIA: Santiago, Chile, 2017; p. 210.
- Toma de Muestra y Proceso de Datos, 15 octubre 2021; Laboratorio Física de Suelos, Departamento Ingeneria y Suelos, Facultad de Ciencias Agronómicas, Universidad de Chile: Santiago, Chile, 2021.
- Sandoval, M.; Döner, J.; Seguel, O.; Cuevas, J.; Rivera, D. Métodos de Análisis Físicos de Suelos; Universidad de Concepción: Chillan, Chile, 2012; p. 80. [Google Scholar]
- The International Seed Testing Association (ISTA). Reglas Internacionales para el Análisis de las Semillas; ISTA: Wallisellen, Switzerland, 2016; p. 192. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | |||
|---|---|---|---|
| P. chilensis | Q. saponaria | C. alba | |
| Ѱm (kPa) | Germinative Capacity (%) | ||
| 0 | 96.0 (±1.2) a | 57.9 (±7.8) ab | 66.1 (±3.5) a |
| −6 | 86.3 (±3.9) ab | 72.8 (±6.9) a | 61.8 (±7.5) a |
| −33 | 78.2 (±5.0) ab | 40.6 (±6.7) b | 62.2 (±9.6) a |
| −750 | 69.3 (±9.2) ab | 28.5 (±5.8) c | 65.1 (±7.8) a |
| −1000 | 35.9 (±5.3) bc | NA | NA |
| −1250 | 0.0 (±0.00) c | 0.0 (±0.0) d | 0.0 (±0.0) b |
| p-values water potential | <0.0001 | <0.0001 | <0.0001 |
| F-value and significance level | |||
| Seed Source (SS) | 3.43 * | 55.40 *** | 20.75 *** |
| Water potential (Ѱm) | 133.12 *** | 391.40 *** | 152.50 *** |
| SS × Ѱm | 4.02 *** | 12.30 *** | 7.22 *** |
| Geographical Origin | Critical Matric Potential (Ѱm kPa) | Mean for Specie | |
|---|---|---|---|
| Prosopis chilensis | Ovalle (OV) | −1158.1 (±19.8) b | −1191 (±19.9) A |
| Chalinga (LG) | −1262.3 (±12.2) a | ||
| Chacabuco (CH) | −1154.2 (±23.2) b | ||
| Quillaja saponaria | El Melón (ME) | −901.3 (±109.9) cd | −1010 (±32.8) B |
| La Dormida (LD) | −1037.3 (±19.4) bc | ||
| Cantalao (AL) | −1209.0 (±24.9) ab | ||
| Quebrada de la Plata (QL) | −1035.7 (±92.0) bc | ||
| Cantillana (CT) | −964.7 (±32.4) cd | ||
| Pomaire (PO) | −883.3 (±30.2) cd | ||
| Camino el Cobre (CU) | −794.3 (±8.7) d | ||
| Villa Prat (VP) | −1272.0 (±12.7) a | ||
| Ñuble (LB) | −996.0 (±38.4) c | ||
| Cryptocarya alba | Hijuelas (HJ) | −1056.8 (±27.3) ab | −1061 (±34.7) B |
| Antumapu (AN) | −1021.9 (±25.6) ab | ||
| Coya (YA) | −1192.6 (±43.9) a | ||
| Infiernillo (FI) | −1188.3 (±62.6) a | ||
| Loncomilla (LO) | −1005.6 (±107.4) ab | ||
| San Pedro (SP) | −904.1 (±59.6) b |
| Seed Size (g) | Health Status | Precipitation (mm) | Martonne Index | Altitude (m.a.s.l.) | |
|---|---|---|---|---|---|
| P. chilensis | −0.36 (0.3306) | 0.71 (0.0336) | −0.71 (0.0336) | −0.7 (0.0345) | −0.53 (0.1383) |
| Q. saponaria | −0.05 (0.8088) | −0.66 (0.0002) | −0.011 (0.5808) | 0.11 (0.5906) | −0.08 (0.6917) |
| C. alba | 0.8 (0.0001) | −0.21 (0.3954) | −0.42 (0.0845) | −0.41(0.0904) | 0.32 (0.2006) |
