
Long−Distance Wind Dispersal Drives Population Range Expansion of Solidago canadensis
Abstract
:1. Introduction
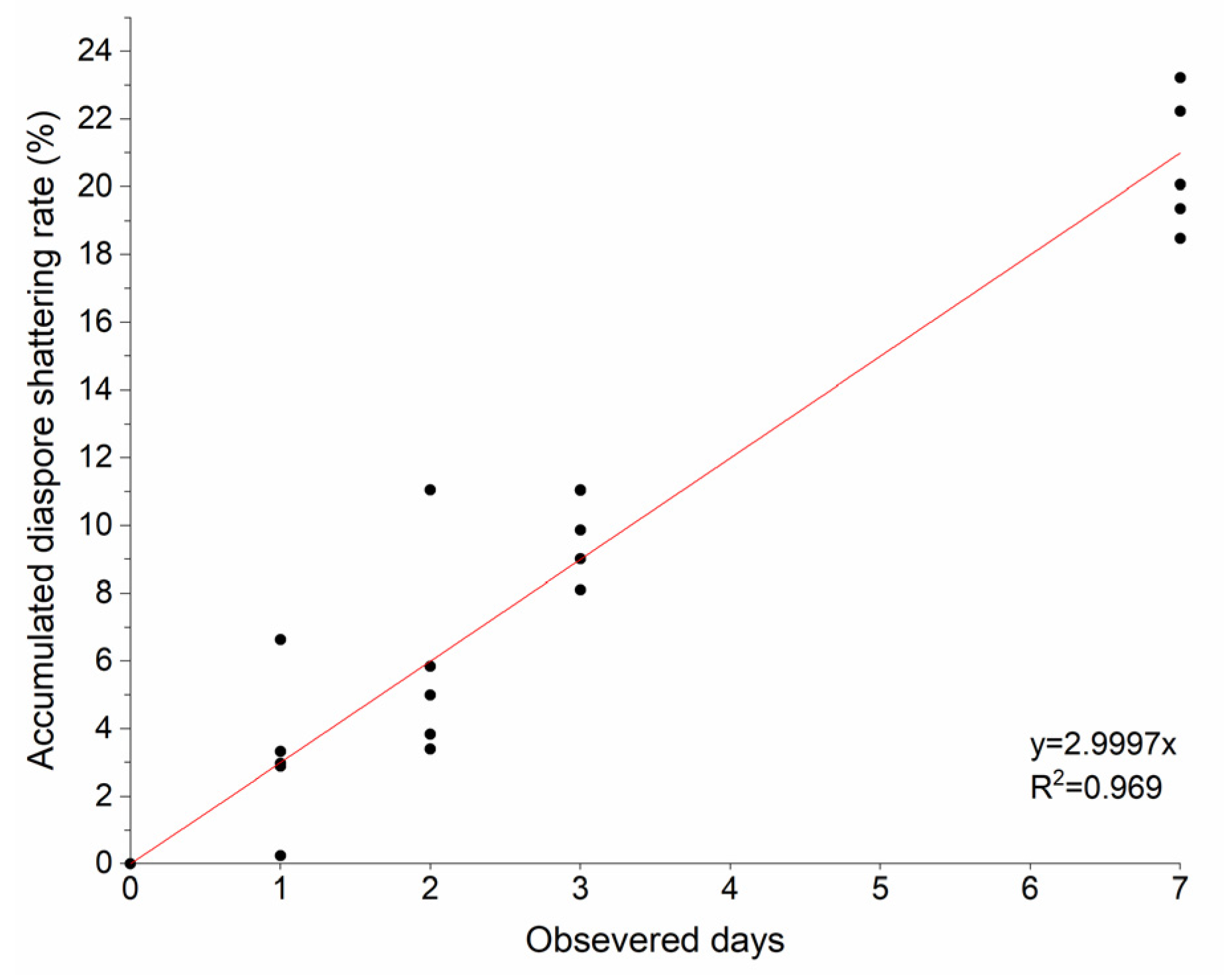
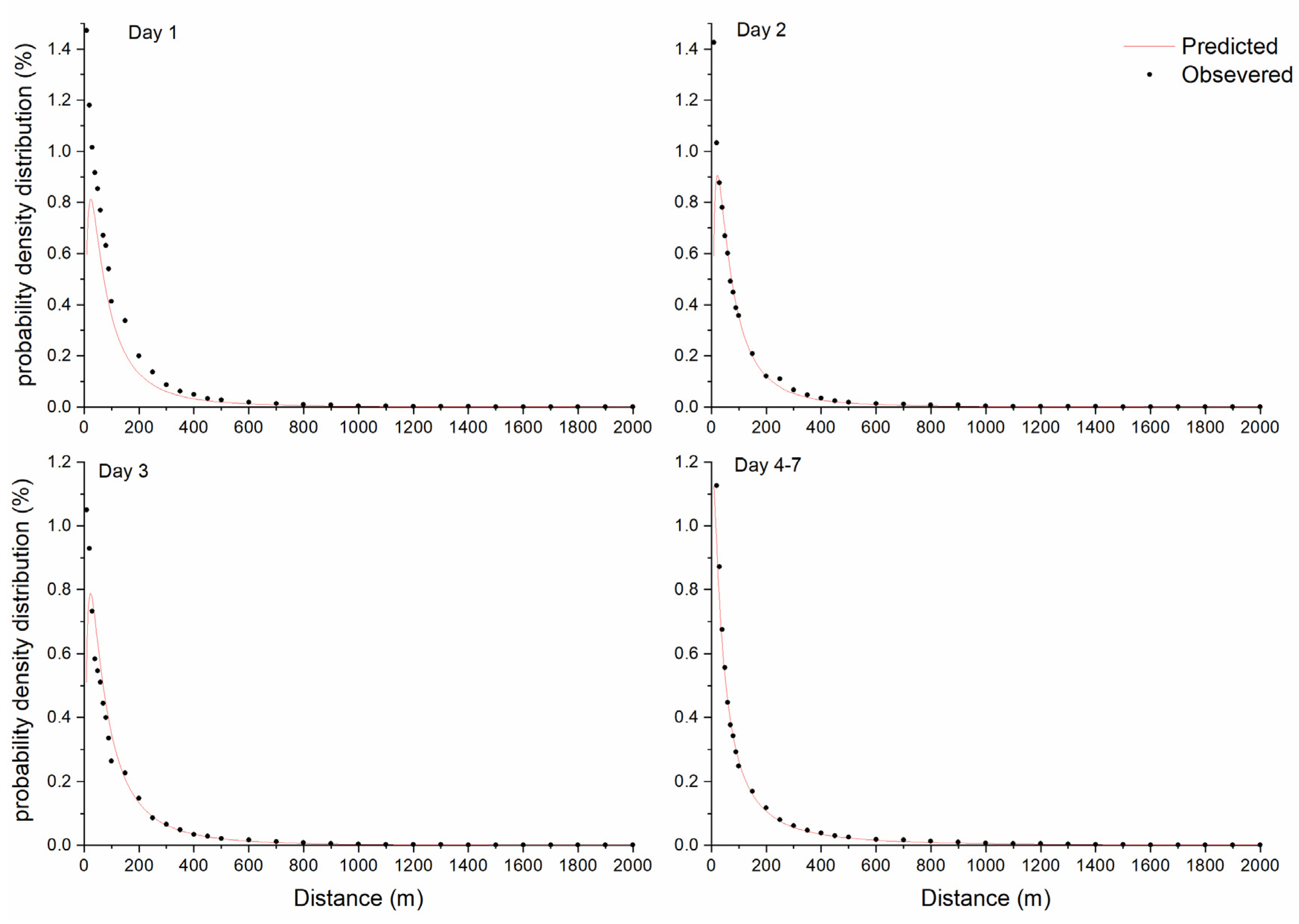
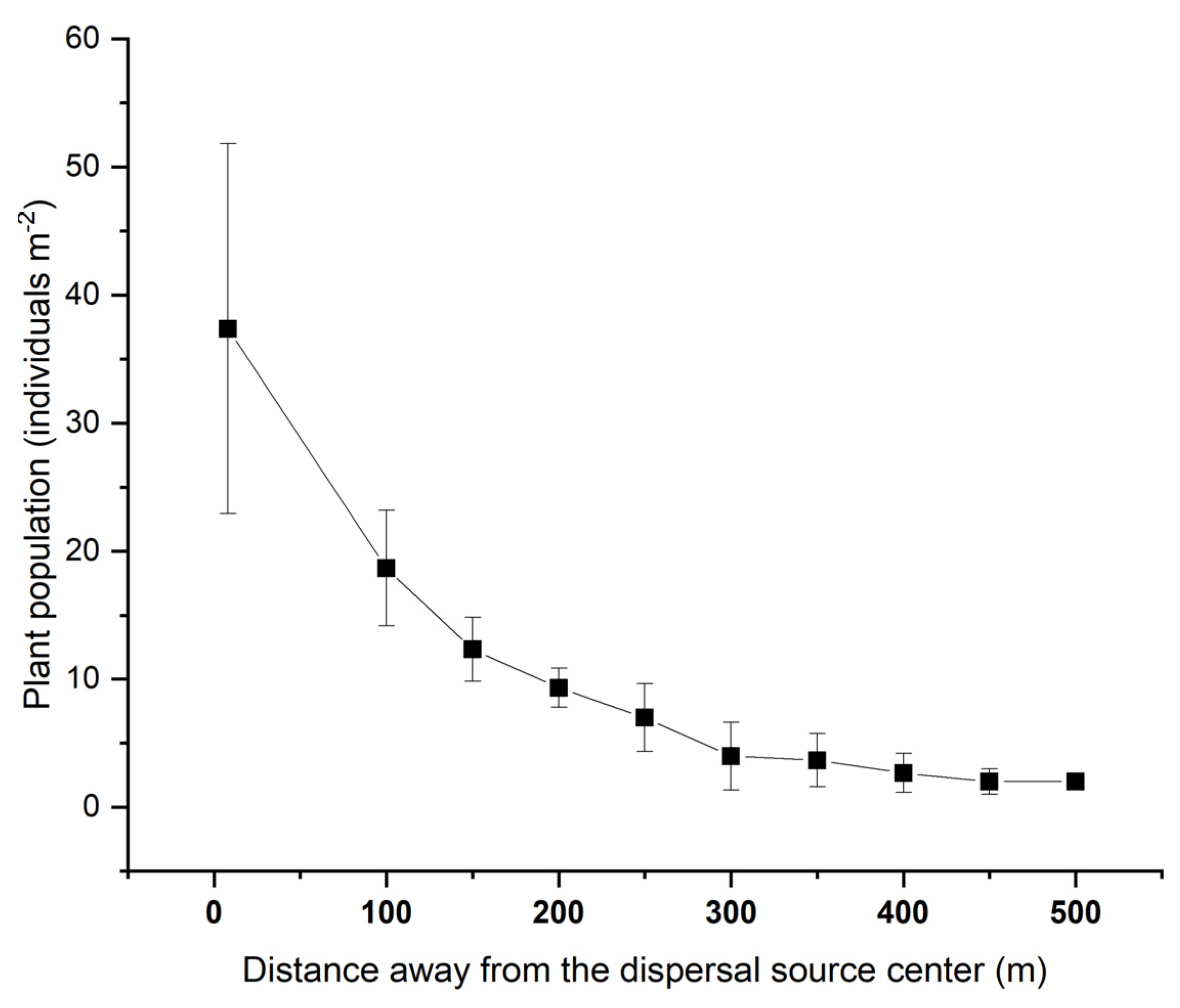
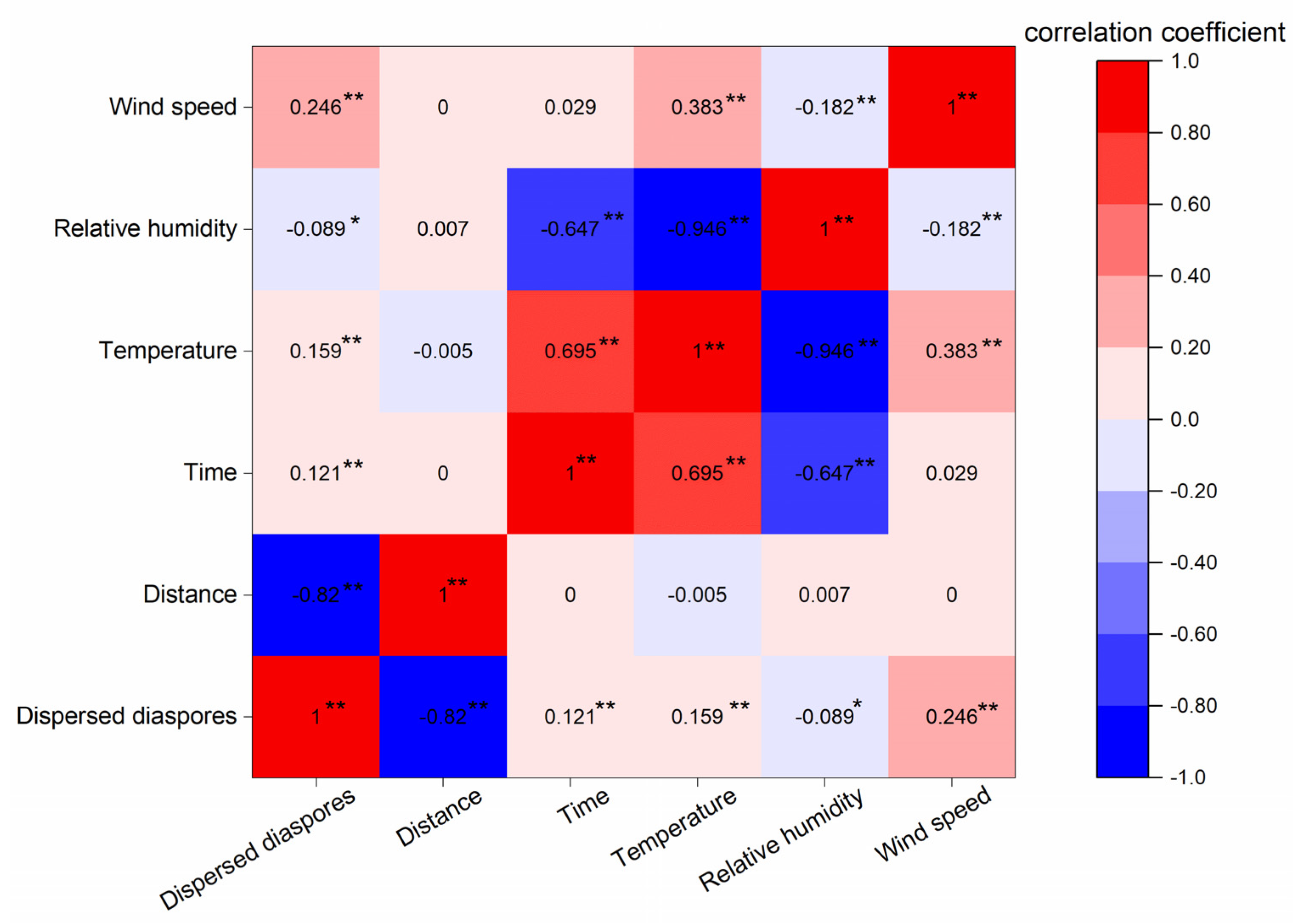
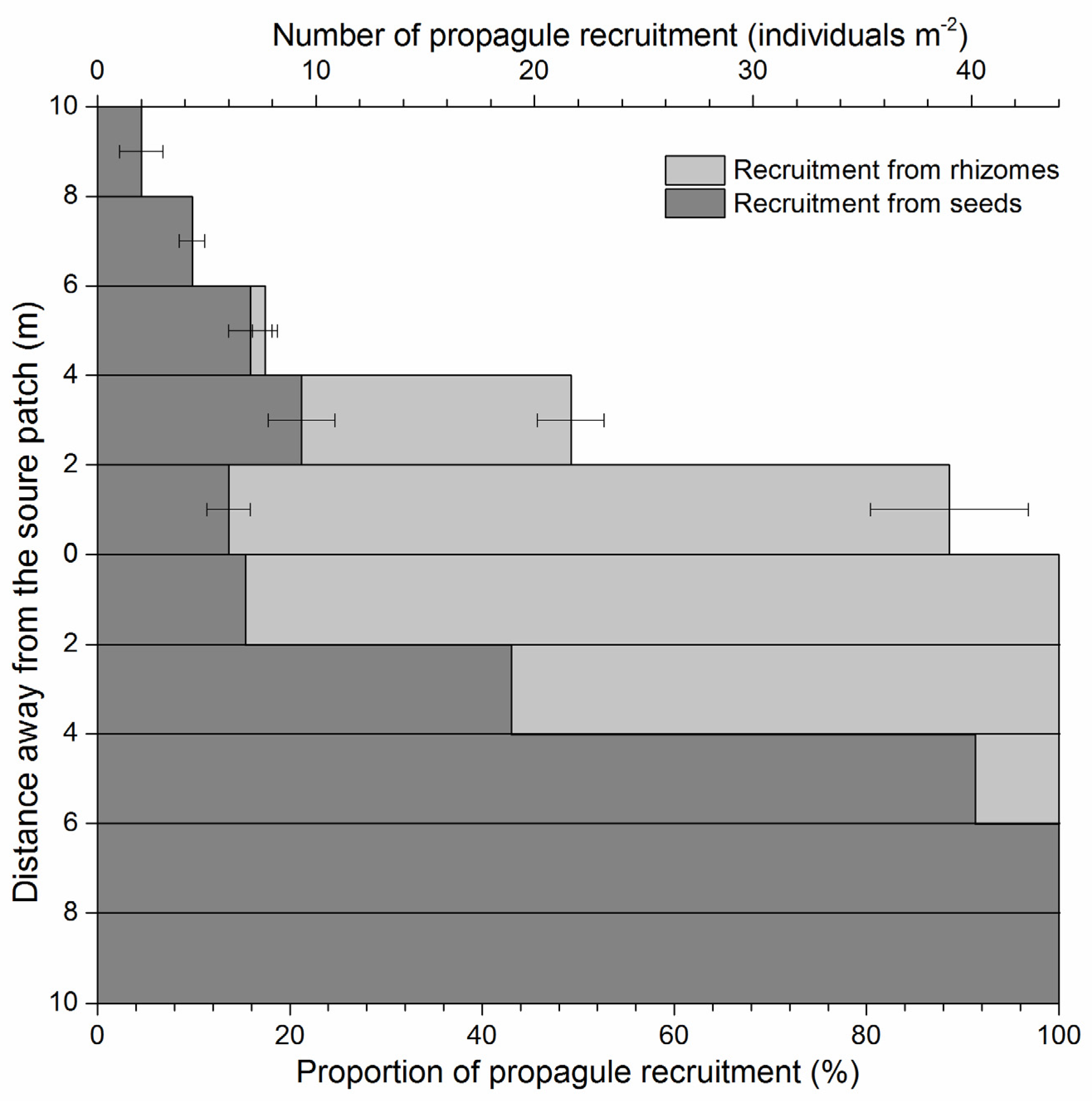
2. Results
The Distribution of Dispersed Diaspores in All Sampled Directions
3. Discussion
3.1. Seed Dispersal and Seedling Recruitment of S. canadensis
3.2. Factors Influencing Wind Release and Dispersal
4. Materials and Methods
