
Seasonal Variation of Tropical Savanna Altered Agronomic Adaptation of Stock-6-Derived Inducer Lines

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Analysis of Variance
2.2. Seasonal Variation and Plant Phenology
2.3. Performance of Maize Haploid Inducers against Seasonal Variation
2.4. Interaction between Genotype and Season in Maize Haploid Inducers
3. Materials and Methods
3.1. Plant Materials
3.2. Field Experiment
3.3. Haploid Induction and Ploidy Identification
3.4. Data Collection
3.5. Statistical Analysis
4. Discussion
4.1. Phenotype of Maize Haploid inducers Is Affected by Genotype, Season, and their Interaction
4.2. Seasonal Variation Is Responsible for Unstable Performance of Maize Haploid Inducers
4.3. Crossover Performance of Maize Haploid Inducers and Implication for Haploid Induction
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaikam, V.; Molenaar, W.; Melchinger, A.E.; Prasanna, M.B. Doubled haploid technology for line development in maize: Technical advances and prospects. Theor. Appl. Genet. 2019, 132, 3227–3243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Fuente, G.N.; Frei, U.K.; Trampe, B.; Nettleton, D.; Zhang, W.; Lübberstedt, T. A diallel analysis of a maize donor population response to in vivo maternal haploid induction: I. Inducibility. Crop. Sci. 2018, 58, 1830–1837. [Google Scholar] [CrossRef] [Green Version]
- Coe, E.H. A line of maize with high haploid frequency. Am. Nat. 1959, 93, 381–382. [Google Scholar] [CrossRef]
- Tyrnov, V.S.; Zavalishina, A.N. High-frequency induction of matroclinal haploids in maize. Dokl. Biol. Sci.–Akad. Nauk. SSSR 1984, 276, 353–355. [Google Scholar]
- Lashermes, P.; Beckert, M. Genetic control of maternal haploidy in maize (Zea mays L.) and selection of haploid inducing lines. Theor. Appl. Genet. 1988, 76, 405–410. [Google Scholar] [CrossRef]
- Chalyk, S.T. Creating new haploid-inducing lines of maize. Maize Genet. Coop. News Lett. 1999, 73, 53–54. [Google Scholar]
- Röber, F.K.; Gordillo, G.A.; Geiger, H.H. In vivo haploid induction in maize—Performance of new inducers and significance of doubled haploid lines in hybrid breeding. Maydica 2005, 50, 275–283. [Google Scholar]
- Chang, M.T.; Coe, E.H. Doubled haploids. In Molecular Genetic Approaches to Maize Improvement. Biotechnology in Agriculture and Forestry; Kriz, A.L., Larkins, B.A., Eds.; Springer: Berlin, Germany, 2009; Volume 63, pp. 127–142. [Google Scholar]
- Rotarenco, V.A.; Dicu, G.; State, D.; Fuia, S. New inducers of maternal haploids in maize. Maize Genet. Coop. News Lett. 2010, 84, 21–22. [Google Scholar]
- De La Fuente, G.N. Improvement to the Maize (Zea mays L.) In Vivo Maternal Doubled Haploid System. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2015; pp. 38–51. [Google Scholar]
