
Insights from a Multi-Omics Integration (MOI) Study in Oil Palm (Elaeis guineensis Jacq.) Response to Abiotic Stresses: Part Two—Drought
 , ,
, , 
Abstract
:1. Introduction
2. Results
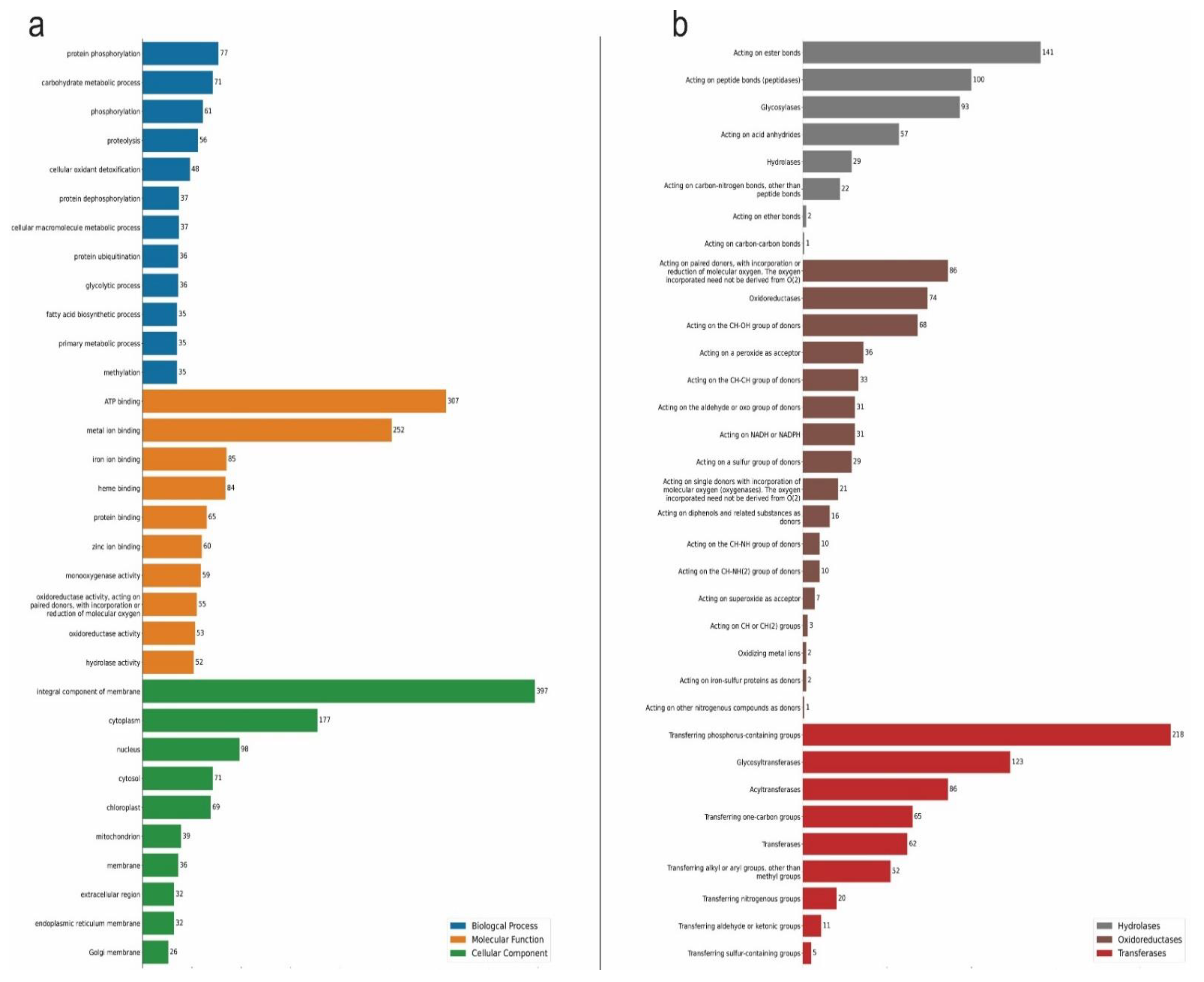
2.1. Oil Palm Transcriptome under Drought Stress
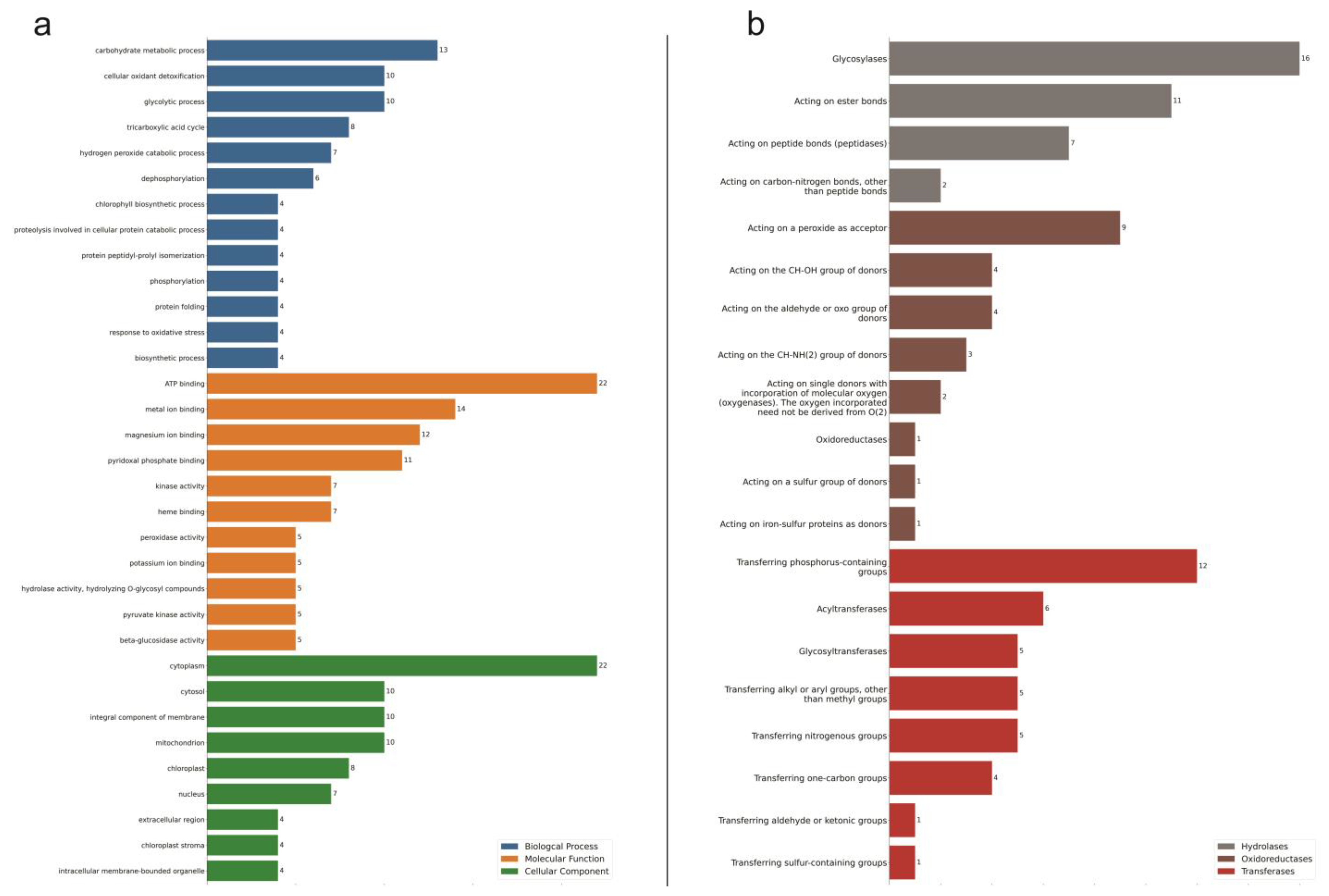
2.2. Oil Palm Proteome under Drought Stress
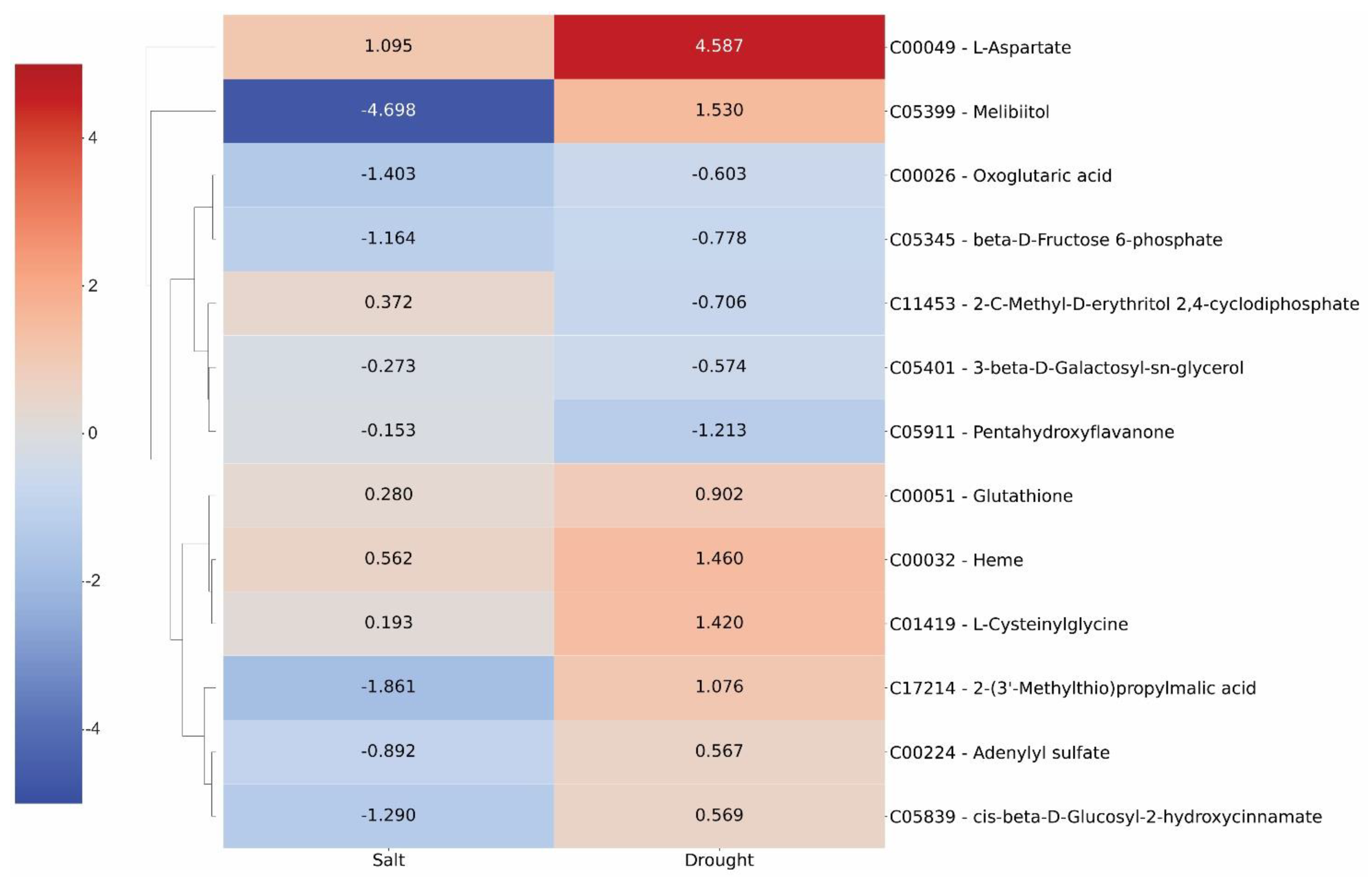
2.3. Oil Palm Metabolome under Drought Stress
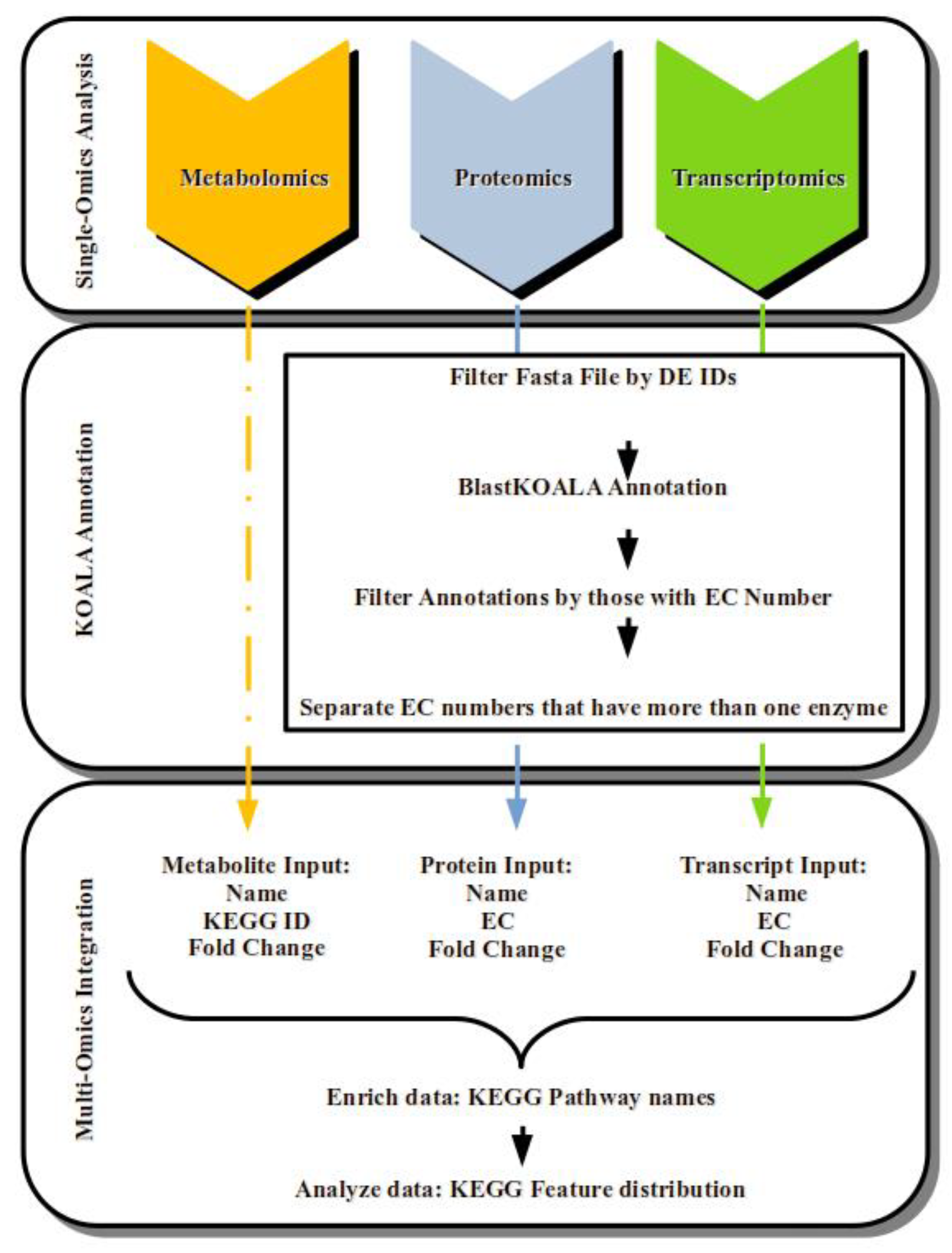
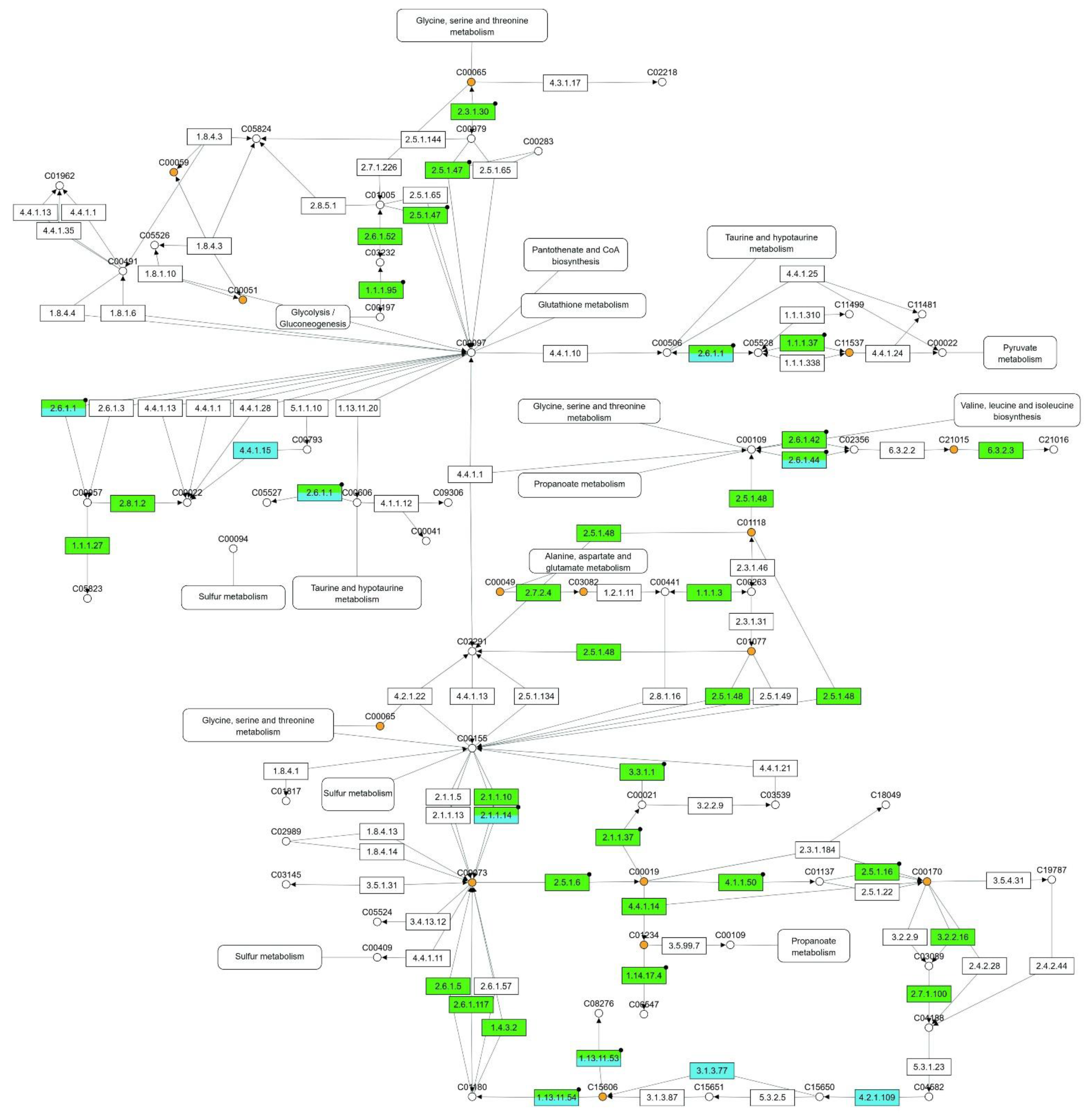
2.4. Integrating Oil Palm Transcriptome, Proteome and Metabolome under Drought Stress
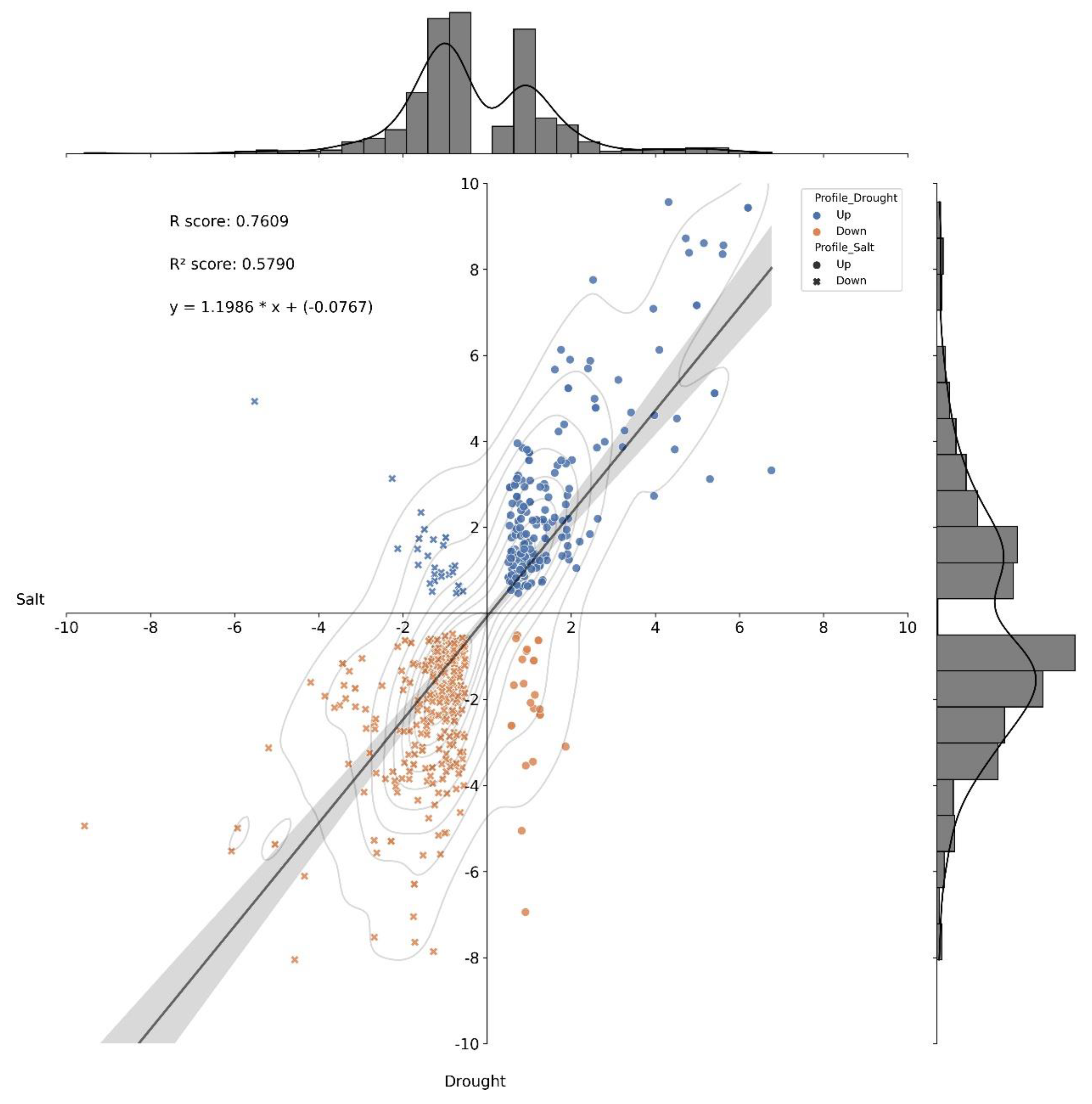
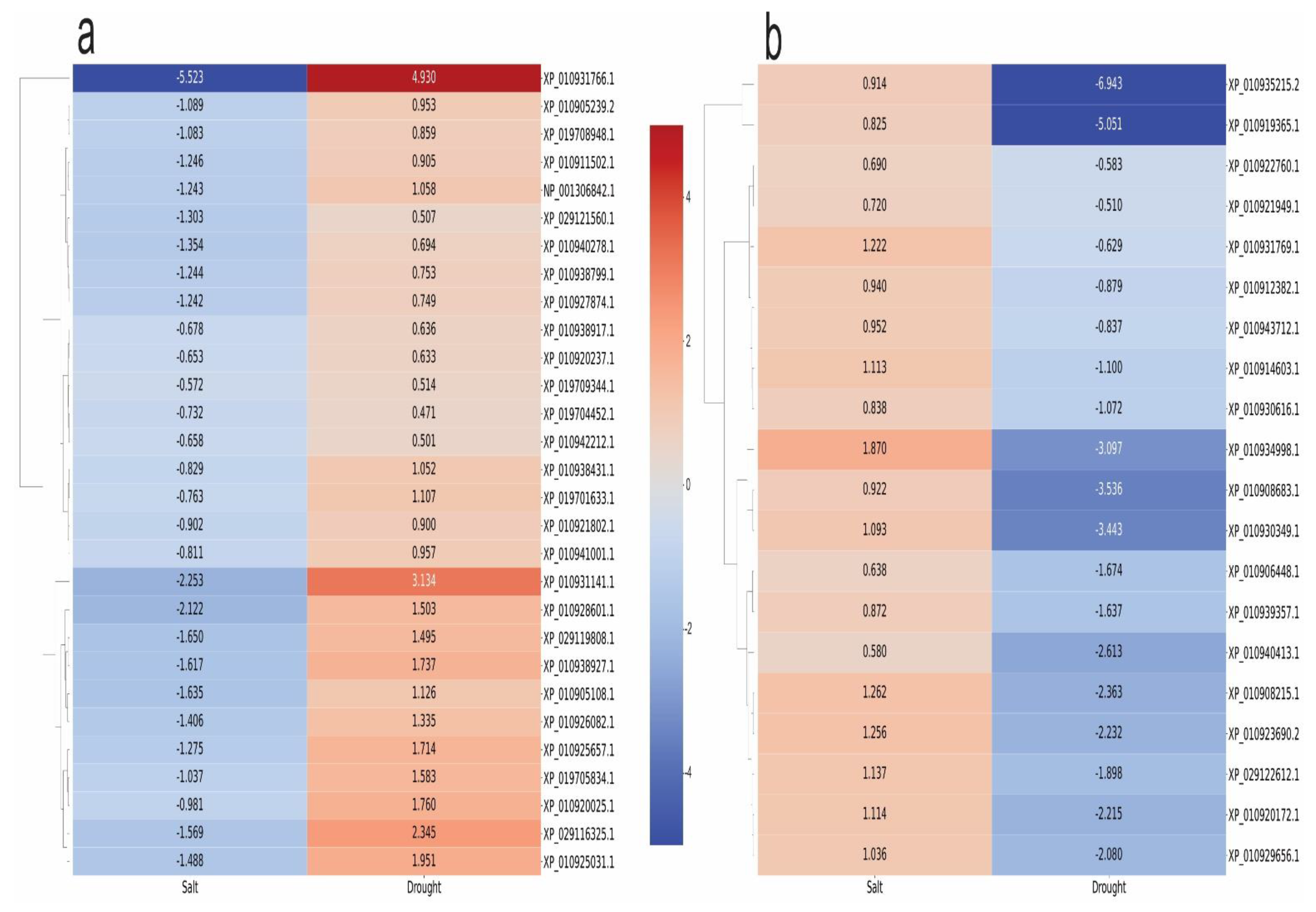
2.5. Commonalities and Dissimilarity in the Transcriptome, Proteome, and Metabolome Profiles of Young Oil Palm Plants Separately Submitted to Drought and Salinity Stresses
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions, Experimental Design, and Drought Stress
4.2. Single-Omics Data Analysis, Functional Annotation, and Integratomics Analysis
4.3. Correlation Analysis of Differentially Expressed Molecules under Two Distinct Scenarios—Salinity and Drought Stresses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- UNCCD. UN Convention to Combat Desertification. Drought in Numbers 2022-Restoration to Readiness and Resilience. 2022. Available online: www.unccd.int/sites/default/files/2022-06/Drought%20in%20Numbers%20%28English%29.pdf (accessed on 31 August 2022).
- Yang, X.; Cushman, J.C.; Borland, A.M.; Liu, Q. Editorial: Systems Biology and Synthetic Biology in Relation to Drought Tolerance or Avoidance in Plants. Front. Plant Sci. 2020, 11, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinwari, Z.K.; Jan, S.A.; Nakashima, K.; Yamaguchi-Shinozaki, K. Genetic engineering approaches to understanding drought tolerance in plants. Plant Biotechnol. Rep. 2020, 14, 151–162. [Google Scholar] [CrossRef]
- Cavill, R.; Jennen, D.; Kleinjans, J.; Briedé, J.J. Transcriptomic and Metabolomic Data Integration. Brief. Bioinform. 2016, 17, 891–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamil, I.N.; Remali, J.; Azizan, K.A.; Nor Muhammad, N.A.; Arita, M.; Goh, H.-H.; Aizat, W.M. Systematic Multi-Omics Integration (MOI) Approach in Plant Systems Biology. Front. Plant Sci. 2020, 11, 944. [Google Scholar] [CrossRef] [PubMed]
