2.1. Production and Germination of Artificial Seeds
According to the analysis of variance, the percentage of germination and the number of days until germination started were not affected by the triple interaction of the treatments.
On the other hand, the interaction of the germination medium at the time of ion exchange did not have a significant effect on the percentage of germination, but the main effects of the treatments, the interaction of the germination medium at the concentration of Sodium alginate and the interaction of the concentration of sodium alginate at the time of ion exchange had a significant effect on these growth parameters (
Table 1).
Figure 1.
(A) S. pennata in the desert habitat. (B) Seeds of wild growing plants. (C) Temporary immersion bioreactor system with germinated artificial seeds. (D) Temporary immersion bioreactor (TIB) system. (E) Shoot growth in solid media. (F) Artificial seeds of S. pennata. (G) Germinated artificial seed of S. pennata. Scale bar: (A) is 10 cm; (B–G) is 1 cm.
Figure 1.
(A) S. pennata in the desert habitat. (B) Seeds of wild growing plants. (C) Temporary immersion bioreactor system with germinated artificial seeds. (D) Temporary immersion bioreactor (TIB) system. (E) Shoot growth in solid media. (F) Artificial seeds of S. pennata. (G) Germinated artificial seed of S. pennata. Scale bar: (A) is 10 cm; (B–G) is 1 cm.
In addition, the main effects of the interaction of the germination medium at the time of ion exchange, the interaction of the germination medium at the concentration of sodium alginate, and the interaction of the concentration of sodium alginate at the time of ion exchange (
Table 1) significantly influenced the number of days to germination.
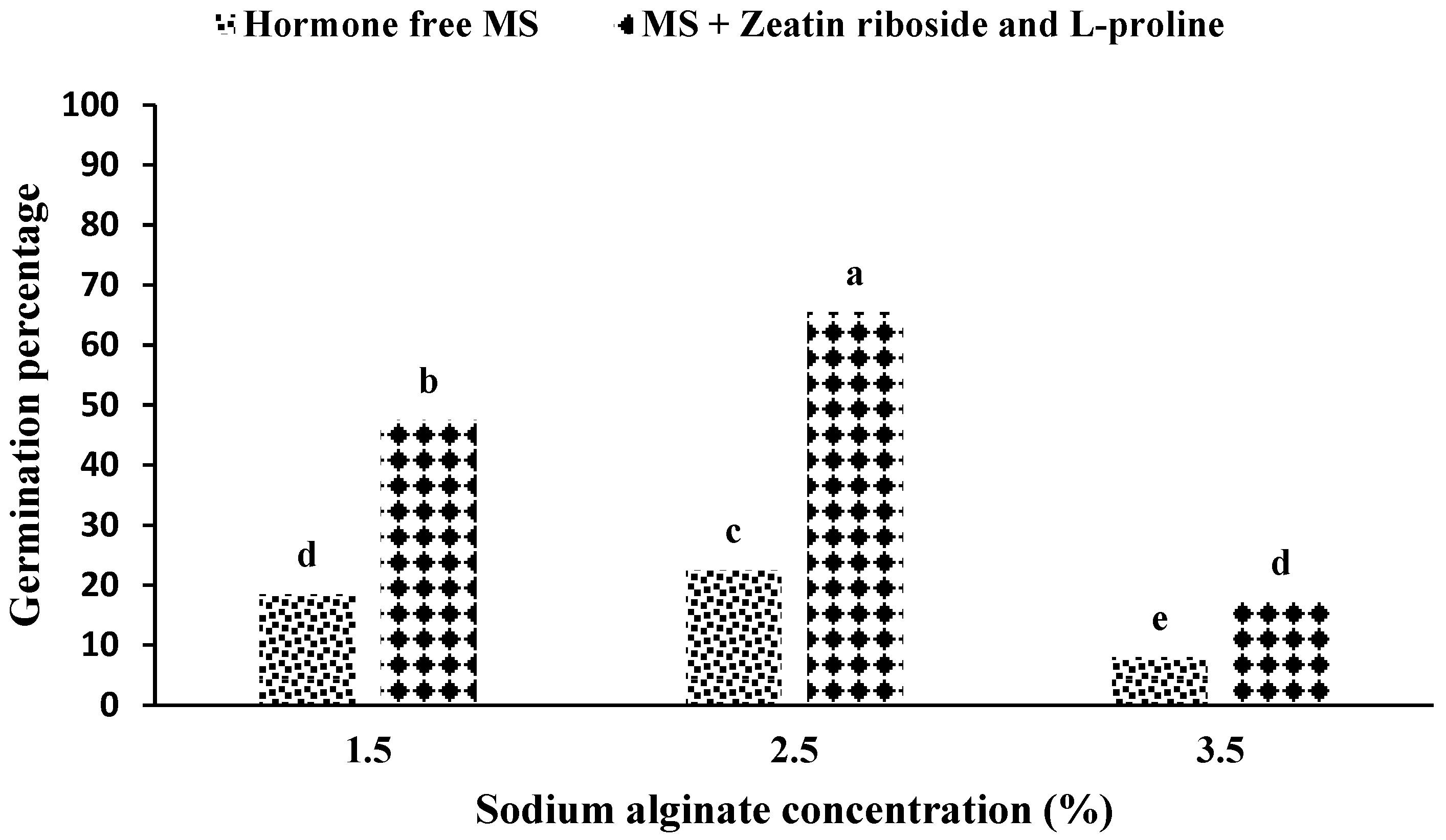
A comparison of the means showed the highest percentage of germination was related to the use of 2.5% sodium alginate in an MS medium containing zeatin riboside and L-proline. In addition, the lowest germination percentage was observed in the MS medium without growth hormone and 3.5% sodium alginate application (
Figure 2).
