1. Introduction
Rice (
Oryza sativa L.) is one of the most important food crops that grows in tropical to temperate regions worldwide [
1]. Low temperature is a severe environmental restriction that most strongly impacts rice growth and development, especially at the early seedling and reproductive stages [
2]. Cold stress at the early vegetative stage can result in stunted growth and increased seedling mortality, which in turn, leads to uneven seedling stand establishment, delayed panicle development, spikelet sterility, and subsequently, decreased yield of rice [
3,
4]. Therefore, it is a crucial object in rice cultivation and breeding to improve cold tolerance at seedling stage.
Cold tolerance of rice is a complex quantitative trait controlled by multiple quantitative trait loci (QTL), which is directly related to a large amount of physiological and biochemical processes and environmental factors [
3,
4]. To date, numerous backcross inbred lines, recombinant inbred lines, and near-isogenic lines have been developed using many cold tolerance varieties as donors, and a massive number of cold tolerance-related QTLs have been identified on 12 chromosomes at different development stages in rice [
5,
6]. However, these are not sufficient to mine the useful genes and reveal the molecular mechanisms of cold tolerance. In recent years, the methods to obtain a single nucleotide polymorphism (SNP) molecular marker based on the high-throughput sequencing technologies, such as genotyping-by-sequencing (GBS), genome-wide association studies (GWAS), and bulked segregant RNA-seq (BSR), are increasingly used to identify QTLs across the whole genome [
7,
8]. In particular, as GBS technology is rapid, simple, and easy to implement for QTL mapping, genomic selection, and genetic diversity, it has been successfully applied in QTL mapping for cold tolerance in rice [
9,
10].
Dongxiang wild rice (
Oryza rufpogon Grif, DXWR) is a relative ancestor of the cultivated rice, which can be used as a donor of novel and favorable alleles for rice breeding [
11,
12,
13]. DXWR has a high tolerance to cold stress at all growth stages, and its underground stem can tolerate temperatures up to −12.8 °C [
14]. Many QTLs associated with cold tolerance have been identified in DXWR [
15,
16,
17,
18,
19]. Therefore, DXWR is an elite material for enhancing the cold tolerance of rice by hybridization, backcrossing or genetic transformation.
Brassinosteroids (BR) are important steroid hormones, which play vital roles in the growth, development, and tolerance to various stresses in plants. The exogenous BR application can enhance low-temperature tolerance in plants [
20,
21,
22]. It was shown that BRASSINOSTEROID INSENSITIVE 1 (BRI1) and BRASSINAZOLE RESISTANT1 (BZR1) positively regulate cold tolerance, whereas BRASSINOSTEROID INSENSITIVE 2 (BIN2) negatively regulates cold tolerance in
Arabidopsis thaliana. In addition, several genes related to BR biosynthesis,
BR6ox2,
DWF4, and
CPD, are rapidly downregulated under low-temperature stress [
23,
24,
25,
26]. In rice, the treatments with BR can also effectively improve cold tolerance at germination and seedling stages [
27,
28,
29,
30]. To date, however, few genes associated with both cold tolerance and BR treatments in rice have been studied. The regulatory relationship between cold tolerance and BR remain largely unclear.