- Scossa, F.; Alseekh, S.; Fernie, A.R. Integrating multi-omics data for crop improvement. J. Plant Physiol. 2021, 257, 153352. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Ueta, R.; Osakabe, Y. Precision genome editing in plants: State-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 2020, 20, 234. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.N.; Hossain, M.A.; Burritt, D.J. Salinity and drought stress: Similarities and differences in oxidative responses and cellular redox regulation. In Water Stress and Crop Plants: A Sustainable Approach; Ahmad, P., Ed.; Wiley: Chichester, UK, 2016; Volume 1, pp. 86–101. [Google Scholar]
- Forni, C.; Duca, D.; Glick, B.R. Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 2017, 410, 335–356. [Google Scholar] [CrossRef]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef]
- Brini, F.; Hanin, M.; Mezghani, I.; Berkowitz, G.A.; Masmoudi, K. Overexpression of wheat Na+/H+ antiporter TNHX1 and H+-pyrophosphatase TVP1 improve salt- and drought-stress tolerance in Arabidopsis thaliana plants. J. Exp. Bot. 2007, 58, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, Z.; Li, X.; Gao, X.; Dai, Z.; Cui, Y.; Zhi, Y.; Liu, Q.; Zhai, H.; Gao, S.; et al. The IbBBX24-IbTOE3-IbPRX17 module enhances abiotic stress tolerance by scavenging reactive oxygen species in sweet potato. New Phytol. 2022, 233, 1133–1152. [Google Scholar] [CrossRef]
- Feng, X.H.; Zhang, H.X.; Ali, M.; Gai, W.X.; Cheng, G.X.; Yu, Q.H.; Yang, S.B.; Li, X.X.; Gong, Z.H. A small heat shock protein CaHsp25.9 positively regulates heat, salt, and drought stress tolerance in pepper (Capsicum annuum L.). Plant Physiol. Biochem. 2019, 142, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.C.; Bohra, A.; Nayyar, H. Advances in “omics” approaches to tackle drought stress in grain legumes. Plant Breed. 2020, 139, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Zargar, S.M.; Mir, R.A.; Ebinezer, L.B.; Masi, A.; Hami, A.; Manzoor, M.; Salgotra, R.K.; Sofi, N.R.; Mushtaq, R.; Rohila, J.S.; et al. Physiological and Multi-Omics Approaches for Explaining Drought Stress Tolerance and Supporting Sustainable Production of Rice. Front. Plant Sci. 2022, 12, 803603. [Google Scholar] [CrossRef] [PubMed]
- Muthuramalingam, P.; Jeyasri, R.; Rakkammal, K.; Satish, L.; Shamili, S.; Karthikeyan, A.; Valliammai, A.; Priya, A.; Selvaraj, A.; Gowri, P.; et al. Multi-Omics and Integrative Approach towards Understanding Salinity Tolerance in Rice: A Review. Biology 2022, 11, 1022. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt, C.B.; Carvalho da Silva, T.L.; Rodrigues Neto, J.C.; Vieira, L.R.; Leão, A.P.; de Aquino Ribeiro, J.A.; Abdelnur, P.V.; de Sousa, C.A.F.; Souza, M.T., Jr. Insights from a Multi-Omics Integration (MOI) Study in Oil Palm (Elaeis guineensis Jacq.) Response to Abiotic Stresses: Part One—Salinity. Plants 2022, 11, 1755. [Google Scholar] [CrossRef]