- Prigge, V.; Schipprack, W.; Mahuku, G.; Atlin, G.N.; Melchinger, A.E. Development of in vivo haploid inducers for tropical maize breeding programs. Euphytica 2012, 185, 481–490. [Google Scholar] [CrossRef]
- Prasanna, B.M. Doubled haploid (DH) technology in maize breeding: An overview. In Doubled Haploid Technology in Maize Breeding: Theory and Practice; Prasanna, B.M., Chaikam, V., Mahuku, G., Eds.; CIMMYT: Texcoco, Mexico, 2012; pp. 1–8. [Google Scholar]
- Chaikam, V.; Nair, S.K.; Martinez, L.; Lopez, L.A.; Utz, H.F.; Melchinger, A.E.; Prasanna, B.M. Marker-assisted breeding of improved maternal haploid inducers in maize for the tropical/subtropical regions. Front. Plant Sci. 2018, 9, 1527. [Google Scholar] [CrossRef] [Green Version]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- Kebede, A.Z.; Dhillon, B.S.; Schipprack, W.; Araus, J.L.; Bänziger, M.; Semagn, K.; Alvarado, G.; Melchinger, A.E. Effect of source germplasm and season on the in vivo haploid induction rate in tropical maize. Euphytica 2011, 180, 219–226. [Google Scholar] [CrossRef]
- Dermail, A.; Chankaew, S.; Lertrat, K.; Lübberstedt, T.; Suriharn, K. Selection gain of maize haploid inducers for the tropical savanna environments. Plants 2021, 10, 2812. [Google Scholar] [CrossRef]
- Abadassi, J. Maize agronomic traits needed in tropical zone. Int. J. Environ. Sci. Technol. 2015, 4, 371–392. [Google Scholar]
- Dermail, A.; Fuengtee, A.; Lertrat, K.; Suwarno, W.B.; Lübberstedt, T.; Suriharn, K. Simultaneous selection of sweet-waxy corn ideotypes appealing to hybrid seed producers, growers, and consumers in Thailand. Agronomy 2022, 12, 87. [Google Scholar] [CrossRef]
- Uliana Trentin, H.; Frei, U.K.; Lübberstedt, T. Breeding maize maternal haploid inducers. Plants 2020, 9, 614. [Google Scholar] [CrossRef]
- Elston, R.C. A weight-free index for the purpose of ranking or selection with respect to several traits at a time. Biometrics 1963, 19, 85–97. [Google Scholar] [CrossRef]
- Stewart, D.W.; Dwyer, L.M.; Carrigan, L.L. Phenological temperature response of maize. Agron. J. 1998, 90, 73–79. [Google Scholar] [CrossRef]
- Nanda, D.K.; Chase, S.S. An embryo marker for detecting monoploids of maize (Zea mays L.). Crop. Sci. 1966, 6, 213–215. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedure for Agricultural Research; John Wiley and Sons: Singapore, 1984; pp. 594–600. [Google Scholar]
- Acquaah, G. Principles of Plant Genetics and Breeding; Blackwell: Oxford, UK, 2006; pp. 121–134. [Google Scholar]
