
The Production of Helianthus Haploids: A Review of Its Current Status and Future Prospects


Abstract
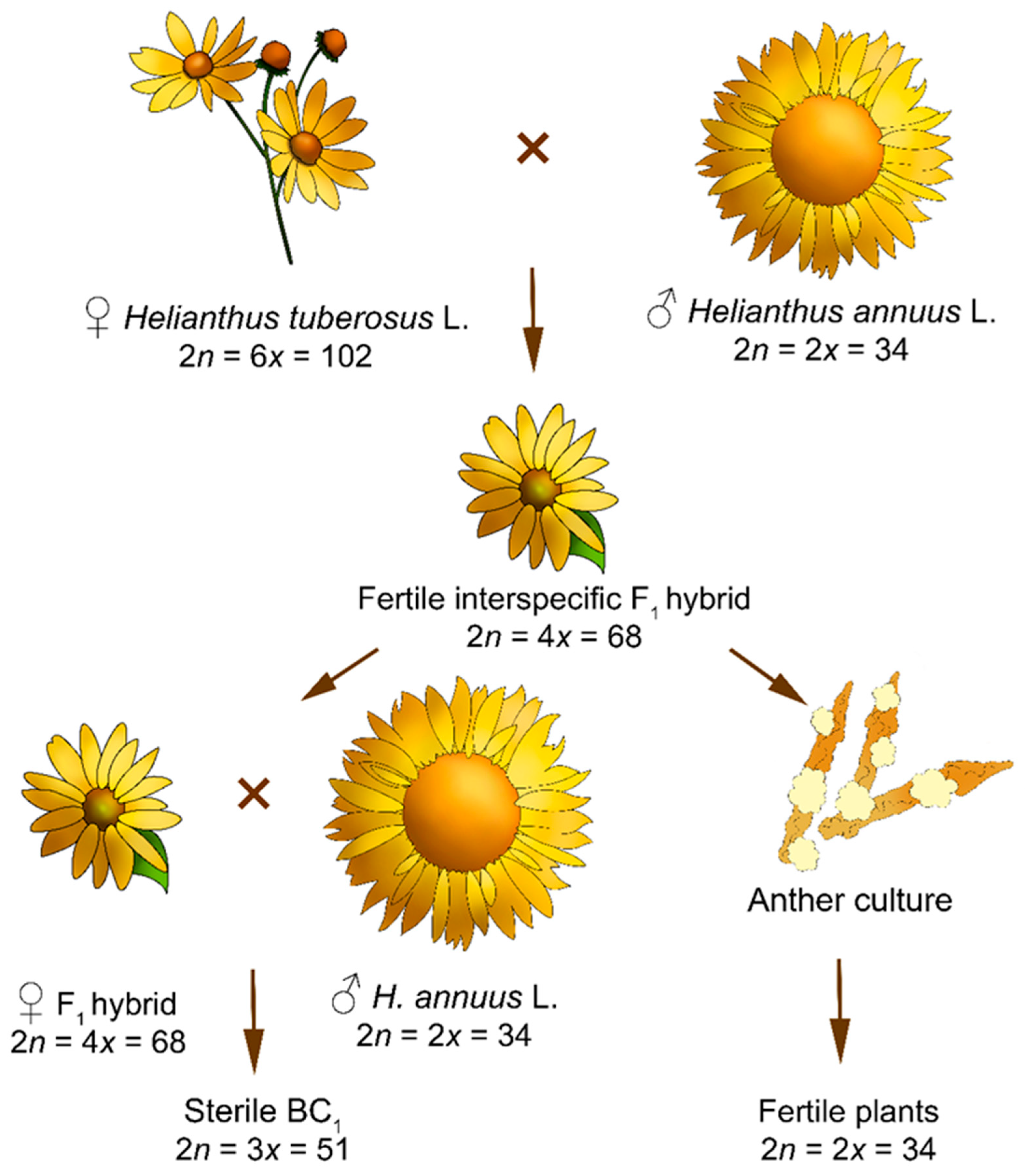
:1. Introduction
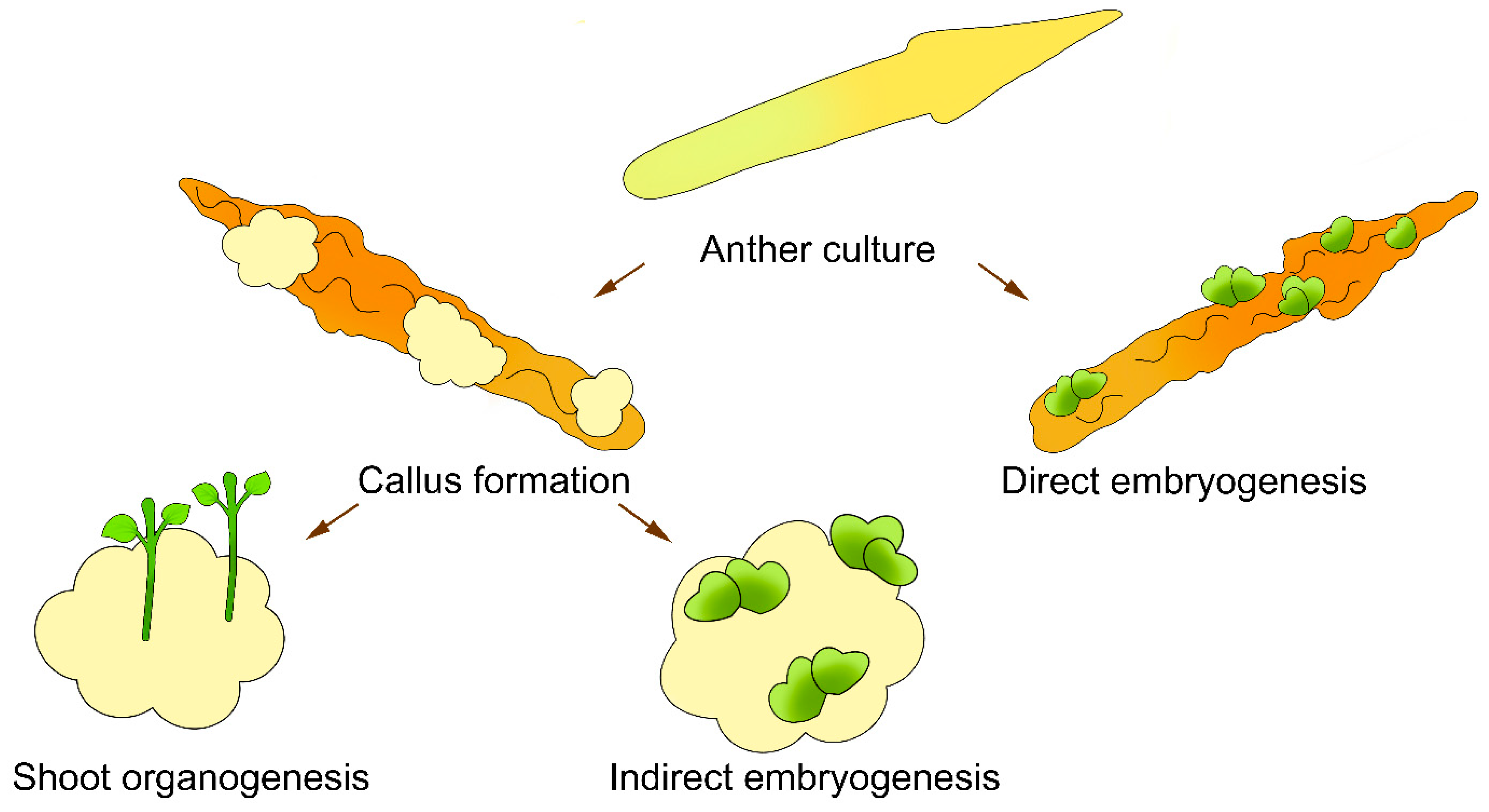
2. Production of Sunflower Haploids in Isolated Anther Cultures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Callus Induction Medium (CIM) | Shoot Induction Medium (SIM) | Results | Reference |
|---|---|---|---|---|
| Cultivar of H. annuus L., 2 F1 interspecific hybrids, 11 species of Helianthus | Modified Murashige and Skoog (MS) medium [55] + White’s vitamins [56] + 1 mg/L 2,4-dichlorophenoxyacetic acid (2,4-D) + 0.2 mg/L kinetin + 30 g/L sucrose | Modified MS + White’s vitamins + 2 mg/L 6-benzylaminopurine (BAP) + 0.2 mg/L indole-3-acetic acid (IAA) + 30 g/L sucrose | Non-morphogenic callus | [9] |
| Two F1 H. annuus L. hybrids | B1 (modified MS + White’s vitamins + 5 mg/L zeatin + 30 g/L sucrose) | P20 (modified N6 [57] + 1 mg/L zeatin + 31.65 g/L maltose) or BRl (modified MS + 0.1 mg/L 1-naphthaleneacetic acid (NAA) + 0.1 mg/L BAP + 0.01 mg/L gibberellic acid (GA3) + 30 g/L sucrose) | Callus formation in 65–68% of anthers, no shoot organogenesis | [45] |
| F1 hybrid H. annuus L. | M1 (½ MS macro- and microsalts + vitamins of Morel and Wetmore [58] + 0.5 mg/L NAA + 0.5 mg/L BAP + 120 g/L sucrose) | P20 or BRl | Callus formation in 95.7% of anthers, 3.4% shoot organogenesis, no data on ploidy level of plants | [45] |
| Two H. annuus L. inbred lines | M3 (½ MS macro- and microsalts + vitamins of Morel and Wetmore + 0.5 mg/L NAA + 0.5 mg/L BAP + 120 g/L sucrose) | P20 or BRl | Callus formation in 20–70%, 1.4–3.3% shoot organogenesis, no data on ploidy level of plants | [45] |
| One species of Helianthus, two interspecific F1 hybrids | MS + 0.5 mg/L NAA + 0.5 mg/L BAP | Gamborg (B5) medium [59] | Callus formation in 14.7–52.3%, 180 plantlets regenerated and rooted in one hybrid, mostly diploids | [52] |
| Interspecific hybrid | MS-I1 (MS + 0.5 mg/L BAP + 0.5 mg/L NAA + 30 g/L sucrose) | MS-R1 (MS + 0.5 mg/L BAP + 0.5 mg/L NAA + 30 g/L sucrose) | Callus formation in 96%, 187 regenerated shoots, 44 rooted plantlets, androgenetic origin of examined plants | [38] |
| Interspecific hybrid | MS-I4 (modified MS + 1 mg/L BAP + 1 mg/L NAA + 30 g/L sucrose) | Non-morphogenic callus in 96% | [38] | |
| Interspecific hybrid | MS-I5 (MS + 0.5 mg/L BAP + 0.5 mg/L NAA + 30 g/L sucrose) | MS-R1 | Callus formation in 75%, 85 regenerated shoots, 41 rooted plantlets, androgenetic origin of examined plants | [38] |
| Six interspecific F1 hybrids | L4 [60] + 30 g/L sucrose | B5 + 10 g/L sucrose | Callus formation in 50–100% with shoot regeneration in four hybrids, no data on ploidy level of plants | [51] |
| Cultivar of H. annuus L., two interspecific hybrids, three wild species | MS + 1 mg/L NAA + 2 mg/L 2,4-D + 0.5 mg/L BAP + 30 g/L sucrose | MS + 0.5 mg/L BAP + 30 g/L sucrose | Only one interspecific hybrid developed plantlets, no data on ploidy level of plants | [40] |