- Singh, R.; Ong-Abdullah, M.; Low, E.-T.L.; Manaf, M.A.A.; Rosli, R.; Nookiah, R.; Ooi, L.C.-L.; Ooi, S.; Chan, K.-L.; Halim, M.A.; et al. Oil Palm Genome Sequence Reveals Divergence of Interfertile Species in Old and New Worlds. Nature 2013, 500, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Ong, A.-L.; Teh, C.-K.; Mayes, S.; Massawe, F.; Appleton, D.R.; Kulaveerasingam, H. An Improved Oil Palm Genome Assembly as a Valuable Resource for Crop Improvement and Comparative Genomics in the Arecoideae Subfamily. Plants 2020, 9, 1476. [Google Scholar] [CrossRef]
- Vieira, L.R.; Silva, V.N.B.; Casari, R.A.D.C.N.; Carmona, P.A.O.; Sousa, C.A.F.D.; Souza Júnior, M.T. Morphophysiological Responses of Young Oil Palm Plants to Salinity Stress. Pesq. Agropec. Bras. 2020, 55, 1835. [Google Scholar] [CrossRef]
- Rodrigues-Neto, J.C.R.; Vieira, L.R.; de Aquino Ribeiro, J.A.; de Sousa, C.A.F.; Júnior, M.T.S.; Abdelnur, P.V. Metabolic Effect of Drought Stress on the Leaves of Young Oil Palm (Elaeis guineensis) Plants Using UHPLC–MS and Multivariate Analysis. Sci. Rep. 2021, 11, 18271. [Google Scholar] [CrossRef]
- Salgado, F.F.; Vieira, L.R.; Silva, V.N.B.; Leão, A.P.; Grynberg, P.; do Carmo Costa, M.M.; Togawa, R.C.; de Sousa, C.A.F.; Júnior, M.T.S. Expression Analysis of MiRNAs and Their Putative Target Genes Confirm a Preponderant Role of Transcription Factors in the Early Response of Oil Palm Plants to Salinity Stress. BMC Plant Biol. 2021, 21, 518. [Google Scholar] [CrossRef]
- Salgado, F.F.; Carvalho da Silva, T.L.; Vieira, L.R.; Silva, V.N.B.; Leão, A.P.; do Carmo Costa, M.M.; Togawa, R.C.; de Sousa, C.A.F.; Grynberg, P.; Júnior, M.T.S. The early response of oil palm (Elaeis guineensis Jacq.) plants to water deprivation: Expression analysis of miRNAs and their putative target genes, and similarities with the response to salinity stress. Front. Plant Sci. 2022; in press. [Google Scholar]
- Luo, F.; Deng, X.; Liu, Y.; Yan, Y. Identification of phosphorylation proteins in response to water deficit during wheat flag leaf and grain development. Bot. Stud. 2018, 59, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damaris, R.N.; Yang, P. Protein Phosphorylation Response to Abiotic Stress in Plants. Methods Mol. Biol. 2021, 2358, 17–43. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Zhang, J.; Pan, C. Integrated Physiological, Proteomic, and Metabolomic Analyses of Pecan Cultivar ‘Pawnee’ Adaptation to Salt Stress. Sci. Rep. 2022, 12, 1841. [Google Scholar] [CrossRef] [PubMed]
