
Effect of CO2 Content in Air on the Activity of Carbonic Anhydrases in Cytoplasm, Chloroplasts, and Mitochondria and the Expression Level of Carbonic Anhydrase Genes of the α- and β-Families in Arabidopsis thaliana Leaves

Abstract
:1. Introduction
2. Results
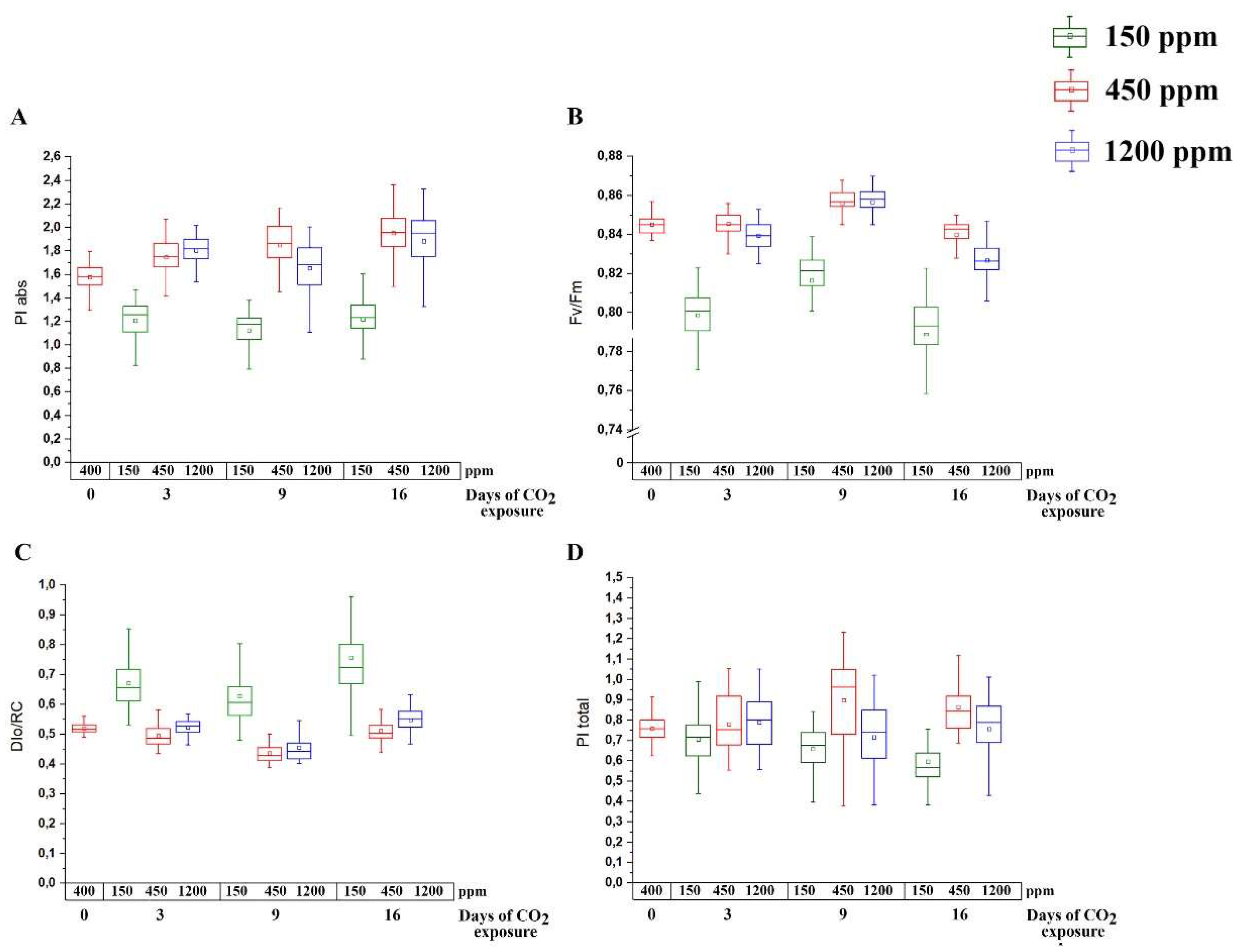
2.1. Photosynthetic and Stress State Characteristics of Arabidopsis Plants during Acclimation to Changed CO2 Conditions
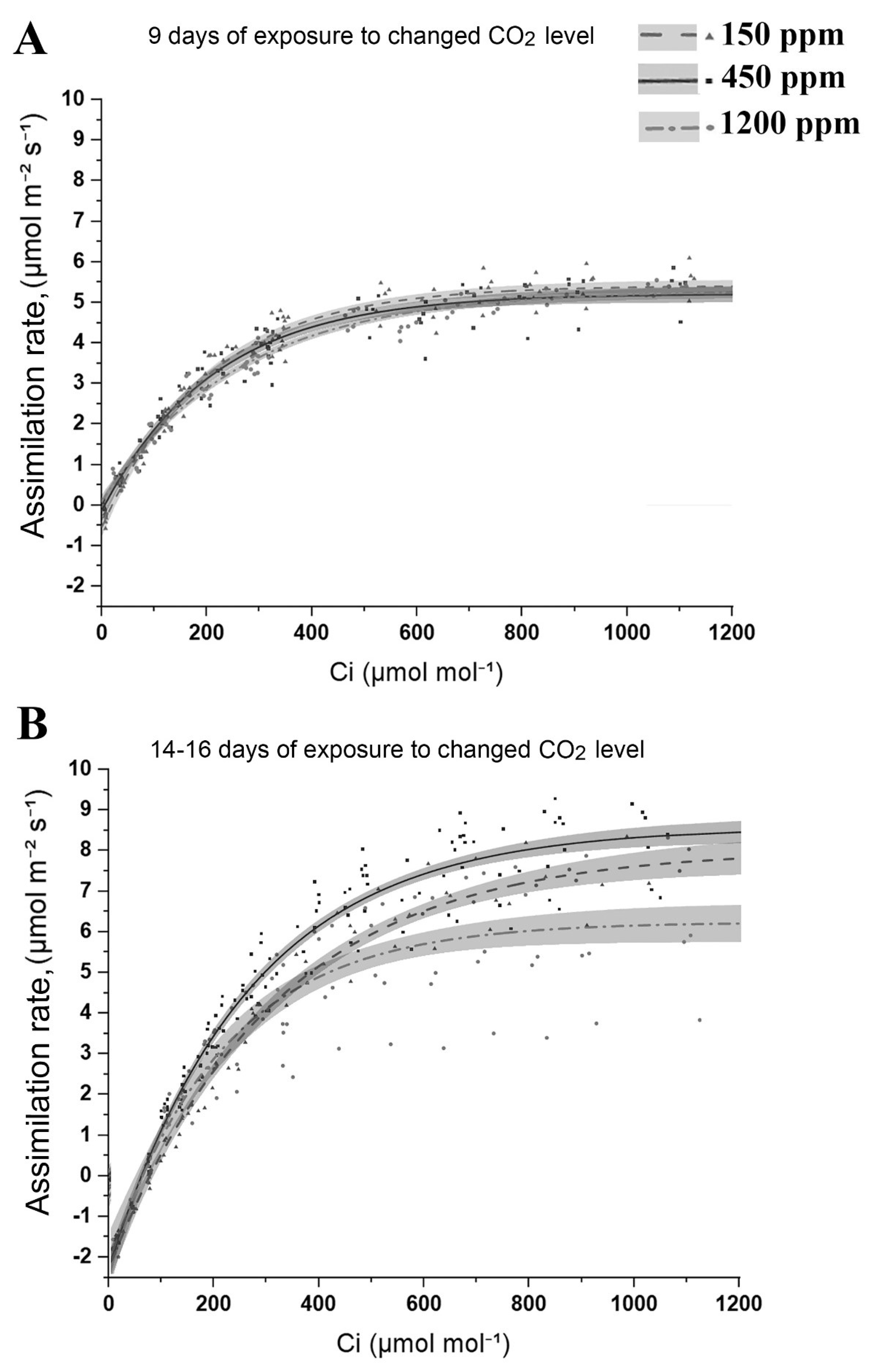
2.2. CO2 Assimilation and Carbon Levels in Leaves of Arabidopsis Plants Acclimated to Changed CO2 Content in the Air
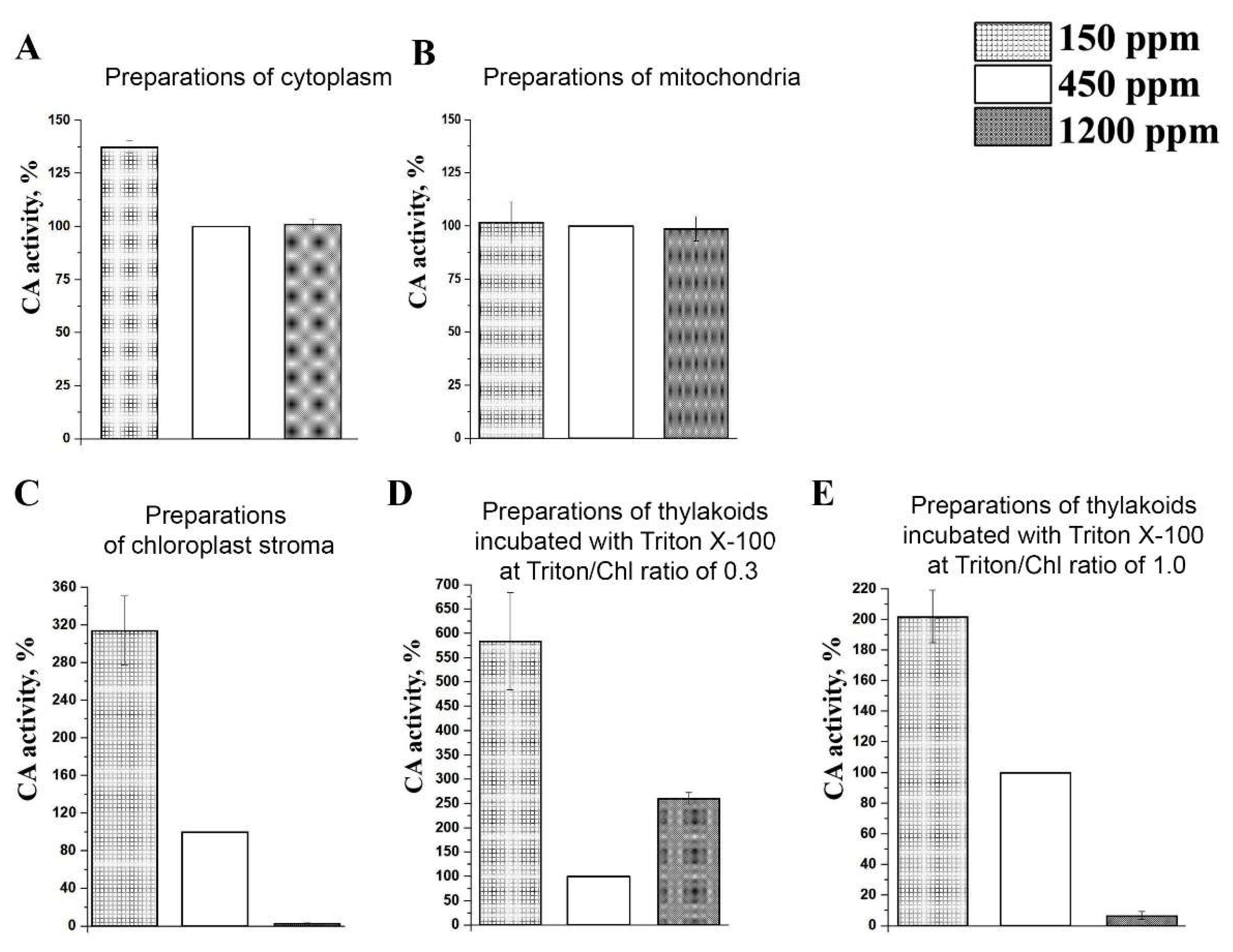
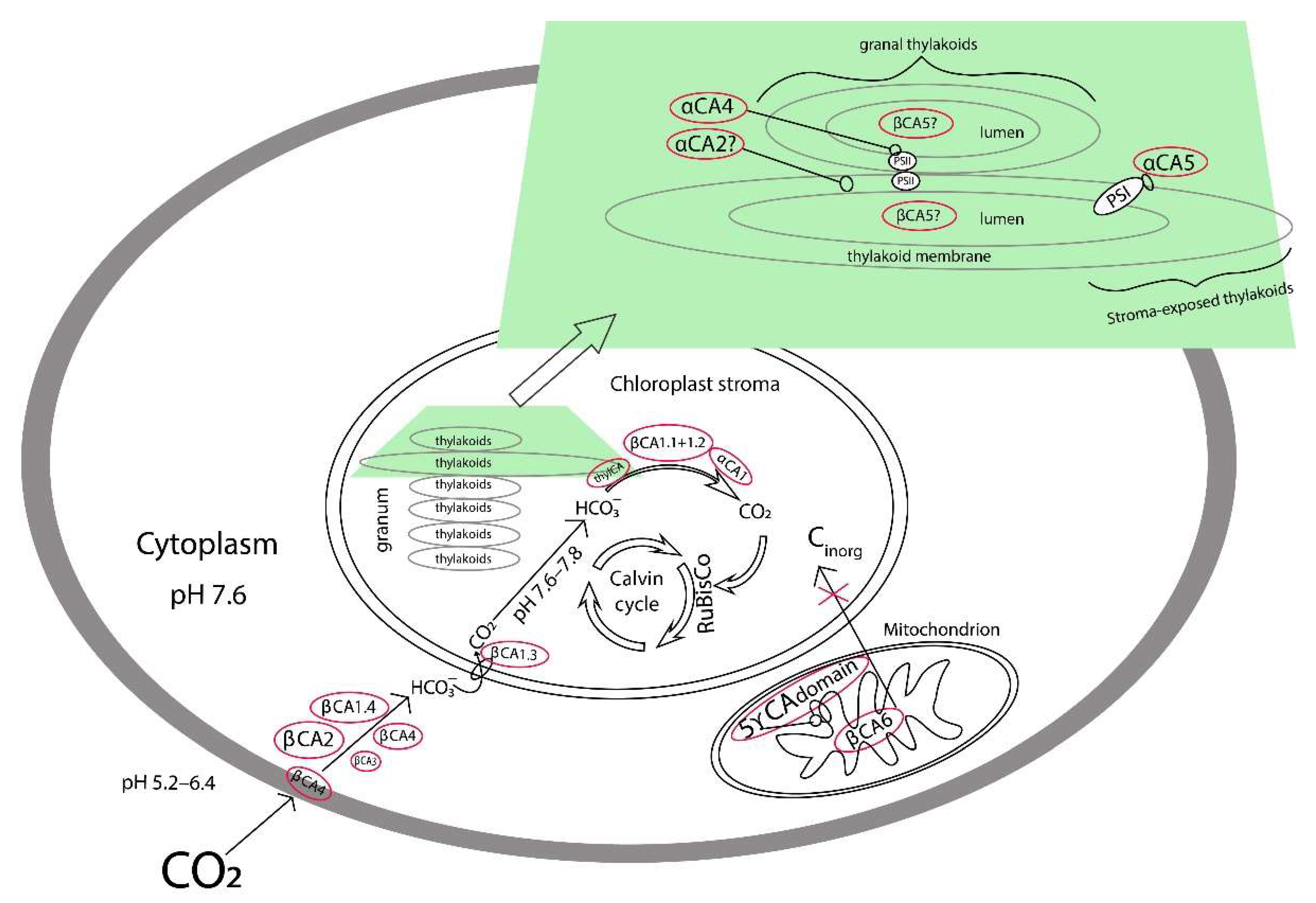
2.3. CA Activity in Preparations of Cytoplasm, Mitochondria, and Chloroplasts Isolated from Leaves of Plants Acclimated to Low, Normal, and High CO2 Level in the Air
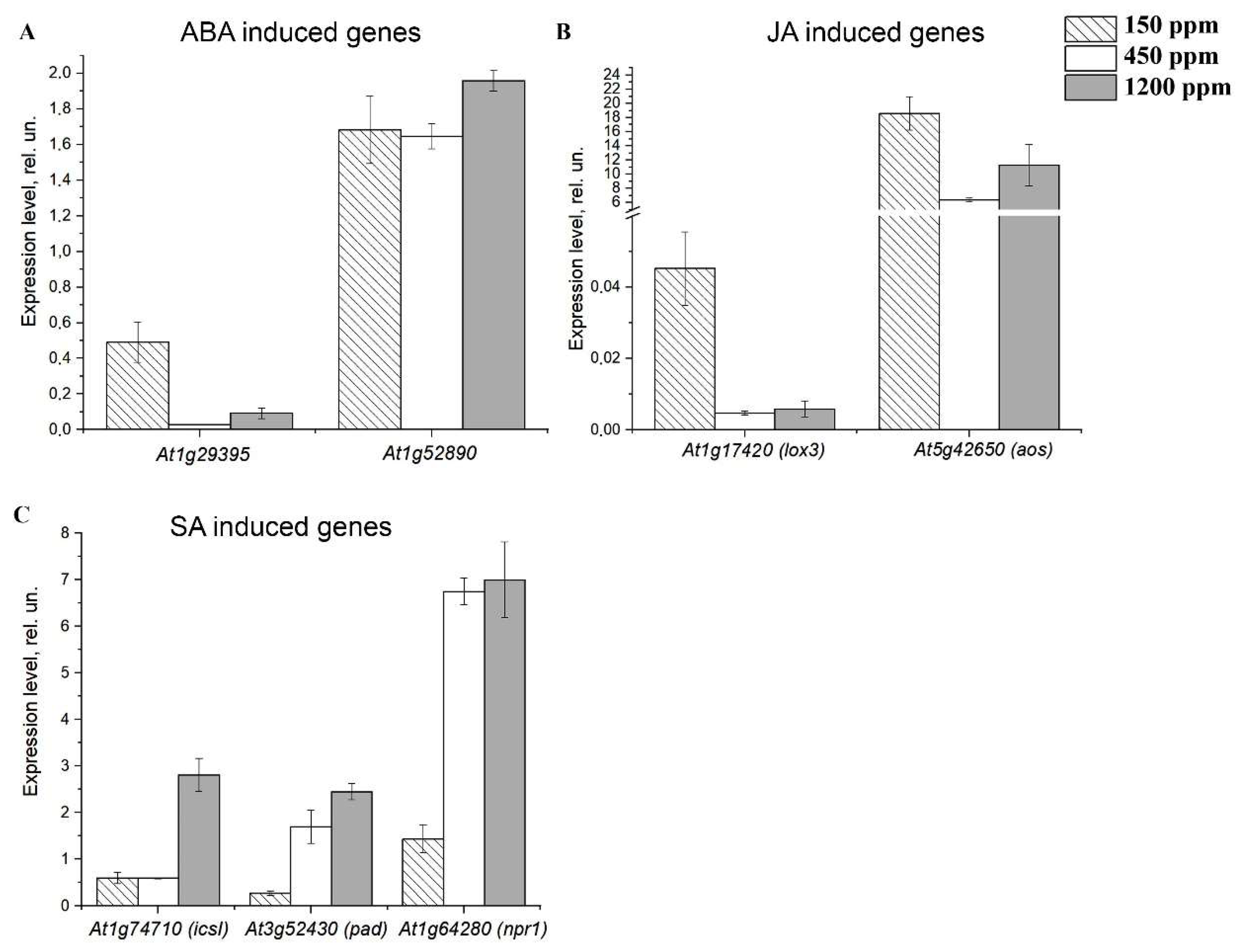
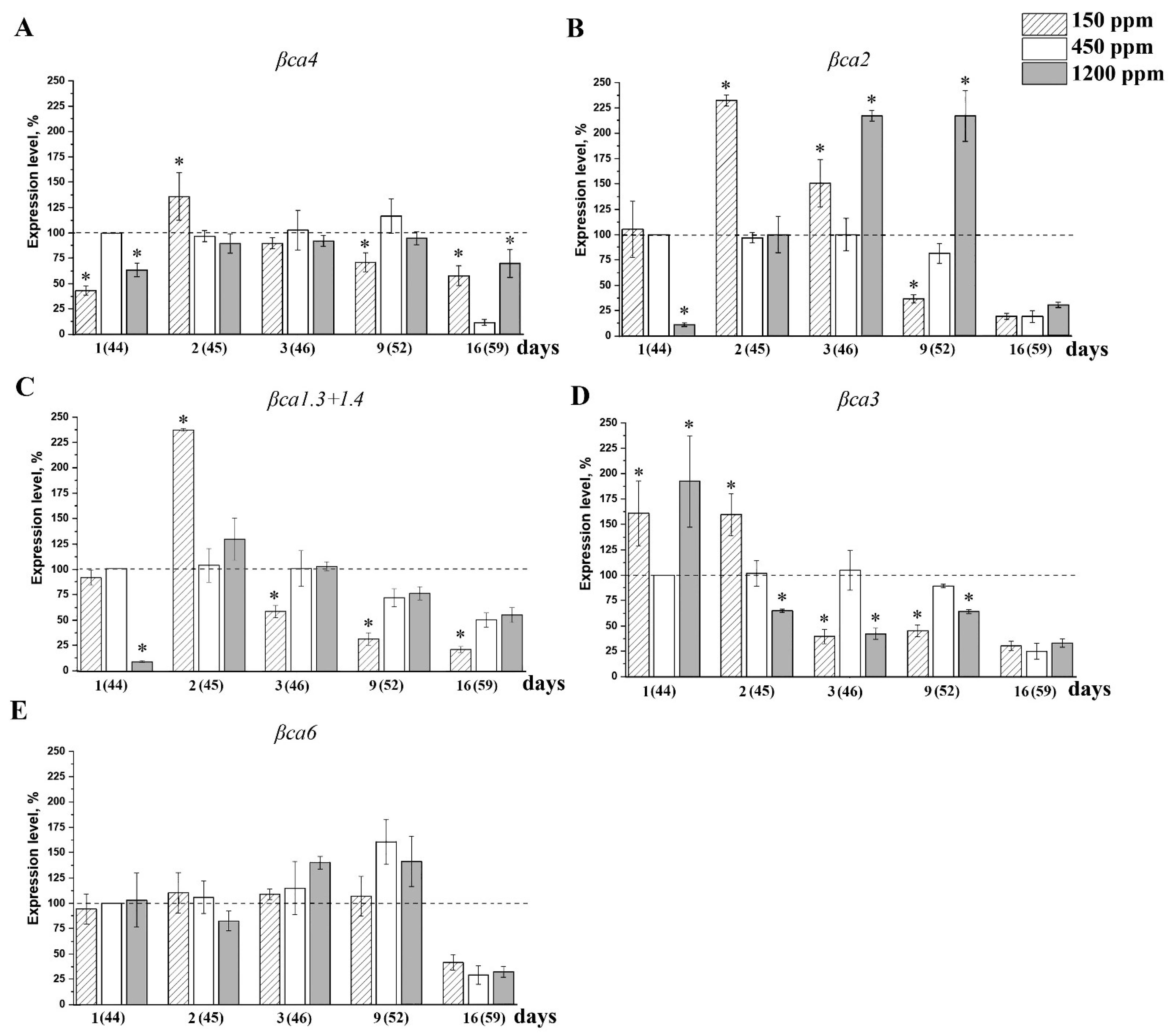
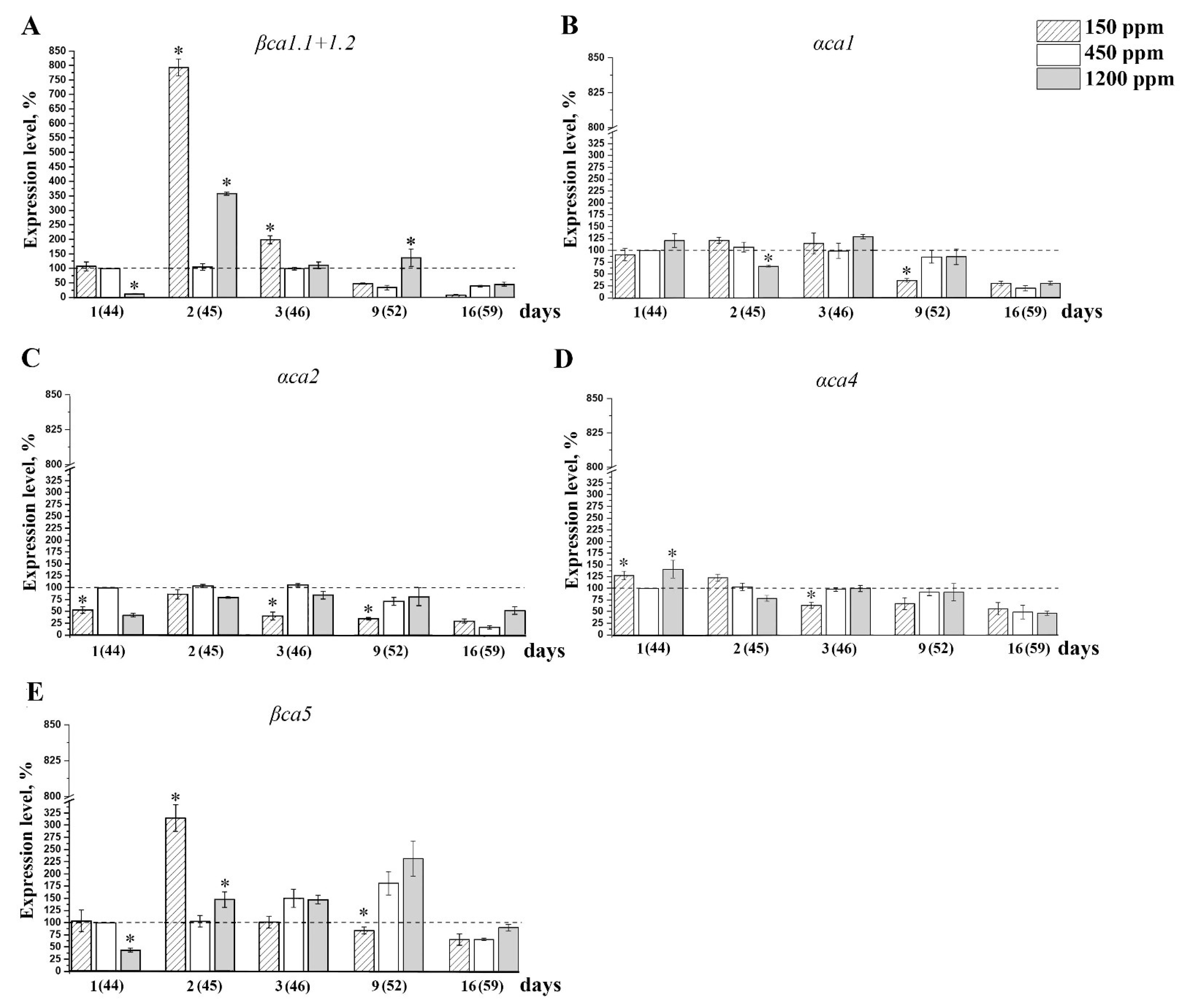
2.4. The Effect of Acclimation of Adult nCO2 Plants to Changed CO2 Content in the Air on the Expression Level of the Genes Encoding CAs of α- and β-Families
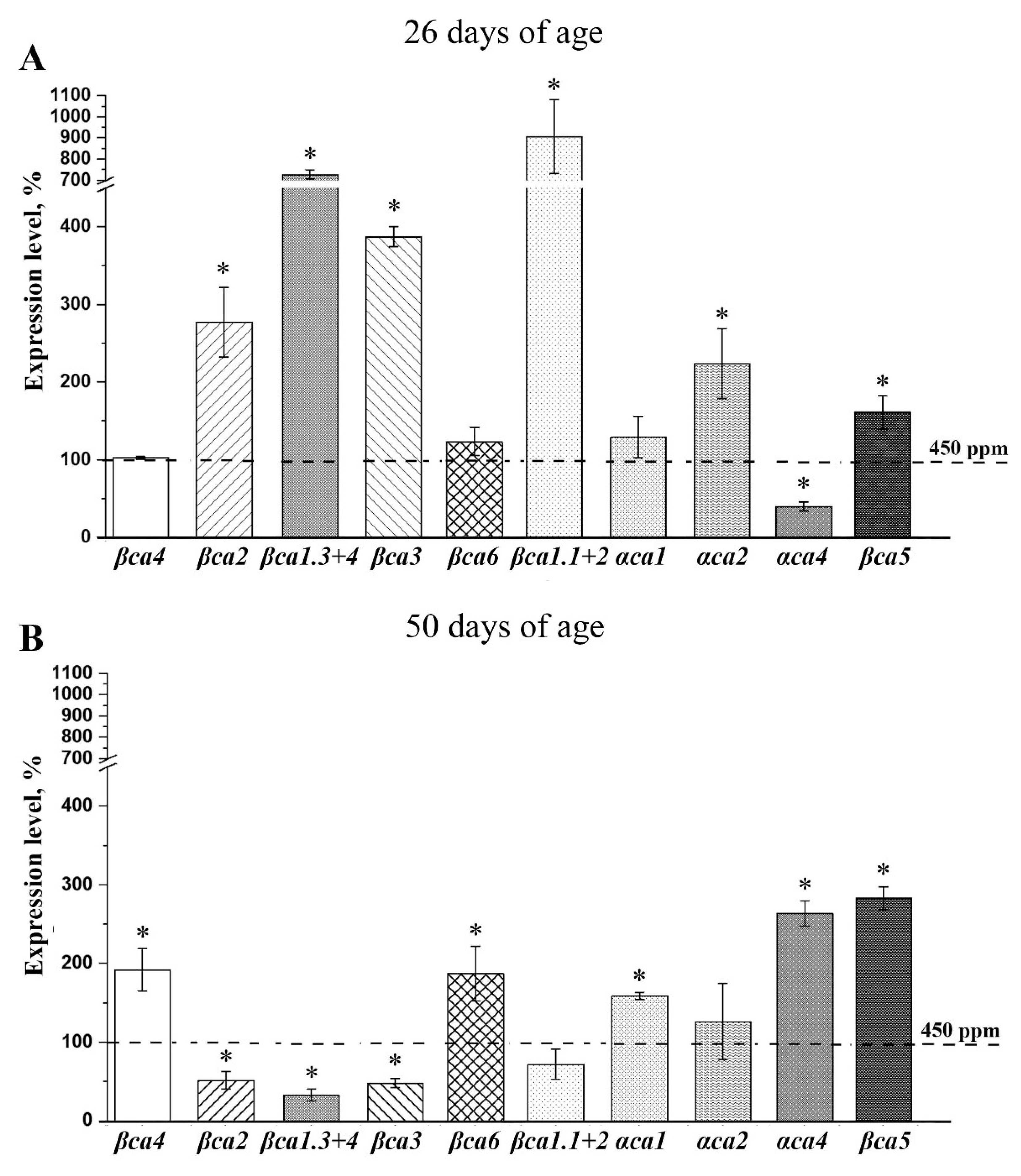
2.5. The effect of Acclimation of Young Plants to low CO2 Content in the Air on the Expression Level of the Genes Encoding CAs of α- and β-Families
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Measurement of Chlorophyll a Fluorescence
4.3. Measurement of CO2 Assimilation Rate
4.4. Determination of Starch and Soluble Carbohydrates Content
4.5. Isolation of Cell Preparations
4.6. Determination of the Protein Content
4.7. Determination of the Chlorophyll Content
4.8. Measurement of Carbonic Anhydrase Activity
4.9. Quantitative Reverse Transcription PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fabre, N.; Reiter, I.M.; Becuwe-Linka, N.; Genty, B.; Rumeau, D. Characterization and expression analysis of genes encoding alfa- and beta carbonic anhydrases in Arabidopsis. Plant Cell Environ. 2007, 30, 617–629. [Google Scholar] [CrossRef] [PubMed]
- DiMario, R.J.; Quebedeaux, J.C.; Longstreth, D.J.; Dassanayake, M.; Hartman, M.M.; Moroney, J.V. The cytoplasmic carbonic anhydrases βCA2 and βCA4 are required for optimal plant growth at low CO2. Plant Physiol. 2016, 171, 280293. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Li, Z.; Fu, Y.; Liang, J. Identification and molecular characterization of the alternative spliced variants of beta carbonic anhydrase 1 (βCA1) from Arabidopsis thaliana. PeerJ 2021, 9, e12673. [Google Scholar] [CrossRef] [PubMed]