| Interspecific F1 hybrid | Modified MS + 1 mg/L BAP + 1 mg/L NAA + 30 g/L sucrose | MS + 0.5 mg/L BAP | Callus formation with 1.2% shoot regeneration, haploid plants | [17] |
| Five fertility restorer lines of H. annuus L., five wild species | MS + 1 mg/L NAA + 2 mg/L 2,4-D + 0.5 mg/L BAP + 30 g/L sucrose | MS + 0.5 mg/L BAP + 0.5 mg/L kinetin | Anthers of all genotypes were showed callus, shoot, and root regeneration in only two species, all plants were haploids | [48] |
| 74 cultivated sunflower plants in BC2 backcross generation | MS + 0.5 mg/L BAP + 0.5 mg/L NAA + 30 g/L sucrose | Shoot regeneration of 9.39% in 48.65% of the tested genotypes, no data on ploidy level of plants | [10] | |
| Two H. annuus L. cultivars | MS + 2 mg/L NAA + 1 mg/L BAP | Callus formation in 9–99% | [53] | |
| Five H. annuus L. lines | Modified MS + 1 mg/L IAA + 2 mg/L 2,4-D + 0.5 mg/L BAP + 30 g/L sucrose | MS + 1 mg/L kinetin + 0.1 mg/L IAA | Shoot regeneration of 21.03%, no data on ploidy level of plants | [49] |
| F1 H. annuus L. hybrids | MS + 2 mg/L NAA + 1 mg/L BAP | Callus formation in 90%, no shoot organogenesis, callus contained haploid and diploid cells | [14] | |
| Two F1 H. annuus L. hybrids | MS + 2 mg/L NAA + 1 mg/L BAP + 30 g/L sucrose or MS + 2 mg/L NAA + 0.5 mg/L BAP + 30 g/L sucrose | Callus formation in 34.95%, no data on ploidy level of plants | [50] | |
| Seven H. annuus L. F2 segregation populations | MS + 2 mg/L NAA + 2 mg/L 2,4-D + 0.5 mg/L BAP + 30 g/L sucrose | Callus formation in 8.3–66.7%, shoot regeneration of 0–6.67%, no data on ploidy level of plants | [50] | |
| Genotypes | Embryogenesis Induction Medium (EIM) | Embryo Development Medium (EDM) | Results | Reference |
|---|---|---|---|---|
| One Helianthus species, one interspecific hybrid | Modified MS medium + White’s vitamins + 5 mg/L zeatin + 30 g/L sucrose | Direct embryogenesis, 1–3 regenerated plants, different chromosome numbers | [9] | |
| Three H. annuus L. lines, five F1 H. annuus L. hybrids | ½ MS + vitamins of Morel and Wetmore + 0.5 mg/L NAA + 0.5 mg/L BAP + 120 g/L sucrose | Liquid medium with filter paper: macro- and microsalts of Monnier [61] + vitamins of Morel and Wetmore + 15 g/L sucrose | Embryogenic anthers in 2.1–54.9%, plants had haploid and diploid chromosome numbers | [13] |
| Two interspecific hybrids | M1 (½ MS macro- and microsalts + vitamins of Morel and Wetmore + 0.5 mg/L NAA + 0.5 mg/L BAP + 120 g/L sucrose) | P20 (modified N6 + 1 mg/L zeatin + 31.65 g/L maltose) or BRl (modified MS + 0.1 mg/L NAA + 0.1 mg/L BAP + 0.01 mg/L GA3 + 30 g/L sucrose) | Callus formation in both hybrids, one embryoid formed shoots, no data on ploidy level | [45] |
| Four H. annuus L. genotypes | MS + 1 mg/L 2,4-D + 0.5 mg/L BAP + 40 g/L sucrose | MS + 0.5 mg/L kinetin + 0.5 mg/L BAP | All genotypes produced callus, 10–15% of embryos germinated into plantlets for one genotype, no data on ploidy level of plants | [41] |
