
The Role of Carbonic Anhydrase αCA4 in Photosynthetic Reactions in Arabidopsis thaliana Studied, Using the Cas9 and T-DNA Induced Mutations in Its Gene
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Analysis of Plants Carrying Site-Specific Mutations. Production of Homozygous Lines
2.2. Photosynthetic Characteristics of Arabidopsis Plants Lacking αCA4
2.3. Daily Dynamics in Starch Content in αCA4-KO
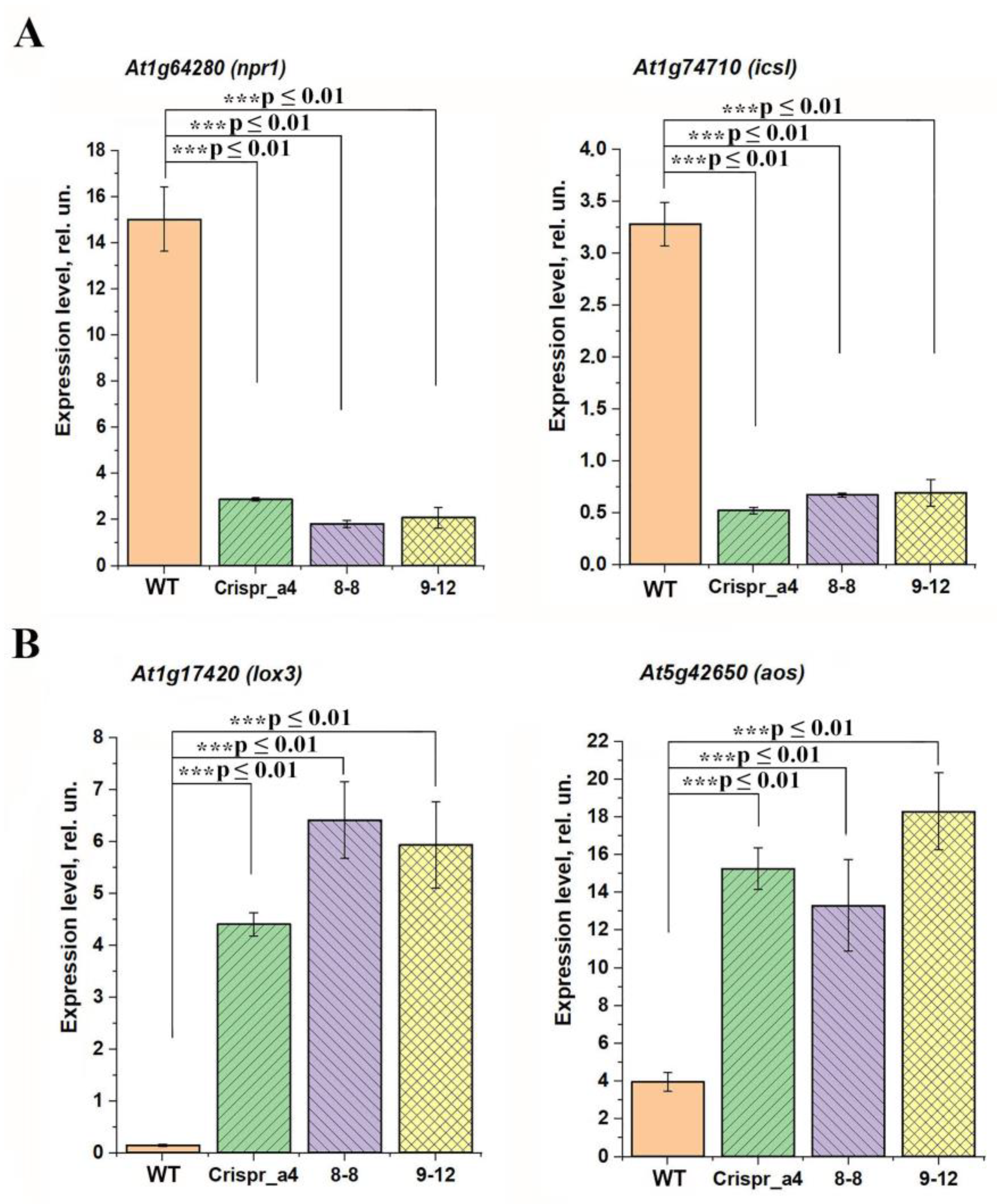
2.4. Effect of αCA4 Absence on the Expression Levels of the Cytoplasmic and Chloroplast Carbonic Anhydrases
2.5. Influence of the Knockout of αCA4 Gene on the Changes in the Content of Lhcb Encoding Genes Transcripts
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Plasmids for Gene Editing
4.3. Selection of gRNA
4.4. Creation of the Genetic Construct pDGE347-218 to Introduce Double-Strand Breaks in the αCA4 GENE Sequence
4.5. Transformation of A. thaliana
4.6. Analysis of Site-Specific Mutations in the αCA4 Gene
4.7. Determination of Starch Content
4.8. Measurement of Chlorophyll a Fluorescence at Room Temperature
4.9. Quantitative Reverse-Transcription PCR
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strisciuglio, P.; Sartorio, R.; Pecoraro, C.; Lotito, F.; Sly, W.S.; Pansini, V.S. Variable clinical presentation of carbonic anhydrase deficiency: Evidence for heterogeneity? Eur. J. Pediatr. 1990, 149, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Sly, W.S.; Hu, Y. Human carbonic anhydrases and carbonic anhydrase deficiencies. Annu. Rev. Biochem. 1995, 64, 375–401. [Google Scholar] [CrossRef] [PubMed]
- DiMario, R.J.; Machingura, M.C.; Waldrop, G.L.; Moroney, J.V. The many types of carbonic anhydrases in photosynthetic organisms. Plant Sci. 2018, 268, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Polishchuk, O.V. Stress-Related Changes in the Expression and Activity of Plant Carbonic Anhydrases. Planta 2021, 253, 58. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Ivanov, B.N. Unsolved Problems of Carbonic Anhydrases Functioning in Photosynthetic Cells of Higher C3 Plants. Biochemistry 2021, 86, 1243–1255. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.N.; Fedorchuk, T.P.; Terentyev, V.V.; Dymova, O.V.; Naydov, I.A.; Golovko, T.K.; Borisova-Mubarakshina, M.M.; Ivanov, B.N. The role of carbonic anhydrase α-CA4 in the adaptive reactions of photosynthetic apparatus: The study with α-CA4 knockout plants. Protoplasma 2020, 257, 489–499. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Fedorchuk, T.P.; Vetoshkina, D.V.; Zhurikova, E.M.; Ignatova, L.K.; Ivanov, B.N. Influence of knockout of At4g20990 gene encoding α-CA4 on photosystem II light-harvesting antenna in plants grown under different light intensities and day lengths. Protoplasma 2018, 255, 69–78. [Google Scholar] [CrossRef]
- Ignatova, L.K.; Rudenko, N.N.; Mudrik, V.A.; Fedorchuk, T.P.; Ivanov, B.N. Carbonic anhydrase activity in Arabidopsis thaliana thylakoid membrane and fragments enriched with PSI or PSII. Photosynth. Res. 2011, 110, 89–98. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Ignatova, L.K.; Ivanov, B.N. Multiple sources of carbonic anhydrase activity in pea thylakoids: Soluble and membrane-bound forms. Photosynth. Res. 2007, 91, 81–89. [Google Scholar] [CrossRef]
