
Effect of Salinity and Silicon Doses on Onion Post-Harvest Quality and Shelf Life
 , , and
, , and
Abstract
:1. Introduction
2. Results
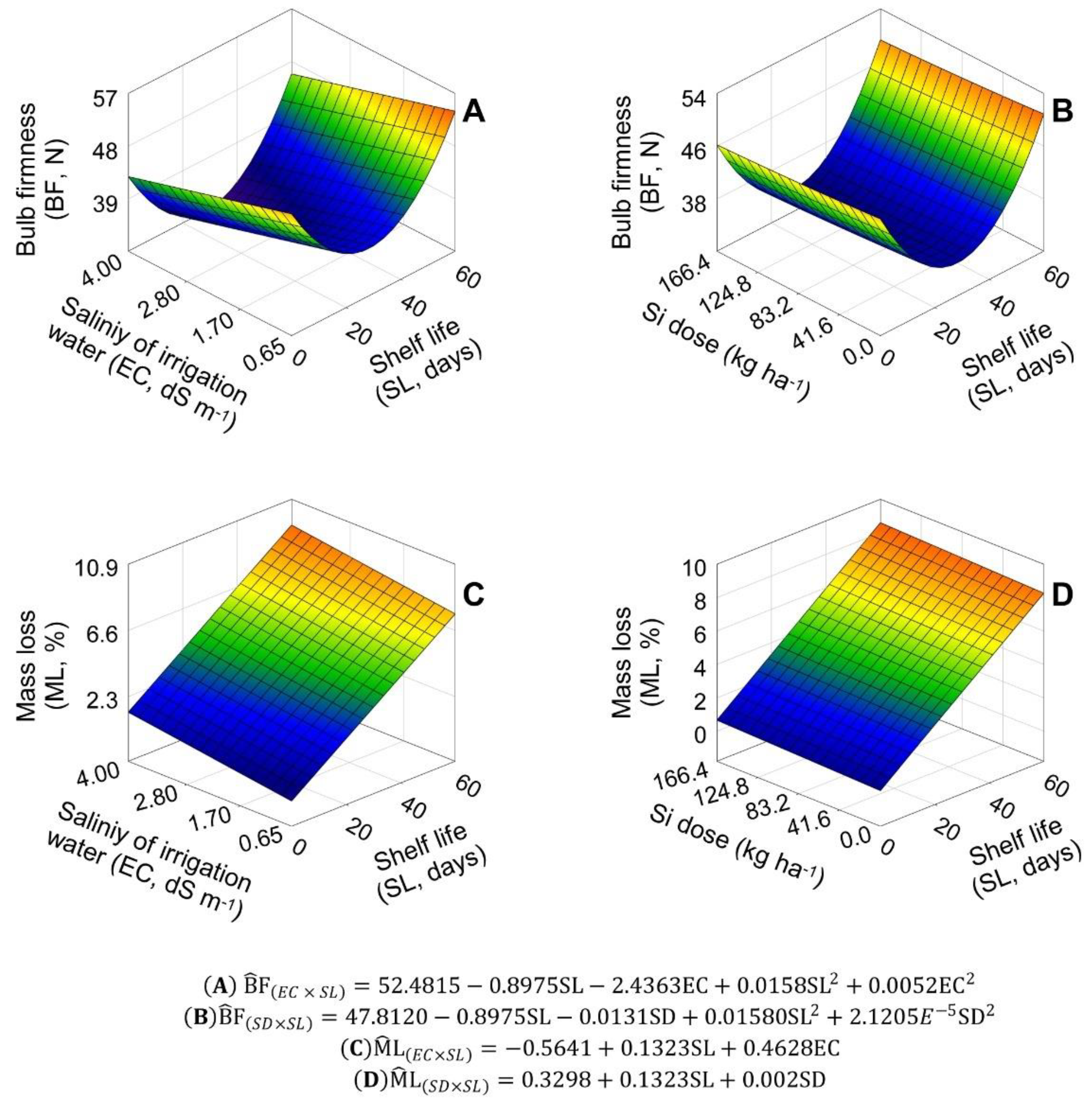
2.1. Bulb Firmness and Mass Loss
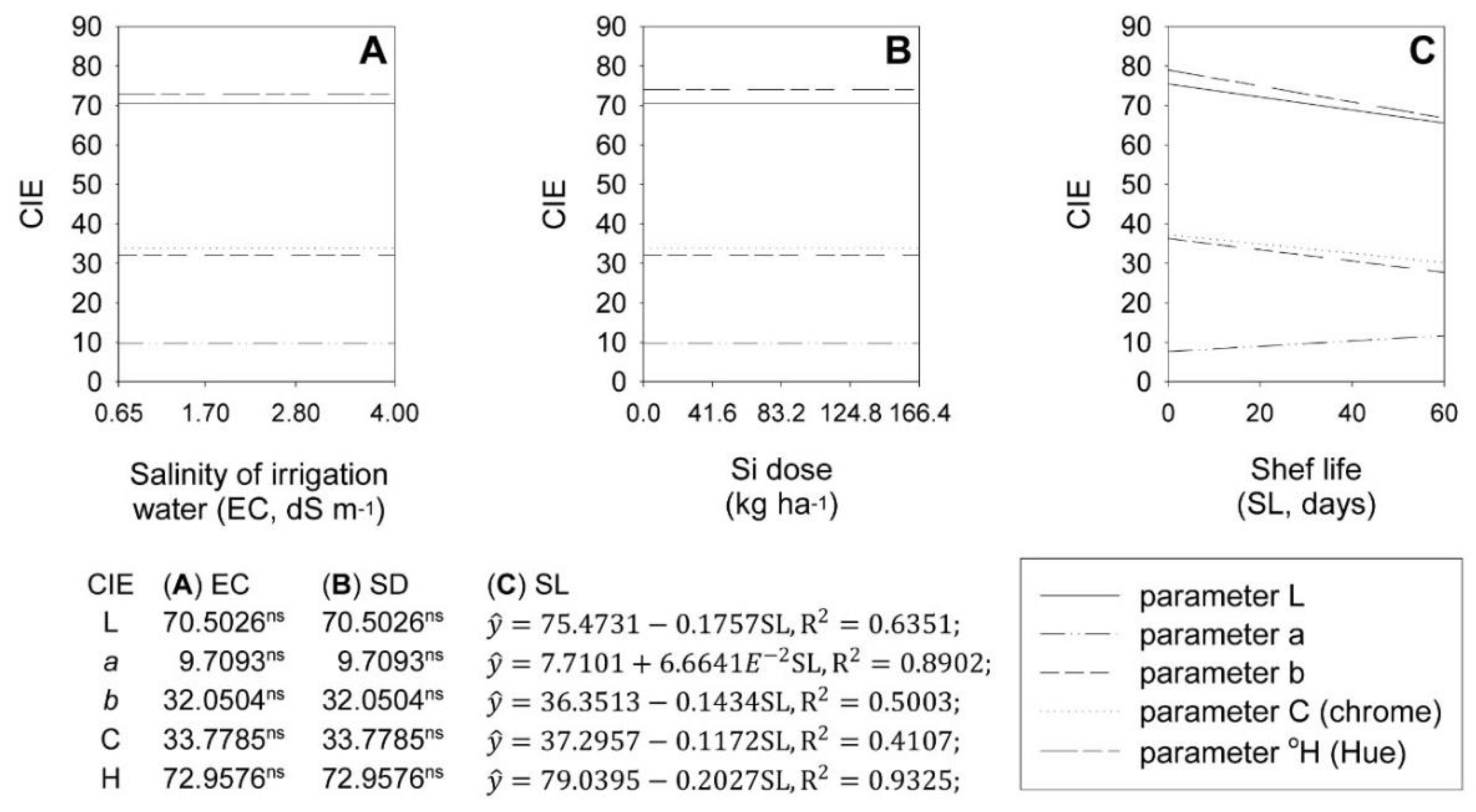
2.2. Tunic Color
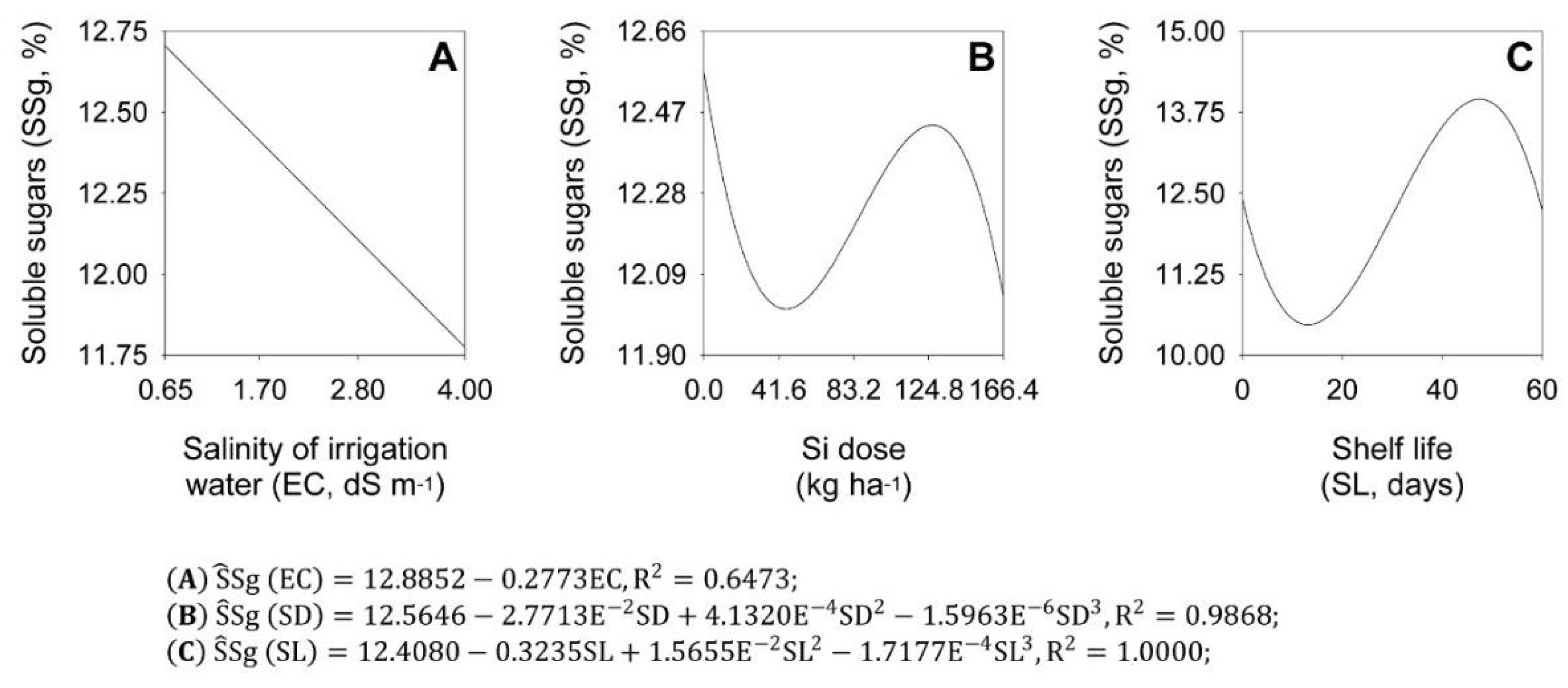
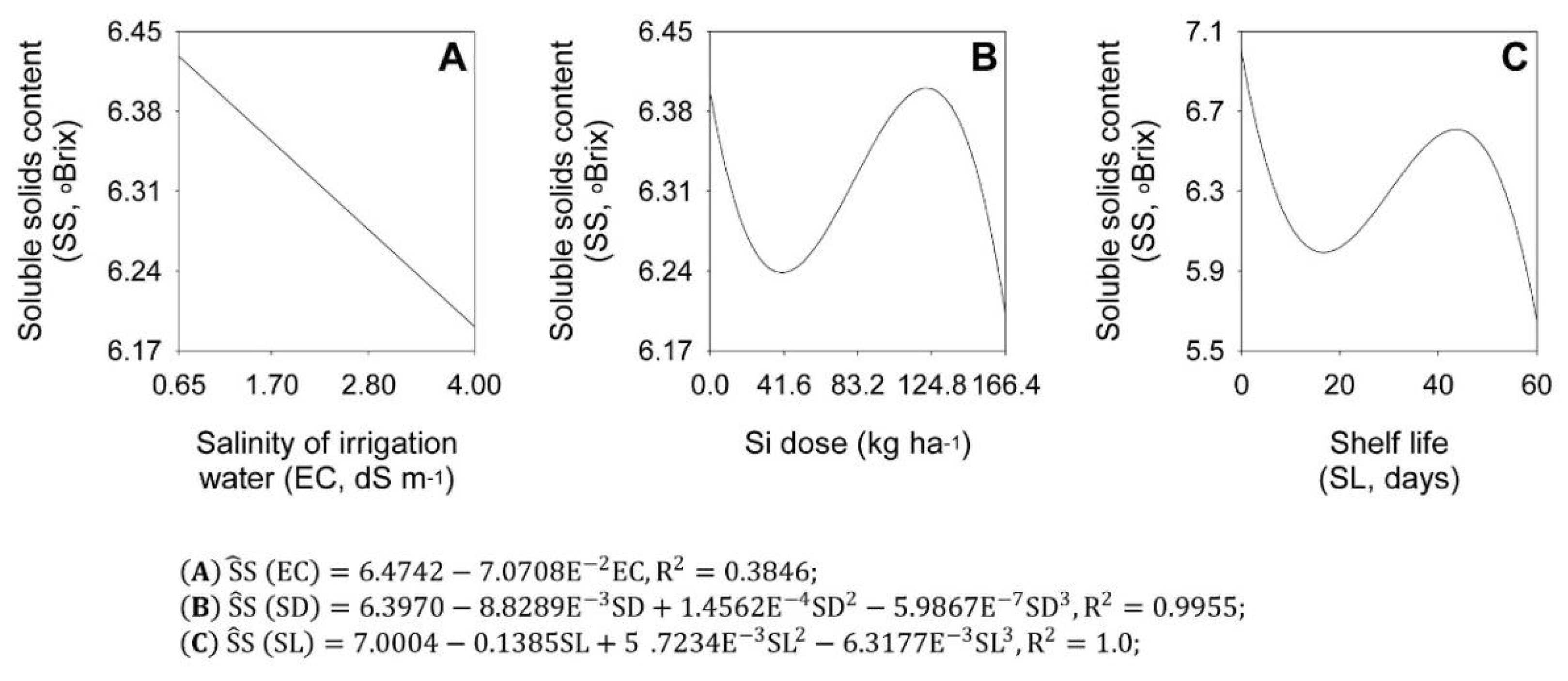
2.3. Soluble Sugars and Total Soluble Solids
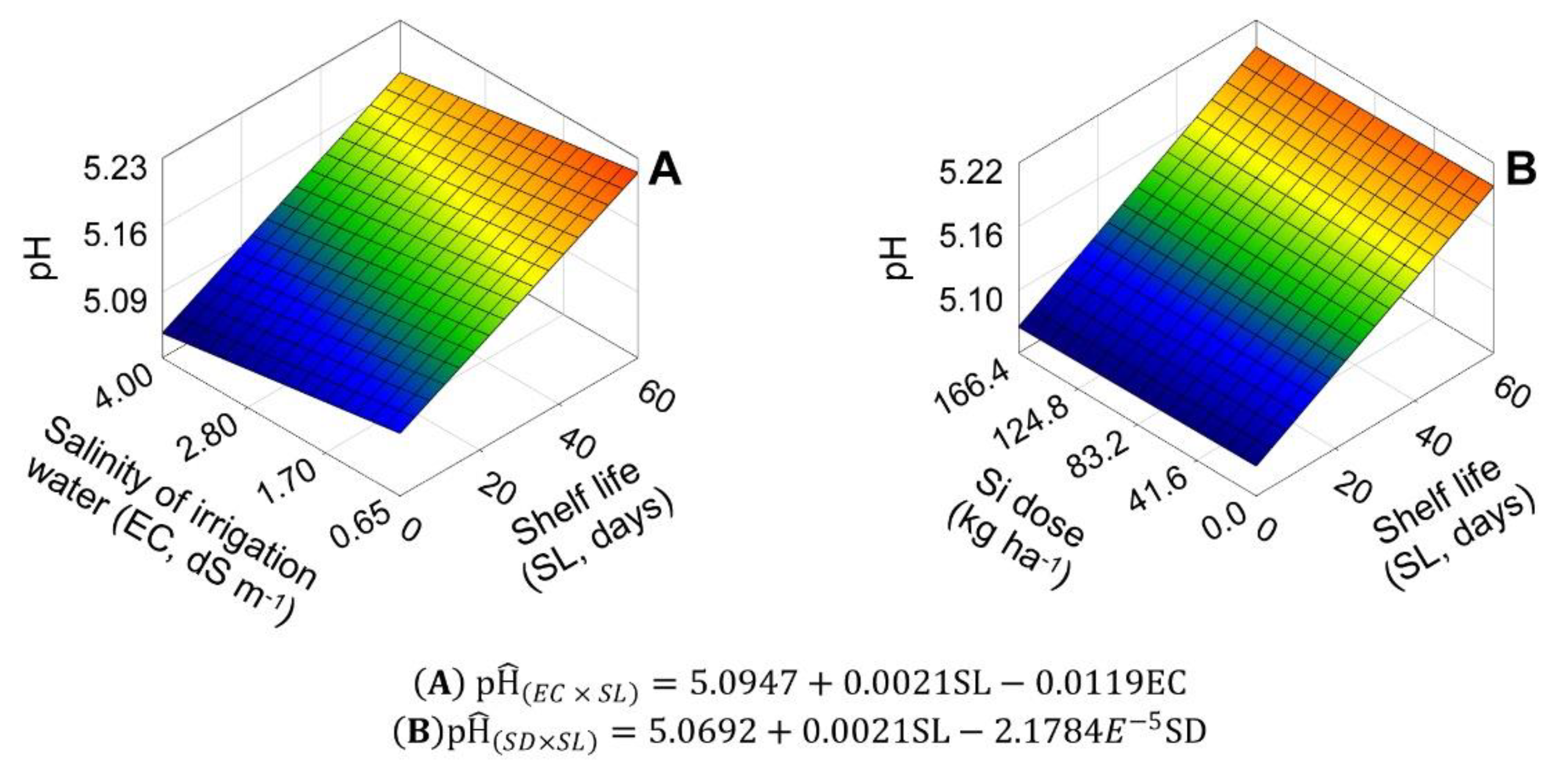
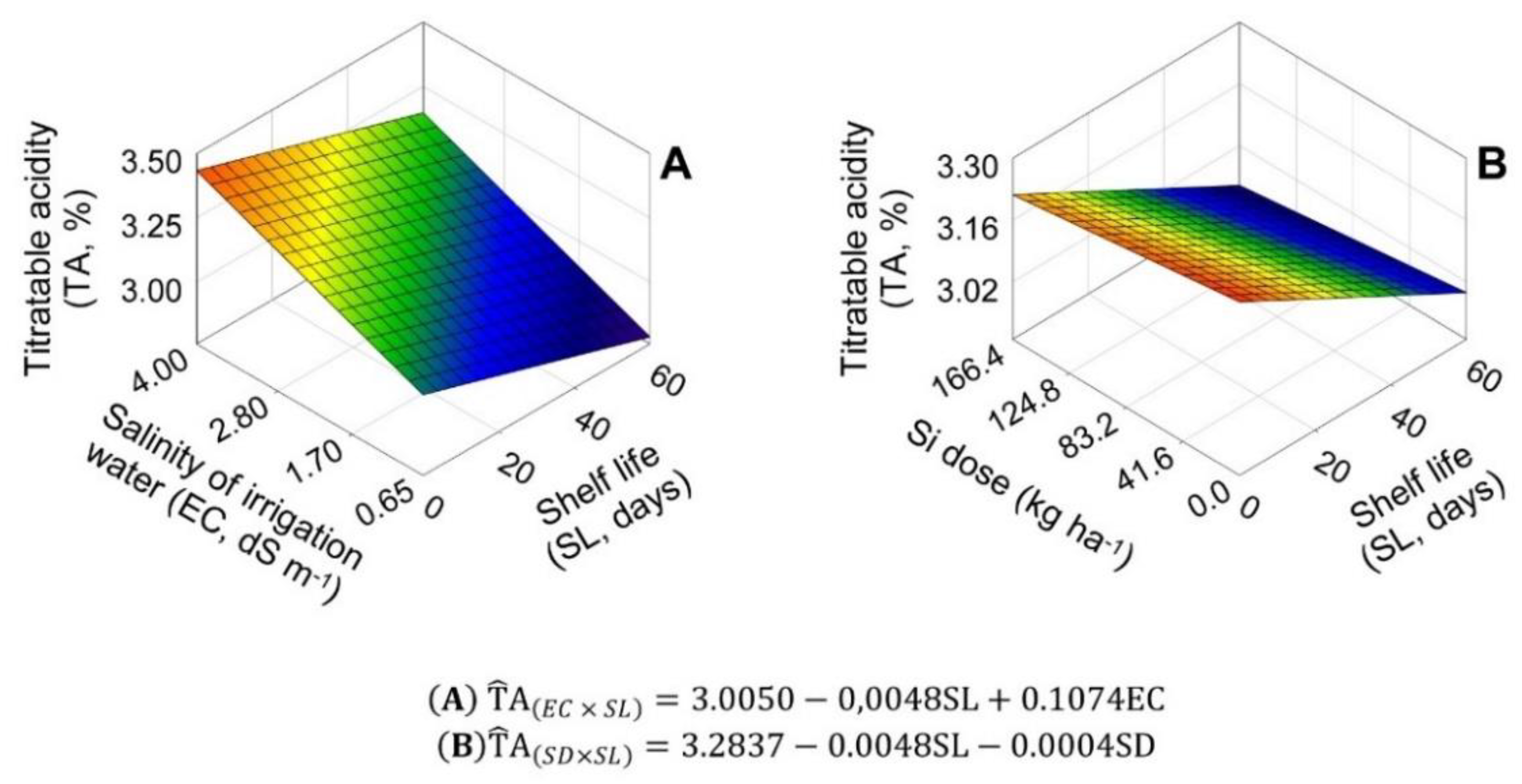
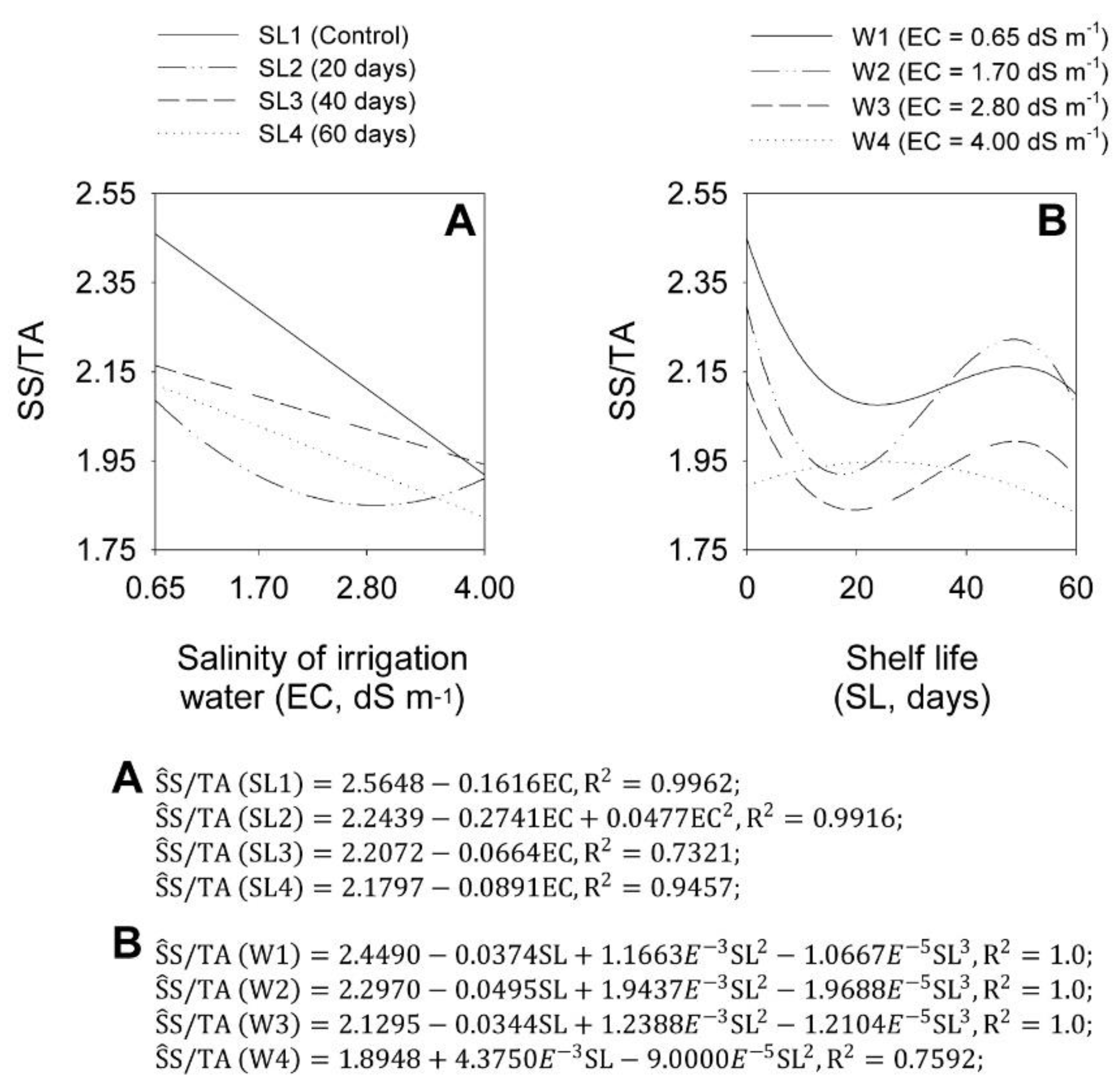
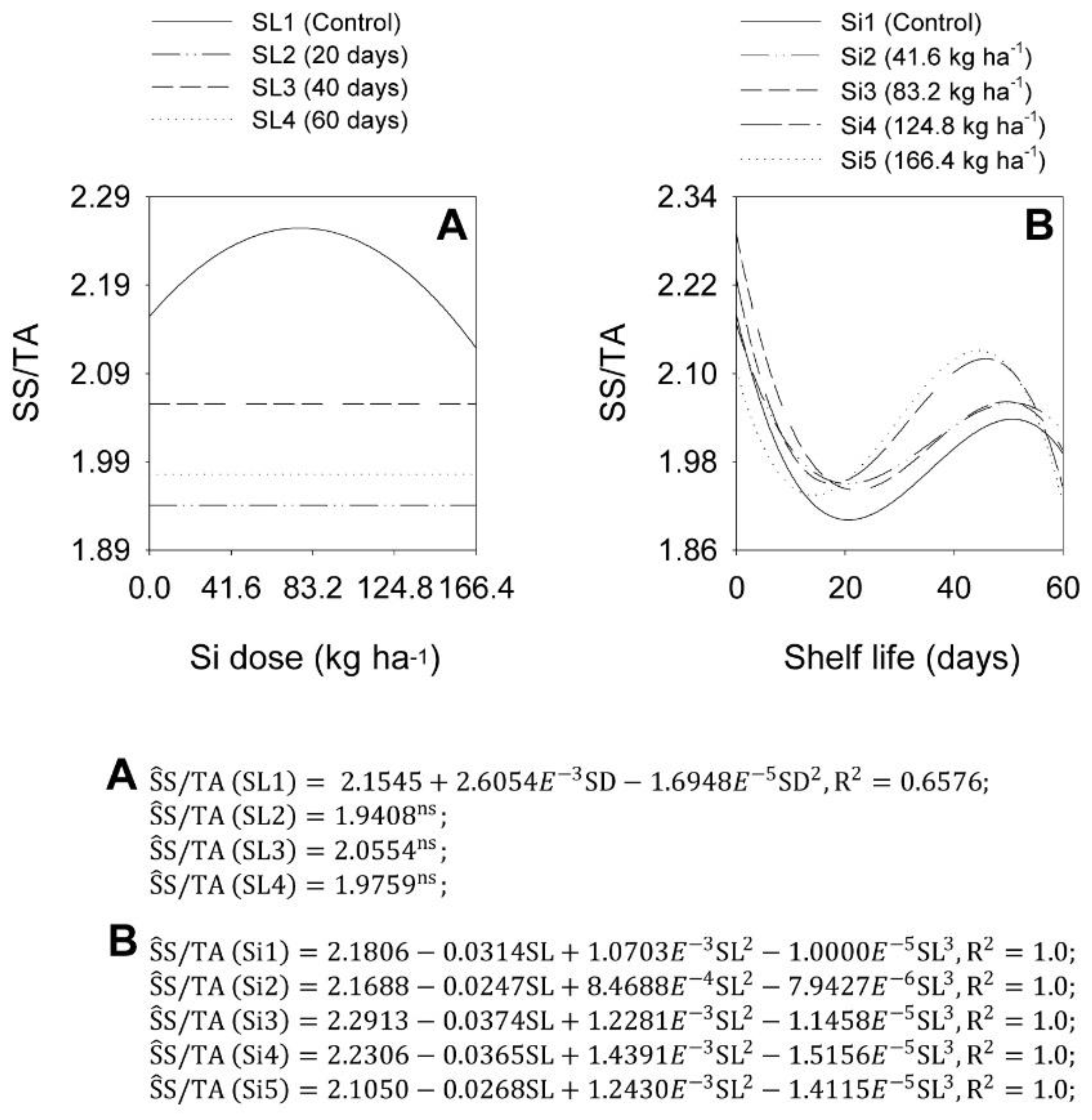
2.4. pH, Titratable Acidity, and SS/TA Ratio in Bulbs
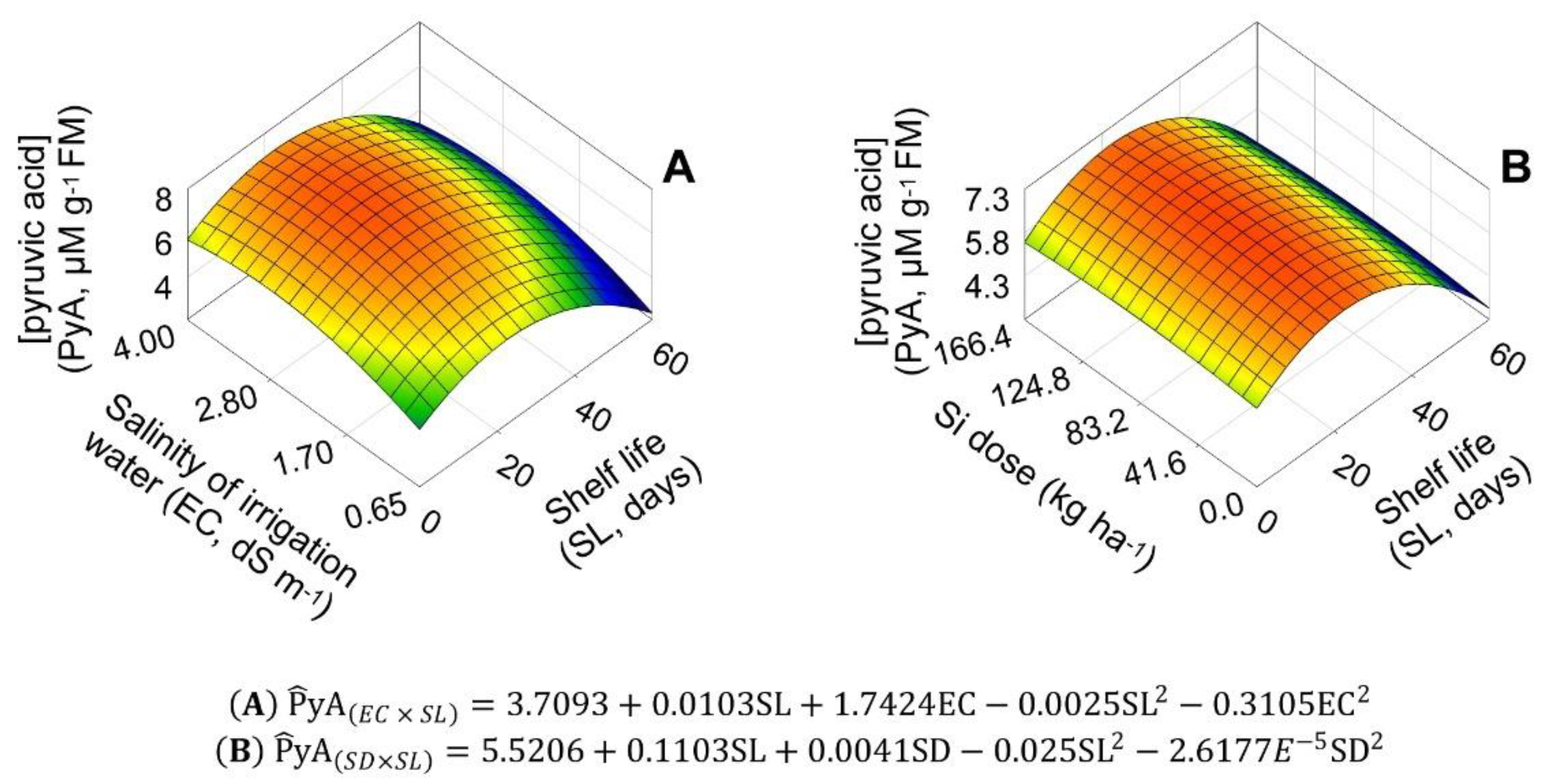
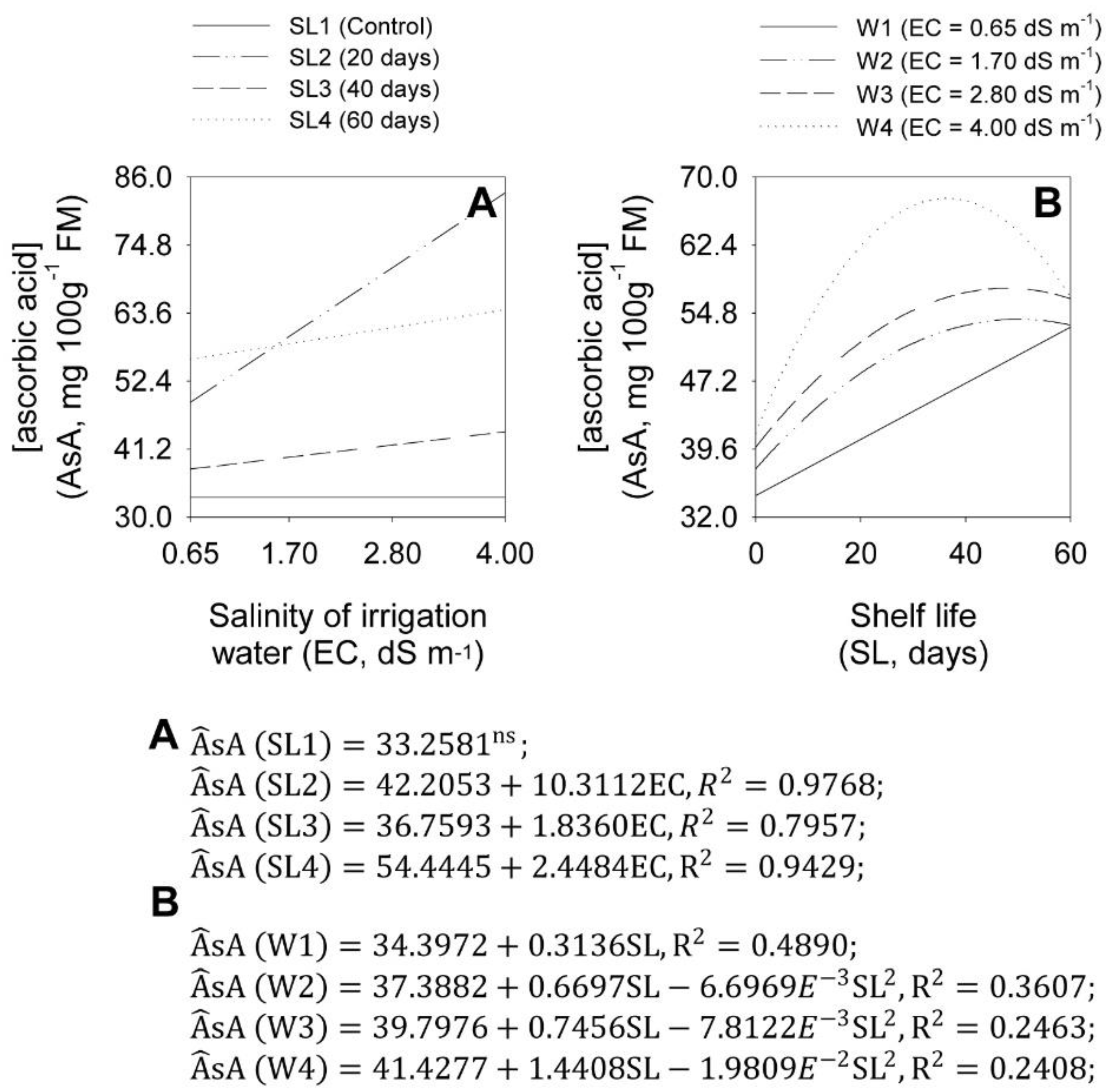
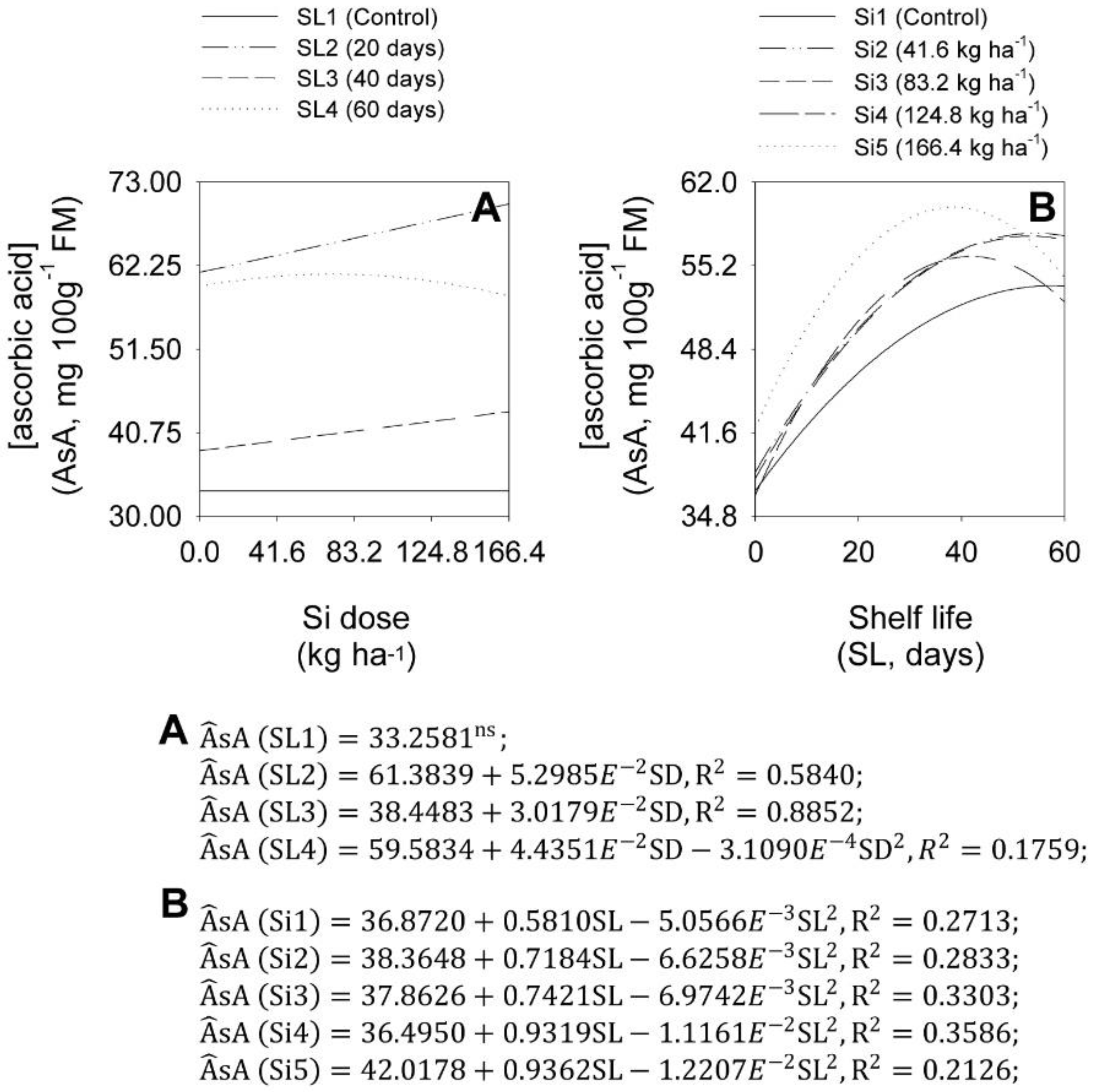
2.5. Concentrations of Pyruvic and Ascorbic Acids
3. Discussion
4. Materials and Methods
4.1. Location and Experimental Design
4.2. Plant Material, Pre-Harvest Treatments, and Post-Harvest Storage
4.3. Evaluated Characteristics
4.3.1. Evaluation of Bulb Firmness and Mass Loss
4.3.2. Evaluation of Bulb Color
4.3.3. Evaluation of Soluble Sugars (SSg), Soluble Solids (SS), Titratable Acidity (TA), SS/TA Ratio, and Hydrogen Potential (pH)
4.3.4. Evaluation of Pyruvic Acid and Ascorbic Acid Concentrations
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rady, M.O.A.; Semida, W.M.; Abd El-Mageed, T.A.; Hemida, K.A.; Rady, M.M. Up-regulation of antioxidative defense systems by glycine betaine foliar application in onion plants confer tolerance to salinity stress. Sci. Hortic. 2018, 240, 614–622. [Google Scholar] [CrossRef]
- Ricciardi, L.; Mazzeo, R.; Marcotrigiano, A.R.; Rainaldi, G.; Iovieno, P.; Zonno, V.; Pavan, S.; Lotti, C. Assessment of Genetic Diversity of the “Acquaviva Red Onion” (Allium cepa L.) Apulian Landrace. Plants 2020, 9, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohanenye, I.C.; Alamar, M.C.; Thompson, A.J.; Terry, L.A. Fructans redistribution prior to sprouting in stored onion bulbs is a potential marker for dormancy break. Postharvest Biol. Technol. 2019, 149, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Sharma, S.R.; Dhall, R.K.; Mittal, T.C. Effect of γ-radiation on post-harvest storage life and quality of onion bulb under ambient condition. J. Food Sci. Technol. 2020, 57, 2534–2544. [Google Scholar] [CrossRef]