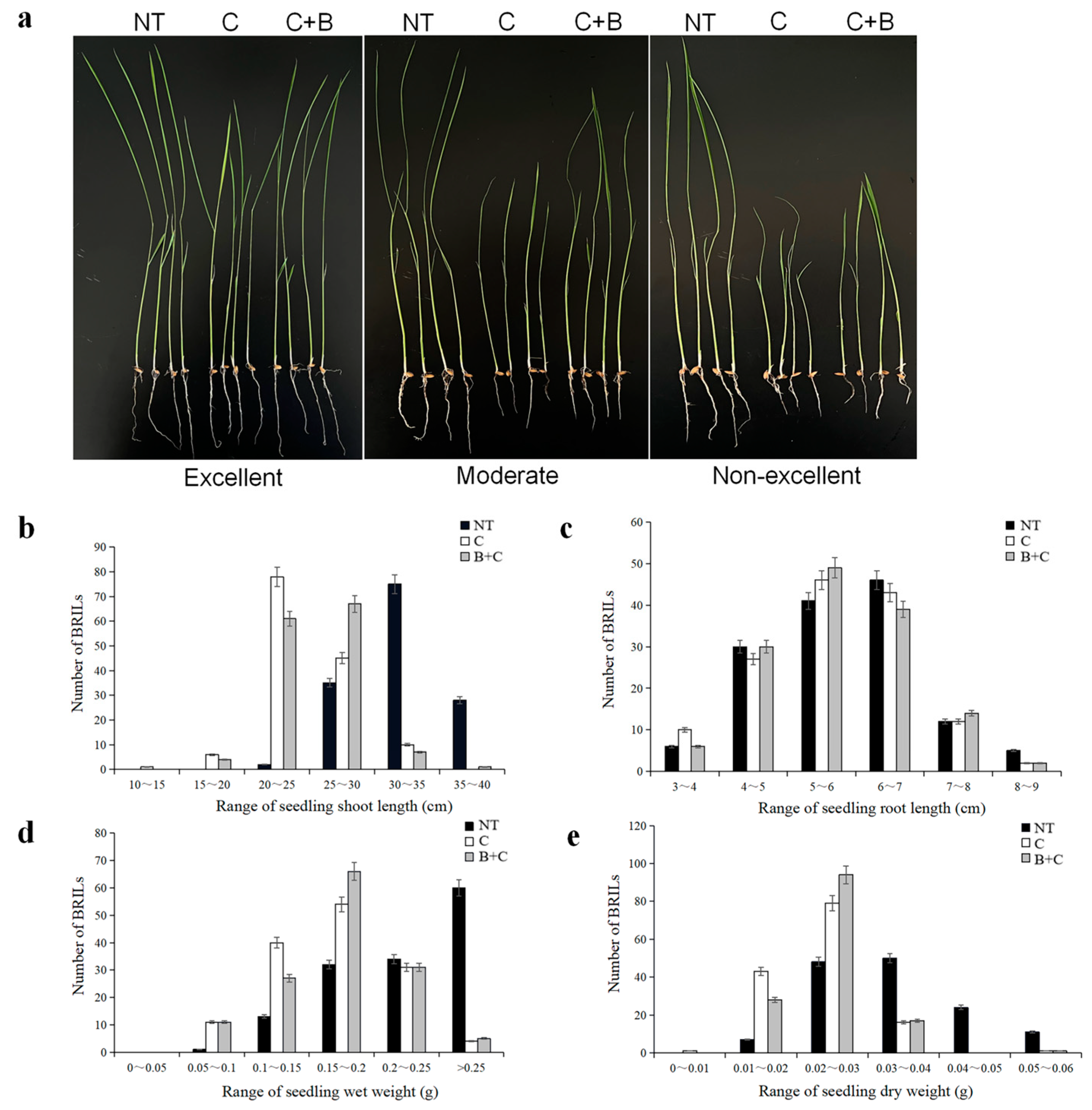
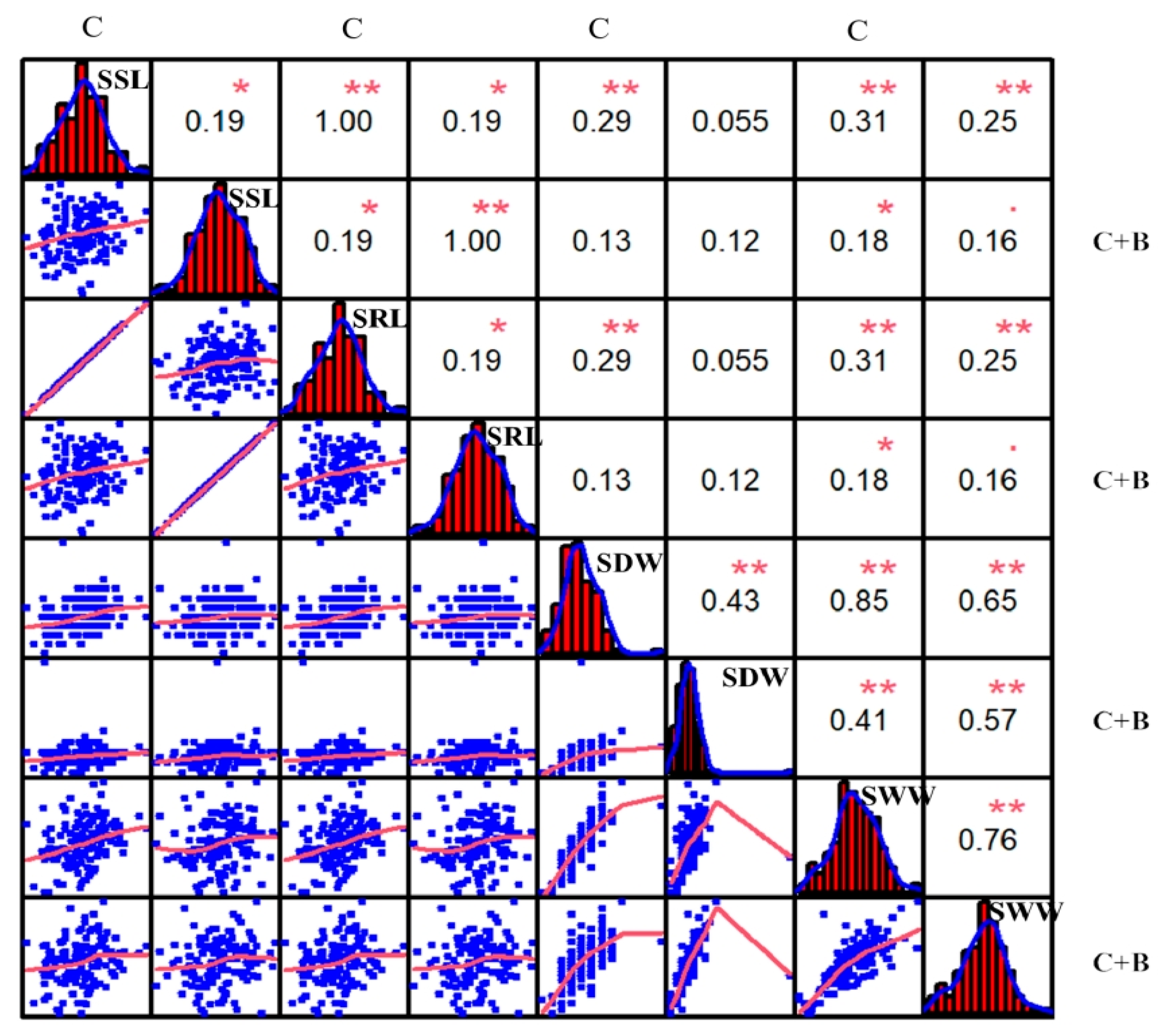
In the current study, a backcross recombinant inbred lines (BRIL) population of 140 individuals, derived from a cross between the DXWR and a super rice SN265, was constructed. Based on this BRIL, a high-resolution genome-wide SNP genetic map was developed using the GBS technique. Furthermore, we identified some QTLs and candidate genes under cold stress and BR-combined cold treatment at the seedling stage. The results have an important significance for the discovery of the genes involved in cold tolerance and for understanding the molecular mechanism that regulates the cold tolerance related to the BR signaling pathway in rice.