- Chaikam, V.; Nair, S.K.; Babu, R.; Martinez, L.; Tejomurtula, J.; Prasanna, M.B. Analysis of effectiveness of R1-nj anthocyanin marker for in vivo haploid identification in maize and molecular markers for predicting the inhibition of R1-nj expression. Theor. Appl. Genet. 2015, 128, 159–171. [Google Scholar] [CrossRef]
- Dermail, A.; Suriharn, B.; Chankaew, S.; Sanitchon, J.; Lertrat, K. Hybrid prediction based on SSR-genetic distance, heterosis and combining ability on agronomic traits and yields in sweet and waxy corn. Sci. Hortic. 2020, 259, 108817. [Google Scholar] [CrossRef]
- Khamphasan, P.; Lomthaisong, K.; Harakotr, B.; Scott, M.P.; Lertrat, K.; Suriharn, B. Effects of mass selection on husk and cob color in five purple corn populations segregating for purple husks. Agriculture 2020, 10, 311. [Google Scholar] [CrossRef]
- Dudley, J.W.; Moll, R.H. Interpretation and use of estimates of heritability and genetic variances in plant breeding. Crop. Sci. 1969, 9, 257–262. [Google Scholar] [CrossRef]
- Lobell, D.B.; Bänziger, M.; Magorokosho, C.; Vivek, B. Nonlinear heat effects on African maize as evidenced by historical yield trials. Nat. Clim. Change 2011, 1, 42–45. [Google Scholar] [CrossRef]
- Yin, Y.; Deng, H.; Wu, S. Spatial-temporal variations in the thermal growing degree-days and season under climate warming in China during 1960–2011. Int. J. Biometeorol. 2019, 63, 649–658. [Google Scholar] [CrossRef]
- Liu, Y.E.; Hou, P.; Xie, R.Z.; Li, S.K.; Zhang, H.B.; Ming, B.; Ma, D.L.; Liang, S.M. Spatial adaptabilities of spring maize to variation of climatic conditions. Crop. Sci. 2013, 53, 1693–1703. [Google Scholar] [CrossRef]
- Dahmardeh, M. Effects of sowing date on the growth and yield of maize cultivars (Zea mays L.) and the growth temperature requirements. Afr. J. Biotechnol. 2012, 11, 12450–12453. [Google Scholar] [CrossRef]
- Hou, P.; Liu, Y.; Xie, R.; Ming, B.; Ma, D.; Li, S.; Mei, X. Temporal and spatial variation in accumulated temperature requirements of maize. Field Crop. Res. 2014, 158, 55–64. [Google Scholar] [CrossRef]
- Yang, H.S.; Dobermann, A.; Lindquist, J.L.; Walters, D.T.; Arkebauer, T.J.; Cassman, K.G. Hybrid-Maize—A maize simulation model that combines two crop modeling approaches. Field Crop. Res. 2004, 87, 131–154. [Google Scholar] [CrossRef] [Green Version]
- Bonhomme, R.; Derieux, M.; Edmeades, G.O. Flowering of diverse maize cultivars in relation to temperature and photoperiod in multilocation field trials. Crop. Sci. 1994, 34, 156–164. [Google Scholar] [CrossRef]
- Jiang, W.; Thapa, S.; Jessup, K.; Hao, B.; Hou, X.; Marek, T.; Becker, J.; Bell, J.; Xue, Q. Corn response to later than traditional planting dates in the Texas high plains. Crop. Sci. 2020, 60, 1004–1020. [Google Scholar] [CrossRef]