- Sharma, K.; Ko, E.Y.; Assefa, A.D.; Nile, S.H.; Park, S.W. A comparative study of anaerobic and aerobic decomposition of quercetin glucosides and sugars in onion at an ambient temperature. Front. Life Sci. 2015, 8, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Mendonza, C.; Vega-García, M.O.; Guevara-Aguilar, A.; Sánchez, E.; Alvarado-González, M.; Flores-Córdova, M. Effect of prolonged storage in controlled atmospheres on the conservation of the onion (Allium cepa L.) quality. Emir. J. Food Agric. 2016, 28, 842–852. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.N.; Wang, A.; Pedersen, J.S.; Edelenbos, M. Microclimate Tools to Monitor Quality Changes in Stored Onions. Acta Hortic. 2017, 1154, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Dhumal, K.; Datir, S.; Pandey, R. Assessment of bulb pungency level in different Indian cultivars of onion (Allium cepa L.). Food Chem. 2007, 100, 1328–1330. [Google Scholar] [CrossRef]
- Chitarra, M.I.F.; Chitarra, A.B. Pós-Colheita de Frutas e Hortaliças: Fisiologia e Manuseio, 2nd ed.; UFLA: Lavras, Brazil, 2005. [Google Scholar]
- Valverde-Miranda, D.; Díaz-Pérez, M.; Gómez-Galán, M.; Callejón-Ferre, Á.J. Total soluble solids and dry matter of cucumber as indicators of shelf life. Postharvest Biol. Technol. 2021, 180, 111603. [Google Scholar] [CrossRef]
- Hasegawa, P.M. Sodium (Na+) homeostasis and salt tolerance of plants. Environ. Exp. Bot. 2013, 92, 19–31. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of sodium chloride toxicity in Solanum lycopersicum L. by supplementation of jasmonic acid and nitric oxide. J. Plant Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Tahara, M.; Yamada, Y.; Mitsudomi, Y.; Koga, K. Characterization of the polyamine biosynthetic pathways and salt stress response in Brachypodium distachyon. J. Plant Growth Regul. 2018, 37, 625–634. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Qin, H.; Li, Z.; Liu, H.; Wang, J.; Zhang, H.; Quan, R.; Huang, R.; Zhang, Z. The synthesis of ascorbic acid in rice roots plays an important role in the salt tolerance of rice by scavenging ROS. Int. J. Mol. Sci. 2018, 19, 3347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wu, F.; Li, Y.; Qian, Y.; Pan, X.; Li, F.; Wang, Y.; Wu, Z.; Fu, C.; Lin, H.; et al. NtMYB4 and NtCHS1 are critical factors in the regulation of flavonoid biosynthesis and are involved in salinity responsiveness. Front. Plant Sci. 2019, 10, 178. [Google Scholar] [CrossRef]
- Venâncio, J.B.; Dias, N.S.; Medeiros, J.F.; Morais, P.L.D.; Nascimento, C.W.A.; Sousa Neto, O.N.; Sá, F.V.S. Yield and Morphophysiology of onion grown under salinity and fertilization with silicon. Sci. Hortic. 2022, 301, 111095. [Google Scholar] [CrossRef]
- Amal, A.; Aly, A. Alteration of some secondary metabolites and enzymes activity by using exogenous antioxidant compound in onion plants growth under seawater salt stress. Am.-Eurasian J. Sci. Res. 2008, 3, 139–146. [Google Scholar]