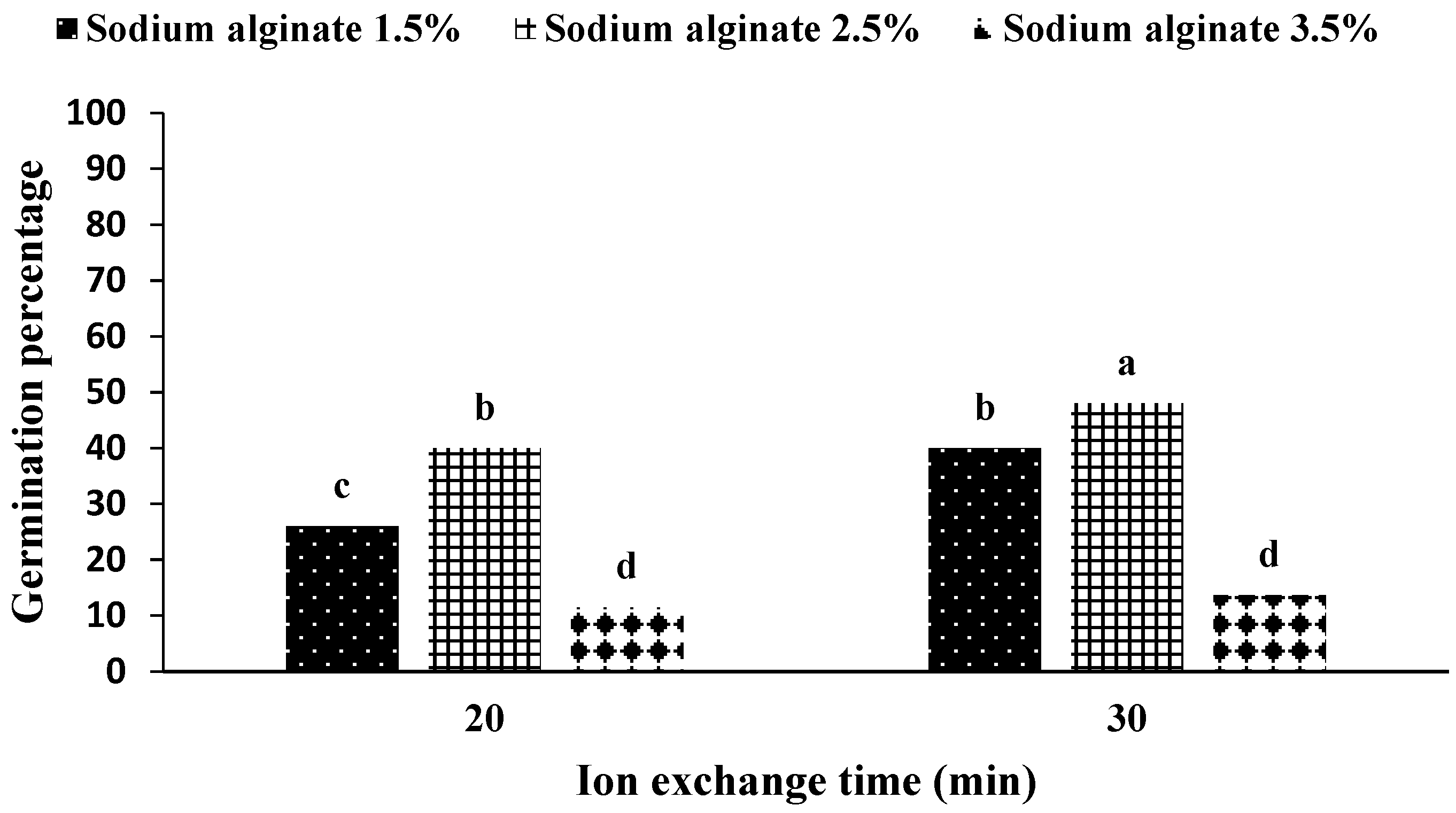
The application of 2.5% sodium alginate and 30 min of ion exchange time increased the germination percentage of artificial seeds, and the highest amount of germination was in the interaction of sodium alginate application at ion exchange time. On the other hand, the use of 3.5% sodium alginate in both ion exchange times caused the lowest germination percentage (
Figure 3).