- Žalud, Z.; Dubrovsky, M. Modelling climate change impacts on maize growth and development in the Czech Republic. Theor. Appl. Climatol. 2002, 72, 85–102. [Google Scholar] [CrossRef]
- Lizaso, J.I.; Ruiz-Ramos, M.; Rodríguez, L.; Gabaldon-Leal, C.; Oliveira, J.A.; Lorite, I.J.; Sánchez, D.; García, E.; Rodríguez, A. Impact of high temperatures in maize: Phenology and yield components. Field Crop. Res. 2018, 216, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Derieux, M.; Bonhomme, R. Heat unit requirement for maize hybrids in Europe. Results of the European FAO subnetwork: II. Period from silking to maturity. Maydica 1982, 27, 79–96. [Google Scholar]
- Zhou, B.; Yue, Y.; Sun, X.; Wang, X.; Wang, Z.; Ma, W.; Zhao, M. Maize grain yield and dry matter production responses to variations in weather conditions. Agron. J. 2016, 108, 196–204. [Google Scholar] [CrossRef]
- Skaugen, T.E.; Tveito, O.E. Growing-season and degree-day scenario in Norway for 2021–2050. Clim. Res. 2004, 26, 221–232. [Google Scholar] [CrossRef]
- Iannucci, A.; Terribile, M.R.; Martiniello, P. Effects of temperature and photoperiod on flowering time of forage legumes in a Mediterranean environment. Field Crop. Res. 2008, 106, 156–162. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, X.; Wang, E.; Chen, S.; Shao, L.; Qin, W. Assessing the contribution of weather and management to the annual yield variation of summer maize using APSIM in the north China plain. Field Crop. Res. 2016, 194, 94–102. [Google Scholar] [CrossRef]
- Zhou, B.; Yue, Y.X.; Sun, Z.D.; Ma, W.; Zhao, M. Maize kernel weight responses to sowing date-associated variation in weather conditions. Crop. J. 2017, 5, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Tsimba, R.; Edmeades, G.O.; Millner, J.P.; Kemp, P.D. The effect of planting date on maize: Phenology, thermal time durations and growth rates in a cool temperate climate. Field Crop. Res. 2013, 150, 145–155. [Google Scholar] [CrossRef]
- Chase, S.S.; Nanda, D.K. Number of leaves and maturity classification in Zea mays L. Crop. Sci. 1967, 7, 431–432. [Google Scholar] [CrossRef]
- Birch, C.J.; Hammer, G.L.; Rickert, K.G. Improved methods for predicting individual leaf area and leaf senescence in maize (Zea mays). Aust. J. Agric. Res. 1998, 49, 249–262. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, X.; Chen, S.; Pei, D.; Liu, C. Effects of harvest and sowing time on the performance of the rotation of winter wheat–summer maize in the North China Plain. Ind. Crop. Prod. 2007, 25, 239–247. [Google Scholar] [CrossRef]