- Dahuja, A.; Kumar, R.R.; Sakhare, A.; Watts, A.; Singh, B.; Goswami, S.; Sachdev, A.; Praveen, S. Role of ATP-binding Cassette Transporters in Maintaining Plant Homeostasis under Abiotic and Biotic Stresses. Physiol. Plant. 2021, 171, 785–801. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wang, L.; Zhai, H.; Song, X.; He, S.; Liu, Q. A novel α/β-hydrolase gene IbMas enhances salt tolerance in transgenic sweetpotato. PLoS ONE 2014, 9, e115128. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Fang, J.; Han, X.; He, X.; Liu, M.; Hu, J.; Zhuo, R. Overexpression of quinone reductase from Salix matsudana Koidz enhances salt tolerance in transgenic Arabidopsis thaliana. Gene 2016, 576, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Stavridou, E.; Voulgari, G.; Michailidis, M.; Kostas, S.; Chronopoulou, E.G.; Labrou, N.E.; Madesis, P.; Nianiou-Obeidat, I. Overexpression of A Biotic Stress-Inducible Pvgstu Gene Activates Early Protective Responses in Tobacco under Combined Heat and Drought. Int. J. Mol. Sci. 2021, 22, 2352. [Google Scholar] [CrossRef]
- Bonner, E.R.; Cahoon, R.E.; Knapke, S.M.; Jez, J.M. Molecular basis of cysteine biosynthesis in plants: Structural and functional analysis of O-acetylserine sulfhydrylase from Arabidopsis thaliana. J. Biol. Chem. 2005, 280, 38803–38813. [Google Scholar] [CrossRef] [Green Version]
- Sekowska, A.; Ashida, H.; Danchin, A. Revisiting the methionine salvage pathway and its paralogues. Microb. Biotechnol. 2019, 12, 77–97. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Wang, Y.; Gu, D.; Nan, J.; Chen, S.; Li, H. Overexpression of S-Adenosyl-l-Methionine Synthetase 2 from Sugar Beet M14 Increased Arabidopsis Tolerance to Salt and Oxidative Stress. Int. J. Mol. Sci. 2017, 18, 847. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Nadira, U.A.; Qiu, C.W.; Cao, F.; Chen, Z.H.; Vincze, E.; Wu, F. The Barley S-Adenosylmethionine Synthetase 3 Gene HvSAMS3 Positively Regulates the Tolerance to Combined Drought and Salinity Stress in Tibetan Wild Barley. Cells 2020, 9, 1530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yan, S.; Zhang, S.; Yan, P.; Wang, J.; Zhang, H. Glutathione, carbohydrate and other metabolites of Larix olgensis A. Henry reponse to polyethylene glycol-simulated drought stress. PLoS ONE 2021, 16, e0253780. [Google Scholar] [CrossRef] [PubMed]