- Hanci, F.; Cebeci, E.; Uysal, E.; Dasgan, H. Effects of salt stress on some physiological parameters and mineral element contents of onion (Allium cepa L.) plants. Acta Hortic. 2016, 1143, 179–186. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef]
- Bybordi, A.; Saadat, S.; Zargaripour, P. The effect of zeolite, selenium and silicon on qualitative and quantitative traits of onion grown under salinity conditions. Arch. Agron. Soil Sci. 2018, 64, 520–530. [Google Scholar] [CrossRef]
- Coolong, T.W.; Randle, W.M.; Wicker, L. Structural and chemical differences in the cell wall regions in relation to scale firmness of three onion (Allium cepa L.) selections at harvest and during storage. J. Sci. Food Agric. 2008, 88, 1277–1286. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Rady, M.O.A.; Marey, R.A.; Abd El-Mageed, T.A. Exogenously applied proline enhances growth and productivity of drought stressed onion by improving photosynthetic efficiency, water use efficiency and up-regulating osmoprotectants. Sci. Hortic. 2020, 272, 109580. [Google Scholar] [CrossRef]
- Safar, A.; Wyatt, H.L.; Mihai, L.A. Debonding of cellular structures under shear deformation. In Proceedings of the 25th Conference of the UK Association for Computational Mechanics, University of Birmingham, Birmingham, UK, 11–13 April 2017. [Google Scholar]
- Ng, A.; Parker, M.L.; Parr, A.J.; Saunders, P.K.; Smith, A.C.; Waldron, K.W. Physicochemical Characteristics of Onion (Allium cepa L.) Tissues. J. Agric. Food Chem. 2000, 48, 5612–5617. [Google Scholar] [CrossRef] [PubMed]
- Maw, B.W.; Mullinix, B.G. Moisture loss of sweet onions during curing. Postharvest Biol. Technol. 2005, 35, 223–227. [Google Scholar] [CrossRef]
- Eshel, D.; Teper-Bamnolker, P.; Vinokur, Y.; Saad, I.; Zutahy, Y.; Rodov, V. Fast curing: A method to improve postharvest quality of onions in hot climate harvest. Postharvest Biol. Technol. 2014, 88, 34–39. [Google Scholar] [CrossRef]
- Díaz-Pérez, J.C. Transpiration. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Sawston, UK, 2018; pp. 157–173. [Google Scholar] [CrossRef]
- Madani, B.; Mirshekari, A.; Imahori, Y. Physiological Responses to Stress. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 405–423. [Google Scholar] [CrossRef]
- Galindo, F.G.; Herppich, W.; Gekas, V.; Sjöholm, I. Factors Affecting Quality and Postharvest Properties of Vegetables: Integration of Water Relations and Metabolism. Crit. Rev. Food Sci. Nutr. 2004, 44, 139–154. [Google Scholar] [CrossRef]
- Fleck, A.T.; Schulze, S.; Hinrichs, M.; Specht, A.; Waßmann, F.; Schreiber, L.; Schenk, M.K. Silicon Promotes Exodermal Casparian Band Formation in Si-Accumulating and Si-Excluding Species by Forming Phenol Complexes. PLoS ONE 2015, 10, e0138555. [Google Scholar] [CrossRef]