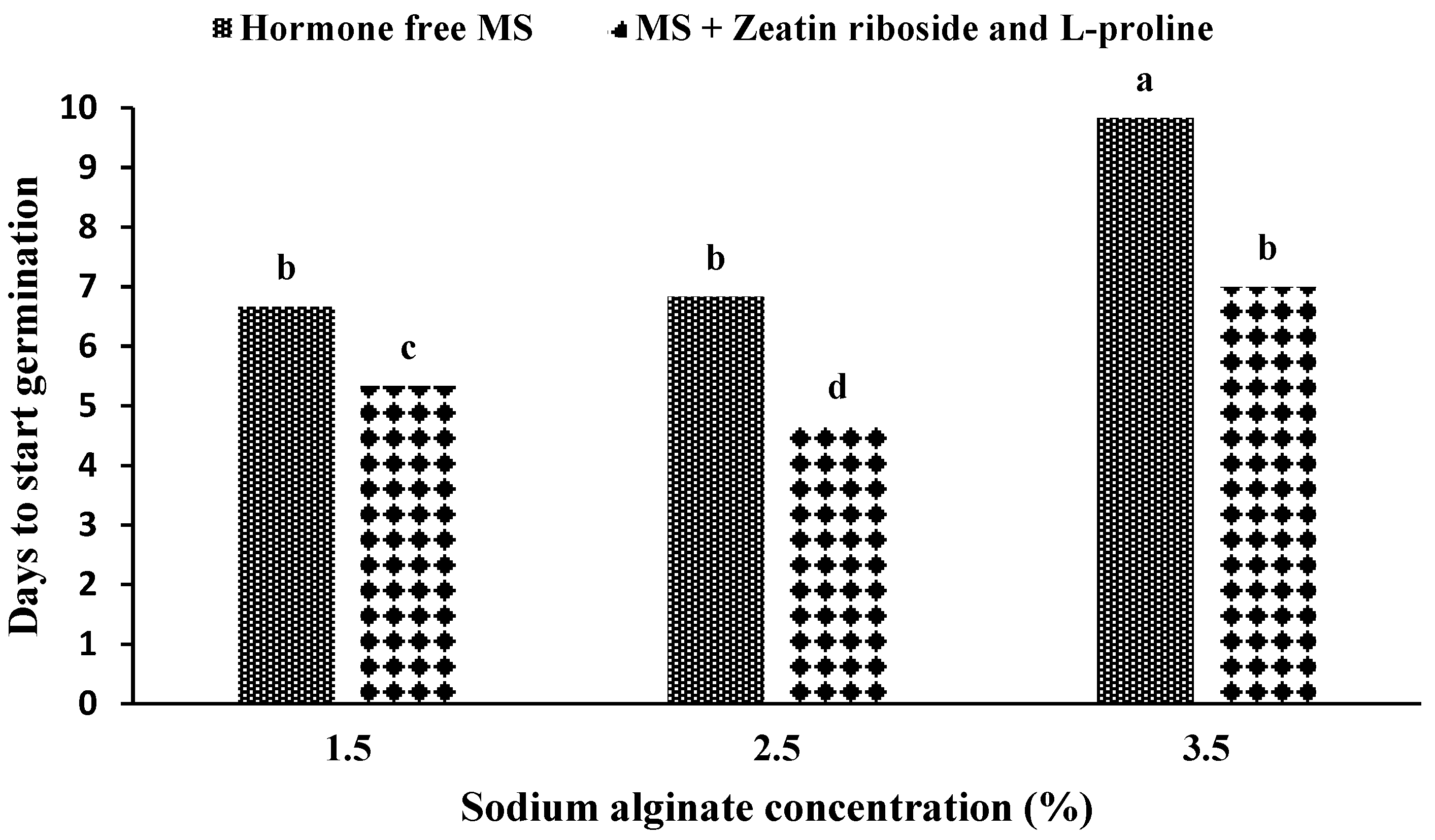
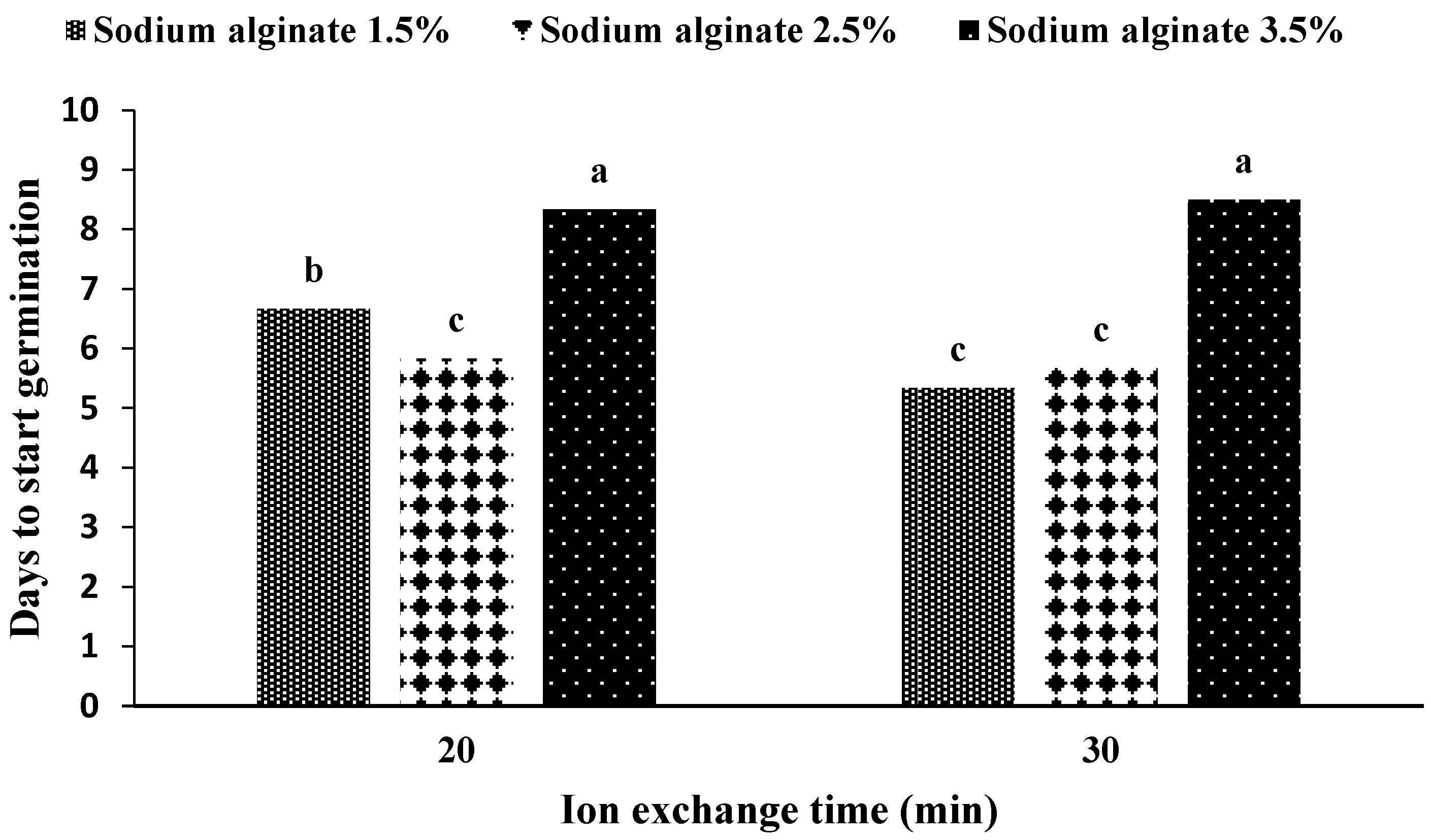
The maximum time until the germination of artificial seeds was obtained in an MS medium without growth hormones with the use of 3.5% sodium alginate (
Figure 4), and the use of 2.5% sodium alginate in the medium containing zeatin riboside and L-proline showed the shortest time for germination. The results were statistically similar in the hormone-free medium and the application of 1.5% and 2.5% sodium alginate, and the medium containing zeatin riboside and L-proline and the application of 3.5% sodium alginate were statistically similar.