- Okemo, P.; Long, H.; Cheng, Y.; Mundree, S.; Williams, B. Stachyose triggers apoptotic like cell death in drought sensitive but not resilient plants. Sci. Rep. 2021, 11, 7099. [Google Scholar] [CrossRef] [PubMed]
- John, R.; Ganeshan, U.; Singh, B.N.; Kaul, T.; Reddy, M.K.; Sopory, S.K.; Rajam, M.V. Over-expression of Topoisomerase II Enhances Salt Stress Tolerance in Tobacco. Front. Plant Sci. 2016, 7, 1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandian, B.A.; Sathishraj, R.; Djanaguiraman, M.; Prasad, P.; Jugulam, M. Role of Cytochrome P450 Enzymes in Plant Stress Response. Antioxidants 2020, 9, 454. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucl. Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Carvalho, P.C.; Fischer, J.S.G.; Xu, T.; Cociorva, D.; Balbuena, T.S.; Valente, R.H.; Perales, J.; Yates, J.R.; Barbosa, V.C. Search Engine Processor: Filtering and Organizing Peptide Spectrum Matches. Proteomics 2012, 12, 944–949. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, 1002. [Google Scholar] [CrossRef]
- Chong, J.; Xia, J. Using MetaboAnalyst 4.0 for Metabolomics Data Analysis, Interpretation, and Integration with Other Omics Data. In Computational Methods and Data Analysis for Metabolomics. Methods in Molecular Biology; Li, S., Ed.; Humana: New York, NY, USA, 2020; Volume 2104, pp. 337–360. [Google Scholar]
- Brink, B.G.; Seidel, A.; Kleinbölting, N.; Nattkemper, T.W.; Albaum, S.P. Omics Fusion–A Platform for Integrative Analysis of Omics Data. J. Integr. Bioinform. 2016, 13, 43–46. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Stressed | Total | |
|---|---|---|---|
| Peptide Spectrum Match (PSM) | 6733 | 6495 | 13,065 |
| Total number of peptides | 3659 | 3824 | 5182 |
| Number of unique peptides | 2098 | 2088 | 3068 |
| Total number of proteins entries | 1353 | 1617 | 1859 |
| Total number of proteins using the maximum parsimony criterion | 762 | 934 | 1085 |
| Pathway Name | Pathway ID | Occurrence of | Unique Molecules | ||
|---|---|---|---|---|---|
| Transcripts | Proteins | Metabolites | |||
| Cysteine and methionine metabolism | 270 | 30 | 8 | 14 | 47 |
| Purine metabolism | 230 | 25 | 3 | 17 | 42 |
| Porphyrin and chlorophyll metabolism | 860 | 24 | 4 | 12 | 37 |
| Amino sugar and nucleotide sugar metabolism | 520 | 25 | 4 | 9 | 35 |
| Glycine, serine, and threonine metabolism | 260 | 24 | 7 | 9 | 34 |
| Pyrimidine metabolism | 240 | 17 | 1 | 16 | 33 |