- Kumar, S.; Soukup, M.; Elbaum, R. Silicification in grasses: Variation between different cell types. Front. Plant Sci. 2017, 8, 438. [Google Scholar] [CrossRef]
- Ferrón-Carrillo, F.; Urrestarazu, M. Effects of Si in nutrient solution on leaf cuticles. Sci. Hortic. 2021, 278, 109863. [Google Scholar] [CrossRef]
- Ko, E.Y.; Nile, S.H.; Sharma, K.; Li, G.H.; Park, S.W. Effect of different exposed lights on quercetin and quercetin glucoside content in onion (Allium cepa L.). Saudi J. Biol. Sci. 2015, 22, 398–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahama, U. Oxidation of vacuolar and apoplastic phenolic substrates by peroxidase: Physiological significance of the oxidation reactions. Phytochem. Rev. 2004, 3, 207–219. [Google Scholar] [CrossRef]
- Downes, K.; Chope, G.A.; Terry, L.A. Effect of curing at different temperatures on biochemical composition of onion (Allium cepa L.) skin from three freshly cured and cold stored UK-grown onion cultivars. Postharvest Biol. Technol. 2009, 54, 80–86. [Google Scholar] [CrossRef]
- Hu, M.; Shi, Z.; Zhang, Z.; Zhang, Y.; Li, H. Effects of exogenous glucose on seed germination and antioxidant capacity in wheat seedlings under salt stress. Plant Growth Regul. 2012, 68, 177–188. [Google Scholar] [CrossRef]
- Obata, T.; Geigenberger, P.; Fernie, A.R. Redox control of plant energy metabolism: The complex intertwined regulation of redox and metabolism in plant cells. Biochemist 2015, 37, 14–18. [Google Scholar] [CrossRef]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 17037. [Google Scholar] [CrossRef] [Green Version]
- Khanna-Chopra, R.; Semwal, V.K.; Lakra, N.; Pareek, A. 5 proline—A key regulator conferring plant tolerance to salinity and drought. In Plant Tolerance to Environmental Stress: Role of Phytoprotectants; Hasanuzzaman, M., Fujita, M., Oku, H., Islam, T., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2019; pp. 59–80. [Google Scholar]
- Sami, F.; Yusuf, M.; Faizan, M.; Faraz, A.; Hayat, S. Role of sugars under abiotic stress. Plant Physiol. Biochem. 2016, 109, 54–61. [Google Scholar] [CrossRef]
- Yahia, E.M.; Carrillo-López, A.; Bello-Perez, L.A. Chapter 9–Carbohydrates. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 175–205. [Google Scholar] [CrossRef]
- Foyer, C.H.; Ruban, A.V.; Noctor, G. Viewing oxidative stress through the lens of oxidative signaling rather than damage. Biochem. J. 2017, 474, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Pei, Z.F.; Ming, D.F.; Liu, D.; Wan, G.L.; Geng, X.X.; Gong, H.J.; Zhou, W.J. Silicon Improves the Tolerance to Water-Deficit Stress Induced by Polyethylene Glycol in Wheat (Triticum aestivum L.) Seedlings. J. Plant Growth Regul. 2010, 29, 106–115. [Google Scholar] [CrossRef]
