
Investigating the Effect of the Interaction of Maize Inducer and Donor Backgrounds on Haploid Induction Rates

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. The Performance of Inducers Belonging to Different Genetic Backgrounds
2.2. The Inducibility of Donors Belonging to Different Genetic Backgrounds
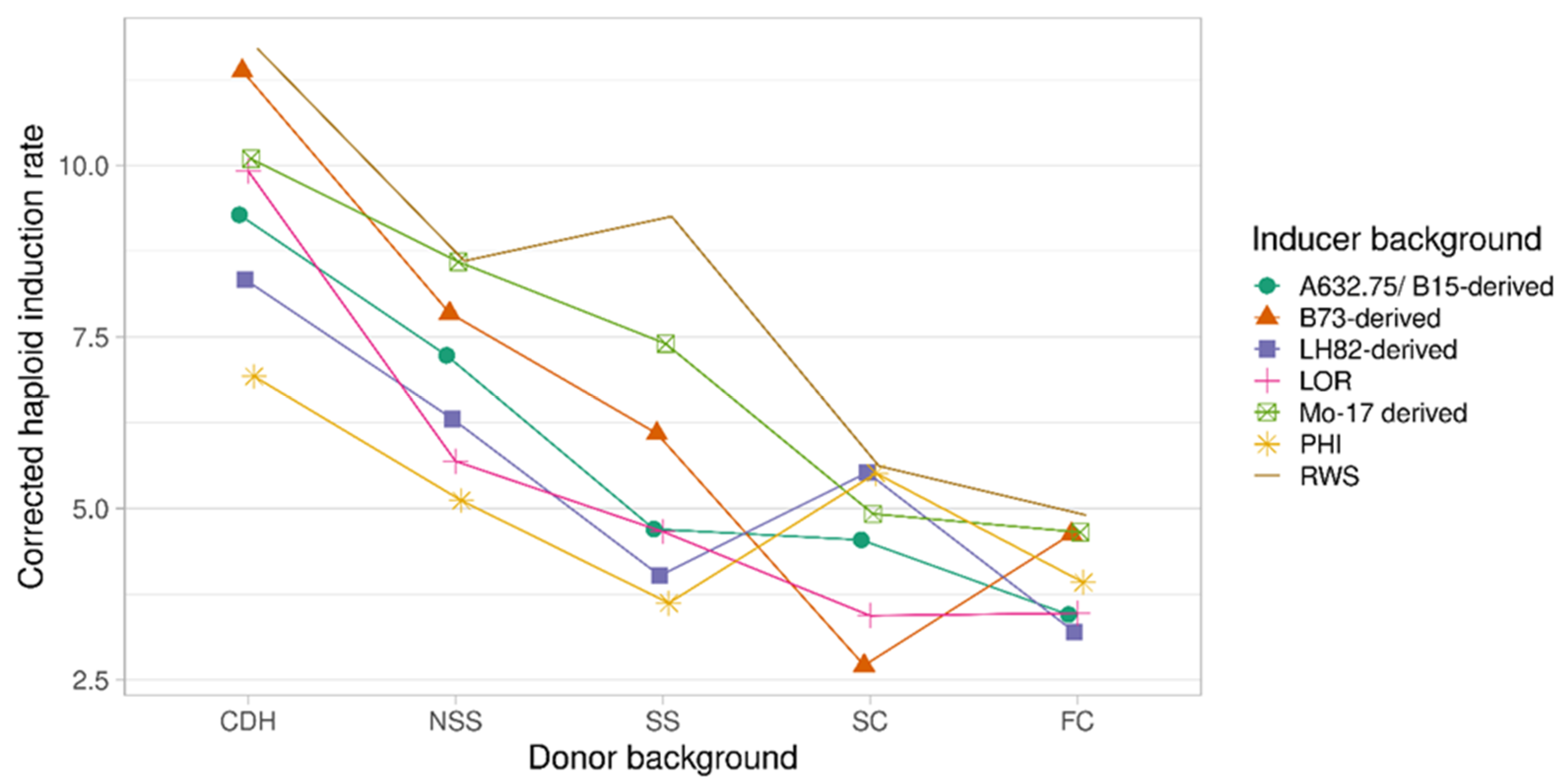
2.3. The Performance of Inducers Belonging to Different Genetic Backgrounds on Specific Donor Backgrounds
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Experimental Design
4.3. Phenotypic Evaluation
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Melchinger, A.E.; Schipprack, W.; Mi, X.; Mirdita, V. Oil content is superior to oil mass for identification of haploid seeds in maize produced with high-oil inducers. Crop Sci. 2015, 55, 188–195. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Carena, M.J.; de Miranda Filho, J.B. Quantitative Genetics in Maize Breeding; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Chaikam, V.; Mahuku, G. (Eds.) Doubled Haploid Technology in Maize Breeding: Theory and Practice; CIMMYT: El Batan, Mexico, 2012; ISBN 978-607-95844-9-8. [Google Scholar]
- Röber, F.K.; Gordillo, G.A.; Geiger, H.H. In vivo haploid induction in maize: Performance of new inducers and significance for doubled haploid lines in hybrid breeding. Maydica 2005, 50, 275–283. [Google Scholar]
- Coe, E.H. A Line of Maize with High Haploid Frequency. Am. Nat. 1959, 93, 381–382. [Google Scholar] [CrossRef]
- Geiger, H.H. Doubled haploids. In Handbook of Maize, Genetics and Genomics; Bennetzen, J.L., Hake, S., Eds.; Springer: New York, NY, USA, 2009; Volume 2, pp. 641–657. [Google Scholar] [CrossRef]
- Germanà, M.A. Anther culture for haploid and doubled haploid production. Plant Cell Tissue Organ Cult. 2010, 104, 283–300. [Google Scholar] [CrossRef]
- Evans, M.M. The indeterminate gametophyte1 gene of maize encodes a LOB domain protein required for embryo Sac and leaf development. Plant Cell 2007, 19, 46–62. [Google Scholar] [CrossRef] [Green Version]
- Kindiger, B.; Hamann, S. Generation of Haploids in Maize: A Modification of the Indeterminate Gametophyte (ig) System. Crop Sci. 1993, 33, 342–344. [Google Scholar] [CrossRef]
- Rotarenco, V.; Dicu, G.; State, D.; Fuia, S. New Inducers of Maternal Haploids in Maize. Maize Genet. Coop. News Lett. 2010, 84, 21–22. [Google Scholar]
- Chaikam, V.; Nair, S.K.; Martinez, L.; Lopez, L.A.; Utz, H.F.; Melchinger, A.E.; Boddupalli, P.M. Marker Assisted Breeding of Improved Maternal Haploid Inducers in Maize for the Tropical/Subtropical Regions. Front. Plant Sci. 2018, 9, 1527. [Google Scholar] [CrossRef] [Green Version]