- Zhurikova, E.M.; Ignatova, L.K.; Rudenko, N.N.; Mudrik, V.A.; Vetoshkina, D.V.; Ivanov, B.N. Participation of two carbonic anhydrases of the alpha family in photosynthetic reactions in Arabidopsis thaliana. Biochemistry 2016, 81, 1182–1187. [Google Scholar] [CrossRef]
- Fedorchuk, T.P.; Kireeva, I.A.; Opanasenko, V.K.; Terentyev, V.V.; Rudenko, N.N.; Borisova-Mubarakshina, M.M.; Ivanov, B.N. Alpha Carbonic Anhydrase 5 Mediates Stimulation of ATP Synthesis by Bicarbonate in Isolated Arabidopsis Thylakoids. Front. Plant Sci. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Friso, G.; Giacomelli, L.; Ytterberg, A.J.; Peltier, J.B.; Rudella, A.; Sun, Q.; Van Wijk, K.J. In-Depth Analysis of the Thylakoid Membrane Proteome of Arabidopsis thaliana Chloroplasts: New Proteins, New Functions, and a Plastid Proteome Database. Plant Cell 2004, 16, 478–499. [Google Scholar] [CrossRef] [Green Version]
- Ignatova, L.; Zhurikova, E.; Ivanov, B. The presence of the low molecular mass carbonic anhydrase in photosystem II of C3 higher plants. J. Plant Physiol. 2019, 232, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Zhurikova, E.M.; Ignatova, L.K.; Semenova, G.A.; Rudenko, N.N.; Mudrik, V.A.; Ivanov, B.N. Effect of knockout of α-carbonic anhydrase 4 gene on photosynthetic characteristics and starch accumulation in leaves of Arabidopsis thaliana. Russ. J. Plant Physiol. 2015, 62, 564–569. [Google Scholar] [CrossRef]
- Shi, Y.; Ke, X.; Yang, X.; Liu, Y.; Hou, X. Plants response to light stress. J. Genet. Genom. 2022, 49, 735–747. [Google Scholar] [CrossRef]
- Li, X.-P.; Bjorkman, O.; Shih, C.; Grossman, A.R.; Rosenquist, M.; Jansson, S.; Niyogi, K.K. A pigment-binding protein essential for regulation of photosynthetic light harvesting. Nature 2000, 403, 391–395. [Google Scholar] [CrossRef]
- Ruban, A.V. Nonphotochemical chlorophyll fluorescence quenching: Mechanism and effectiveness in protecting plants from photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [Green Version]
- Nilkens, M.; Kress, E.; Lambrev, P.; Miloslavina, Y.; Müller, M.; Holzwarth, A.R.; Jahns, P. Identification of a slowly inducible zeaxanthin-dependent component of non-photochemical quenching of chlorophyll fluorescence generated under steady-state conditions in Arabidopsis. Biochim. Biophys. Acta (BBA)-Bioenerg. 2010, 1797, 466–475. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, N.N.; Vetoshkina, D.V.; Fedorchuk, T.P.; Ivanov, B.N. Effect of light intensity under different photoperiods on expression level of carbonic anhydrase genes of the α- and β-families in Arabidopsis thaliana leaves. Biochemistry 2017, 82, 1025–1035. [Google Scholar] [CrossRef]
- Shelake, R.M.; Kadam, U.S.; Kumar, R.; Pramanik, D.; Singh, A.K.; Kim, J.Y. Engineering drought and salinity tolerance traits in crops through CRISPR-mediated genome editing: Targets, tools, challenges, and perspectives. Plant Commun. 2022, 3, 100417. [Google Scholar] [CrossRef]