- Soto, D.; Córdoba, J.P.; Villarreal, F.; Bartoli, C.; Schmitz, J.; Maurino, V.G.; Braun, H.P.; Pagnussat, G.C.; Zabaleta, E. Functional characterization of mutants affected in the carbonic anhydrase domain of the respiratory complex I in Arabidopsis thaliana. Plant J. 2015, 83, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Villarejo, A.; Burén, S.; Larsson, S.; Déjardin, A.; Monné, M.; Rudhe, C.; Karlsson, J.; Jansson, S.; Lerouge, P.; Rolland, N.; et al. Evidence for a protein transported through the secretory pathway enroute to the higher plant chloroplast. Nat. Cell Biol. 2005, 7, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.N.; Ignatova, L.K.; Ivanov, B.N. Multiple sources of carbonic anhydrase activity in pea thylakoids: Soluble and membrane-bound forms. Photosynth. Res. 2007, 91, 81–89. [Google Scholar] [CrossRef]
- Fedorchuk, T.; Rudenko, N.; Ignatova, L.; Ivanov, B. The presence of soluble carbonic anhydrase in the thylakoid lumen of chloroplasts from Arabidopsis leaves. J. Plant Physiol. 2014, 171, 903–906. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Ignatova, L.K.; Fedorchuk, T.P.; Ivanov, B.N. Carbonic anhydrases in photosynthetic cells of higher plants. Biochemistry 2015, 80, 798–813. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Ignatova, L.K.; Nadeeva-Zhurikova, E.M.; Fedorchuk, T.P.; Ivanov, B.N.; Borisova-Mubarakshina, M.M. Advances in understanding the physiological role and locations of carbonic anhydrases in C3 plant cells. Protoplasma 2021, 258, 249–262. [Google Scholar] [CrossRef]
- Friso, G.; Giacomelli, L.; Ytterberg, A.J.; Peltier, J.B.; Rudella, A.; Sun, Q.; Wijk, K.J. In-depth analysis of the thylakoid membrane proteome of Arabidopsis thaliana chloroplasts: New proteins, new functions, and a plastid proteome database. Plant Cell 2004, 16, 478–499. [Google Scholar] [CrossRef]
- Ignatova, L.; Zhurikova, E.; Ivanov, B. The presence of the low molecular mass carbonic anhydrase in photosystem II of C3 higher plants. J. Plant Physiol. 2019, 232, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Zhurikova, E.M.; Ignatova, L.K.; Rudenko, N.N.; Mudrik, V.A.; Vetoshkina, D.V.; Ivanov, B.N. Participation of two carbonic anhydrases of the alpha family in photosynthetic reactions in Arabidopsis thaliana. Biochemistry 2016, 81, 1182–1187. [Google Scholar] [CrossRef]
- Fedorchuk, T.P.; Kireeva, I.A.; Opanasenko, V.K.; Terentyev, V.V.; Rudenko, N.N.; Borisova-Mubarakshina, M.M.; Ivanov, B.N. Alpha carbonic anhydrase 5 mediates stimulation of ATP synthesis by bicarbonate in isolated Arabidopsis thylakoids. Front. Plant Sci. 2021, 12, 662082. [Google Scholar] [CrossRef]