- Chase, S.S.; Nanda, D.K. Screening for monoploids of maize by use of a purple embryo marker. Maize Genet. Coop. Newsl. 1965, 39, 59–60. [Google Scholar]
- Kebede, A.Z.; Dhillon, B.S.; Schipprack, W.; Jose, L.A.; Bänziger, M.; Semagn, K.; Alvarado, G.; Melchinger, A.E. Effect of source germplasm and season on the in vivo haploid induction rate in tropical maize. Euphytica 2011, 180, 219–226. [Google Scholar] [CrossRef]
- Prigge, V.; Sánchez, C.; Dhillon, B.S.; Schipprack, W.; Araus, J.L.; Bänziger, M.; Melchinger, A.E. Doubled haploids in tropical maize. I. Effects of inducers and source germplasm on in vivo haploid induction rates. Crop Sci. 2011, 51, 1498–1506. [Google Scholar] [CrossRef]
- Prigge, V.; Xu, X.X.; Li, L.; Babu, R.; Chen, S.J.; Atlin, G.N.; Melchinger, A.E. New insights into the genetics of in vivo in-duction of maternal haploids, the backbone of doubled haploid technology in maize. Genetics 2012, 190, 781–793. [Google Scholar] [CrossRef] [Green Version]
- Paz-Ares, J.; Ghosal, D.; Saedler, H. Molecular analysis of the C1-I allele from Zea mays: A dominant mutant of the regulatory C1 locus. EMBO J. 1990, 9, 315–321. [Google Scholar] [CrossRef]
- Burr, F.A.; Burr, B.; Scheffler, B.E.; Blewitt, M.; Wienand, U.; Matz, E.C. The maize repressor-like gene intensifier1 shares homology with the r1/b1 multigene family of transcription factors and exhibits missplicing. Plant Cell 1996, 8, 1249–1259. [Google Scholar] [CrossRef] [Green Version]
- Della Vedova, C.B.; Lorbiecke, R.; Kirsch, H.; Schulte, M.B.; Scheets, K.; Borchert, L.M.; Scheffler, B.E.; Wienand, U.; Cone, K.C.; Birchler, J.A. The Dominant Inhibitory Chalcone Synthase Allele C2-Idf (Inhibitor Diffuse) from Zea mays (L.) Acts via an Endogenous RNA Silencing Mechanism. Genetics 2005, 170, 1989–2002. [Google Scholar] [CrossRef] [Green Version]
- Chaikam, V.; Nair, S.K.; Babu, R.; Martinez, L.; Tejomurtula, J.; Boddupalli, P.M. Analysis of effectiveness of R1-nj anthocyanin marker for in vivo haploid identification in maize and molecular markers for predicting the inhibition of R1-nj expression. Theor. Appl. Genet. 2015, 128, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Birchler, J.A. A Green Fluorescent Protein-Engineered Haploid Inducer Line Facilitates Haploid Mutant Screens and Doubled Haploid Breeding in Maize. Mol. Breed. 2016, 36, 5. [Google Scholar] [CrossRef]
- Coe, E.H. Anthocyanin Genetics. In The Maize Handbook; Freeling, M., Waibot, V., Eds.; Springer: New York, NY, USA, 1994; pp. 279–281. [Google Scholar] [CrossRef]
- Chaikam, V.; Martinez, L.; Melchinger, A.E.; Schipprack, W.; Prasanna, B.M. Development and Validation of Red Root Marker-Based Haploid Inducers in Maize. Crop Sci. 2016, 56, 1678–1688. [Google Scholar] [CrossRef]