| Seven F1 hybrids, four H. annuus L. inbred lines | ½ MS macrosalts + MS microsalts + vitamins of Morel and Wetmore + 0.5 mg/L NAA + 0.5 mg/L BAP | Monnier medium + 0.05 mg/L BAP + reduced sucrose concentrations (10% for first week, 6% for second, 3% thereafter) | Callogenic anthers in 16–18%, embryogenic anthers in 1–11%, direct and indirect embryogenesis, all plantlets were diploid and originated from somatic cells | [39] |
| Two interspecific hybrids | MS-I3 (modified MS + 0.5 mg/L BAP + 0.5 mg/L NAA + 30 g/L sucrose) | MS-R3 (modified MS + 30 g/L sucrose) | Direct embryogenesis in up to 92.7%, androgenetic origin of examined plants | [38] |
| One cultivar, three H. annuus L. hybrids | Callus formation: MS + 2 mg/L NAA + 1 mg/L BAP + 30 g/L sucrose; indirect embryo initiation: MS + 0.1 mg/L NAA + 0.5 mg/L BAP | MS + 0.5 mg/L BAP | Calli produced in >90% of anthers, 44% of calli differentiated into embryos, low rate of embryo development, plants were haploid and diploid | [42] |
| Interspecific F1 hybrid | Modified MS + 1 mg/L BAP + 1 mg/L NAA + 30 g/L sucrose | MS + 0.5 mg/L BAP | All anthers developed embryos, regeneration of 98.7%, haploid plants | [17] |
| Six sunflower cultivars | MS + 0.5 mg/L BAP + 0.5 mg/L NAA + 30 g/L sucrose | Callus formation in 50–90%, embryo formation in 0–51.7%, no data on ploidy level of produced plants | [46] | |
| H. annuus L. cultivar | MS + 1 mg/L BAP + 2 mg/L NAA + 30 g/L sucrose | Callus formation in 60.44%, indirect embryogenesis in 20.45%, no data on ploidy level of produced plants | [43] | |
| Two H. annuus L. cultivars | MS + 0.5 mg/L 2,4-D + 0.5 mg/L BAP + 30 g/L sucrose | MS medium + 0.5 mg/L kinetin | Embryogenic callus formation in 81–88%, regeneration in 2–9%, no data on ploidy level of plants | [44] |
2.1. Genotype
2.2. Conditions for Growing Donor Plants
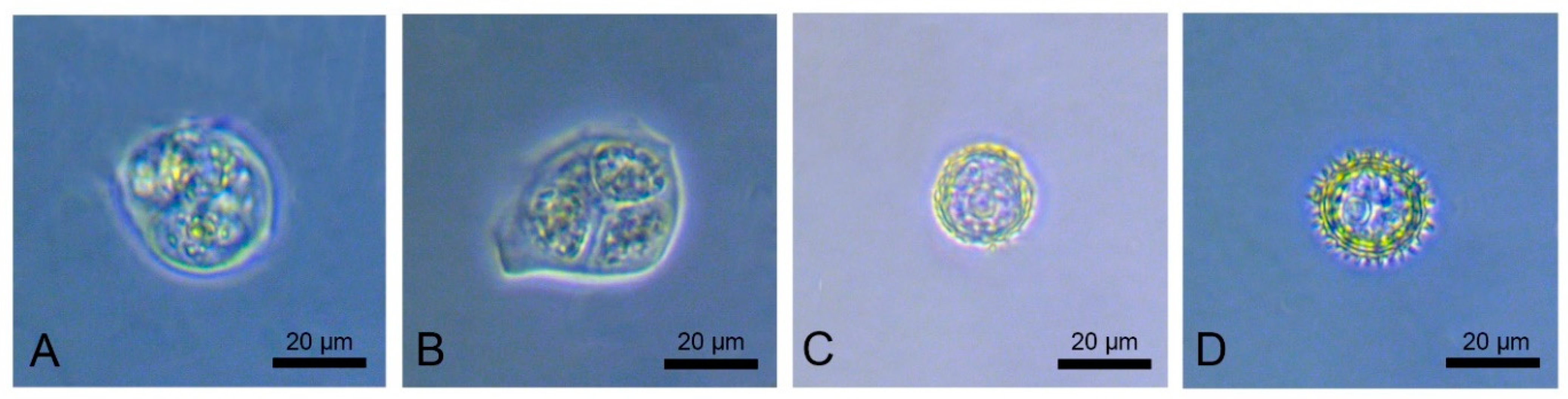
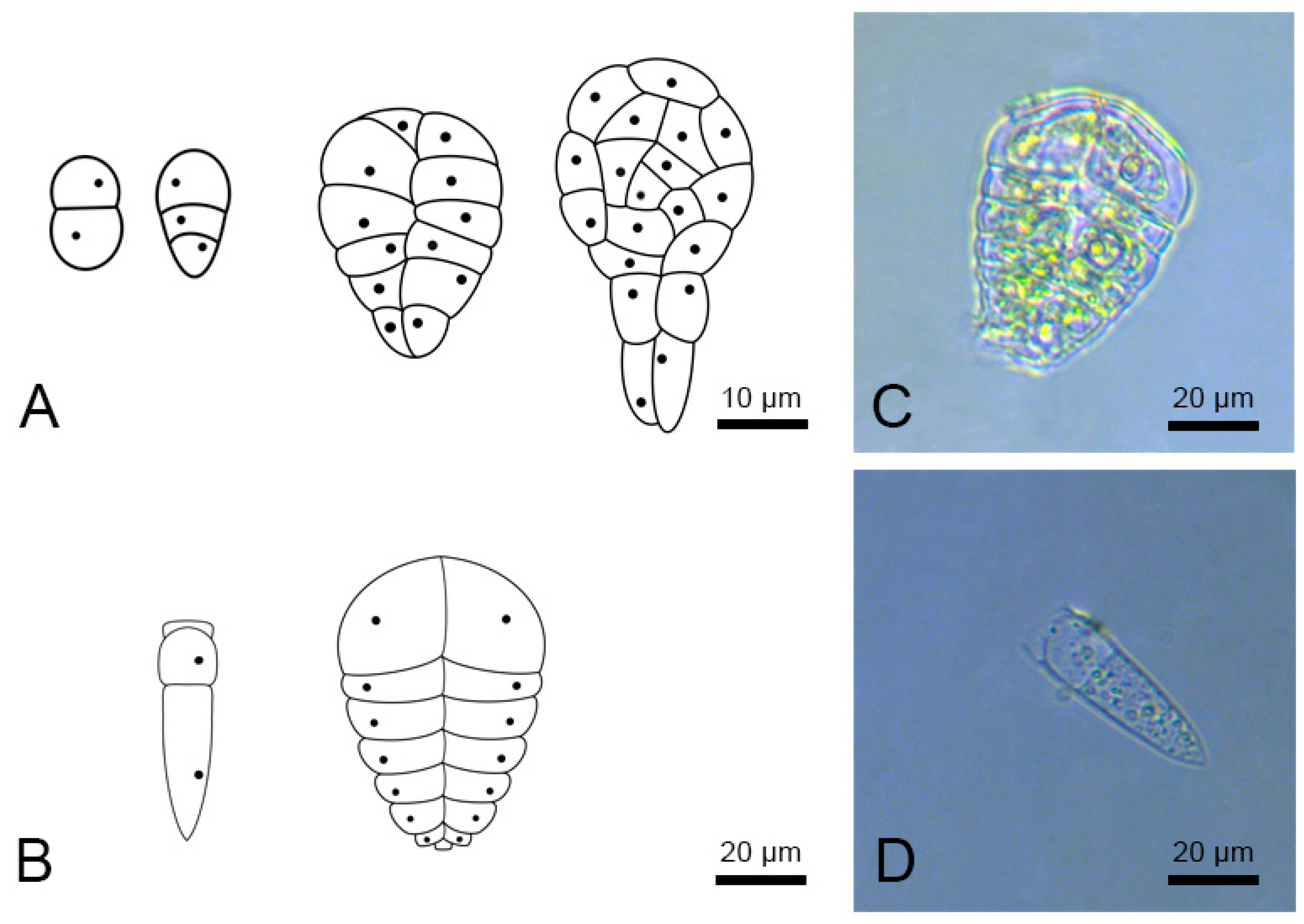
2.3. Stages of Microspore Development
2.4. Cold Pretreatment of Inflorescences and Flower Buds
2.5. Composition of Culture Medium
2.6. Culture Conditions
3. Producing Sunflower Haploids in Isolated Microspore Culture
4. Producing Sunflower Haploids by the In Vitro Culture of Unpollinated Ovaries and Ovules
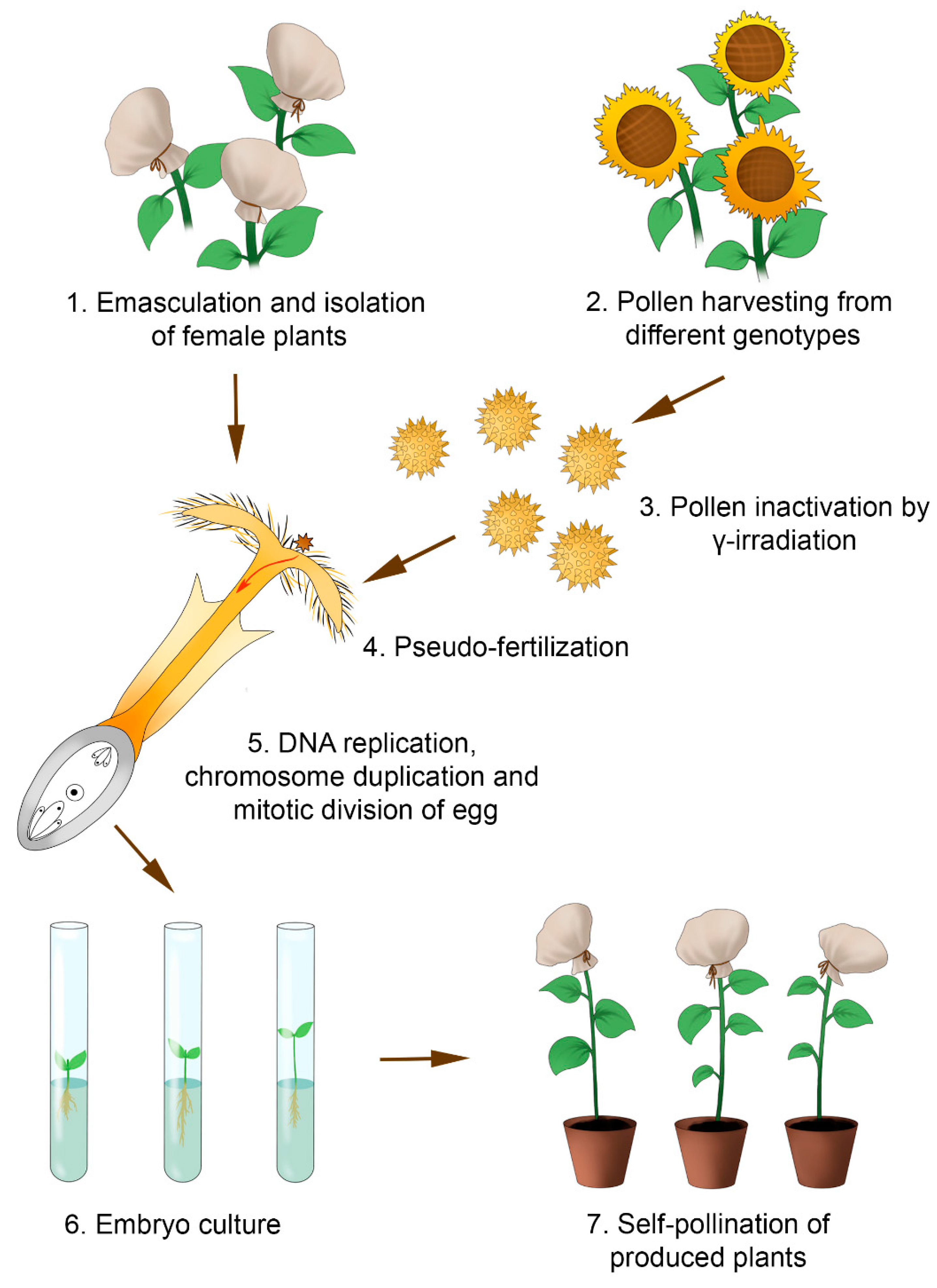
5. Induced Parthenogenesis by γ-Irradiated Pollen
6. Induced Parthenogenesis by Distant Hybridization
7. Methods for Doubling the Sunflower Haploid Chromosome Set and Determining the Ploidy Level
8. Achievements in Sunflower Breeding Programs by the Implementation of Haploid and Doubled Haploid Plants
9. Conclusions and Future Prospects
- (1)
- Inducing parthenogenesis by γ-irradiated pollen is the most efficient method for producing haploid sunflower plants. The dihaploid lines obtained by this method have been involved in commercial breeding programs to produce high-yielding F1 hybrids with an enhanced resistance to abiotic and/or biotic stresses. However, several parameters (the dose of γ-irradiation, dependence on the genotype, embryo survival) mean that this method is substantially limited compared to the classical production of inbred parental lines through cycles of self-pollination.