- Yin, L.; Wang, S.; Li, J.; Tanaka, K.; Oka, M. Application of silicon improves salt tolerance through ameliorating osmotic and ionic stresses in the seedling of Sorghum bicolor. Acta Physiol. Plant. 2013, 35, 3099–3107. [Google Scholar] [CrossRef]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.-X.; Xu, X.-B.; Hu, Y.-H.; Han, W.-H.; Yin, J.-L.; Li, H.-L.; Gong, H.-J. Silicon improves salt tolerance by increasing root water uptake in Cucumis sativus L. Plant Cell Rep. 2015, 34, 1629–1646. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-X.; Gong, H.-J.; Yin, J.-L. Role of Silicon in Mediating Salt Tolerance in Plants: A Review. Plants 2019, 8, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltveit, M.E. Chapter 4—Respiratory Metabolism. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 73–91. [Google Scholar] [CrossRef]
- Coca, C.A.; Carranza, C.E.; Miranda, D.; Rodríguez, M.H. Efecto del NaCl sobre los parámetros de crecimiento, rendimiento y calidad de la cebolla de bulbo (Allium cepa L.) bajo condiciones controladas (NaCl effects on growth, yield and quality parameters in the onion (Allium cepa L.) under controlled conditions). Rev. Colomb. Ciências Hortícolas 2012, 6, 196–212. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.-T.; Randle, W.M. Sodium Chloride Timing and Length of Exposure Affect Onion Growth and Flavor. J. Plant Nutr. 2005, 28, 1755–1766. [Google Scholar] [CrossRef]
- Chope, G.A.; Terry, L.A.; White, P.J. Effect of controlled atmosphere storage on abscisic acid concentration and other biochemical attributes of onion bulbs. Postharvest Biol. Technol. 2006, 39, 233–242. [Google Scholar] [CrossRef]
- Sohany, M.; Sarker, M.K.U.; Mahmud, M.S. Physiological changes in red onion bulbs at different storage temperature. World J. Eng. Technol. 2016, 4, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Vallarino, J.G.; Osorio, S. Chapter 10—Organic Acids. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 207–224. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Eprintsev, A.T. Organic Acids: The Pools of Fixed Carbon Involved in Redox Regulation and Energy Balance in Higher Plants. Front. Plant Sci. 2016, 7, 1042. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U.; Bykova, N.V. Role of organic acids in integrating cellular redox metabolism and mediation of redox signalling in photosynthetic tissues of higher plants. Free Radic. Biol. Med. 2018, 122, 74–85. [Google Scholar] [CrossRef]