- Wang, C.; Hu, H.; Qin, X.; Zeise, B.; Xu, D.; Rappel, W.J.; Boron, W.F.; Schroeder, J.I. Reconstitution of CO2 regulation of SLAC1 anion channel and function of CO2 Permeable PIP2;1 Aquaporin as Carbonic anhydrase 4 interactor. Plant Cell 2016, 28, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Boisson-Dernier, A.; Israelsson-Nordström, M.; Böhmer, M.; Xue, S.; Ries, A.; Godoski, J.; Kuhn, J.M.; Schroeder, J.I. Carbonic anhydrases are upstream regulators of CO2-controlled stomatal movements in guard cells. Nat. Cell Biol. 2010, 12, 87–93. [Google Scholar] [CrossRef]
- Coleman, J.R. Carbonic anhydrase and its role in photosynthesis. In Photosynthesis: Physiology and Metabolism; Leegood, R.C., Sharkey, T.D., Caemmerer, S., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 353–367. [Google Scholar]
- Jebanathirajah, J.A.; Coleman, J.R. Association of carbonic anhydrase with a Calvin cycle enzyme complex in Nicotiana tabacum. Planta 1998, 204, 177–182. [Google Scholar] [CrossRef]
- Anderson, L.; Carol, A. Enzyme co-localization with rubisco in pea leaf chloroplasts. Photosynth. Res. 2004, 82, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Majeau, N.; Coleman, J.R. Correlation of Carbonic Anhydrase and Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase Expression in Pea. Plant Physiol. 1994, 104, 1393–1399. [Google Scholar] [CrossRef]
- Lazova, G.N.; Stemler, A.J. A 160 kDa protein with carbonic anhydrase activity is complexed with rubisco on the outer surface of thylakoids. Cell Biol. Int. 2008, 32, 646–653. [Google Scholar] [CrossRef]
- Wang, L.; Jin, X.; Li, Q.; Wang, X.; Li, Z.; Wu, X. Comparative proteomics reveals that phosphorylation of β carbonic anhydrase 1 might be important for adaptation to drought stress in Brassica napus. Sci. Rep. 2016, 6, 39024. [Google Scholar] [CrossRef]
- Price, G.D.; Von Caemmerer, S.; Evans, J.R.; Yu, J.-W.; Lloyd, J.; Oja, V.; Kell, P.; Harrison, K.; Gallagher, A.; Badger, M.R. Specific reduction of chloroplast carbonic anhydrase activity by antisense RNA in transgenic tobacco plants has a minor effect on photosynthetic CO2 assimilation. Planta 1994, 193, 331–340. [Google Scholar] [CrossRef]
- Ivanov, B.N.; Ignatova, L.K.; Romanova, A.K. Diversity in forms and functions of carbonic anhydrase in terrestrial higher plants. Russ. J. Plant Physiol. 2007, 54, 143–162. [Google Scholar] [CrossRef]
- Suss, K.H.; Arkona, C.; Manteuffel, R.; Adler, K. Calvin cycle multienzyme complexes are bound to chloroplast thylakoid membranes of higher plants in situ. Proc. Natl. Acad. Sci. USA 1993, 90, 5514–5518. [Google Scholar] [CrossRef]
- Anderson, L.E.; Gibbons, J.T.; Wang, X. Distribution of ten enzymes of carbon metabolism in pea (Pisum sativum) chloroplasts. Int. J. Plant Sci. 1996, 157, 525–538. [Google Scholar] [CrossRef]