- Gelvin, S.B. Integration of Agrobacterium T-DNA into the Plant Genome. Annu. Rev. Genet. 2017, 51, 195–217. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Boisson-Dernier, A.; Israelsson-Nordström, M.; Xue, S.; Ries, A.; Godoski, J.; Kuhn, J.M.; Julian, I. Carbonic Anhydrases are Upstream Regulators in Guard Cells of CO2 -Controlled Stomatal Movements. Nat. Cell Biol. 2010, 12, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiMario, R.J.; Quebedeaux, J.C.; Longstreth, D.J.; Dassanayake, M.; Hartman, M.M.; Moroney, J.V. The cytoplasmic carbonic anhydrases βCA2 and βCA4 are required for optimal plant growth at low CO2. Plant Physiol. 2016, 171, 280–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, K.A. T-DNA insertion mutagenesis in Arabidopsis: Mutational spectrum. Plant J. 1991, 1, 71–82. [Google Scholar] [CrossRef]
- Gang, H.; Liu, G.; Zhang, M.; Zhao, Y.; Jiang, J.; Chen, S. Comprehensive characterization of T-DNA integration induced chromosomal rearrangement in a birch T-DNA mutant. BMC Genom. 2019, 20, 311. [Google Scholar] [CrossRef] [Green Version]
- Jupe, F.; Rivkin, A.C.; Michael, T.P.; Zander, M.; Motley, S.T.; Sandoval, J.P.; Keith Slotkin, R.; Chen, H.; Castanon, R.; Nery, J.R.; et al. The complex architecture and epigenomic impact of plant T-DNA insertions. PLoS Genet. 2019, 15, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Pucker, B.; Kleinbölting, N.; Weisshaar, B. Large scale genomic rearrangements in selected Arabidopsis thaliana T-DNA lines are caused by T-DNA insertion mutagenesis. BMC Genom. 2021, 22, 599. [Google Scholar] [CrossRef]
- Lopez, L.S.; Völkner, C.; Day, P.M.; Lewis, C.M.; Lewis, C.L.; Schneider, D.; Correa Galvis, V.; Cruz, J.A.; Armbruster, U.; Kramer, D.M.; et al. The Arabidopsis T-DNA mutant SALK_008491 carries a 14-kb deletion on chromosome 3 that provides rare insights into the plant response to dynamic light stress. Plant Direct 2022, 6, e429. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxe. Biol. Chem. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Shen, J.; Li, Z.; Fu, Y.; Liang, J. Identification and molecular characterization of the alternative spliced variants of beta carbonic anhydrase 1 (βCA1) from Arabidopsis thaliana. PeerJ 2021, 9, e12673. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van Der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Stuttmann, J.; Barthel, K.; Martin, P.; Ordon, J.; Erickson, J.L.; Herr, R.; Ferik, F.; Kretschmer, C.; Berner, T.; Keilwagen, J.; et al. Highly efficient multiplex editing: One-shot generation of 8× Nicotiana benthamiana and 12× Arabidopsis mutants. Plant J. 2021, 106, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Castel, B.; Tomlinson, L.; Locci, F.; Yang, Y.; Jones, J.D.G. Optimization of T-DNA architecture for Cas9-mediated mutagenesis in Arabidopsis. PLoS ONE 2019, 14, e0204778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; McKenna, A.; Schreiber, J.; Haeussler, M.; Yin, Y.; Agarwal, V.; Noble, W.S.; Shendure, J. Massively parallel profiling and predictive modeling of the outcomes of CRISPR/Cas9-mediated double-strand break repair. Nucleic Acids Res. 2019, 47, 7989–8003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soyars, C.L.; Peterson, B.A.; Burr, C.A.; Nimchuk, Z.L. Cutting Edge Genetics: CRISPR/Cas9 Editing of Plant Genomes. Plant Cell Physiol. 