3. Discussion
The rich genetic diversity is an important basis for improving the agronomic trait and stresses tolerance in plants. Previous studies regarding evolution and genetic divergence showed that cultivated rice was domesticated from wild rice species. However, during the domestication process with thousands of years of evolution and natural selection, the diversity of many morphological traits was reduced, and many gene resources were lost in cultivated rice [
14,
31,
32,
33]. As the relative ancestor of the cultivated rice, common wild rice has high diversity of desirable genes, which are very important genetic resources for improving the agronomic trait and stresses tolerance in cultivated rice [
34,
35,
36]. DXWR (
O. rufipogon) was discovered in Dongxiang County, Jiangxi Province, China, which has the northernmost distribution in all species of wild rice (28°14′ N). Since DWXR plays important roles in basic rice research and industrial development, it is known as “the panda of wild plants” [
17,
18,
37]. DXWR possesses abundant genetic resources associated with wide cross-compatibility, fertility restoration, cytoplasmic male sterility, high grain yield, and resistance to a range of biotic and abiotic stresses, especially low-temperature tolerance.
Although wild rice and cultivated rice have the same genome type, they exhibit many differences in their genome sequences. If the temporary genetic populations, such as F
2 or BC
1 populations and the permanent primary genetic populations, such as recombinant inbred line were constructed and used for QTL mapping, they could show not only a low QTL detection efficiency, but also poor stability. Therefore, a stable and reliable mapping population is essential for utilizing wild rice [
32,
33,
38]. To date, multiple types of mapping populations have been constructed using DXWR as paternal parent, while a large number of QTLs associated with cold tolerance at the germination, seedling, booting, and flowering stages have been mapped to all chromosomes [
16,
17,
18,
19,
30]. In this study, the female parent SN265 was used to backcross for an additional four times after a cross between the female parent SN265 and female parent DXWR (BC
4F
1). Furthermore, the BC
4F
1 population was self-crossed for eight times to obtain advanced BRIL populations (BC
4F
8). The back- and self-crossing for multiple generations reduced the genetic differences caused by different genetic bases between cultivated rice and wild rice, which made the result of QTL mapping more stable and reliable.
It is well known that the low-temperature tolerance of plants is a complicated quantitative trait controlled by polygenes. To cope with low-temperature stress, plants modify their physiology, metabolism, and growth by means of signal transduction and expression regulation of many genes associated with cold tolerance when plants are exposed to low-temperature stress [
5]. A substantial number of genes that facilitate cold signaling and control the expression of cold regulons have been identified in many plants. A cascade signaling pathway, ICE–CBF–COR, is the most intensively studied, and is thought to be pretty important for cold tolerance in plants. This pathway contains the core components CBF/
DREB (C-repeat binding factors/dehydration-responsive element-binding proteins) transcriptional factor, ICE (inducer of CBF expression) activated factor, and diverse downstream functional proteins called cold-regulated (COR) proteins. The CBFs belong to the AP2/ERF (apetala 2/ethylene response factor) family of transcription factors, which are the most important in the ICE–CBF–COR pathway. The CBFs control the expression of
COR genes in response to cold stress, while the ICE acts as a positive upstream regulator of
CBF genes [
39,
40,
41].
In rice, 10
CBF/DREB homologous genes (
OsDREB1A-OsDREB1J) have been identified. Among them,
OsDREB1A,
OsDREB1B,
OsDREB1D,
OsDREB1F, and
OsDREB1G enhanced cold tolerance in Arabidopsis or rice [
42,
43]. In the case of QTL mapping studies, Os
DREB1G and
OsDREB1J were identified as the candidate genes related to cold tolerance for three QTLs,
qLOP2,
qPSR2-
1 and
qSR8-3, using the DXWR populations as the donor parent, respectively [
16,
30]. In addition, an ICE1-like gene,
OrbHLH001, was isolated from DXWR, which enhanced cold tolerance when expressed in transgenic Arabidopsis [
44]. Apart from these core components, many other activators or repressors are related to this pathway that directly or indirectly affect the low-temperature tolerance of plants. A MYB transcriptional factor, MYB15, interacted with ICE1 and was bound to MYB recognition sequences in the promoters of CBF genes, which repressed the expression of CBF genes. ICE1 activated the transcription of CBF3 by binding to MYC recognition elements in the promoter [
39,
45].