- Chassaigne-Ricciulli, A.A.; Mendoza-Onofre, L.E.; Córdova-Téllez, L.; Carballo-Carballo, A.; San Vicente-García, F.M.; Dhliwayo, T. Effective seed yield and flowering synchrony of parents of CIMMYT three-way-cross tropical maize hybrids. Agriculture 2021, 11, 161. [Google Scholar] [CrossRef]
- Hunter, R.B.; Tollenaar, M.; Breuer, C.M. Effects of photoperiod and temperature on vegetative and reproductive growth of a maize (Zea mays) hybrid. Can. J. Plant Sci. 1977, 57, 1127–1133. [Google Scholar] [CrossRef]
- Baum, M.E.; Archontoulis, S.V.; Licht, M.A. Planting date, hybrid maturity, and weather effects on maize yield and crop stage. Agron. J. 2019, 111, 303–313. [Google Scholar] [CrossRef]
- Holzkämper, A.; Fuhrer, P.C.J. Identifying climatic limitations to grain maize yield potentials using a suitability evaluation approach. Agric. For. Meteorol. 2013, 168, 149–159. [Google Scholar] [CrossRef]
- Tanaka, W.; Angel Maddonni, G. Maize kernel oil and episodes of shading during the grain-filling period. Crop. Sci. 2009, 49, 2187–2197. [Google Scholar] [CrossRef]
- Tao, F.; Yokozawa, M.; Xu, Y.; Hayashi, Y.; Zhang, Z. Climate changes and trends in phenology and yields of field crops in China, 1981–2000. Agric. For. Meteorol. 2006, 138, 82–92. [Google Scholar] [CrossRef]
- Liu, S.; Mo, X.; Lin, Z.; Xu, Y.; Ji, J.; Wen, G.; Richey, J. Crop yield responses to climate change in the Huang-Huai Hai Plain of China. Agric. Water Manag. 2010, 97, 1195–1209. [Google Scholar] [CrossRef]
- Wei, T.; Zhang, T.; De Bruin, K.; Glomrød, S.; Shi, Q. Extreme weather impacts on maize yield: The case of Shanxi province in China. Sustainability 2017, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Peters, D.; Pendleton, J.; Hageman, R.; Brown, C. Effect of night air temperature on grain yield of corn, wheat, and soybeans. Agron. J. 1971, 63, 809. [Google Scholar] [CrossRef]
- Wang, Y.; Tao, H.; Tian, B.; Sheng, D.; Xu, C.; Zhou, H.; Huang, S.; Wang, P. Flowering dynamics, pollen, and pistil contribution to grain yield in response to high temperature during maize flowering. Environ. Exp. Bot. 2019, 158, 80–88. [Google Scholar] [CrossRef]
- Bassetti, P.; Westgate, M.E. Water deficit affects receptivity of maize silks. Crop. Sci. 1993, 33, 279–282. [Google Scholar] [CrossRef]
- Rattalino Edreira, J.I.; Carpici, E.B.; Sammarro, D.; Otegui, M.E. Heat stress effects around flowering on kernel set of temperate and tropical maize hybrids. Field Crop. Res. 2011, 123, 62–73. [Google Scholar] [CrossRef]
- Jones, R.J.; Gengenbach, B.G.; Cardwell, V.B. Temperature effects on in vitro kernel development of maize. Crop. Sci. 1981, 21, 761–766. [Google Scholar] [CrossRef]
- Yan, P.; Chen, Y.Q.; Sui, P.; Vogel, A.; Zhang, X.P. Effect of maize plant morphology on the formation of apical kernels at different sowing dates and under different plant densities. Field Crop. Res. 2018, 223, 83–92. [Google Scholar] [CrossRef]