- Wydrzynski, T. Govindjee A new site of bicarbonate effect in Photosystem II of photosynthesis: Evidence from chlorophyll fluorescence transients in spinach chloroplasts. Biochim. Biophys. Acta Bioenerg. 1975, 387, 403–408. [Google Scholar] [CrossRef]
- Van Rensen, J.J.S.; Tonk, W.J.M.; de Bruijn, S.M. Involvement of bicarbonate in the protonation of the secondary quinone electron acceptor of photosystem II via the non-haem iron of the quinone-iron acceptor complex. FEBS Lett. 1988, 226, 347–351. [Google Scholar] [CrossRef]
- Klimov, V.V.; Hulsebosch, R.J.; Allakhverdiev, S.I.; Wincencjusz, H.; Van Gorkom, H.J.; Hoff, A.J. Bicarbonate may be required for ligation of manganese in the oxygen-evolving complex of photosystem II. Biochemistry 1997, 36, 16277–16281. [Google Scholar] [CrossRef]
- Klimov, V.; Baranov, S. Bicarbonate requirement for the water-oxidizing complex of photosystem II. Biochim. Biophys. Acta Bioenerg. 2001, 1503, 187–196. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Fedorchuk, T.P.; Vetoshkina, D.V.; Zhurikova, E.M.; Ignatova, L.K.; Ivanov, B.N. Influence of knockout of At4g20990 gene encoding α-CA4 on photosystem II light-harvesting antenna in plants grown under different light intensities and day lengths. Protoplasma 2018, 255, 69–78. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Fedorchuk, T.P.; Terentyev, V.V.; Dymova, O.V.; Naydov, I.A.; Golovko, T.K.; Borisova-Mubarakshina, M.M.; Ivanov, B.N. The role of carbonic anhydrase α-CA4 in the adaptive reactions of photosynthetic apparatus: The study with α-CA4 knockout plants. Protoplasma 2020, 257, 489–499. [Google Scholar] [CrossRef]
- Zhurikova, E.; Ignatova, L.; Semenova, G.; Rudenko, N.; Mudrik, V.; Ivanov, B. Effect of knockout of α-carbonic anhydrase 4 gene on photosynthetic characteristics and starch accumulation in leaves of Arabidopsis thaliana. Russ. J. Plant Physiol. 2015, 62, 564–569. [Google Scholar] [CrossRef]
- Villarejo, A.; Shutova, T.; Moskvin, O.; Forssen, M.; Klimov, V.V.; Samuelsson, G. A photosystem II-associated carbonic anhydrase regulates the efficiency of photosynthetic oxygen evolution. EMBO J. 2002, 21, 1930–1938. [Google Scholar] [CrossRef]
- Slaymaker, D.; Navarre, D.; Clark, D.; del Pozo, O.; Martin, G.; Klessig, D. The tobacco salicylic acid binding protein 3 (SABP3) is the chloroplast carbonic anhydrase which exhibits antioxidant activity and plays a role in the hypersensitive defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 11640–11645. [Google Scholar] [CrossRef]