OsbHLH002, a homolog of ICE1 in rice, positively regulated cold tolerance by promoting the expression of OsTPP1, which encoded a key enzyme for trehalose biosynthesis [
46]. Two F-box proteins, EIN3-binding F-box 1/2 (EBF1/2), positively regulated the expression of
CBF genes by regulating the degradation of EIN3 and PIF3 [
47,
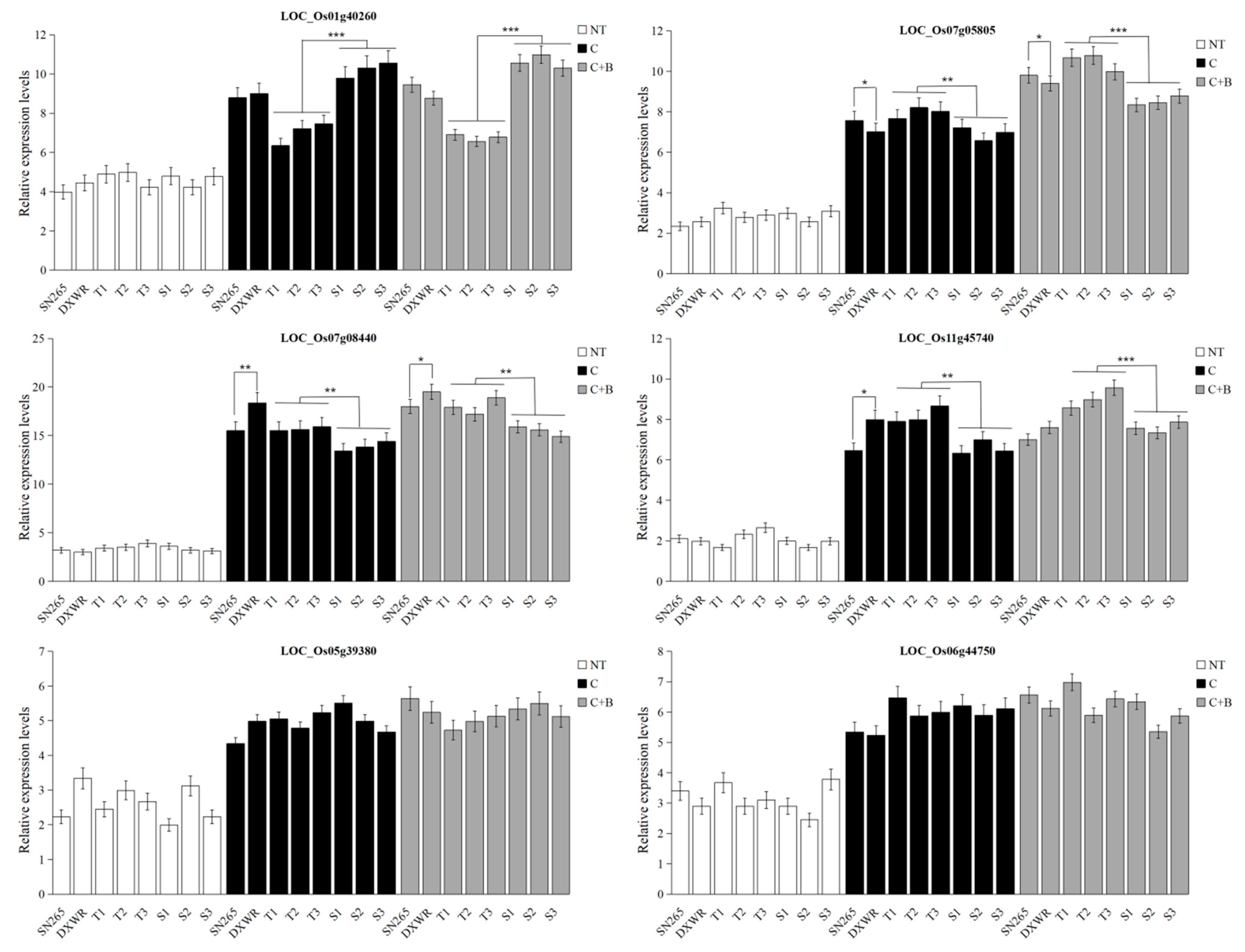
48]. In the current study, we identified some genes similar to those described above, such as bHLH transcription factor (LOC_Os07g08440), F-box and another domain-containing protein (LOC_Os09g27660 and LOC_Os10g04590), MYB family transcription factor (LOC_Os11g45740), and AP2 domain-containing protein (LOC_Os06g44750). The transcript levels of these genes except for LOC_Os09g27660 and LOC_Os10g04590 (F-box) were upregulated both under cold stress and BR-combined cold treatment. In the representative BRILs, LOC_Os07g08440 (bHLH), LOC_Os11g45740 (MYB), and LOC_Os06g44750 (AP2) showed higher transcript levels in the excellent BRILs than the non-excellent BRILs under cold stress and BR-combined cold treatment.