2018, 59, 1608–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauser, F.; Schiml, S.; Puchta, H. Both CRISPR/Cas-based nucleases and nickases can be used efficiently for genome engineering in Arabidopsis thaliana. Plant J. 2014, 79, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Mao, Y.; Xu, N.; Zhang, B.; Wei, P.; Yang, D.-L.; Wang, Z.; Zhang, Z.; Zheng, R.; Yang, L.; et al. Multigeneration analysis reveals the inheritance, specificity, and patterns of CRISPR/Cas-induced gene modifications in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 4632–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Bianchi, S.; Betterle, N.; Kouril, R.; Cazzaniga, S.; Boekema, E.; Bassi, R.; Dall’Osto, L. Arabidopsis mutants deleted in the light-harvesting protein Lhcb4 have a disrupted photosystem II macrostructure and are defective in photoprotection. Plant Cell 2011, 23, 2659–2679. [Google Scholar] [CrossRef] [Green Version]
- Teardo, E.; de Laureto, P.P.; Bergantino, E.; Dalla Vecchia, F.; Rigoni, F.; Szabò, I.; Giacometti, G.M. Evidences for interaction of PsbS with photosynthetic complexes in maize thylakoids. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Floris, M.; Bassi, R.; Robaglia, C.; Alboresi, A.; Lanet, E. Post-transcriptional control of light-harvesting genes expression under light stress. Plant Mol. Biol. 2013, 82, 147–154. [Google Scholar] [CrossRef]
- Borisova-Mubarakshina, M.M.; Vetoshkina, D.V.; Rudenko, N.N.; Shirshikova, G.N.; Fedorchuk, T.P.; Naydov, I.A.; Ivanov, B.N. The size of the light-harvesting antenna of higher plant photosystem ii is regulated by illumination intensity through transcription of antenna protein genes. Biochemistry 2014, 79, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Ignatova, L.K.; Rudenko, N.N.; Khristin, M.S.; Ivanov, B.N. Heterogeneous origin of carbonic anhydrase activity of thylakoid membranes. Biochemistry 2006, 71, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Streb, S.; Zeeman, S.C. Starch Metabolism in Arabidopsis. Arab. Book 2012, 10, e0160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolleter, D.; Chochois, V.; Poiré, R.; Dean Price, G.; Badger, M.R. Measuring CO2 and HCO3− permeabilities of isolated chloroplasts using a MIMS-18O approach. J. Exp. Bot. 2017, 68, 3915–3924. [Google Scholar] [CrossRef] [Green Version]
- Perez-Martin, A.; Michelazzo, C.; Torres-Ruiz, J.M.; Flexas, J.; Fernández, J.E.; Sebastiani, L.; Diaz-Espejo, A. Regulation of photosynthesis and stomatal and mesophyll conductance under water stress and recovery in olive trees: Correlation with gene expression of carbonic anhydrase and aquaporins. J. Exp. Bot. 2014, 65, 3143–3156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wildermuth, M.C.; Dewdney, J.; Wu, G.; Ausubel, F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001, 414, 562–565. [Google Scholar] [CrossRef]