- Restrepo, S.; Myers, K.L.; del Pozo, O.; Martin, G.B.; Hart, A.L.; Buell, C.R.; Fry, W.E.; Smart, C.D. Gene Profiling of a Compatible Interaction Between Phytophthora infestans and Solanum tuberosum Suggests a Role for Carbonic Anhydrase. Mol. Plant-Microbe Interact. 2005, 18, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Medina-Puche, L.; Castelló, M.; Canet, J.; Lamilla, J.; Colombo, M.; Tornero, P. β-Carbonic anhydrases play a role in salicylic acid perception in Arabidopsis. PLoS ONE 2017, 12, e0181820. [Google Scholar] [CrossRef] [PubMed]
- Hines, K.M.; Chaudhari, V.; Edgeworth, K.; Owens, T.; Hanson, M.R. Absence of carbonic anhydrase in chloroplasts affects C 3 plant development but not photosynthesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2107425118. [Google Scholar] [CrossRef]
- Dąbrowska-Bronk, J.; Komar, D.N.; Rusaczonek, A.; Kozłowska-Makulska, A.; Szechyńska-Hebda, M.; Karpiński, S. β-carbonic anhydrases and carbonic ions uptake positively influence Arabidopsis photosynthesis, oxidative stress tolerance and growth in light dependent manner. J. Plant Physiol. 2016, 203, 44–54. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, F.; Hao, L.; Yu, J.; Guo, L.; Zhou, H.; Ma, C.; Zhang, X.; Xu, M. Elevated CO2 concentration induces photosynthetic down-regulation with changes in leaf structure, non-structural carbohydrates and nitrogen content of soybean. BMC Plant Biol. 2019, 19, 255–273. [Google Scholar] [CrossRef]
- Moskvin, O.V.; Ignatova, L.K.; Ovchinnikova, V.I.; Ivanov, B.N. Membrane-Bound Carbonic Anhydrase in Pea Thylakoids. Biochemistry 1995, 60, 859–864. [Google Scholar]
- Ignatova, L.K.; Rudenko, N.N.; Mudrik, V.A.; Fedorchuk, T.P.; Ivanov, B.N. Carbonic anhydrase activity in Arabidopsis thaliana thylakoid membrane and fragments enriched with PSI or PSII. Photosynth. Res. 2011, 110, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.N.; Ignatova, L.K.; Kamornitskaya, V.B.; Ivanov, B.N. Pea leaf thylakoids contain several carbonic anhydrases. Dokl. Biochem. Biophys. 2006, 408, 155–157. [Google Scholar] [CrossRef]
- Ignatova, L.K.; Ivanov, B.N. Carbonic anhydrases of higher plant thylakoids and their participation in photosynthesis. In Handbook of Photosynthesis; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 193–200. [Google Scholar] [CrossRef]
- Ignatova, L.; Rudenko, N.; Zhurikova, E.; Borisova-Mubarakshina, M.; Ivanov, B. Carbonic Anhydrases in photosynthesizing cells of c3 higher plants. Metabolites 2019, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.N.; Ivanov, B.N. Unsolved problems of carbonic anhydrases functioning in photosynthetic cells of higher C3 plants. Biochemistry 2021, 86, 1243–1255. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.L.; Graham, D. Carbonic anhydrase in plants: Distribution, properties and possible physiological roles. In Progress in Phytochemistry; Reinhold, L., Harborne, J.B., Swain, T., Eds.; Pergammon Press: Oxford, UK, 1981; pp. 47–94. [Google Scholar]