4.1. Study Site
4.2. Staining In Situ
4.3. Seed Sampling
4.4. Plant Investigation of S. canadensis
4.5. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Antonio, C.M.; Vitousek, P.M. Biological Invasions by Exotic Grasses, the Grass/Fire Cycle, and Global Change. Annu. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar] [CrossRef]
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; RejmÁNek, M.; Westbrooks, R. Introduced species: A significant component of human−caused global change. N. Z. J. Ecol. 1997, 21, 1–16. [Google Scholar]
- Byers, J.E.; Reichard, S.; Randall, J.M.; Parker, I.M.; Smith, C.S.; Lonsdale, W.M.; Atkinson, I.A.E.; Seastedt, T.R.; Williamson, M.; Chornesky, E.; et al. Directing Research to Reduce the Impacts of Nonindigenous Species. Conserv. Biol. 2002, 16, 630–640. [Google Scholar] [CrossRef] [Green Version]
- Pejchar, L.; Mooney, H.A. Invasive species, ecosystem services and human well−being. Trends Ecol. Evol. 2009, 24, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Harter, D.E.; Irl, S.D.; Seo, B.; Steinbauer, M.J.; Gillespie, R.; Triantis, K.A.; Fernández-Palacios, J.-M.;; Beierkuhnlein, C. Impacts of global climate change on the floras of oceanic islands—Projections, implications and current knowledge. Perspect. Plant Ecol. 2015, 17, 160–183. [Google Scholar] [CrossRef]
- Wu, S.-H.; Wang, H.-H. Potential Asteraceae invaders in Taiwan: Insights from the flora and herbarium records of casual and naturalized alien species. Taiwania 2005, 50, 62–70. [Google Scholar]
- Pyšek, P. Alien and native species in Central European urban floras: A quantitative comparison. J. Biogeogr. 1998, 25, 155–163. [Google Scholar] [CrossRef]
- Vanderhoeven, S.; Dassonville, N.; Chapuis−Lardy, L.; Hayez, M.; Meerts, P. Impact of the invasive alien plant Solidago giganteaon primary productivity, plant nutrient content and soil mineral nutrient concentrations. Plant Soil 2006, 286, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Scharfy, D.; Güsewell, S.; Gessner, M.O.; Venterink, H.O. Invasion of Solidago gigantea in contrasting experimental plant communities: Effects on soil microbes, nutrients and plant−soil feedbacks. J. Ecol. 2010, 98, 1379–1388. [Google Scholar] [CrossRef]
- Del Fabbro, C.; Güsewell, S.; Prati, D. Allelopathic effects of three plant invaders on germination of native species: A field study. Biol. Invasions 2014, 16, 1035–1042. [Google Scholar] [CrossRef]
- Xu, H.; Sheng, Q. Invasive Alien Species in China; Science Press: Beijing, China, 2018. [Google Scholar]
- Notice on the Publication of the List of the Second Invasive Alien Species in China. Available online: http://www.mee.gov.cn/gkml/hbb/bwj/201001/t20100126_184831.htm (accessed on 12 June 2021).