- (2)
- Conventional in vitro methods for haploid induction in Helianthus plants using male (isolated microspore and anther cultures) or female (unpollinated ovaries and ovules culture) gametophytes are still not widely used in biotechnological practice due to their low embryogenic response. A radical change in this situation could be achieved by identifying genotypes with a strong ability to undergo gametic embryogenesis using a fine-mapping approach with quantitative trait loci and improving them through various genetic engineering strategies, as well as optimizing physiological factors such as the culture conditions of mother plants, the stage of development of the gametic cells, the culture media composition, and the culture conditions, etc.
- (3)
- Alternative in vivo methods for haploid induction in Helianthus plants via induced parthenogenesis by distant hybridization, including manipulations with the centromere-specific histone (CENH3), which plays a crucial role in uniparental genome elimination during early embryogenesis, will be of great fundamental and practical value in the future.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rauf, S. Breeding Strategies for Sunflower (Helianthus annuus L.) Genetic Improvement. In Advances in Plant Breeding Strategies: Industrial and Food Crops; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Springer: Cham, Switzerland, 2019; Volume 6, pp. 637–673. [Google Scholar]
- Atlagic, J.; Secerov-Fiser, V.; Marinkovic, R. Interspecific Hybridisation and Cytogenetic Studies in Ornamental Sunflower Breeding. Aust. J. Exp. Agric. 2005, 45, 93–97. [Google Scholar] [CrossRef]
- Fernández-Martínez, J.M.; Pérez-Vich, B.; Velasco, L. Sunflower. In Oil Crops. Handbook of Plant Breeding; Vollmann, J., Rajcan, I., Eds.; Springer: New York, NY, USA, 2009; Volume 4, pp. 155–232. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 18 July 2022).
- Adeleke, B.S.; Babalola, O.O. Oilseed Crop Sunflower (Helianthus annuus) as a Source of Food: Nutritional and Health Benefits. Food Sci. Nut. 2020, 8, 4666–4684. [Google Scholar] [CrossRef] [PubMed]
- Kaya, Y. Sunflower. In Alien Gene Transfer in Crop Plants; Pratap, A., Kumar, J., Eds.; Springer: New York, NY, USA, 2014; Volume 2, pp. 281–315. [Google Scholar]
- Dimitrijevic, A.; Horn, R. Sunflower Hybrid Breeding: From Markers to Genomic Selection. Front. Plant Sci. 2018, 8, 2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drumeva, M.; Yankov, P.; Nenova, N.; Shindrova, P. Investigation on the Resistance of Doubled Haploid Sunflower Lines to Some Biotic Factors. Agric. Sci. Technol. 2014, 6, 11–13. [Google Scholar]
- Bohorova, N.E.; Atanassov, A.I. Sunflower (Helianthus annuus L.): In vitro Production of Haploids. In Haploids in Crop Improvement I; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1990; Volume 12, pp. 428–441. [Google Scholar]
- Miladinovic, D.; Kovacevic, B.; Dimitrijevic, A.; Imerovski, I.; Jocic, S.; Cvejic, S.; Miklic, V. Towards Dihaploid Production in Sunflower- Selection of Regeneration Medium. In Proceedings of the 18th International Sunflower Conference, Mar del Plata, Argentina, 27 February–1 March 2012; International Sunflower Association: Paris, France, 2012; pp. 674–677. [Google Scholar]
- Drumeva, M.; Yankov, P.; Nenova, N.; Shindroova, P.; Encheva, V. New Sunflower Restorer Lines Developed by γ-induced Parthenogenesis from Helianthus annuus Hybrids- Disease Resistance, Combining Ability. I. Disease Resistance. In Proceedings of the International Conference Conventional and Molecular Breeding of Field and Vegetable Crops, Novi Sad, Serbia, 24–27 November 2008. [Google Scholar]
- Dağüstü, N.; Bayram, G.; Sincik, M.; Bayraktaroğlu, M. The Short Breeding Cycle Protocol Effective on Diverse Genotypes of Sunflower (Helianthus annuus L.). Turk. J. Field Crops 2012, 17, 124–128. [Google Scholar]
- Jonard, R.; Mezzarobba, A. Sunflower (Helianthus spp.): Anther Culture and Field Studies on Haploids. In Legumes and Oilseed Crops I. Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1990; Volume 10, pp. 485–501. [Google Scholar]
- Dayan, S.; Arda, H. Effects of Naphthaleneacetic Acid and N6- benzyladenine on Androgenesis in Helianthus annuus L. In Proceedings of the 19th International Sunflower Conference, Edirne, Turkey, 30 May–1 June 2016; pp. 177–184. [Google Scholar]
- Nurhidayah, T.; Koehler, H.; Dahlhoff, M.; Friedt, W. Anther Culture of Interspecific Sunflower Hybrids and Examination of Regenerants by Biochemical and Molecular Methods. Biotechnol. Biotechnol. Equip. 1993, 7, 113–116. [Google Scholar] [CrossRef]
- Drumeva, M. Developing sunflower fertility restorer lines from commercial hybrids by using in vitro technique. Agric. Sci. Technol. 2012, 4, 361–364. [Google Scholar]
- Sujatha, M.; Prabakaran, A.J. Ploidy Manipulation and Introgression of Resistance to Alternaria helianthi from Wild Hexaploid Helianthus Species to Cultivated Sunflower (H. annuus L.) Aided by Anther Culture. Euphytica 2006, 152, 201–215. [Google Scholar] [CrossRef]
- Rokka, V.M. Potato Haploids and Breeding. In Advances in Haploid Production in Higher Plants; Touraev, A., Forster, B.P., Jain, S.M., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 199–208. [Google Scholar]
- Germana, M.A. Gametic Embryogenesis and Haploid Technology as Valuable Support to Plant Breeding. Plant Cell Rep. 2011, 30, 839–857. [Google Scholar] [CrossRef]
- Davey, M.R.; Jan, M. Sunflower (Helianthus annuus L.): Genetic Improvement Using Conventional and in vitro Technologies. J. Crop Improv. 2010, 24, 349–391. [Google Scholar] [CrossRef]
- National Sunflower Association. Available online: https://www.sunflowernsa.com/Research/NSA-Funded-Research/ (accessed on 19 July 2022).