- Wang, M.; Zhang, Q.; Liu, F.C.; Xie, W.F.; Wang, G.D.; Wang, J.; Gao, Q.H.; Duan, K. Family wide expression characterization of Arabidopsis beta carbonic anhydrase genes using qRT PCR and promoter::GUS fusions. Biochimie 2014, 97, 219–227. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Vetoshkina, D.V.; Fedorchuk, T.P.; Ivanov, B.N. Effect of light intensity under different photoperiods on expression level of Carbonic Anhydrase genes of the α and βfamilies in Arabidopsis thaliana leaves. Biochemistry 2017, 82, 1025–1035. [Google Scholar] [CrossRef]
- Sage, R.F.; Coleman, J.R. Effects of low atmospheric CO2 on plants: More than a thing of the past. Trends Plant Sci. 2001, 6, 18–24. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.Z.; Schjoerring, J.K. Effects of elevated atmospheric CO2 on physiology and yield of wheat (Triticum aestivum L.): A meta-analytic test of current hypotheses. Agric. Ecosyst Environ. 2013, 178, 57–63. [Google Scholar] [CrossRef]
- Medlyn, B.E.; Barton, C.V.M.; Broadmeadow, M.S.J.; Ceulemans, R.; De Angelis, P.; Forstreuter, M.; Freeman, M.; Jackson, S.B.; Kellomäki, S.; Laitat, E.; et al. Stomatal conductance of forest species after long-term exposure to elevated CO2 concentration: A synthesis. New Phytol. 2001, 149, 247–264. [Google Scholar] [CrossRef]
- Porter, M.A.; Grodzinski, B. Acclimation to high CO2 in bean: Carbonic anhydrase and ribulose bisphosphate carboxylase. Plant Physiol. 1984, 74, 413–416. [Google Scholar] [CrossRef]
- Missner, A.; Pohl, P. 110 years of the Meyer-Overton rule: Predicting membrane permeability of gases and other small compounds. ChemPhysChem 2009, 10, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Boron, W.F.; Endeward, V.; Gros, G.; Musa-Aziz, R.; Pohl, P. Intrinsic CO2 permeability of cell membranes and potential biological relevance of CO2 channels. ChemPhysChem 2011, 12, 1017–1019. [Google Scholar] [CrossRef] [PubMed]
- Endeward, V.; Al-Samir, S.; Itel, F.; Gros, G. How does carbon dioxide permeate cell membranes? A discussion of concepts, results and methods. Front. Physiol. 2014, 4, 382. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Barbour, M.M.; Brendel, O.; Cabrera, H.M.; Carriquí, M.; DíazEspejo, A.; Douthe, C.; Dreyer, E.; Ferrio, J.P.; Gago, J.; et al. Mesophyll diffusion conductance to CO2: An unappreciated central player in photosynthesis. Plant Sci. 2012, 193–194, 70–84. [Google Scholar] [CrossRef]
- Sunderhaus, S.; Dudkina, N.V.; Jänsch, L.; Klodmann, J.; Heinemeyer, J.; Perales, M.; Zabaleta, E.; Boekema, E.J.; Braun, H.P. Carbonic anhydrase subunits form a matrix-exposed domain attached to the membrane arm of mitochondrial complex I in plants. J. Biol. Chem. 2006, 281, 6482–6488. [Google Scholar] [CrossRef]