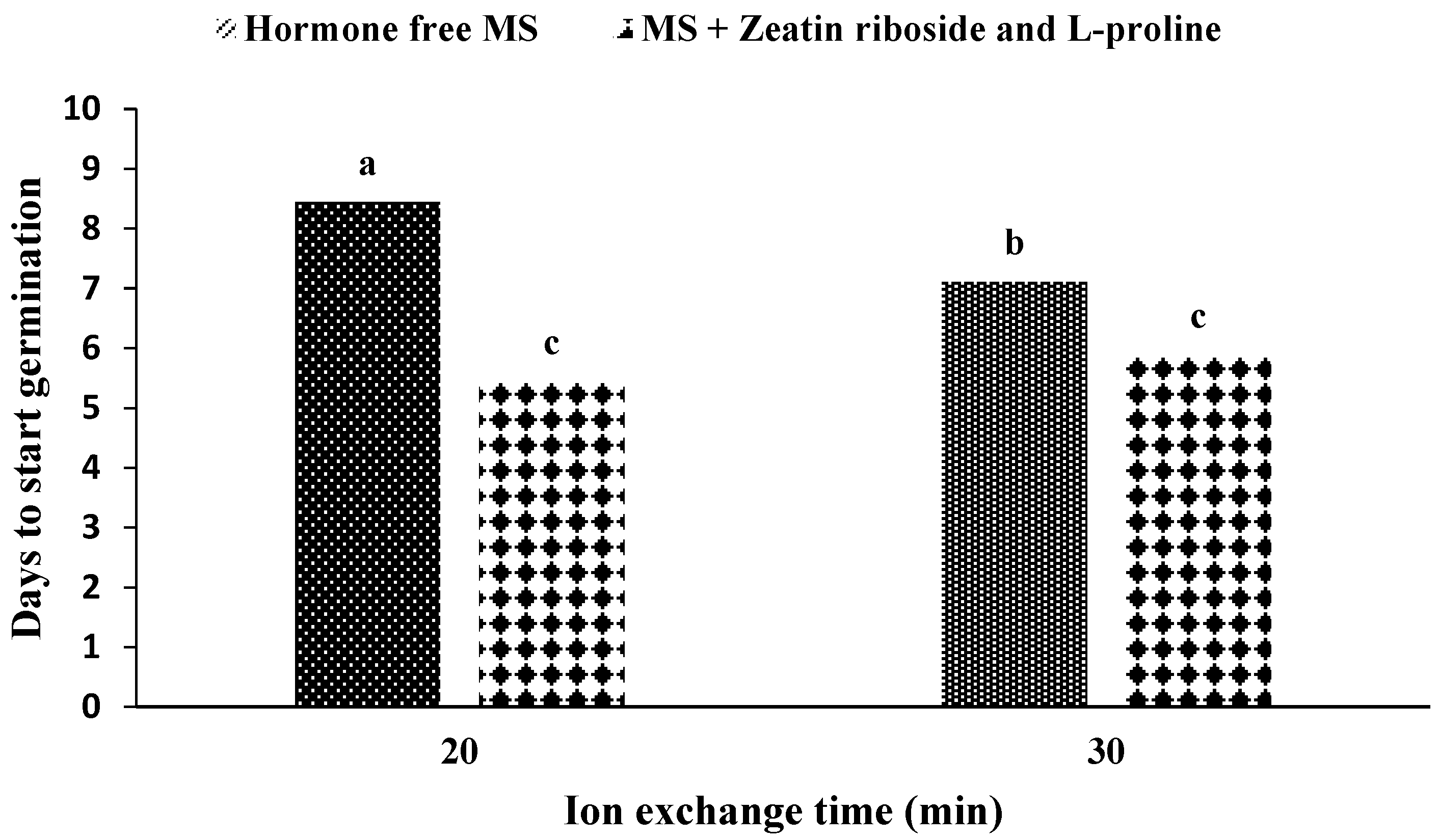
In the mean comparison of the medium during ion exchange, it was observed that if the medium is free of hormones and the ion exchange time is determined to be 20 min, the maximum time would be required until germination begins (
Figure 5). In addition, if the medium contains zeatin riboside and L-proline (no difference in ion exchange time at 20 min and 30 min), the embryo takes the least time to break out of the capsule.
The interaction between sodium alginate concentration and ion exchange time on the number of days until the beginning of germination showed that for the concentrations of 2.5% sodium alginate (with both ion exchange times) and 1.5% sodium alginate (with 30 min of ion exchange), the artificial seed needs the least number of days for germination (
Figure 6). On the other hand, the longest time for the embryo to germinate corresponds to the concentration of 3.5% sodium alginate (both times of ion exchange).
The use of sodium alginate is important in the formation of artificial seeds. An increase in sodium alginate concentration causes hardness of the explant protective gel and will hurt the percentage of germination and the time to sprout seedlings from the capsule. This is despite the fact that sodium alginate in very low concentrations does not help to maintain the proper shape and storage of artificial seeds because the formed capsule is very soft and flexible and cannot support the explant well against movement and physical pressure Redenbaugh et al. (1987) [
23].
In an experiment by Haque and Ghosh (2016) [
24], three concentrations of sodium alginate, 1.5%, 3%, and 4.5%, were used in the preparation of
Ledebouria revoluta artificial seeds. Even though the highest percentage of germination was obtained in the concentration of 1.5% sodium alginate, it was found that the created capsules were not well polymerized and were not suitable for carrying. A concentration of 3% was considered the optimal condition. On the other hand, the concentration of 4.5% sodium alginate greatly reduced the germination percentage, and the embryo was not able to exit the capsule due to its high hardness. Similar results have been obtained by [
25], where sodium alginate’s optimal concentration was 3%.
The research of Inpuay and Te-chato (2012) [
26] on oil palm explants showed that the interaction of 2.5% sodium alginate concentration and 15 min of ion exchange time had the best germination percentage of artificial seeds. In addition, the excessive softness of the capsule at a two percent concentration and the high hardness of the gel matrix at a three percent concentration of sodium alginate were stated as reasons for the decrease in germination.