- Friedt, W.; Nichterlein, K.; Dahlhoff, M.; Köhler, H.; Gürel, A. Recent Progress and Prospects of Biotechnology in Sunflower Breeding. Lipid/Fett 1991, 93, 368–374. [Google Scholar] [CrossRef]
- Friedt, W. Present State and Future Prospects of Biotechnology in Sunflower Breeding. Field Crops Res. 1992, 30, 425–442. [Google Scholar] [CrossRef] [Green Version]
- Friedt, W.; Nurhidayah, T.; Röcher, T.; Köhler, H.; Bergmann, R.; Horn, R. Haploid Production and Application of Molecular Methods in Sunflower (Helianthus annuus L.). In In Vitro Haploid Production in Higher Plants; Jain, S.M., Sopory, S.K., Veilleux, R.E., Eds.; Springer: Dordrecht, The Netherlands, 1997; Volume 5, pp. 17–35. [Google Scholar]
- Thierfelder, A.; Lühs, W.; Friedt, W. Breeding of Industrial Oil Crops with the Aid of Biotechnology: A Review. Indust. Crops Prod. 1992, 1, 261–271. [Google Scholar] [CrossRef]
- Palmer, C.E.; Keller, W.A. In vitro Culture of Oilseeds. In Plant Cell and Tissue Culture; Vasil, I.K., Thorpe, T.A., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 413–455. [Google Scholar]
- Vasić, D. Use of Methods of Biotechnology and Molecular Biology in Sunflower Breeding: Achievements in the World. Zbornik radova Instituta za Ratarstvo i Povrtarstvo 2001, 35, 259–269. [Google Scholar]
- Ivanov, P.; Encheva, J.; Nenova, N.; Todorova, M. Application of Some Biotechnological Achievements in Sunflower Breeding. Helia 2002, 25, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Nichterlein, K.; Horn, R. Haploids in the Improvement of Linaceae and Asteraceae. In Haploids in Crop Improvement II. Biotechnology in Agriculture and Forestry; Don Palmer, C., Keller, W.A., Kasha, K.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 56, pp. 277–294. [Google Scholar]
- Dagustu, N. In vitro Tissue Culture Studies in Sunflower (Helianthus spp.). Ekin J. Crop Breed. Gen. 2018, 4, 13–21. [Google Scholar]
- Karjee, S.; Mahapatra, S.; Singh, D.; Saha, K.; Viswakarma, P.K. Production of Double Haploids in Ornamental Crops. J. Pharmacogn. Phytochem. 2020, 9, 555–565. [Google Scholar]
- Yang, H.Y.; Yan, H.; Zhou, C. In vitro Production of Haploids in Helianthus. In Legumes and Oilseed Crops I. Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1990; Volume 10, pp. 472–484. [Google Scholar]
- Bohorova, N.; Atanassov, A.; Antonova, G. In vitro Isolation of Anthers from Interspecific Hybrids in the Helianthus Genus. CR Acad. Sci. Bulg. 1980, 33, 1545–1548. [Google Scholar]
- Bohorova, N.; Atanassov, A.; Georgieva-Todorova, J. In vitro Organogenesis, Androgenesis and Embryo Culture, in the Genus Helianthus L. Z. Pflanzenzüchtg. 1985, 95, 35–44. [Google Scholar]
- Alissa, A.; Serieys, H.; Jonard, R. Wild Species and Interspecific Hybrid Regeneration in Helianthus Genus by in vitro Androgenesis. Compt. Rend. Acad. Sci. 1985, 300, 25–30. [Google Scholar]
- Bohorova, N.; Landjeva, S. A Cytogenetical Study on Anther Ca1lus Culture of Helianthus Species in Different Ploidy Level. Genet Breed 1987, 19, 509–516. [Google Scholar]
- Mix, G. Anther and Ovary Culture of Helianthus annuus L. Landbauforsch Volkenrode 1985, 35, 153–156. [Google Scholar]
- Nurhidayah, T.; Horn, R.; Röcher, T.; Friedt, W. High Regeneration Rates in Anther Culture of Interspecific Sunflower Hybrids. Plant Cell Rep. 1996, 16, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Michaux-Ferriére, N.; Coumans, M. Assay for Doubled Haploid Sunflower (Helianthus annuus) Plant Production by Androgenesis: Fact or Artifact? Part 1. In vitro Anther Culture. Plant Cell Tissue Organ Cult. 1995, 41, 91–97. [Google Scholar] [CrossRef]
- Priya, K.V.; Sassikumar, D.; Sudhagar, R.; Gopalan, A. Androgenetic Response of Sunflower in Different Culture Environments. Helia 2003, 26, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Thengane, S.R.; Joshi, M.S.; Khuspe, S.S.; Mascarenhas, A.F. Anther Culture in Helianthus annuus L., Influence of Genotype and Culture Conditions on Embryo Induction and Plant Regeneration. Plant Cell Rep. 1994, 13, 222–226. [Google Scholar] [CrossRef]
- Saji, K.; Sujatha, M. Embryogenesis and Plant Regeneration in Anther Culture of Sunflower (Helianthus annuus L.). Euphytica 1998, 103, 1–7. [Google Scholar] [CrossRef]
- Saensee, K.; Machikowa, T.; Kaya, Y. Relationship between Floret Size and Anther Culture Response in an Ornamental Sunflower. Asia-Pac. J. Sci. Technol. 2018, 23, 1–9. [Google Scholar]
- Çakmak, E.; Uncuoğlu, A.A.; Aydın, Y. Evaluation of in vitro Genotoxic Effects Induced by in vitro Anther Culture Conditions in Sunflower. Plant Signal. Behav. 2019, 14, 1633885. [Google Scholar] [CrossRef]
- Gürel, A.; Nichterlein, K.; Friedt, W. Shoot Regeneration from Anther Culture of Sunflower (Helianthus annuus) and Some Interspecific Hybrids as Affected by Genotype and Culture Procedure. Plant Breed. 1991, 106, 68–76. [Google Scholar] [CrossRef]
- Vasić, D.; Skorić, D.; Jocić, S. Anther Culture of Sunflower Cultivars. In Proceedings of the 15th International Sunflower Conference, Toulouse, France, 12–15 June 2000; pp. 52–55. [Google Scholar]
- Nenova, N.; Ivanov, P.; Christov, M. Anther Culture Regeneration of F1 Hybrids of Helianthus annuus × Helianthus smitii and Helianthus annuus × Helianthus eggertii. In Proceedings of the 13th International Sunflower Conference, Piza, Italy, 7–11 September 1992; pp. 1509–1514. [Google Scholar]
- Chigrin, T.V.; Zadorozhnaya, O.A.; Yushkina, L.L. Androgenesis Ability under in vitro Cultivation of Anthers of Different Sunflower Species. In Innovative Research Trends in Breeding and Cultivation Technology of Oil Crops, Proceedings of the 6th International Conference of the Young Scientist and Experts, Krasnodar, Russia, 24–25 February 2011; VNIIMK: Krasnodar, Russia, 2011; pp. 357–361. [Google Scholar]
- Kostina, E.E.; Lobachev, Y.V.; Tkachenko, O.V. Androgenesis in the in vitro Anther Culture of Genetically Marked Sunflower Lines. Sovr. Probl. Nauki Obraz. 2015, 3, 613. (In Russian) [Google Scholar]
- Garkusha, S.; Savenko, E.; Glazyrina, V.; Demurin, Y.; Gorlova, L.; Martynova, E.; Goryunova, S.; Goryunov, D.; Mukhina, Z. Development of Methodological Procedures for Culturing Sunflower Anthers in vitro. J. Biotech. Res. 2017, 8, 138–150. [Google Scholar]
- Todorova, M.; Nenova, N.; Ivanov, P.; Christov, M. Plant Regeneration through Anther Culture and Induced Parthenogenesis in Genus Helianthus. Biotechnol. Biotechnol. Equip. 1997, 11, 27–30. [Google Scholar] [CrossRef]
- Nenova, N.; Ivanov, P.; Christov, M. Anther Culture Regeneration from Helianthus mollis (M-020) and its Hybrid Combinations. Biotechnol. Biotechnol. Equip. 1998, 12, 58–60. [Google Scholar] [CrossRef]
- Krudnak, A.; Muangsan, N.; Machikowa, T. High Frequency Callus Induction through Anther Culture in High Oil Sunflower (Helianthus annuus L.). Asia-Pac. J. Sci. Technol. 2013, 18, 62–72. [Google Scholar]
- Prabakaran, A.J.; Sujatha, M. Breeding for Alternaria Resistance in Sunflower: Approaches for Introgression from Wild Sunflowers. In Proceedings of the 15th International Sunflower Conference, Toulouse, France, 12–15 June 2000; pp. 31–36. [Google Scholar]