Previous studies have shown that the exogenous application of BR can improve low-temperature tolerance at different growth stages in various plants, including rice [
27,
28], maize [
49], wheat [
50], winter rye [
51], and Arabidopsis [
24,
26], etc. To date, the molecular mechanisms regarding both cold stress and BR signaling pathway were only intensively studied in Arabidopsis. Moreover, the cold tolerance in BR signaling pathway was involved in ICE–CBF–COR signaling pathway. Under cold stress, BR directed a bHLH transcription factor CESTA (CES) to regulate the expression of the CBF and downstream COR genes in Arabidopsis [
45]. BZR1 was a very important transcription factor in BR signaling pathway, acting upstream of CBF1 and CBF2 to directly enhance low-temperature tolerance in Arabidopsis. Interestingly, OsBZR1 (LOC_Os07g05805) was predicted for
qSSL7-1 under BR-combined cold treatment in the current study. The expression level of OsBZR1 was upregulated both under cold stress and BR-combined cold treatment, and it showed higher transcript levels in the excellent BRILs than the non-excellent BRILs under cold stress and BR-combined cold treatment. Moreover, BZR1 regulated other genes uncoupled with CBFs, such as WKRY6, PYL6, and SOC1, etc., to improve cold tolerance in Arabidopsis [
24,
26]. As a negative modulator in BR signaling pathway, the protein kinase BIN2 interacted with and phosphorylated ICE1 under cold stress, which facilitated the interaction between ICE1 and the E3 ubiquitin ligase HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE1 (HOS1), and thereby promoted ICE1 degradation. It was suggested that BIN2 mainly downregulated ICE1 abundance when the expression levels of
CBF genes were attenuated [
26]. In this study, OsWRKY77 (LOC_Os01g40260), a WRKY transcription factor, was also identified for qSSL1-2 under BR-combined cold treatment. It was shown that the expression level of
OsWRKY77 was upregulated in the sensitive genotype, suggesting its negative role in cold tolerance at the germination stage in rice [
52]. The expression levels of
OsWRKY77 were upregulated both under cold stress and BR-combined cold treatment, and its expression levels in the excellent BRILs were lower than the non-excellent BRILs under cold stress and BR-combined cold treatment. In addition to the transcription factors and genes related to cold tolerance and BR signaling pathway mentioned above, some other genes related to cold tolerance were identified, such as zinc finger domain containing protein, auxin-induced protein, oxygen evolving enhancer protein, heat shock protein, etc. The precise function and regulatory mechanism of these genes still needs further investigation.
In conclusion, many QTLs and candidate genes related to cold tolerance or BR pathway were identified by mapping analysis and qRT-PCR. This study provided a basis for further mining the genes involved in low-temperature tolerance or BR signaling pathway and investigating the mechanism regulating low-temperature tolerance in rice. Further studies were needed to thoroughly investigate whether these genes are associated with both cold tolerance and BR signaling pathway in rice.







{kind=link}
{kind=link}
{kind=link}
{kind=link}