- Khan, N.A.; Khan, M.I.R.; Asgher, M.; Fatma, M.; Masood, A.; Syeed, S. Salinity tolerance in plants: Revisiting the role of sulfur metabolites. J. Plant Biochem. Physiol. 2014, 2, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Schwarzenberg, A.; Yvin, J.-C.; Hosseini, S.A. Regulatory Role of Silicon in Mediating Differential Stress Tolerance Responses in Two Contrasting Tomato Genotypes Under Osmotic Stress. Front. Plant Sci. 2018, 9, 1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maillard, A.; Ali, N.; Schwarzenberg, A.; Jamois, F.; Yvin, J.-C.; Hosseini, S.A. Silicon transcriptionally regulates sulfur and ABA metabolism and delays leaf senescence in barley under combined sulfur deficiency and osmotic stress. Environ. Exp. Bot. 2018, 155, 394–410. [Google Scholar] [CrossRef]
- Réthoré, E.; Ali, N.; Yvin, J.-C.; Hosseini, S.A. Silicon Regulates Source to Sink Metabolic Homeostasis and Promotes Growth of Rice Plants under Sulfur Deficiency. Int. J. Mol. Sci. 2020, 21, 3677. [Google Scholar] [CrossRef] [PubMed]
- Do, H.; Kim, I.-S.; Jeon, B.W.; Lee, C.W.; Park, A.K.; Wi, A.R.; Shin, S.C.; Park, H.; Kim, Y.-S.; Yoon, H.-S.; et al. Structural understanding of the recycling of oxidized ascorbate by dehydroascorbate reductase (OsDHAR) from Oryza sativa L. japonica. Sci. Rep. 2016, 6, 19498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Sehrawat, A.; Deswal, R. Asada-Halliwell pathway maintains redox status in Dioscorea alata tuber which helps in germination. Plant Sci. 2016, 250, 20–29. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic acid—A potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef]
- Cao, S.; Du, X.H.; Li, L.H.; Liu, Y.D.; Zhang, L.; Pan, X.; Li, Y.; Li, H.; Lu, H. Overexpression of Populus tomentosa cytosolic ascorbate peroxidase enhances abiotic stress tolerance in tobacco plants. Russ. J. Plant Physiol. 2017, 64, 224–234. [Google Scholar] [CrossRef]
- Sharma, S.; Kiran, U.; Sopory, S.K. Developing stress-tolerant plants by manipulating components involved in oxidative stress. In Plant Biotechnology: Principles and Applications; Abdin, M.Z., Kiran, U., Kamaluddin, A.A., Eds.; Springer: Singapore, 2017; pp. 233–248. [Google Scholar] [CrossRef]
- Yoshimura, K.; Ishikawa, T. Chemistry and metabolism of ascorbic acid in plants. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Hossain, M.A., Munné-Bosch, S., Burritt, D.J., Diaz-Vivancos, P., Fujita, M., Lorence, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–23. [Google Scholar] [CrossRef]
- Ishikawa, T.; Maruta, T.; Yoshimura, K.; Smirnoff, N. Biosynthesis and regulation of ascorbic acid in plants. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 163–179. [Google Scholar] [CrossRef]
- Eskling, M.; Arvidsson, P.-O.; Åkerlund, H.-E. The xanthophyll cycle, its regulation and components. Physiol. Plant. 1997, 100, 806–816. [Google Scholar] [CrossRef]