- Murashige, T.; Skoog, F.A. Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Culture. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- White, P.R. The Cultivation of Animal and Plant Cells; The Ronald Press: New York, NY, USA, 1963; p. 228. [Google Scholar]
- Chu, C.C.; Wang, C.C.; Sun, C.S.; Hsu, C.; Yin, K.C.; Chu, C.Y.; Bi, F.Y. Establishment of an Efficient Medium for Anther Culture of Rice Through Comparative Experiments on the Nitrogen Sources. Sci. Sinica. 1975, 18, 659–669. [Google Scholar]
- Morel, G.; Wetmore, R.H. Fern Callus Tissue Culture. Am. J. Bot. 1951, 38, 141–143. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.; Ojima, K. Nutrient Requirements of Suspension Cultures of Soybean Root Cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Lenée, P.; Chupeau, Y. Isolation and Culture of Sunflower Protoplasts (Helianthus annuus L.): Factors Influencing the Viability of Cell Colonies Derived from Protoplasts. Plant Sci. 1986, 43, 69–75. [Google Scholar] [CrossRef]
- Monnier, M. Croissance et Développement des Embryons Globulaires de Capsella Barsa-pastoris Cultivés in vitro Dans un Milieu à Base D’une Nouvelle Solution Minérale. Bull. Société Bot. Fr. 1973, 120 (Suppl. S1), 179–194. [Google Scholar] [CrossRef]
- Sarrafi, A.; Bolandi, A.R.; Serieys, H.; Berville, A.; Alibert, G. Analysis of Cotyledon Culture to Measure Genetic Variability for Organogenesis Parameters in Sunflower (Helianthus annuus L.). Plant Sci. 1996, 121, 213–219. [Google Scholar] [CrossRef]
- Deglene, L.; Lesignes, P.; Alibert, G.; Sarrafi, A. Genetic Control of Organogenesis in Cotyledons of Sunflower (Helianthus annuus). Plant Cell Tissue Org. Cult. 1997, 48, 127–130. [Google Scholar] [CrossRef]
- Flores Berrios, E.; Gentzbittel, L.; Kayyal, H.; Alibert, G.; Sarrafi, A. AFLP Mapping of QTLs for in vitro Organogenesis Traits Using Recombinant Inbred Lines in Sunflower (Helianthus annuus L.). Theor. Appl. Gen. 2000, 101, 1299–1306. [Google Scholar] [CrossRef]
- Petitprez, M.; Sarrafi, A.; Flores-Berrios, E.; XuHan, X.; Briere, C.; Gentzbittel, L. Somatic Embryogenesis by Liquid Culture of Epidermal Layers in Sunflower: From Genetic Control to Cell Development. In Liquid Culture Systems for in vitro Plant Propagation; Hvoslef-Eide, A.K., Preil, W., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 313–322. [Google Scholar]
- Miladinović, D.; Kovačević, B.; Jocić, S.; Imerovski, I.; Dimitrijević, A.; Cvejić, S.; Balalić, I. The Effect of Sowing Date and Density on Callus Induction and Shoot Regeneration from Sunflower Anthers. In Proceedings of the 19th International Sunflower Conference, Edirne, Turkey, 29 May–3 June 2016; International Sunflower Association: Paris, France, 2016; pp. 509–514. [Google Scholar]
- Schneiter, A.A.; Miller, J.F. Description of Sunflower Growth Stages 1. Crop Sci. 1981, 21, 901–903. [Google Scholar] [CrossRef]
- Ferrie, A.M.R.; Caswell, K.L. Isolated Microspore Culture Techniques and Recent Progress for Haploid and Doubled Haploid Plant Production. Plant Cell Tissue Org. Cult. 2011, 104, 301–309. [Google Scholar] [CrossRef]
- Gürel, A.; Kontowski, S.; Nichterlein, K.; Friedt, W. Embryogenesis in Microspore Culture of Sunflower (Helianthus annuus L.). Helia 1991, 14, 123–128. [Google Scholar]
- Todorova, M.; Dalhoff, M.; Friedt, W. Microspore Culture in Sunflower (Helianthus annuus L.). Biotechnol. Biotechnol. Equip. 1993, 7, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Coumans, M.; Zhong, D. Doubled Haploid Sunflower (Helianthus annuus) Plant Production by Androgenesis: Fact or Artifact? Part 2. In vitro Isolated Microspore Culture. Plant Cell Tissue Org. Cult. 1995, 41, 203–209. [Google Scholar] [CrossRef]
- Bal, U.; Shariatpanahi, M.E.; Castro, A.J.; Emery, D.; Clement, C.; Dehestani-Ardakani, M.; Mozaffari, K.; Touraev, A. Pseudo-embryogenic Structures in Anther and Isolated Microspore Cultures in vitro: A Cautionary Guide. Czech J. Genet. Plant Breed. 2012, 48, 51–60. [Google Scholar] [CrossRef] [Green Version]
- De Leonardis, A.; Albanese, T.; Macciola, V. Biodegradation in vivo and in vitro of Chlorogenic Acid by a Sunflower-seedling (Helianthus annuus) Like-polyphenoloxidase Enzyme. Eur. Food Res. Technol. 2006, 223, 295–301. [Google Scholar] [CrossRef]
- Baimukhametova, E.A.; Kuluev, B.R. Darkening of Plant Tissues during in vitro Cultivation and Methods for its Prevention. Biotekhnologiya 2020, 36, 26–42. [Google Scholar] [CrossRef]
- Cai, D.T.; Zhou, C. In vitro Induction of Haploid Embryoids and Plantlets from Unpollinated Young Florets and Ovules of Helianthus annuus L. Kexue Tongbao 1984, 29, 680–682. [Google Scholar]
- Gelebart, P.; San, L.H. Production of Haploid Plants in Sunflower (Helianthus annuus L.) by in vitro Culture of Nonfertilized Ovaries and Ovules. Agronomie 1987, 7, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Dong, J.; Zhou, C.; Yang, H. Regulation of in vitro Parthenogenesis and Somatic Proliferation in Sunflower by Several Factors. Acta Bot. Sin. 1987, 29, 580–587. [Google Scholar]
- Yan, H.; Zhou, C.; Yang, H. Experimental Studies on Several Factors Affecting Unfertilized Ovule Culture in Sunflower. J. Wuhan Bot. Res. 1988, 6, 319–326. [Google Scholar]
- Yang, H.Y. The Use of a Whole Stain-Clearing Technique for Observations on Embryo Sac, Embryo, Endosperm and Embryoid. Acta Bot. Sin. 1986, 28, 575–581. [Google Scholar]
- Wei, Z.; Yang, H. Histochemical Studies on in vitro Parthenogenesis in Helianthus annuus L. Acta Bot. Res. 1986, 28, 117–122. [Google Scholar]
- Yan, H.; Yang, H.Y.; Jensen, W.A. An Electron Microscope Study on in vitro Parthenogenesis in Sunflower. Sex. Plant Reprod. 1989, 2, 154–166. [Google Scholar] [CrossRef]
- Yan, H.; Wu, Y.C.X.; Wei, Z.; Zhou, C.; Yang, H. Microscopical Observations on the Embryoid Formation in Cultured Unfertilized Ovules of Helianthus annuus L. Acta Bot. Sin. 1985, 27, 13–18. [Google Scholar]
- Ferrant, V.; Bouharmont, J. Origin of Gynogenetic Embryos of Beta vulgaris L. Sex. Plant Reprod. 1994, 7, 12–16. [Google Scholar] [CrossRef]
- Musial, K.; Bohanec, B.; Jakse, M.; Przywara, L. The Development of Onion (Allium cepa L.) Embryo Sacs In vitro and Gynogenesis Induction in Relation to Flower Size. Vitr. Cell. Dev. Biol. Plant. 2005, 41, 446–452. [Google Scholar] [CrossRef]
- Li, J.W.; Si, S.W.; Cheng, J.Y.; Li, J.X.; Liu, J.Q. Thidiazuron and Silver Nitrate Enhanced Gynogenesis of Unfertilized Ovule Cultures of Cucumis sativus. Biol. Plant. 2013, 57, 164–168. [Google Scholar] [CrossRef]
- Lentini, Z.; González, Á.; Tabares, E.; Buitrago, M.E.; Wêdzony, M. Studies on Gynogenesis Induction in Cassava (Manihot esculenta Crantz) Unpollinated Ovule Culture. Front. Plant Sci. 2020, 11, 365. [Google Scholar] [CrossRef] [Green Version]
- Hause, B.; Van Veenendaal, W.L.H.; Hause, G.; Van Lammeren, A.A.M. Expression of Polarity during Early Development of Microspore-Derived and Zygotic Embryos of Brassica napus L. cv. Topas. Bot. Acta 1994, 107, 407–415. [Google Scholar] [CrossRef]