- De La Fuente, G.N.; Frei, U.K.; Trampe, B.; Nettleton, D.; Zhang, W.; Lübberstedt, T. A diallel analysis of a maize donor population response to in vivo maternal haploid induction I: Inducibility. Crop Sci. 2018, 58, 1830–1837. [Google Scholar] [CrossRef] [Green Version]
- Seaney, R.R. Monoploids in Maize. Maize Genet. Coop. News Lett. 1954, 28, 22. [Google Scholar]
- Chase, S.S. Monoploids and monoploid-derivatives of maize (Zea mays L.). Bot. Rev. 1969, 35, 117–168. [Google Scholar] [CrossRef]
- Chase, S.S. Utilization of haploids in plant breeding: Breeding diploid species. In Haploids in Higher Plants: Advances and Potential: Proceedings of the First International Symposium; Kasha, K.J., Ed.; Guelph University Press: Guelph, ON, Canada, 1974; pp. 211–230. [Google Scholar]
- Tyrnov, V.S. Producing of parthenogenetic forms of maize. Maize Genet. Coop. News Lett. 1997, 71, 73–74. [Google Scholar]
- Rotarenco, V.A. Production of matroclinous maize haploids following natural and artificial pollination with a haploid inducer. Maize Genet. Coop. News Lett. 2002, 76, 16. [Google Scholar]
- Rotarenco, V.A.; Mihailov, M.E. The influence of ear age on the frequency of maternal haploids produced by a haploid-inducing line. Maize Genet. Coop. News Lett. 2007, 81, 9–10. [Google Scholar]
- Chalyk, S.T.; Baumann, A.; Daniel, G.; Eder, J. Aneuploidy as a possible cause of haploid-induction in maize. Maize Genet. Coop. News Lett. 2003, 77, 29. [Google Scholar]
- Rotarenco, V.; Eder, J. Possible effects of heterofertilization on the induction of maternal haploids in maize. Maize Genet. Coop. News Lett. 2003, 77, 30. [Google Scholar]
- Wu, P.; Li, H.; Ren, J.; Chen, S. Mapping of maternal QTLs for in vivo haploid induction rate in maize (Zea mays L.). Euphytica 2014, 196, 413–421. [Google Scholar] [CrossRef]
- Randolph, L.F. Note on haploid frequencies. Maize Genet. Coop. News Lett. 1940, 14, 51. [Google Scholar]
- Chase, S.S. Production of Homozygous Diploids of Maize from Monoploids. Agron. J. 1952, 44, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Lashermes, P.; Beckert, M. Genetic Control of Maternal Haploidy in Maize (Zea Mays L.) and Selection of Haploid Inducing Lines. Theor. Appl. Genet. 1988, 76, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Eder, J.; Chalyk, S. In vivo haploid induction in maize. Theor. Appl. Genet. 2002, 104, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Geiger, H.H.; Gordillo, G.A. Doubled Haploids in Hybrid Maize Breeding. Maydica 2009, 54, 485–499. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 21 January 2022).