- Medina-Puche, L.; Castelló, M.J.; Canet, J.V.; Lamilla, J.; Colombo, M.L.; Tornero, P. β-carbonic anhydrases play a role in salicylic acid perception in Arabidopsis. PLoS ONE 2017, 12, e0181820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borzenkova, R.A.; Borovkova, M.P. Developmental Patterns of Phytohormone Content in the Cortex and Pith of Potato Tubers as Related to Their Growth and Starch Content. Russ. J. Plant Physiol. 2003, 50, 119–124. [Google Scholar] [CrossRef]
- Sunderhaus, S.; Dudkina, N.V.; Jänsch, L.; Klodmann, J.; Heinemeyer, J.; Perales, M.; Zabaleta, E.; Boekema, E.J.; Braun, H.P. Carbonic anhydrase subunits form a matrix-exposed domain attached to the membrane arm of mitochondrial complex I in plants. J. Biol. Chem. 2006, 281, 6482–6488. [Google Scholar] [CrossRef] [Green Version]
- Concordet, J.P.; Haeussler, M. CRISPOR: Intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.; Zhao, W.; Richards, J.T.; Wheeler, R.M.; Guy, C.L.; Levine, L.H. Transcriptional and metabolic insights into the differential physiological responses of arabidopsis to optimal and supraoptimal atmospheric co2. PLoS ONE 2012, 7, e0043583. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Starch Content, % | ||
|---|---|---|---|
| After 16 h of Darkness | After 8 h of Light | ||
| WT | 100 | 232.0 ± 25.6 | |
| αCA4-KO | Crispr_α4 | 63.2 ± 5.9 | 295.7 ± 38.5 |
| 8-8 | 80.1 ± 11.4 | 390.1 ± 11.4 | |
| 9-12 | 85.6 ± 14.9 | 392.0 ± 15.4 | |
| Name of the Oligonucleotide | Nucleotide Sequence 5′-3′ |
|---|---|
| Ath guide218 F | ATTGAAAACGGAAAAGGGACCAGA |
| Ath guide218 R | AAACTCTGGTCCCTTTTCCGTTTT |
| pDGE forward | CGAATCAAAAGTTGAGCTCC |
| Ath αCA4 Test1 F | ATTTCACACGCTCATTCTGAAGTCG |
| Ath αCA4 Test1 R | AAGGAGGAACAGTGAGAGAGCCAA |
| Ath αCA4 Test2 R | GGTCGTACCTGTAATAATGCCAAAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudenko, N.N.; Permyakova, N.V.; Ignatova, L.K.; Nadeeva, E.M.; Zagorskaya, A.A.; Deineko, E.V.; Ivanov, B.N. The Role of Carbonic Anhydrase αCA4 in Photosynthetic Reactions in Arabidopsis thaliana Studied, Using the Cas9 and T-DNA Induced Mutations in Its Gene. Plants 2022, 11, 3303. https://doi.org/10.3390/plants11233303
Rudenko NN, Permyakova NV, Ignatova LK, Nadeeva EM, Zagorskaya AA, Deineko EV, Ivanov BN. The Role of Carbonic Anhydrase αCA4 in Photosynthetic Reactions in Arabidopsis thaliana Studied, Using the Cas9 and T-DNA Induced Mutations in Its Gene. Plants. 2022; 11(23):3303. https://doi.org/10.3390/plants11233303
Chicago/Turabian StyleRudenko, Natalia N., Natalya V. Permyakova, Lyudmila K. Ignatova, Elena M. Nadeeva, Alla A. Zagorskaya, Elena V. Deineko, and Boris N. Ivanov. 2022. "The Role of Carbonic Anhydrase αCA4 in Photosynthetic Reactions in Arabidopsis thaliana Studied, Using the Cas9 and T-DNA Induced Mutations in Its Gene" Plants 11, no. 23: 3303. https://doi.org/10.3390/plants11233303