- Ilic-Grubor, K.; Attree, S.M.; Fowke, L.C. Comparative Morphological Study of Zygotic and Microspore-Derived Embryos of Brassica napus L. as Revealed by Scanning Electron Microscopy. Ann. Bot. 1998, 82, 157–165. [Google Scholar] [CrossRef]
- Seldimirova, O.A.; Kruglova, N.N.; Titova, G.E.; Batygina, T.B. Comparative Ultrastructural Analysis of the in vitro Microspore Embryoids and in vivo Zygotic Embryos of Wheat as a Basis for Understanding of Cytophysiological Aspects of Their Development. Russ. J. Dev. Biol. 2017, 48, 185–197. [Google Scholar] [CrossRef]
- Encheva, J.; Ivanov, P. Sunflower Commercial Hybrids, Developed by Biotechnologically Obtained Lines. In Proceedings of the Congress Book of The 2nd International Balkan Agriculture Congress, Tekirdağ, Turkey, 16–18 May 2017; Namık Kemal Üniversitesi: Tekirdag, Turkey, 2017; pp. 717–726. [Google Scholar]
- Todorova, M.; Ivanov, P.; Shindrova, P.; Christov, M.; Ivanova, I. Doubled Haploid Production of Sunflower (Helianthus annuus L.) Through Irradiated Pollen-Induced Parthenogenesis. Euphytica 1997, 97, 249–254. [Google Scholar] [CrossRef]
- Aktaş, Y.E.; Uncuoğlu, A.A.; Aydi, Y. Induction of Parthenogenetic Haploid Embryos and Plants after Pollination by Irradiated Pollen in Sunflower. In Proceedings of the International, Agricultural, Biological and Life Science Conference, Edirne, Turkey, 2–5 September 2018; pp. 547–551. [Google Scholar]
- Todorova, M.; Ivanov, P. Induced Parthenogenesis in Sunflower: Effect of Pollen Donor. Helia 1999, 22, 49–56. [Google Scholar]
- Drumeva, M.; Berville, A.; Ivanov, P.; Nenova, N.; Encheva, J. Molecular Investigations on the Doubled Haploid Origin of Sunflower Lines (Helianthus annuus L.) Developed Through Gamma-Induced Parthenogenesis. Biotechnol. Biotechnol. Equip. 2005, 19, 46–50. [Google Scholar] [CrossRef]
- Todorova, M.; Ivanov, P.; Nenova, N.; Encheva, J. Effect of Female Genotype on the Efficiency of γ-induced Parthenogenesis in Sunflower (Helianthus annuus). Helia 2004, 27, 67–74. [Google Scholar] [CrossRef]
- Drumeva, M.; Yankov, P. Investigation on the Parthenogenetic Response of Sunflower Lines and Hybrids. Agric. Sci. Technol. 2015, 7, 415–418. [Google Scholar]
- Drumeva, M.; Yankov, P. Parthenogenetic Responsiveness of Sunflower Hybrid Combinations with Expressed Tolerance to Herbicides. Agric. Sci. Technol. 2017, 9, 190–193. [Google Scholar] [CrossRef]
- Todorova, M.; Nenova, N.; Encheva, J. Study on In vitro Pollen Germination of Sunflower (Helianthus annuus L.) Before and After Gamma- irradiation. Bulg. J. Agric. Sci. 2004, 10, 65–70. [Google Scholar]
- Kalinowska, K.; Chamas, S.; Unkel, K.; Demidov, D.; Lermontova, I.; Dresselhaus, T.; Kumlehn, J.; Dunemann, F.; Houben, A. State-of-the-art and Novel Developments of In vivo Haploid Technologies. Theor. Appl. Genet. 2019, 132, 593–605. [Google Scholar] [CrossRef] [Green Version]
- Watts, A.; Kumar, V.; Raipuria, R.K.; Bhattachatarya, R.C. In vivo Haploid Production in Crop Plants: Methods and Challenged. Plant Mol. Biol. Rep. 2018, 36, 685–694. [Google Scholar] [CrossRef]
- Chaudhary, H.K.; Kaila, V.; Rather, S.A.; Tayeng, T. Distant Hybridization and Doubled Haploid Breeding. In Alien Gene Transfer in Crop Plants; Pratap, A., Kumar, J., Eds.; Springer: New York, NY, USA, 2014; Volume 1, pp. 143–164. [Google Scholar]
- Gupta, S.B. Duration of Mitotic Cycle and Regulation of DNA Replication in Nicotiana plumbaginifolia and a hybrid derivative of N. tabacum showing chromosome instability. Can. J. Genet. Cytol. 1969, 11, 133–142. [Google Scholar] [CrossRef]
- Davies, D.R. Chromosome Elimination in Inter-specific Hybrids. Heredity 1974, 32, 267–270. [Google Scholar] [CrossRef] [Green Version]
- Laurie, D.A.; Bennett, M.D. The Timing of Chromosome Elimination in Hexaploid Wheat x Maize Crosses. Genome 1989, 32, 953–961. [Google Scholar] [CrossRef]
- Linde-Laursen, I.; Bothmer, R.V. Aberrant Meiotic Divisions of a Hordeum lechleri x Hordeum vulgare Hybrids. Hereditas 1999, 118, 145–153. [Google Scholar] [CrossRef]
- Jin, W.; Melo, J.R.; Nagaki, K.; Talbert, P.B.; Henikoff, S.; Dawe, R.K.; Jiang, J. Maize Centromeres: Organization and Functional Adaptation in the Genetic Background of Oat. Plant Cell 2004, 16, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Mochida, K.; Tsujimoto, H.; Sasakuma, T. Confocal Analysis of Chromosome Behavior in Wheat x Maize Zygotes. Genome 2004, 47, 199–205. [Google Scholar] [CrossRef]
- Gernand, D.; Rutten, T.; Varshney, A.; Rubtsova, M.; Prodanovic, S.; Brüß, C.; Kumlehn, J.; Matzk, F.; Houben, A. Uniparental Chromosome Elimination at Mitosis and Interphase in Wheat and Pearl millet Crosses Involves Micronucleus Formation, Progressive Heterochromatinization and DNA Fragmentation. Plant Cell 2005, 17, 2431–2438. [Google Scholar] [CrossRef] [Green Version]
- Ishii, T.; Ueda, T.; Tanaka, H.; Tsujimoto, H. Chromosome Elimination by Wide Hybridization Between Triticeae or Oat Plant and Pearl millet: Pearl millet Chromosome Dynamics in Hybrid Embryo Cells. Chromosome Res. 2010, 18, 821–831. [Google Scholar] [CrossRef]
- Surikov, I.M.; Dunaeva, S.E. Elimination of Chromosomes in Distant Hybridization in the Grass Family and Its Use in Production of Haploids. Zh. Obshch. Biol. 1989, 50, 158. [Google Scholar]
- Sanei, M.; Pickering, R.; Kumke, K.; Houben, A. Loss of Centromeric Histone H3 (CENH3) from Centromeres Precedes Uniparental Chromosome Elimination in Interspecific Barley Hybrids. Proc. Natl. Acad. Sci. USA 2011, 108, 498–505. [Google Scholar] [CrossRef] [Green Version]
- Kuppu, S.; Tan, E.H.; Nguyen, H.; Rodgers, A.; Comai, L.; Chan, S.W.; Britt, A.B. Point Mutations in Centromeric Histone Induce Post-zygotic Incompatibility and Uniparental Inheritance. PLoS Genet. 2015, 11, 1005494. [Google Scholar] [CrossRef]
- Jacquier, N.M.; Gilles, L.M.; Pyott, D.E.; Martinant, J.P.; Rogowsky, P.M.; Widiez, T. Puzzling out Plant Reproduction by Haploid Induction for Innovations in Plant Breeding. Nat. Plants 2020, 6, 610–619. [Google Scholar] [CrossRef]
- Hayes, P.; Corey, A.; DeNoma, J. Doubled haploid Production in Barley Using the Hordeum bulbosum (L.) Technique. In Doubled Haploid Production in Crop Plants; Maluszynski, M., Kasha, K.J., Forster, B.P., Szarejko, I., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 5–14. [Google Scholar]
- Patial, M.; Chaudhary, H.K.; Sharma, N.; Sundaresha, S.; Kapoor, R.; Pal, D.; Pramanick, K.K.; Shukla, A.K.; Kumar, J. Effect of Different in vitro and in vivo Variables on the Efficiency of Doubled Haploid Production in Triticum aestivum L. Using Imperata cylindrica- mediated Chromosome Elimination Technique. Cereal Res. Commun. 2021, 49, 133–140. [Google Scholar] [CrossRef]
- Eudes, F.; Chugh, A. An Overview of Triticale Doubled Haploids. In Advances in Haploid Production in Higher Plants; Touraev, A., Forster, B.P., Jain, S.M., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 87–96. [Google Scholar]
- National Sunflower Association. Available online: https://www.sunflowernsa.com/uploads/17/jan_present.futureplansdoubledhaploid.pdf (accessed on 27 July 2022).