- Weber, E. Current and potential ranges of three exotic goldenrods (Solidago) in Europe. Conserv. Biol. 2001, 15, 122–128. [Google Scholar] [CrossRef]
- Cheng, J.; Li, J.; Zhang, Z.; Lu, H.; Chen, G.; Yao, B.; Dong, Y.; Ma, L.; Yuan, X.; Xu, J. Autopolyploidy−driven range expansion of a temperate−originated plant to pan−tropic under global change. Ecol. Monogr. 2021, 91, e01445. [Google Scholar] [CrossRef]
- Fryxell, P.A. Mode of reproduction of higher plants. Bot. Rev. 1957, 23, 135–233. [Google Scholar] [CrossRef]
- Mulligan, G.A.; Findlay, J.N. Reproductive systems and colonization in Canadian weeds. Can. J. Bot. 1970, 48, 859–860. [Google Scholar] [CrossRef]
- Jian−Zhong, L.; En−Sheng, W.; Xiao−Wen, W.; Weber, E.; Bin, Z.; Bo, L. Potential distribution of Solidago canadensis in China. J. Syst. Evol. 2007, 45, 670–674. [Google Scholar]
- Mei, D.; Jian−Zhong, L.; Wen−Ju, Z.; Jia−Kuan, C.; Bo, L. Canada goldenrod (Solidago canadensis): An invasive alien weed rapidly spreading in China. J. Syst. Evol. 2006, 44, 72–85. [Google Scholar]
- Baker, H. Patterns of plant invasion in North America. In Ecology of Biological Invasions of North America and Hawaii; Springer: Berlin/Heidelberg, Germany, 1986; pp. 44–57. [Google Scholar]
- Newsome, A.; Noble, I. Ecological and physiological characters of invading species. In Ecology of Biological Invasions; Cambridge University Press: Cambridge, UK, 1986; pp. 1–20. [Google Scholar]
- Baker, H.G. The evolution of weeds. Annu. Rev. Ecol. Syst. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Meyer, A.H.; Schmid, B. Seed dynamics and seedling establishment in the invading perennial Solidago altissima under different experimental treatments. J. Ecol. 1999, 87, 28–41. [Google Scholar] [CrossRef]
- Huang, H.; Guo, S.; Chen, G. Reproductive biology in an invasive plant Solidago canadensis. Front. Biol. China 2007, 2, 196–204. [Google Scholar] [CrossRef]
- Andersen, M.C. Diaspore morphology and seed dispersal in several wind−dispersed Asteraceae. Am. J. Bot. 1993, 80, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Gravuer, K.; Von Wettberg, E.J.; Schmitt, J. Dispersal biology of Liatris scariosa var. novae−angliae (Asteraceae), a rare New England grassland perennial. Am. J. Bot. 2003, 90, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Imbert, E. The effects of achene dimorphism on the dispersal in time and space in Crepis sancta (Asteraceae). Can. J. Bot. 1999, 77, 508–513. [Google Scholar]
- Pazos, G.E.; Greene, D.F.; Katul, G.; Bertiller, M.B.; Soons, M.B. Seed dispersal by wind: Towards a conceptual framework of seed abscission and its contribution to long−distance dispersal. J. Ecol. 2013, 101, 889–904. [Google Scholar] [CrossRef]
- Zhang, Z.; Shi, X.; Li, R.; Qiang, S. A new technique for stain−marking of seeds with safranine to track seed dispersal and seed bank dynamics. Front. Plant Sci. 2022, 13, 959046. [Google Scholar] [CrossRef]
- Reluga, T.C.; Shaw, A.K. Resource distribution drives the adoption of migratory, partially migratory, or residential strategies. Theor. Ecol. 2015, 8, 437–447. [Google Scholar] [CrossRef]
- Jordano, P. What is long−distance dispersal? And a taxonomy of dispersal events. J. Ecol. 2017, 105, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Nathan, R. Long−distance dispersal of plants. Science 2006, 313, 786–788. [Google Scholar] [CrossRef] [Green Version]
- Cain, M.L.; Milligan, B.G.; Strand, A.E. Long−distance seed dispersal in plant populations. Am. J. Bot. 2000, 87, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Nathan, R.; Perry, G.; Cronin, J.T.; Strand, A.E.; Cain, M.L. Methods for estimating long−distance dispersal. Oikos 2003, 103, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, K.; Fenner, M. Ecology of seedling regeneration. In Seeds: The Ecology of Regeneration in Plant Communities; CABI Publishing: Wallingford, UK, 2000; pp. 331–359. [Google Scholar]