- Cerrudo, A.; Di Matteo, J.; Fernandez, E.; Robles, M.; Pico, L.O.; Andrade, F.H. Yield components of maize as affected by short shading periods and thinning. Crop. Pasture Sci. 2013, 64, 580–587. [Google Scholar] [CrossRef]
- Seebauer, J.R.; Moose, S.P.; Fabbri, B.J.; Crossland, L.D.; Below, F.E. Amino acid metabolism in maize earshoots. Implications for assimilate preconditioning and nitrogen signaling. Plant Physiol. 2004, 136, 4326–4334. [Google Scholar] [CrossRef] [Green Version]
- Scharf, P.C.; Kitchen, N.R.; Sudduth, K.A.; Davis, J.G. Spatially variable corn yield is a weak predictor of optimal nitrogen rate. Soil Sci. Soc. Am. J. 2006, 70, 2154–2160. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Tremblay, N.; Tremblay, G.; Bourgeois, G.; Bouroubi, M.; Wei, Z. Weather effects on corn response to in-season nitrogen rates. Can. J. Plant Sci. 2013, 93, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Marković, M.; Šoštarić, J.; Josipović, M.; Atilgan, A. Extreme weather events affect agronomic practices and their environmental impact in maize cultivation. Appl. Sci. 2021, 11, 7352. [Google Scholar] [CrossRef]
- Westgate, M.; Lisazo, J.; Batchelor, W. Quantitative relationship between pollen-shed density and grain yield in maize. Crop. Sci. 2003, 43, 934–942. [Google Scholar] [CrossRef]
- Carcova, J.; Uribelarrea, M.; Borras, L.; Otegui, M.E.; Westgate, M.E. Synchronous pollination within and between ears improves kernel set in maize. Crop. Sci. 2000, 40, 1056–1061. [Google Scholar] [CrossRef]



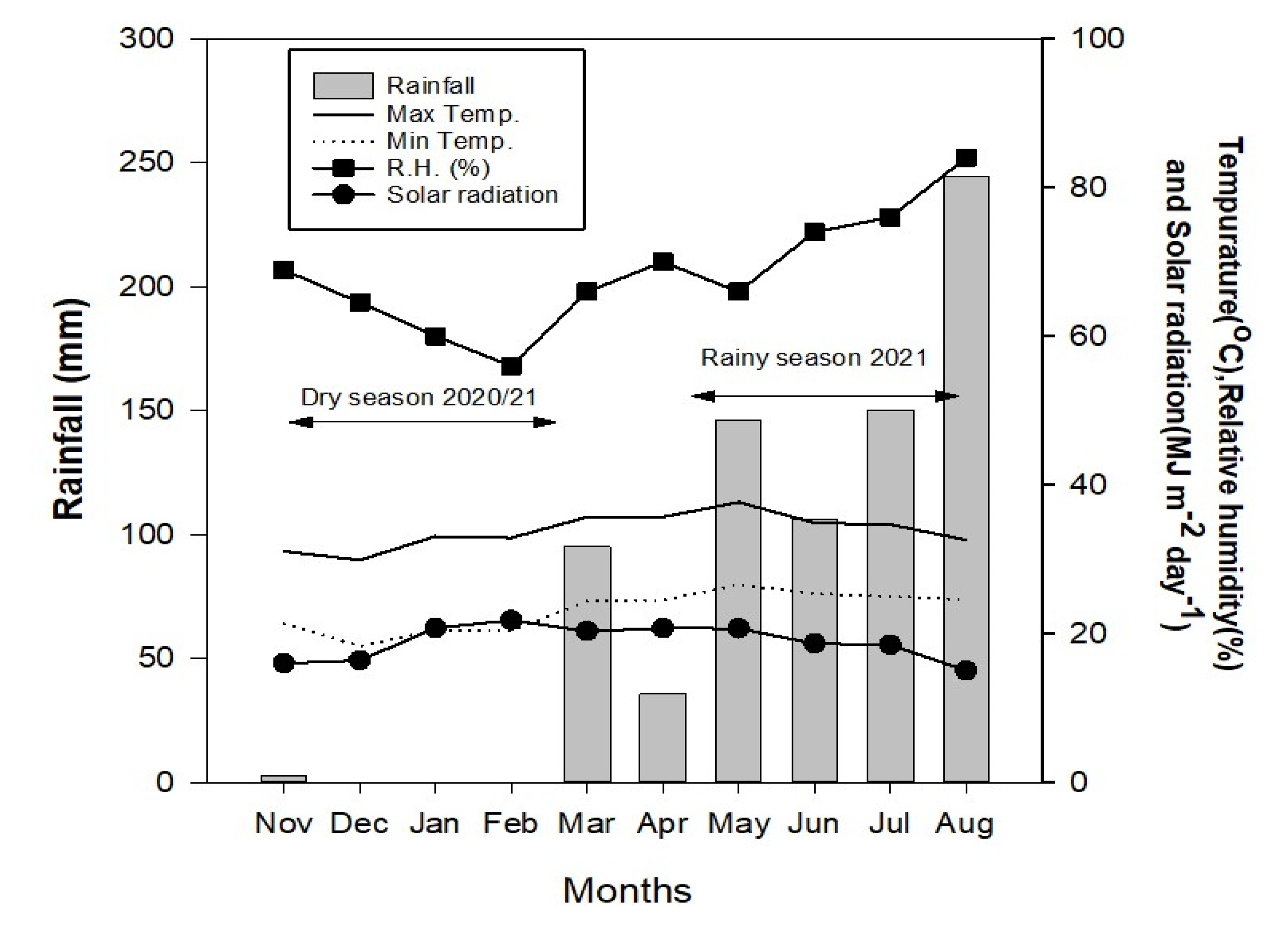{kind=link}
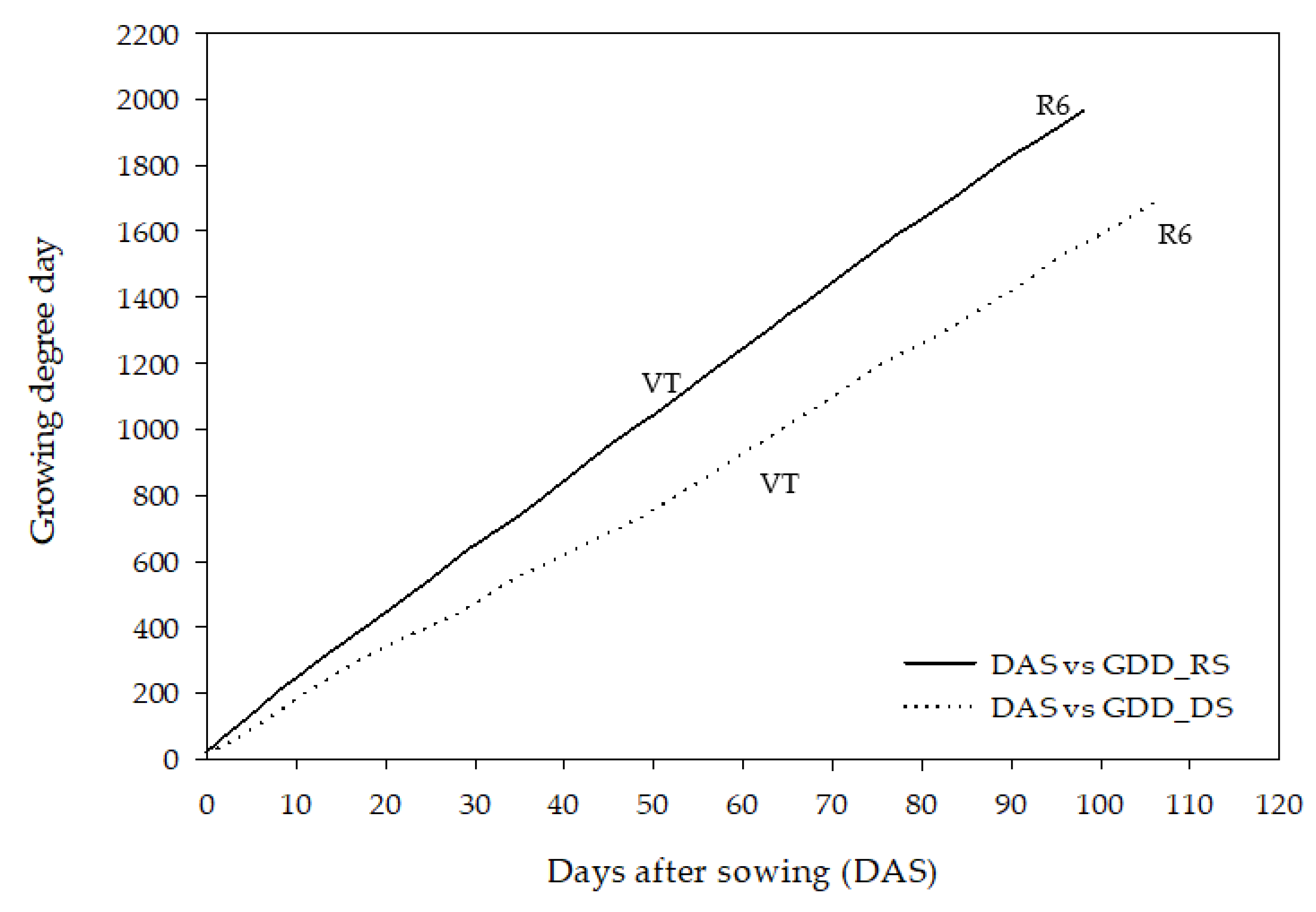{kind=link}
| SOV | df | HIR | ISR | Yield Components | |||||||
| EW | EL | ED | HC | NRE | NKR | CD | KD | ||||
| Season (S) | 1 | 10.1 ** | 164.7 ns | 57,150.1 ** | 76.9 ** | 40.3 ** | 6.3 ** | 403.3 ** | 1931.5 ** | 10.9 ** | 1.3 ** |
| (14.1) | (0.5) | (79.5) | (13.3) | (60.4) | (10.4) | (47.4) | (54.1) | (47.7) | (46.5) | ||
| Rep/S (a) | 4 | 0.1 | 41.9 | 3.7 | 0.3 | 0.0 | 0.1 | 0.4 | 1.9 | 0.0 | 0.0 |
| (0.7) | (0.5) | (0.0) | (0.2) | (0.2) | (0.7) | (0.2) | (0.2) | (0.2) | (0.9) | ||
| Genotype (G) | 13 | 1.8 ** | 1674.0 ** | 856.0 ** | 20.9 ** | 1.0 ** | 2.8 ** | 14.1** | 87.2 ** | 0.5 ** | 0.1 ** |
| (33.0) | (64.4) | (15.5) | (47.0) | (17.9) | (59.8) | (21.5) | (31.7) | (26.6) | (28.0) | ||
| G × S | 13 | 2.3 ** | 767.9 ** | 171.6 ** | 14.8 ** | 1.0 ** | 1.0 ** | 16.9 ** | 25.7 ** | 0.4 ** | 0.1 ** |
| (41.5) | (29.5) | (3.1) | (33.4) | (18.7) | (22.6) | (25.8) | (9.4) | (22.1) | (21.8) | ||
| Pooled error (b) | 52 | 0.1 | 33.3 | 25.9 | 0.7 | 0.0 | 0.1 | 0.8 | 3.2 | 0.0 | 0.0 |