- Zabaleta, E.; Martin, M.V.; Braun, H.P. A basal carbon concentrating mechanism in plants? Plant Sci. 2012, 187, 97–104. [Google Scholar] [CrossRef]
- Wickell, D.; Kuo, L.-Y.; Yang, H.-P.; Ashok, A.D.; Irisarri, I.; Dadras, A.; de Vries, S.; de Vries, J.; Huang, Y.-M.; Li, Z.; et al. Underwater CAM photosynthesis elucidated by Isoetes genome. Nat. Commun. 2021, 12, 6348–6360. [Google Scholar] [CrossRef]
- Buren, S. Targeting and Function of CAH1 Characterisation of a Novel Protein Pathway to the Plant Cell Chloroplast. Ph.D. Thesis, Umea University, Umeå, Sweden, 2010. [Google Scholar]
- Chaki, M.; Carreras, A.; López-Jaramillo, J.; Begara-Morales, J.C.; Sánchez-Calvo, B.; Valderrama, R.; Corpas, F.J.; Barroso, J.B. Tyrosine nitration provokes inhibition of sunflower carbonic anhydrase (β-CA) activity under high temperature stress. Nitric Oxide 2013, 29, 30–33. [Google Scholar] [CrossRef]
- Ferreira, F.; Guo, C.; Coleman, J. Reduction of plastid-localized carbonic anhydrase activity results in reduced Arabidopsis seedling survivorship. Plant Physiol. 2008, 147, 585–594. [Google Scholar] [CrossRef]
- Hoang, C.; Chapman, K. Biochemical and molecular inhibition of plastidial carbonic anhydrase reduces the incorporation of acetate into lipids in cotton embryos and tobacco cell suspensions and leaves. Plant Physiol. 2002, 128, 1417–1427. [Google Scholar] [CrossRef]
- Badger, M.; Pfanz, H. Effect of carbonic anhydrase inhibition on photosynthesis by leaf pieces of C3 and C4 plants. Funct. Plant Biol. 1995, 22, 45–49. [Google Scholar] [CrossRef]
- Ignatova, L.K.; Moskvin, O.V.; Ivanov, B.N. Effects of carbonic anhydrase inhibitors on proton exchange and photosynthesis in pea protoplasts. Russ. J. Plant Physiol. 2001, 48, 467–472. [Google Scholar] [CrossRef]
- Riazunnisa, K.; Padmavathi, L.; Bauwe, H.; Raghavendra, A.S. Markedly low requirement of added CO2 for photosynthesis by mesophyll protoplasts of pea (Pisum sativum): Possible roles of photorespiratory CO2 and carbonic anhydrase. Physiol. Plant. 2006, 128, 763–772. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Kaplan, F.; Zhao, W.; Richards, J.T.; Wheeler, R.M.; Guy, C.L.; Levine, L.H. Transcriptional and metabolic insights into the differential physiological responses of Arabidopsis to optimal and supraoptimal atmospheric CO2. PLoS ONE 2012, 7, e43583. [Google Scholar] [CrossRef]