| Glycolysis/Gluconeogenesis | 10 | 26 | 9 | 4 | 30 |
| Phenylpropanoid biosynthesis | 940 | 12 | 3 | 18 | 30 |
| Pentose phosphate pathway | 30 | 17 | 8 | 11 | 29 |
| Arginine and proline metabolism | 330 | 16 | 1 | 13 | 29 |
| Aminoacyl-tRNA biosynthesis | 970 | 13 | 1 | 15 | 29 |
| Glyoxylate and dicarboxylate metabolism | 630 | 17 | 8 | 8 | 28 |
| Pyruvate metabolism | 620 | 20 | 6 | 5 | 26 |
| Starch and sucrose metabolism | 500 | 22 | 5 | 3 | 26 |
| Tyrosine metabolism | 350 | 13 | 2 | 12 | 25 |
| Phenylalanine, tyrosine, and tryptophan biosynthesis | 400 | 17 | 1 | 8 | 25 |
| Methane metabolism | 680 | 16 | 5 | 8 | 24 |
| Alanine, aspartate, and glutamate metabolism | 250 | 18 | 5 | 6 | 24 |
| Lysine degradation | 310 | 14 | 3 | 9 | 24 |
| Tryptophan metabolism | 380 | 15 | 4 | 7 | 23 |
| Terpenoid backbone biosynthesis | 900 | 17 | 4 | 5 | 23 |
| Carbon fixation in photosynthetic organisms | 710 | 19 | 6 | 3 | 22 |
| Citrate cycle (TCA cycle) | 20 | 16 | 8 | 5 | 21 |
| Arginine biosynthesis | 220 | 9 | 3 | 11 | 21 |
| Galactose metabolism | 52 | 12 | 3 | 8 | 21 |
| Valine, leucine, and isoleucine degradation | 280 | 15 | 2 | 5 | 21 |
| Carbon fixation pathways in prokaryotes | 720 | 13 | 4 | 6 | 20 |
| Glutathione metabolism | 480 | 13 | 3 | 7 | 20 |
| Glycerolipid metabolism | 561 | 15 | 2 | 4 | 20 |
| Ascorbate and aldarate metabolism | 53 | 13 | 1 | 7 | 20 |
| Protein ID | Gene ID * | KO Number | EC | Source | Profile | Fold Change | Description_KEGG | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Salt | Drought | Salt | Drought | Salt | Drought | |||||
| A0A6J0PH47 | LOC105044080 | K02641 | 1.18.1.2 | Common | Common | Down | Down | −1.885 | −3.730 | ferredoxin-NADP+ reductase |
| B3TLY5 | CAT2 | K03781 | 1.11.1.6 | Common | Common | Up | Up | 1.834 | 3.177 | catalase |
| B3TM49 | LOC105035746 | K08967 | 1.13.11.53 | Only Control | Common | Down | Up | NA | 3.277 | 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase |
| 1.13.11.54 | ||||||||||
| A0A6I9QHS4 | 1.13.11.53 | Down | Up | NA | 2.463 | |||||
| 1.13.11.54 | ||||||||||
| A0A6J0PS13 | LOC105039298 | K15918 | 2.7.1.31 | Only Control | Only Control | Down | Down | NA | NA | D-glycerate 3-kinase |
| A0A6I9RVC5 | LOC105053135 | K05396 | 4.4.1.15 | Only Control | Only Control | Down | Down | NA | NA | D-cysteine desulfhydrase |
| A0A6J0PP21 | LOC105053549 | K00434 | 1.11.1.11 | Only Control | Only Stressed | Down | Up | NA | NA | L-ascorbate peroxidase |
| A0A6I9S451 | ||||||||||