Investigating the interaction of the medium in sodium alginate showed that the highest percentage of germination was in the treatment of a hormone-free medium and a 2.5% concentration of sodium alginate (
Figure 2). In addition, the results of the interaction between ion exchange time and sodium alginate concentration showed that the concentration of 2.5% sodium alginate in the ion exchange period of 30 min (
Figure 3) considering the treatment combination.
Considering these results and comparing them with the results of other researchers, it seems that the concentration of sodium alginate played a central role in the formation of artificial seeds of
S. pennata. According to observations, if the appropriate concentration of sodium alginate (both lack and excess concentration) is not present in the gel matrix, with a concentration of 1.5% and the presence of growth hormones, 27% compared to a concentration of 2.5% and the presence of plant growth hormones reduced germination. This amount of reduction was 74% with a concentration of 3.5% and the presence of the plant growth hormone (
Figure 2). Similarly, the time of 30 min for ion exchange and the concentration of 2.5% sodium alginate showed a 17% increase in the percentage of germination, which was lower than the statistical level (
Figure 3).
According to Ghosh and Sen (1994) [
27], the best treatment for artificial seed production was 3.5% sodium alginate and an ion exchange time of 40 min. It seems that the shorter time in the present experiment (30 min) may be due to the synergistic effects of sodium alginate and calcium as well as their concentration. Both sodium alginate concentration and calcium chloride are the key components in the polymerization process, according to Mujib et al. (2014) [
28].
Extensive results have been obtained regarding the role of nutrients in the culture environment, including various types of sugars, growth hormones, etc. According to Rizkalla et al. (2012) [
29], adding 30 g/L of sucrose to the culture medium for sugar beet explants showed the highest germination percentage, and additionally, the shortest time was needed for starting the germination. Other research also stated that the presence of nutrients in the alginate matrix led to the better regeneration of banana explants (Panis, 1996) [
9].