- Jambhale, N.D. An Attempt of Hybridization between Carthamus tinctorius and Helianthus annuus Aided by Grafting. Indian J. Genet. 1994, 54, 235–241. [Google Scholar]
- Piosik, L. Haploid Embryos of Lettuce (Lactuca sativa) Induced by Alien Pollen or Chemical Factors. Afr. J. Biotech. 2013, 12, 345–352. [Google Scholar] [CrossRef]
- Piosik, L.; Zenkteler, E.; Zenkteler, M. Development of Haploid Embryos and Plants of Lactuca sativa Induced by Distant Pollination with Helianthus annuus and H. tuberosus. Euphytica 2015, 208, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Dozet, B.; Windsor, A.J. Contemporary Challenges in Sunflower Breeding. In Proceedings of the 19th International Sunflower Conference, Edirne, Turkey, 29 May–3 June 2016; pp. 11–20. [Google Scholar]
- Touchell, D.H.; Palmer, I.E.; Ranney, T.G. In vitro Ploidy Manipulation for Crop Improvement. Front. Plant Sci. 2020, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, B.; Ebrahimzadeh, H. In vitro Androgenesis: Spontaneous vs. Artificial Genome Doubling and Characterization of Regenerants. Plant Cell Rep. 2020, 39, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Srivastava, G.K. Autopolyploids of Helianthus annuus L. var. Morden. Cytologia 2002, 67, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Vardar, F.; Aydin, Y.; Ayturk, O.; Unal, M. The Effects of Colchicine on Growth and Development of Sunflower (Helianthus annuus L.) Seedlings. Fresenius Environ. Bull. 2017, 26, 4113–4118. [Google Scholar]
- Singh, R.N. Chromosomal Abnormalities and Fertility in Induced Autotetraploid Helianthus annuus in C1 and C2 Generation. Cytologia 1992, 57, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Feulgen, R.; Rossenbeck, H. Mikroskopisch-chemischer Nachweis einer Nucleinsäure vom Typus der Thymonucleinsäure und die-darauf beruhende elektive Färbung von Zellkernen in mikroskopischen Präparaten. Hoppe-Seyler’s Z. Physiol. Chem. 1924, 135, 203–252. [Google Scholar] [CrossRef]
- Oleszczuk, S.; Rabiza-Swider, J.; Zimny, J.; Lukaszewski, A.J. Aneuploidy among Androgenic Progeny of Hexaploid Triticale (×Triticosecale Wittmack). Plant Cell Rep. 2011, 30, 575–586. [Google Scholar] [CrossRef]
- Ohnoutková, L.; Vlčko, T. Homozygous Transgenic Barley (Hordeum vulgare L.) Plants by Anther Culture. Plants 2020, 9, 918. [Google Scholar] [CrossRef]
- Doğramacı-Altuntepe, M.; Peterson, T.S.; Jauhar, P.P. Anther Culture-derived Regenerants of Durum Wheat and Their Cytological Characterization. J. Hered. 2001, 92, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, H.; Zhao, Z.; Sheng, X.; Yu, H.; Wang, J. Efficient Doubled Haploid Production in Microspore Culture of Loose-curd Cauliflower (Brassica oleracea var. botrytis). Euphytica 2014, 195, 467–475. [Google Scholar] [CrossRef]
- Kozar, E.V.; Domblides, E.A.; Soldatenko, A.V. Factors Affecting DH Plants in vitro Production from Microspores of European Radish. Vavilovskii Zhurnal Genet. Sel. 2020, 24, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Mineykina, A.; Bondareva, L.; Soldatenko, A.; Domblides, E. Androgenesis of Red Cabbage in Isolated Microspore Culture in Vitro. Plants 2021, 10, 1950. [Google Scholar] [CrossRef] [PubMed]
- Christov, M.; Piskov, A.; Encheva, J.; Valkova, D.; Drumeva, M.; Nenova, N.; Nikolova, V.; Encheva, V.; Shindrova, P.; Petrov, P.; et al. Developing Sunflower Hybrid Cultivars with Increased Productivity, Resistant to Disease and Broomrape Using Classical and Biotechnological Methods. Sci.-Tech. Bull. IOC 2009, 14, 74–87. [Google Scholar]
- Peevska, P.; Drumeva, M.; Georgiev, G. Registration of a New Sunflower Hybrid-Sevar. Agric. Sci. Technol. 2018, 10, 29–33. [Google Scholar] [CrossRef]
- Nenova, N. New Bulgarian Sunflower Hybrid «LINZI». Field Crop Stud. 2018, 11, 97–102. [Google Scholar]





| Genotypes | Explants | Culture Conditions | Results | Reference |
|---|---|---|---|---|
| Eight sunflower cultivars | Unpollinated ovules and young florets | N6 + 0.125 mg/L MCPA + 2–6% sucrose | Ovules and young florets produced 96 and 12 embryos, respectively; produced plants were haploids and diploids | [75] |
| Eight sunflower cultivars | Unpollinated ovaries | For callus production and shoot regeneration: modified MS + 0.25 mg/L 2,4-D + 1 mg/L kinetin + 1 mg/L GA3 + 20 g/L sucrose; for direct embryogenesis: modified MS + 0.5 mg/L BAP + 30 g/L sucrose or modified MS + 1 mg/L IAA + 1 mg/L NAA + 0.5 mg/L kinetin + 1 mg/L BAP + 30 g/L sucrose | All genotypes formed calli (10–85%), 19 diploid plants were regenerated for three cultivars | [37] |
| Eight sunflower genotypes | Unpollinated ovaries | Modified MS + 2 mg/L NAA + 10% sucrose for ovaries; ovules were isolated from ovaries during cultivation and transferred to analogous medium | Embryogenesis frequency of 1.19%, 47% of embryos developed into plants: 50% haploid, 25% diploid, and 25% mixoploid | [76] |
| Not presented | Unpollinated ovules | Modified N6 + 12% sucrose for ovules; modified MS + 1 mg/L NAA + 1–2 mg/L BAP + 0.1 mg/L GA3 + 3–6% sucrose for formed embryos; modified MS + 1–2 mg/L BAP + 0.1 mg/L GA3 + 3–6% sucrose for shoot organogenesis | Up to 23.9% gynogenic embryos | [32] |
| Method (Approach) | Description of Method | Results | References |
|---|---|---|---|
| Direct methods for determining ploidy level | |||
| Chromosome count | Stain root tips or young leaves with acetocarmine or aceto-orcein, according to Feulgen [127], count chromosomes in the metaphase plates | Determination of haploids, diploids, aneuploids, and mixoploids | [9,17,39,42,47,48,76] |
| Stain callus cells with acetocarmine, according to Feulgen | Detection of haploid and diploid cells, suggestive of spontaneous chromosome doubling | [14,42] | |
| Flow cytometry | Isolate and stain nucleus, assess amount of DNA using a flow fluorometer | Data on ploidy level | [17,39,52,91,92,93,96] |
| Indirect methods for determining ploidy level | |||
| Determination of morphological characteristics | Check phenotypic traits including height, anthocyanin coloration, leaf blade length and width, and leaf dentation | Differences between plants produced by anther culture and the donor plant | [9,38] |
| Histological analysis | Conduct histological examination of foci in preparations of cultivated anthers, ovules, and ovaries | Gametic embryo and callus from microspores and egg cells, somatic embryo and callus from anther walls, endothelium and integument tissue | [32,39,77] |
| Use of control plants | Use (1) emasculated plants not pollinated with irradiated pollen, or (2) plants pollinated with irradiated pollen but without embryo rescue | First control without seeds proves the effect of pseudo-pollination; second control with shrunken achene proves the absence of fertilization | [93,95] |
| Methods for confirming plant homozygosity | |||
| Use of isozyme markers | Menadione reductase | Distinction between regenerants and donor plants | [38] |
| Esterase and 6-phosphogluconate dehydrogenase | Confirmation of parthenogenetic origin | [91] | |
| Use of molecular markers | Primers SSL26 and SSL46 | Distinction between donor plants, paternal line, and doubled haploids | [94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blinkov, A.O.; Varlamova, N.V.; Kurenina, L.V.; Khaliluev, M.R. The Production of Helianthus Haploids: A Review of Its Current Status and Future Prospects. Plants 2022, 11, 2919. https://doi.org/10.3390/plants11212919
Blinkov AO, Varlamova NV, Kurenina LV, Khaliluev MR. The Production of Helianthus Haploids: A Review of Its Current Status and Future Prospects. Plants. 2022; 11(21):2919. https://doi.org/10.3390/plants11212919
Chicago/Turabian StyleBlinkov, Andrey O., Nataliya V. Varlamova, Ludmila V. Kurenina, and Marat R. Khaliluev. 2022. "The Production of Helianthus Haploids: A Review of Its Current Status and Future Prospects" Plants 11, no. 21: 2919. https://doi.org/10.3390/plants11212919