| (10.7) | (5.1) | (1.9) | (6.1) | (2.8) | (6.6) | (5.2) | (4.6) | (3.3) | (2.8) | ||
| C.V.(a)(%) | 29.0 | 8.4 | 4.2 | 5.4 | 5.9 | 14.9 | 5.9 | 8.3 | 6.0 | 12.4 | |
| C.V.(b)(%) | 30.8 | 7.5 | 11.3 | 8.7 | 6.3 | 13.2 | 8.5 | 10.7 | 6.3 | 6.2 | |
| SOV | df | Agronomic traits | |||||||||
| GR | PSD | DTA | DSI | PH | EH | PTB | TTB | TSL | SPL | ||
| Season (S) | 1 | 342.5 * | 19.6 ** | 1288.6 ** | 3375.7 ** | 29,385.2 ** | 10,906.4 ** | 82.4 ** | 199.3 ** | 2979.6 ** | 1818.3 ** |
| (6.3) | (41.8) | (57.7) | (78.8) | (42.4) | (35.7) | (14.9) | (20.4) | (72.3) | (69.1) | ||
| Rep/S (a) | 4 | 24.7 | 0.1 | 2.9 | 1.5 | 98.7 | 41.9 | 0.2 | 3.4 | 4.2 | 5.9 |
| (1.8) | (0.9) | (0.5) | (0.1) | (0.6) | (0.5) | (0.1) | (1.4) | (0.4) | (0.9) | ||
| Genotype (G) | 13 | 213.9 ** | 0.7 ** | 44.6 ** | 46.8 ** | 2369.2 ** | 1234.6 ** | 24.6 ** | 42.8 ** | 51.0 ** | 29.3 ** |
| (51.4) | (19.2) | (26.0) | (14.2) | (44.5) | (52.5) | (57.6) | (57.0) | (16.1) | (14.5) | ||
| G × S | 13 | 72.0 ** | 0.9 ** | 11.7 ** | 9.3 ** | 400.9 ** | 135.5 ** | 8.6 ** | 11.3 ** | 24.1 ** | 22.8 ** |
| (17.3) | (24.2) | (6.8) | (2.8) | (7.5) | (5.8) | (20.2) | (15.0) | (7.6) | (11.3) | ||
| Pooled error (b) | 52 | 24.1 | 0.1 | 3.8 | 3.3 | 66.9 | 32.5 | 0.8 | 1.2 | 2.9 | 2.2 |
| (23.2) | (13.8) | (8.9) | (4.0) | (5.0) | (5.5) | (7.2) | (6.1) | (3.7) | (4.3) | ||
| C.V.(a)(%) | 5.8 | 8.3 | 3.0 | 2.2 | 7.3 | 9.8 | 4.0 | 14.0 | 7.6 | 14.0 | |
| C.V.(b)(%) | 5.8 | 8.9 | 3.4 | 3.2 | 6.0 | 8.6 | 8.2 | 8.1 | 6.3 | 8.5 | |
| Season | Agronomic Traits | |||||||||
| GR (%) | PSD (d) | DTA (d) | DSI (d) | PH (cm) | EH (cm) | PTB | TTB | TSL (cm) | SPL (cm) | |
| Dry | 83.3 ± 1.2 | 4.5 ± 0.1 | 61.1 ± 0.5 | 63.9 ± 0.6 | 154.1 ± 4.0 | 77.7 ± 2.7 | 11.7 ± 0.4 | 14.8 ± 0.5 | 33.0 ± 0.6 | 22.1 ± 0.5 |
| Rainy | 87.4 ± 1.3 | 3.5 ± 0.1 | 53.3 ± 0.5 | 51.2 ± 0.5 | 116.7 ± 2.7 | 54.9 ± 2.1 | 9.7 ± 0.4 | 11.7 ± 0.4 | 21.1 ± 0.6 | 12.8 ± 0.5 |
| p-value | 0.033 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | 0.004 | <0.001 | <0.001 |
| Season | HIR (%) | ISR (%) | Yield Components | |||||||
| EW (g) | EL (cm) | ED (cm) | HC | NRE | NKR | CD (cm) | KD (cm) | |||
| Dry | 1.6 ± 0.2 | 78.7 ± 1.9 | 71.1 ± 2.4 | 10.4 ± 0.2 | 3.7 ± 0.1 | 2.4 ± 0.1 | 13.0 ± 0.2 | 21.5 ± 0.5 | 2.3 ± 0.1 | 0.8 ± 0.1 |
| Rainy | 0.9 ± 0.2 | 75.9 ± 4.0 | 19.0 ± 1.7 | 8.5 ± 0.5 | 2.3 ± 0.1 | 1.8 ± 0.1 | 8.6 ± 0.5 | 11.9 ± 0.8 | 1.6 ± 0.1 | 0.5 ± 0.1 |
| p-value | 0.008 | 0.562 | <0.001 | 0.003 | <0.001 | 0.007 | <0.001 | <0.001 | <0.001 | <0.001 |
| Group | Haploid Inducers | Dry Season | Rainy Season | ||
|---|---|---|---|---|---|
| HIR (%) | ISR (%) | HIR (%) | ISR (%) | ||