- Giorgia, S.; Giorgetti, A.; Fufezan, C.; Giacometti, G.M.; Bassi, R.; Morosinotto, T. Mutation analysis of violaxanthin de-epoxidase identifies substrate-binding sites and residues involved in catalysis. J. Biol. Chem. 2010, 285, 23763–23770. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-Y.; Lin, S.-T.; Ko, T.-P.; Wu, S.-M.; Lin, T.-H.; Chang, Y.-C.; Huang, K.-F.; Lee, T.-M. Enzymatic characterization and crystal structure analysis of Chlamydomonas reinhardtii dehydroascorbate reductase and their implications for oxidative stress. Plant Physiol. Biochem. 2017, 120, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radic. Biol. Med. 2018, 122, 116–129. [Google Scholar] [CrossRef] [PubMed]
- García-Caparrós, P.; Hasanuzzaman, M.; Lao, M.T. Oxidative Stress and Antioxidant Defense in Plants under Salinity. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2019; pp. 291–309. [Google Scholar] [CrossRef]
- Saqib, M.; Zörb, C.; Schubert, S. Silicon-mediated improvement in the salt resistance of wheat (Triticum aestivum) results from increased sodium exclusion and resistance to oxidative stress. Fun. Plant Biol. 2008, 35, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Al-Huqail, A.A.; Alqarawi, A.A.; Hashem, A.; Malik, J.A.; Abd_Allah, E.F. Silicon supplementation modulates antioxidant system and osmolyte accumulation to balance salt stress in Acacia gerrardii Benth. Saudi J. Biol. Sci. 2019, 26, 1856–1864. [Google Scholar] [CrossRef]
- Farooq, A.; Bukhari, S.A.; Akram, N.A.; Ashraf, M.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Exogenously Applied Ascorbic Acid-Mediated Changes in Osmoprotection and Oxidative Defense System Enhanced Water Stress Tolerance in Different Cultivars of Safflower (Carthamus tinctorious L.). Plants 2020, 9, 104. [Google Scholar] [CrossRef] [Green Version]
- Carmo Filho, F.; Oliveira, O.F. Mossoró: Um Município do Semi-Árido Nordestino, Caracterização Climática e Aspecto Florístico; ESAM: Mossoró, Brazil, 1995; p. 62. [Google Scholar]
- Chauhan, C.P.S.; Shisodia, P.K.; Minhas, P.S.; Chauhan, R.S. Response of onion (Allium cepa) and garlic (Allium sativum) to irrigation with different salinity waters with or without mitigating salinity stress at seedling establishment stage. Indian J. Agric. Sci. 2007, 77, 483–485. [Google Scholar]
- Aguiar Neto, P.; Grangeiro, L.C.; Mendes, A.M.S.; Costa, N.D.; Marrocos, S.T.P.; Sousa, V.F.L. Growth and accumulation of macronutrients in onion crop in Baraúna (RN) and Petrolina (PE). Rev. Bras. Eng. Agrícola E Ambient. 2014, 18, 370–380. [Google Scholar] [CrossRef]