| Geographical Origin | Critical Matric Potential (Ѱm kPa) | Mean for Specie | |
|---|---|---|---|
| Prosopis chilensis | Ovalle (OV) | −1158.1 (±19.8) b | −1191 (±19.9) A |
| Chalinga (LG) | −1262.3 (±12.2) a | ||
| Chacabuco (CH) | −1154.2 (±23.2) b | ||
| Quillaja saponaria | El Melón (ME) | −901.3 (±109.9) cd | −1010 (±32.8) B |
| La Dormida (LD) | −1037.3 (±19.4) bc | ||
| Cantalao (AL) | −1209.0 (±24.9) ab | ||
| Quebrada de la Plata (QL) | −1035.7 (±92.0) bc | ||
| Cantillana (CT) | −964.7 (±32.4) cd | ||
| Pomaire (PO) | −883.3 (±30.2) cd | ||
| Camino el Cobre (CU) | −794.3 (±8.7) d | ||
| Villa Prat (VP) | −1272.0 (±12.7) a | ||
| Ñuble (LB) | −996.0 (±38.4) c | ||
| Cryptocarya alba | Hijuelas (HJ) | −1056.8 (±27.3) ab | −1061 (±34.7) B |
| Antumapu (AN) | −1021.9 (±25.6) ab | ||
| Coya (YA) | −1192.6 (±43.9) a | ||
| Infiernillo (FI) | −1188.3 (±62.6) a | ||
| Loncomilla (LO) | −1005.6 (±107.4) ab | ||
| San Pedro (SP) | −904.1 (±59.6) b |
| Geographical Origin | Coordinates UTM | Altitude (masl) | MAT (ºC) | MAP (mm) | MAI | ETR Annual (mm) | Deficit pp (%) | Soil Series | Texture of Soil | Db | Dr | S (%) | Water Retention (kPa) | UW (%) | Health Status | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| X | Y | (Mg m−3) | (Mg m−3) | 33 | 1500 | |||||||||||||
| Prosopis chilensis | Ovalle (OV) | 289,710 | 6,634,010 | 446 | 15.6 | 66 | 2.6 | 1448 | 89 | Tambillo | Sandy loam | 1.45 | 2.71 | 46 | 0.13 | 0.06 | 7 | 2 |
| Chalinga (LG) | 269,291 | 6,596,327 | 157 | 18.8 | 66 | 2.3 | 1448 | 89 | Tuqui | Silty Clay Loam | 1.62 | 2.68 | 40 | 0.22 | 0.14 | 8 | 2 | |
| Chacabuco (CH) | 340,080 | 6,348,960 | 641 | 19.8 | 259 | 8.7 | 1534 | 75 | Rungue | Clayey | 1.46 | 2.65 | 45 | 0.31 | 0.23 | 8 | 1 | |
| Quillaja saponaria | El Melón (ME) | 289,988 | 6,391,566 | 600 | 14.4 | 429 | 17.6 | 1350 | 75 | Catemu | Loam | 1.35 | 2.65 | 49 | 0.2 | 0.1 | 10 | 3 |
| La Dormida (LD) | 314,475 | 6,340,022 | 695 | 14.8 | 310 | 12.5 | 1380 | 73 | Lo Vásquez | Clay Loam | 1.45 | 2.65 | 45 | 0.19 | 0.12 | 7 | 1 | |
| Cantalao (AL) | 359,604 | 6,295,762 | 465 | 14.7 | 174 | 7 | 1474 | 72 | Asociación Challay | Clay Loam | 1.12 | 2.72 | 59 | 0.31 | 0.18 | 13 | 2 | |
| Quebrada de la Plata (QL) | 321,508 | 6,291,796 | 401 | 14.8 | 86 | 3.5 | 1489 | 70 | Asociación Munsel | Sandy Clay Loam | 1.35 | 2.52 | 46 | 0.29 | 0.12 | 17 | 1 | |
| Cantillana (CT) | 321,558 | 6,250,315 | 915 | 14.6 | 115 | 4.7 | 1455 | 72 | Lo Vásquez | Sandy Clay Loam | 1.46 | 2.68 | 46 | 0.21 | 0.09 | 12 | 3 | |
| Pomaire (PO) | 313,058 | 6,280,254 | 216 | 14.4 | 407 | 16.7 | 1443 | 58 | Pahuilmo | Slimy loam | 1.4 | 2.61 | 46 | 0.35 | 0.25 | 10 | 0 | |