| High | KHI 49 | 1.8 cd (A) | 74.0 bc (A) | 0.0 e (B) | 82.9 b (A) |
| KHI 59 | 0.8 ef (A) | 81.3 b (A) | 0.3 e (B) | 88.7 ab (A) | |
| KHI 54 | 2.5 bc (A) | 62.5 de (B) | 0.9 d (B) | 72.8 c (A) | |
| KHI 42 | 0.8 ef (B) | 93.6 a (A) | 1.8 a (A) | 95.2 a (A) | |
| KHI 47 | 1.7 cd (A) | 94.2 a (A) | 1.6 ab (A) | 92.1 ab (A) | |
| Moderate | KHI 5 | 1.8 cd (A) | 92.0 a (A) | 0.0 e (B) | 95.6 a (A) |
| KHI 65 | 3.4 a (A) | 71.4 cd (A) | 0.0 e (B) | 66.4 cd (A) | |
| KHI 66 | 3.3 ab (A) | 75.2 bc (A) | 1.5 ab (B) | 53.5 e (B) | |
| KHI 50 | 1.1 def (B) | 81.0 bc (A) | 1.8 a (A) | 61.0 de (B) | |
| KHI 80 | 0.6 f (A) | 63.8 de (A) | 0.0 e (B) | 0.0 f (B) | |
| Low | KHI 56 | 1.0 def (B) | 79.4 bc (B) | 1.5 ab (A) | 97.5 a (A) |
| KHI 61 | 0.4 f (B) | 80.9 bc (A) | 0.9 d (A) | 71.9 c (A) | |
| KHI 72 | 1.5 de (A) | 60.7 e (B) | 1.3 bc (A) | 94.3 a (A) | |
| KHI 55 | 1.7 d (A) | 91.8 a (A) | 1.0 cd (B) | 90.6 ab (A) | |
| No. | Genotypes | Pedigree | Group 1 |
|---|---|---|---|
| 1 | KHI 49 | WST/Stock6-S(C6)-IDLT2A-28-B | High |
| 2 | KHI 59 | WST/Stock6-S(C6)-IDLT2A-WS-B | High |
| 3 | KHI 54 | WST/Stock6-S(C6)-IDLT2A-34-1-B | High |
| 4 | KHI 42 | TL/Stock6-S(C6)-IDLT1B-93-B | High |
| 5 | KHI 47 | WST/Stock6-S(C6)-IDLT2A-24-B | High |
| 6 | KHI 5 | NSX/Stock6-S(C6)-IDLT1A-110-B | Moderate |
| 7 | KHI 65 | KND/Stock6-S(C6)-IDLT2B-22-B | Moderate |
| 8 | KHI 66 | TB/Stock6-S(C6)-IDLT3-4-B | Moderate |
| 9 | KHI 50 | WST/Stock6-S(C6)-IDLT2A-29-B | Moderate |
| 10 | KHI 80 | TB/Stock6-S(C6)-IDLT4-4-B | Moderate |
| 11 | KHI 56 | WST/Stock6-S(C6)-IDLT2A-36-B | Low |
| 12 | KHI 61 | KND/Stock6-S(C6)-IDLT2B-15-B | Low |
| 13 | KHI 72 | TB/Stock6-S(C6)-IDLT4-24-B | Low |
| 14 | KHI 55 | WST/Stock6-S(C6)-IDLT2A-35-B | Low |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sintanaparadee, P.; Dermail, A.; Lübberstedt, T.; Lertrat, K.; Chankaew, S.; Ruanjaichon, V.; Phakamas, N.; Suriharn, K. Seasonal Variation of Tropical Savanna Altered Agronomic Adaptation of Stock-6-Derived Inducer Lines. Plants 2022, 11, 2902. https://doi.org/10.3390/plants11212902
Sintanaparadee P, Dermail A, Lübberstedt T, Lertrat K, Chankaew S, Ruanjaichon V, Phakamas N, Suriharn K. Seasonal Variation of Tropical Savanna Altered Agronomic Adaptation of Stock-6-Derived Inducer Lines. Plants. 2022; 11(21):2902. https://doi.org/10.3390/plants11212902
Chicago/Turabian StyleSintanaparadee, Paepan, Abil Dermail, Thomas Lübberstedt, Kamol Lertrat, Sompong Chankaew, Vinitchan Ruanjaichon, Nittaya Phakamas, and Khundej Suriharn. 2022. "Seasonal Variation of Tropical Savanna Altered Agronomic Adaptation of Stock-6-Derived Inducer Lines" Plants 11, no. 21: 2902. https://doi.org/10.3390/plants11212902