- Augspurger, C.K.; Franson, S.E.; Cushman, K.C.; Muller−Landau, H.C. Intraspecific variation in seed dispersal of a Neotropical tree and its relationship to fruit and tree traits. Ecol. Evol. 2016, 6, 1128–1142. [Google Scholar] [CrossRef]
- Yang, X.; Cheng, J.; Yao, B.; Lu, H.; Zhang, Y.; Xu, J.; Song, X.; Sowndhararajan, K.; Qiang, S. Polyploidy−promoted phenolic metabolism confers the increased competitive ability of Solidago canadensis. Oikos 2021, 130, 1014–1025. [Google Scholar] [CrossRef]
- Greene, D.; Quesada, M.; Calogeropoulos, C. Dispersal of seeds by the tropical sea breeze. Ecology 2008, 89, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Soons, M.B.; Bullock, J.M. Non−random seed abscission, long−distance wind dispersal and plant migration rates. J. Ecol. 2008, 96, 581–590. [Google Scholar] [CrossRef]
- Kuparinen, A. Mechanistic models for wind dispersal. Trends Plant Sci. 2006, 11, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Cousens, R.; Dytham, C.; Law, R. Dispersal in Plants: A Population Perspective; Oxford University Press: New York, NY, USA, 2008. [Google Scholar]
- Greene, D.F. The role of abscission in long−distance seed dispersal by the wind. Ecology 2005, 86, 3105–3110. [Google Scholar] [CrossRef]
- Jongejans, E.; Skarpaas, O.; Tipping, P.W.; Shea, K. Establishment and spread of founding populations of an invasive thistle: The role of competition and seed limitation. Biol. Invasions 2007, 9, 317–325. [Google Scholar] [CrossRef]
- Marchetto, K.M.; Jongejans, E.; Shea, K.; Auhl, R. Water loss from flower heads predicts seed release in two invasive thistles. Plant Ecol. Divers. 2012, 5, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Savage, D.; Barbetti, M.J.; MacLeod, W.J.; Salam, M.U.; Renton, M. Seasonal and diurnal patterns of spore release can significantly affect the proportion of spores expected to undergo long−distance dispersal. Microbi. Ecol. 2012, 63, 578–585. [Google Scholar] [CrossRef]
- Isard, S.A.; Gage, S.H. Flow of Life in the Atmosphere; Michigan State University Press: East Lansing, MI, USA, 2001. [Google Scholar]
- Greene, D.; Johnson, E. A model of wind dispersal of winged or plumed seeds. Ecology 1989, 70, 339–347. [Google Scholar] [CrossRef]
- Guo, Q.; Zhu, M.; Xu, L.; Zhai, B. Seed settlement velocity of five weed species. Chin. J. Ecol. 2008, 27, 519–523, (In Chinese with English Abstract). [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters (Unit) | Day 1 | Day 2 | Day 3 | Day 7 |
|---|---|---|---|---|
| Q (seeds) | 1,887,375 | 1,535,267 | 2,346,756 | 6,378,962 |
| H (m) | 1.79 | 1.79 | 1.79 | 1.79 |
| σu (m s−1) | 1.11 | 1.09 | 1.14 | 1.48 |
| UH (m s−1) | 6.46 | 5.78 | 6.71 | 5.68 |
| F (m s−1) | 0.141 | 0.141 | 0.141 | 0.141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Wen, G.; Bu, D.; Sun, G.; Qiang, S. Long−Distance Wind Dispersal Drives Population Range Expansion of Solidago canadensis. Plants 2022, 11, 2734. https://doi.org/10.3390/plants11202734
Zhang Z, Wen G, Bu D, Sun G, Qiang S. Long−Distance Wind Dispersal Drives Population Range Expansion of Solidago canadensis. Plants. 2022; 11(20):2734. https://doi.org/10.3390/plants11202734
Chicago/Turabian StyleZhang, Zheng, Guangyue Wen, Dexiao Bu, Guojun Sun, and Sheng Qiang. 2022. "Long−Distance Wind Dispersal Drives Population Range Expansion of Solidago canadensis" Plants 11, no. 20: 2734. https://doi.org/10.3390/plants11202734