| Camino el Cobre (CU) | 353,356 | 621,490 | 220 | 14.1 | 567 | 23.5 | 1471 | 55 | Pimpinela | Silty Clay Loam | 1.49 | 2.68 | 44 | 0.25 | 0.13 | 12 | 2 | |
| Villa Prat (VP) | 236,739 | 6,101,855 | 404 | 14.5 | 671 | 27.4 | 1470 | 38 | Lontue | Sandy loam | 1.25 | 2.65 | 53 | 0.23 | 0.14 | 9 | 3 | |
| Ñuble (LB) | 725,000 | 5,980,500 | 945 | 13.9 | 858 | 35.9 | 1371 | 33 | Asociación Treguaco | Sandy Clay Loam | 1.26 | 2.65 | 52 | 0.26 | 0.14 | 12 | 2 | |
| Cryptocarya alba | Hijuelas (HJ) | 308,503 | 6,260,416.3 | 473 | 14.4 | 429 | 17.6 | 1350 | 71 | Ocoa | Sandy loam | 1.18 | 2.49 | 53 | 0.25 | 0.15 | 10 | 0 |
| Antumapu (AN) | 348,515 | 6,291,391 | 629 | 14.7 | 174 | 7 | 1474 | 68 | Santiago | Clay loam | 1.15 | 2.57 | 55 | 0.34 | 0.19 | 15 | 0 | |
| Coya (CY) | 360,498 | 6,214,498 | 1047 | 14.1 | 567 | 23.5 | 1471 | 55 | Asociación Sierra Bellavista | Sandy loam | 1.45 | 2.67 | 46 | 0.22 | 0.13 | 9 | 1 | |
| Infiernillo (FN) | 221,871 | 6,076,473 | 150 | 14.5 | 671 | 27.4 | 1470 | 40 | Asociación Pocillas | Loamy Clay Loam | 1.41 | 2.62 | 46 | 0.27 | 0.16 | 11 | 2 | |
| Loncomilla (LN) | 229,719,9 | 6,065,850 | 88 | 13.8 | 720 | 30.3 | 1521 | 33 | Asociación Cauquenes | Clay loam | 1.45 | 2.69 | 46 | 0.25 | 0.16 | 9 | 1 | |
| San Pedro (SP) | 191,758 | 6,066,938 | 230 | 13.3 | 920 | 39.5 | 1271 | 35 | Asociación Constitución | Sandy Clay Loam | 1.37 | 2.58 | 47 | 0.31 | 0.21 | 10 | 3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faúndez, Á.; Magni, C.R.; Martínez-Herrera, E.; Espinoza, S.; Vaswani, S.; Yañez, M.A.; Gréz, I.; Seguel, O.; Abarca-Rojas, B.; Quiroz, I. Effect of the Soil Matric Potential on the Germination Capacity of Prosopis chilensis, Quillaja saponaria and Cryptocarya alba from Contrasting Geographical Origins. Plants 2022, 11, 2963. https://doi.org/10.3390/plants11212963
Faúndez Á, Magni CR, Martínez-Herrera E, Espinoza S, Vaswani S, Yañez MA, Gréz I, Seguel O, Abarca-Rojas B, Quiroz I. Effect of the Soil Matric Potential on the Germination Capacity of Prosopis chilensis, Quillaja saponaria and Cryptocarya alba from Contrasting Geographical Origins. Plants. 2022; 11(21):2963. https://doi.org/10.3390/plants11212963
Chicago/Turabian StyleFaúndez, Ángela, Carlos R. Magni, Eduardo Martínez-Herrera, Sergio Espinoza, Suraj Vaswani, Marco A. Yañez, Iván Gréz, Oscar Seguel, Betsabé Abarca-Rojas, and Iván Quiroz. 2022. "Effect of the Soil Matric Potential on the Germination Capacity of Prosopis chilensis, Quillaja saponaria and Cryptocarya alba from Contrasting Geographical Origins" Plants 11, no. 21: 2963. https://doi.org/10.3390/plants11212963