- Du Bois, M.; Gilles, K.A.; Hamilton, J.K.; Roberts, R.A.; Smith, E. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Murcha, M.W.; Whelan, J. Isolation of intact mitochondria from the model plant species Arabidopsis thaliana and Oryza sativa. Methods Mol. Biol. 2015, 1305, 1–12. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Khristin, M.S.; Ignatova, L.K.; Rudenko, N.N.; Ivanov, B.N.; Klimov, V.V. Photosystem II associated carbonic anhydrase activity in higher plants is situated in core-complex. FEBS Lett. 2004, 577, 305–308. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CO2 Level in Air | Starch Content, mg/g of Fresh Weight | Soluble Carbohydrates Content, mg/g of Fresh Weight |
|---|---|---|
| 150 ppm | 0.82 ± 0.04 (23%) | 2.92 ± 0.35 (74%) |
| 450 ppm | 3.60 ± 0.34 (100%) | 3.95 ± 0.31 (100%) |
| 1200 ppm | 2.19 ± 0.22 (61%) | 4.25 ± 0.36 (108%) |
| Genes | Levels of Expression at | |||
|---|---|---|---|---|
| 26th Day of Age | 44th Day of Age | 50th Day of Age | 59–60th Days of Age | |
| βca4 | 1037.9 ± 12.46 | 60.6 ± 2.7 | 55.89 ± 3.97 | 7.22 ± 0.52 |
| βca2 | 1416.38 ± 12.35 | 195.67 ± 9.41 | 190.88 ± 15.41 | 37.90 ± 6.92 |
| βca1.3+1.4 | 1290.03 ± 7.46 | 153.08 ± 16.1 | 140.74 ± 12.30 | 76.54 ± 5.43 |
| βca3 | 1.52 ± 0.20 | 0.98 ± 0.04 | 0.80 ± 0.02 | 0.25 ± 0.07 |
| βca6 | 10.94 ± 0.09 | 7.99 ± 0.93 | 6.45 ± 0.32 | 2.94 ± 0.06 |
| βca1.1+1.2 | 24.34 ± 4.67 | 19.96 ± 0.96 | 17.91 ± 0.55 | 7.91 ± 0.46 |
| αca1 | 69.48 ± 5.71 | 16.43 ± 0.78 | 14.04 ± 1.44 | 3.31 ± 0.27 |
| αca2 | 3.8 ± 0.28 | 0.93 ± 0.05 | 0.71 ± 0.09 | 0.16 ± 0.05 |
| αca4 | 0.78 ± 0.03 | 0.28 ± 0.01 | 0.26 ± 0.04 | 0.14 ± 0.02 |
| βca5 | 14.8 ± 2.3 | 10.59 ± 0.13 | 10.3 ± 0.20 | 6.98 ± 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudenko, N.N.; Ignatova, L.K.; Naydov, I.A.; Novichkova, N.S.; Ivanov, B.N. Effect of CO2 Content in Air on the Activity of Carbonic Anhydrases in Cytoplasm, Chloroplasts, and Mitochondria and the Expression Level of Carbonic Anhydrase Genes of the α- and β-Families in Arabidopsis thaliana Leaves. Plants 2022, 11, 2113. https://doi.org/10.3390/plants11162113
Rudenko NN, Ignatova LK, Naydov IA, Novichkova NS, Ivanov BN. Effect of CO2 Content in Air on the Activity of Carbonic Anhydrases in Cytoplasm, Chloroplasts, and Mitochondria and the Expression Level of Carbonic Anhydrase Genes of the α- and β-Families in Arabidopsis thaliana Leaves. Plants. 2022; 11(16):2113. https://doi.org/10.3390/plants11162113
Chicago/Turabian StyleRudenko, Natalia N., Lyudmila K. Ignatova, Ilya A. Naydov, Natalia S. Novichkova, and Boris N. Ivanov. 2022. "Effect of CO2 Content in Air on the Activity of Carbonic Anhydrases in Cytoplasm, Chloroplasts, and Mitochondria and the Expression Level of Carbonic Anhydrase Genes of the α- and β-Families in Arabidopsis thaliana Leaves" Plants 11, no. 16: 2113. https://doi.org/10.3390/plants11162113