| A0A6I9RWM2 | LOC105053973 | K00344 | 1.6.5.5 | Only Control | Only Stressed | Down | Up | NA | NA | NADPH: quinone reductase |
| A0A6I9QWL2 | LOC105040656 | K03405 | 6.6.1.1 | Only Stressed | Only Stressed | Up | Up | NA | NA | magnesium chelatase subunit I |
| A0A6I9QXL0 | LOC105041662 | K10525 | 5.3.99.6 | Only Stressed | Only Stressed | Up | Up | NA | NA | allene oxide cyclase |
| A0A6J0PKN5 | LOC105048171 | K03405 | 6.6.1.1 | Only Stressed | Only Stressed | Up | Up | NA | NA | magnesium chelatase subunit I |
| A0A6I9QUV8 | LOC105040597 | K00430 | 1.11.1.7 | Only Stressed | Only Stressed | Up | Up | NA | NA | peroxidase |
| M1H922 | LOC105057156 | K13379 | 2.4.1.- | Only Stressed | Only Stressed | Up | Up | NA | NA | reversibly glycosylated polypeptide/UDP-arabinopyranose mutase |
| 5.4.99.30 | ||||||||||
| A0A6I9R7V0 | LOC105045344 | K25108 | 3.2.1.73 | Only Stressed | Only Stressed | Up | Up | NA | NA | licheninase |
| A0A6I9R784 | LOC105045348 | |||||||||
| A0A6I9QEA6 | LOC105033962 | |||||||||
| A0A6I9R7N0 | LOC105045343 | |||||||||
| A0A6I9QD17 | LOC105033964 | |||||||||
| Name | Type | EC | KO | Salinity | Drought | Description | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| logFC | Profile | Source | logFC | Profile | Source | |||||
| XP_010905665.1 | TRNS | 2.3.1.30 | K00640 | 3.4 | Up | NA | 4.7 | Up | NA | cysE; serine O-acetyltransferase |
| XP_010914049.1 | TRNS | 4.1.1.50 | K01611 | 0.7 | Up | NA | 2.1 | Up | NA | speD, AMD1; S-adenosylmethionine decarboxylase |
| XP_010915297.1 | TRNS | 2.5.1.47 | K01738 | 0.9 | Up | NA | 2.5 | Up | NA | cysK; cysteine synthase |
| XP_010920378.1 | TRNS | 2.6.1.1 | K14454 | 1.0 | Up | NA | 3.8 | Up | NA | GOT1; aspartate aminotransferase, cytoplasmic |
| XP_010921340.1 | TRNS | 2.5.1.6 | K00789 | 1.4 | Up | NA | 2.4 | Up | NA | metK, MAT; S-adenosylmethionine synthetase |
| XP_010924485.1 | TRNS | 2.3.1.30 | K00640 | −0.9 | Down | NA | −0.7 | Down | NA | cysE; serine O-acetyltransferase |
| XP_010927510.1 | TRNS | 1.1.1.95 | K00058 | −0.8 | Down | NA | −1.2 | Down | NA | serA, PHGDH; D-3-phosphoglycerate dehydrogenase/2-oxoglutarate reductase |
| XP_010928399.1 | TRNS | 2.7.1.100 | K00899 | −1.0 | Down | NA | −2.5 | Down | NA | mtnK; 5-methylthioribose kinase |
| XP_010931932.1 | TRNS | 6.3.2.3 | K21456 | −0.5 | Down | NA | −1.1 | Down | NA | GSS; glutathione synthase |
| XP_010938608.1 | TRNS | 2.6.1.44 | K00830 | −0.6 | Down | NA | −3.2 | Down | NA | AGXT; alanine-glyoxylate transaminase/serine-glyoxylate transaminase/serine-pyruvate transaminase |