{kind=link}
| Factor | SS | MS | Num DF | Den DF | F Value | Pr (>F) |
|---|---|---|---|---|---|---|
| Year | 439.8 | 146.6 | 3 | 36.4 | 17.92 | |
| Inducer background | 382.3 | 63.7 | 6 | 68.2 | 7.79 | <0.001 |
| Donor background | 375.1 | 93.8 | 4 | 24.0 | 11.46 | <0.001 |
| Inducer background × donor background | 421.4 | 17.6 | 24 | 336.9 | 2.15 | <0.001 |
| Groups | Variance |
|---|---|
| Screener | 11.650 |
| Year × Inducer | 0.265 |
| Year × Donor | 1.777 |
| Year × Donor × Inducer | 3.102 |
| Residual | 8.179 |
| Inducer Background | Mean (%) | SE | Asymp. LCL | Asymp. UCL | Group |
|---|---|---|---|---|---|
| RWS | 8.02 | 0.780 | 6.49 | 9.55 | A |
| Mo17-derived | 7.13 | 0.675 | 5.81 | 8.46 | A |
| B73-derived | 6.53 | 0.727 | 5.11 | 7.96 | AB |
| A632.75/B15-derived | 5.84 | 0.676 | 4.52 | 7.17 | B |
| LH82-derived | 5.48 | 0.676 | 4.15 | 6.80 | B |
| LOR | 5.44 | 0.710 | 4.04 | 6.83 | B |
| PHI | 5.02 | 0.781 | 3.49 | 6.55 | B |
| Donor Background | Mean (%) | SE | Asymp. LCL | Asymp. UCL | Group |
|---|---|---|---|---|---|
| Commercial dent hybrid | 9.66 | 0.923 | 7.85 | 11.47 | A |
| Non-stiff stalk | 7.06 | 0.767 | 5.55 | 8.56 | B |
| Stiff stalk | 5.68 | 0.766 | 4.18 | 7.18 | BC |
| Sweet corn | 4.61 | 0.948 | 2.75 | 6.47 | BC |
| Flint corn | 4.03 | 0.795 | 2.47 | 5.59 | C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uliana Trentin, H.; Batîru, G.; Frei, U.K.; Dutta, S.; Lübberstedt, T. Investigating the Effect of the Interaction of Maize Inducer and Donor Backgrounds on Haploid Induction Rates. Plants 2022, 11, 1527. https://doi.org/10.3390/plants11121527
Uliana Trentin H, Batîru G, Frei UK, Dutta S, Lübberstedt T. Investigating the Effect of the Interaction of Maize Inducer and Donor Backgrounds on Haploid Induction Rates. Plants. 2022; 11(12):1527. https://doi.org/10.3390/plants11121527
Chicago/Turabian StyleUliana Trentin, Henrique, Grigorii Batîru, Ursula Karoline Frei, Somak Dutta, and Thomas Lübberstedt. 2022. "Investigating the Effect of the Interaction of Maize Inducer and Donor Backgrounds on Haploid Induction Rates" Plants 11, no. 12: 1527. https://doi.org/10.3390/plants11121527