- Behairy, A.G.; Mahmoud, A.R.; Shafeek, M.; Ali, A.H.; Hafez, M.M. Growth, yield and bulb quality of onion plants (Allium cepa L.) as affected by foliar and soil application of potassium. Middle East J. Agric. Res. 2015, 4, 60–66. [Google Scholar]
- Marrocos, S.T.; Grangeiro, L.C.; Sousa, V.F.L.; Ribeiro, R.M.P.; Cordeiro, C.J. Potassium fertilization for optimization of onion production. Rev. Caatinga 2018, 31, 379–384. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Barros, L.; Ferreira, I.C.F.R.; Ntatsi, G. Morphological, nutritional and chemical description of “Vatikiotiko”, an onion local landrace from Greece. Food Chem. 2015, 182, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Instituto Adolfo Lutz. Métodos Físico-Químicos Para Análise de Alimentos, 4th ed.; Instituto Adolfo Lutz: São Paulo, Brasil, 2008.
- Schwimmer, S.; Weston, W.J. Onion Flavor and Odor, Enzymatic Development of Pyruvic Acid in Onion as a Measure of Pungency. J. Agric. Food Chem. 1961, 9, 301–304. [Google Scholar] [CrossRef]
- Strohecker, R.; Zaragoza, F.M.; Henning, H.M. Analisis de Vitaminas: Métodos Comprovados; Editorial Paz Montalvo: Madrid, Spain, 1967. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Ca2+ | Mg2+ | K+ | Na+ | Al3+ | H++Al3+ | P | M.O. | Si | Clay | Silt | Sand |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ───────────cmolcdm−3─────────── | mg dm−3 | g kg−1 | mg dm−3 | --------g kg−1-------- | ||||||||
| 7.30 | 1.60 | 0.41 | 0.14 | 0.07 | 0.00 | 0.44 | 31.08 | 4.65 | 0.10 | 29 | 21 | 950 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venâncio, J.B.; Dias, N.d.S.; Medeiros, J.F.d.; Morais, P.L.D.d.; Nascimento, C.W.A.d.; Sousa Neto, O.N.d.; Andrade, L.M.d.; Pereira, K.T.O.; Peixoto, T.D.C.; Rocha, J.L.A.; et al. Effect of Salinity and Silicon Doses on Onion Post-Harvest Quality and Shelf Life. Plants 2022, 11, 2788. https://doi.org/10.3390/plants11202788
Venâncio JB, Dias NdS, Medeiros JFd, Morais PLDd, Nascimento CWAd, Sousa Neto ONd, Andrade LMd, Pereira KTO, Peixoto TDC, Rocha JLA, et al. Effect of Salinity and Silicon Doses on Onion Post-Harvest Quality and Shelf Life. Plants. 2022; 11(20):2788. https://doi.org/10.3390/plants11202788
Chicago/Turabian StyleVenâncio, Jefferson Bittencourt, Nildo da Silva Dias, José Francismar de Medeiros, Patrícia Lígia Dantas de Morais, Clístenes Williams Araújo do Nascimento, Osvaldo Nogueira de Sousa Neto, Luciara Maria de Andrade, Kleane Targino Oliveira Pereira, Tayd Dayvison Custódio Peixoto, Josinaldo Lopes Araújo Rocha, and et al. 2022. "Effect of Salinity and Silicon Doses on Onion Post-Harvest Quality and Shelf Life" Plants 11, no. 20: 2788. https://doi.org/10.3390/plants11202788