| XP_010939394.1 | TRNS | 2.6.1.117 | K23977 | −0.5 | Down | NA | −0.7 | Down | NA | GTK; L-glutamine---4-(methylsulfanyl)-2-oxobutanoate aminotransferase |
| XP_010941130.1 | TRNS | 1.1.1.27 | K00016 | −3.4 | Down | NA | −1.7 | Down | NA | LDH, ldh; L-lactate dehydrogenase |
| XP_010942212.1 | TRNS | 2.5.1.16 | K00797 | −0.7 | Down | NA | 0.5 | Up | NA | speE, SRM, SPE3; spermidine synthase |
| XP_010943220.1 | TRNS | 1.1.1.37 | K00026 | −0.6 | Down | NA | −1.9 | Down | NA | MDH2; malate dehydrogenase |
| XP_019711215.1 | TRNS | 2.6.1.5 | K00815 | 0.8 | Up | NA | 1.0 | Up | NA | TAT; tyrosine aminotransferase |
| C00051 | METB | - | NA | 0.3 | Up | NA | 0.9 | Up | NA | Glutathione |
| C00049 | METB | - | NA | 1.1 | Up | NA | 4.6 | Up | NA | L-Aspartate |
| A0A6I9QHS4 | PROT | 1.13.11.53 | K08967 | NA | NA | OC | 1.3 | Up | COM | 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase |
| 1.13.11.54 | ||||||||||
| A0A6I9RVC5 | PROT | 4.4.1.15 | K05396 | NA | NA | OC | NA | NA | OC | D-cysteine desulfhydrase |
| B3TM49 | PROT | 1.13.11.53 | K08967 | NA | NA | OC | 1.7 | Up | COM | 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase |
| 1.13.11.54 | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leão, A.P.; Bittencourt, C.B.; Carvalho da Silva, T.L.; Rodrigues Neto, J.C.; Braga, Í.d.O.; Vieira, L.R.; de Aquino Ribeiro, J.A.; Abdelnur, P.V.; de Sousa, C.A.F.; Souza Júnior, M.T. Insights from a Multi-Omics Integration (MOI) Study in Oil Palm (Elaeis guineensis Jacq.) Response to Abiotic Stresses: Part Two—Drought. Plants 2022, 11, 2786. https://doi.org/10.3390/plants11202786
Leão AP, Bittencourt CB, Carvalho da Silva TL, Rodrigues Neto JC, Braga ÍdO, Vieira LR, de Aquino Ribeiro JA, Abdelnur PV, de Sousa CAF, Souza Júnior MT. Insights from a Multi-Omics Integration (MOI) Study in Oil Palm (Elaeis guineensis Jacq.) Response to Abiotic Stresses: Part Two—Drought. Plants. 2022; 11(20):2786. https://doi.org/10.3390/plants11202786
Chicago/Turabian StyleLeão, André Pereira, Cleiton Barroso Bittencourt, Thalliton Luiz Carvalho da Silva, Jorge Cândido Rodrigues Neto, Ítalo de Oliveira Braga, Letícia Rios Vieira, José Antônio de Aquino Ribeiro, Patrícia Verardi Abdelnur, Carlos Antônio Ferreira de Sousa, and Manoel Teixeira Souza Júnior. 2022. "Insights from a Multi-Omics Integration (MOI) Study in Oil Palm (Elaeis guineensis Jacq.) Response to Abiotic Stresses: Part Two—Drought" Plants 11, no. 20: 2786. https://doi.org/10.3390/plants11202786