The presence of nutrients in the gel matrix, which are placed around the explant as a nutritional substrate, probably improved the growth and survival of the S. pennata explants. We can assume that L-proline is needed to provide the energy for germination. On the other hand, the presence of zeatin riboside stimulated the explant growth and the proper development of mature seedlings.
It was shown by Huda et al. (2007) [
30] that sucrose was the most suitable source of carbon supply for eggplant artificial seed production. Similar results were obtained in the production of
Tylophora indica artificial seeds by Gantait et al. (2017) [
31]. The highest percentage of germination and the highest degree of polymerization of the capsule around the explant and, at the same time, the shortest time until the germination occurred when 3% sodium alginate was used.
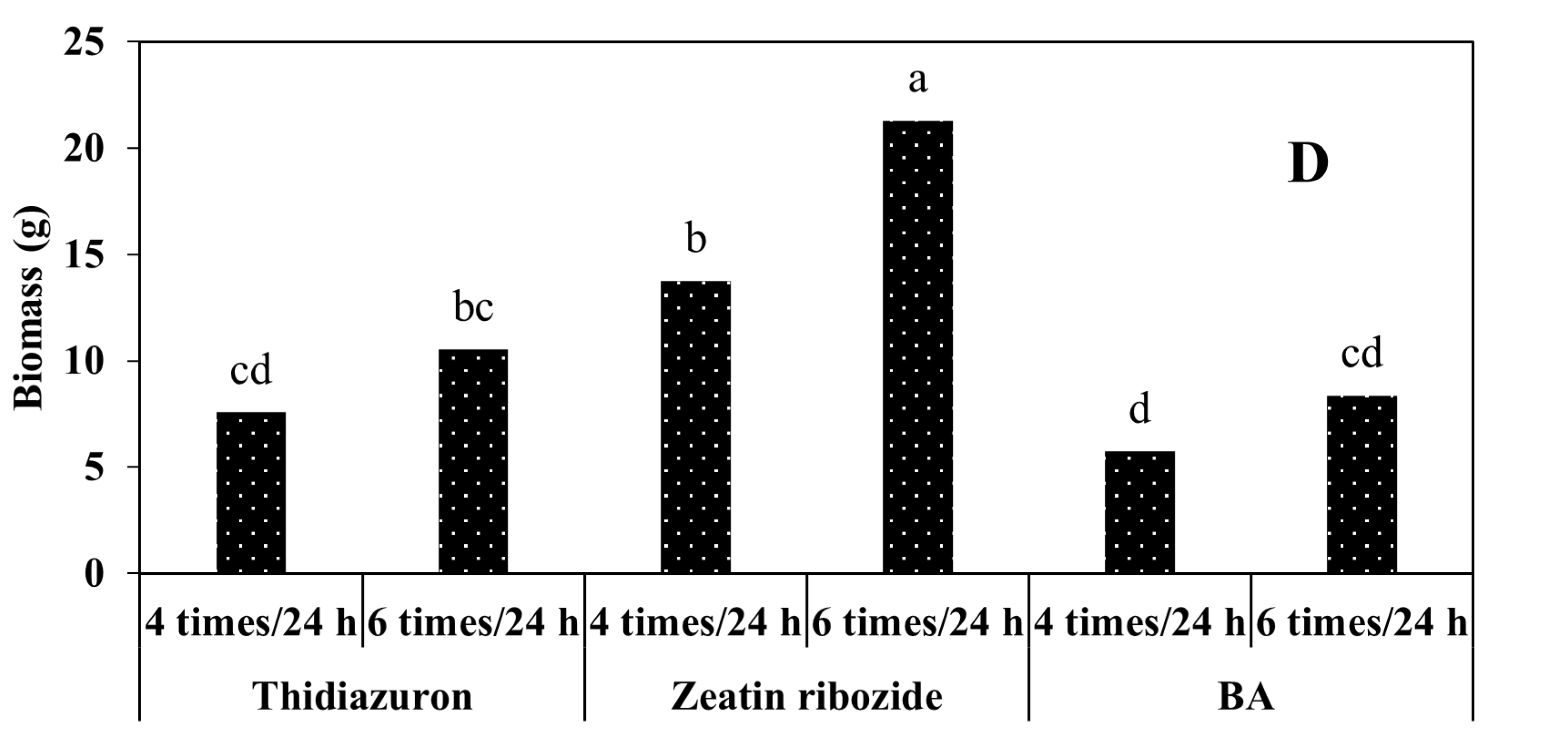
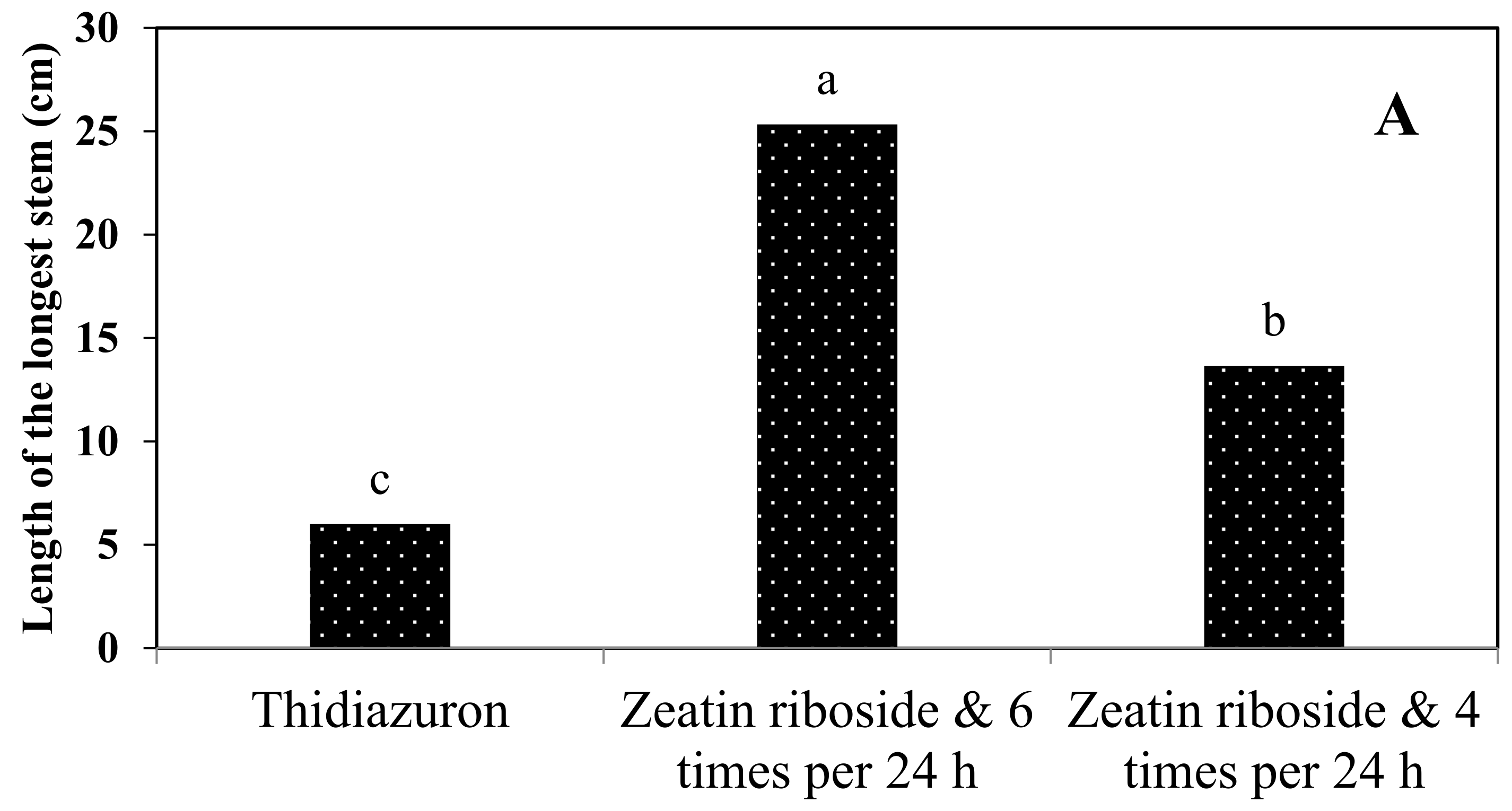
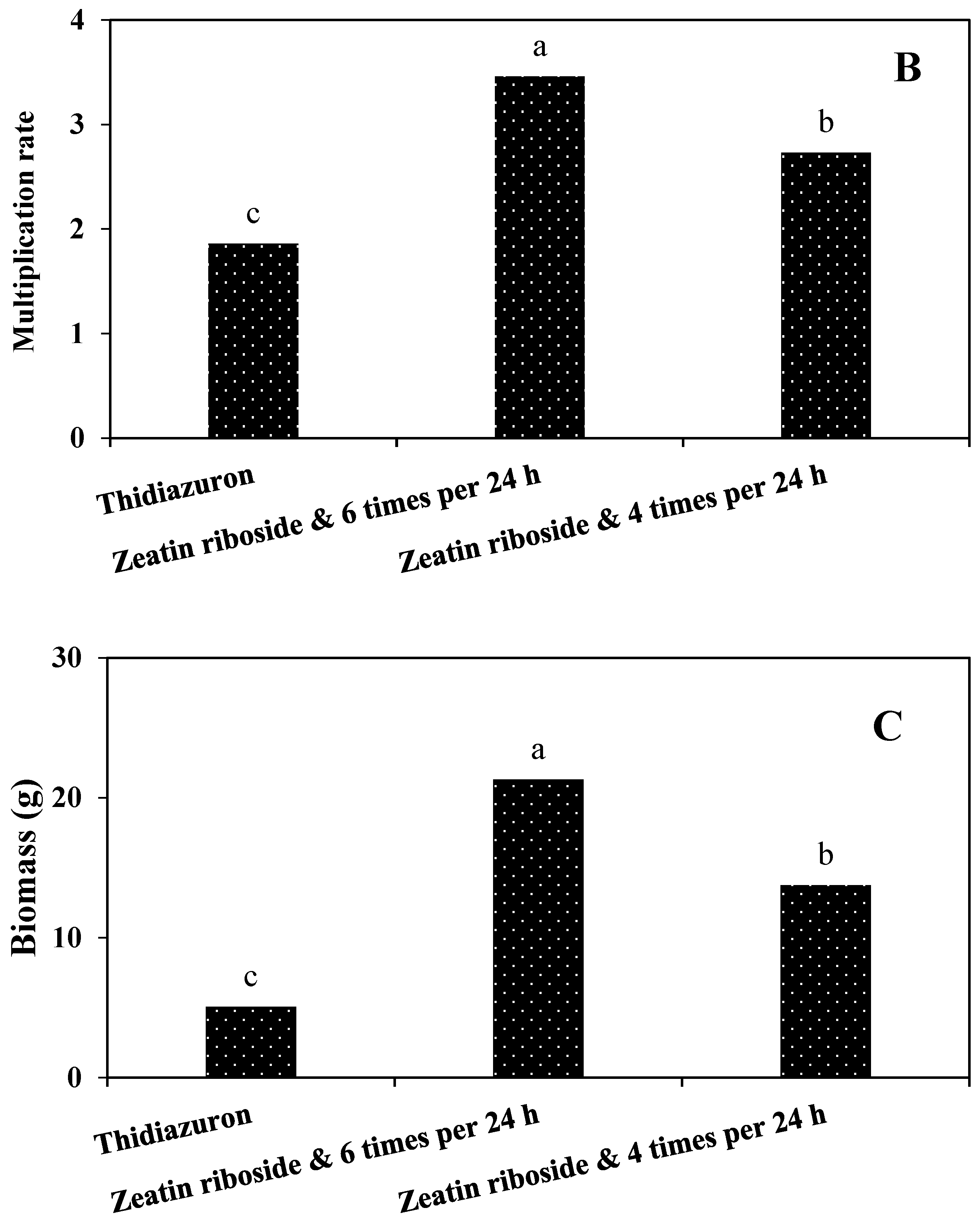
In general, according to our results, the most suitable conditions for the production of artificial seeds were when 2.5% sodium alginate with a duration of ion exchange of 30 min was used and the artificial seeds were placed on the culture medium supplemented with zeatin riboside and L-proline.

 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}