
Chemical Composition of Various Nepeta cataria Plant Organs’ Methanol Extracts Associated with In Vivo Hepatoprotective and Antigenotoxic Features as well as Molecular Modeling Investigations

 , , , ,
, , , ,  and
and 
Abstract
:
1. Introduction
2. Results and Discussion
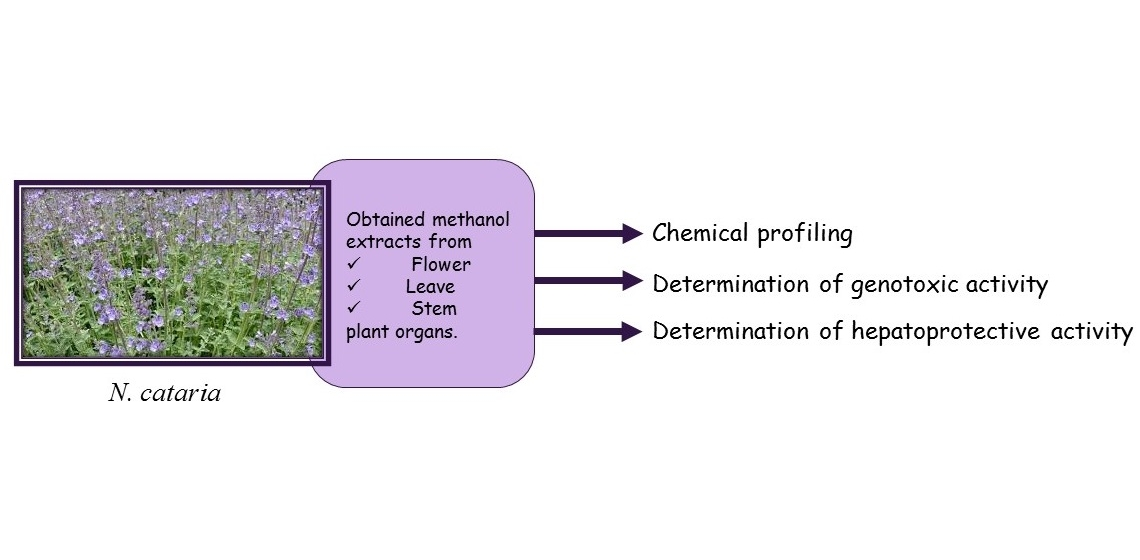
2.1. Chemical Composition of Methanol Extracts of N. cataria
2.1.1. Flowers Methanol Extract (FME) of N. cataria
2.1.2. Leaves Methanol Extract (LME) of N. cataria
2.1.3. Stems Methanol Extract (SME) of N. cataria
2.1.4. Implications of N. cataria Extracts Chemical Composition on the Plant’s Activity
2.2. Hepatotoxic and Hepatoprotective Features of N. cataria Flower (FME), Leaf (LME), and Stem (SME) Methanol Extracts
2.2.1. Hepatotoxic and Hepatoprotective Features of N. cataria Flower Methanol Extract (FME)
The Hepatocytes Redox Status

{kind=link}
{kind=link}
{kind=link}
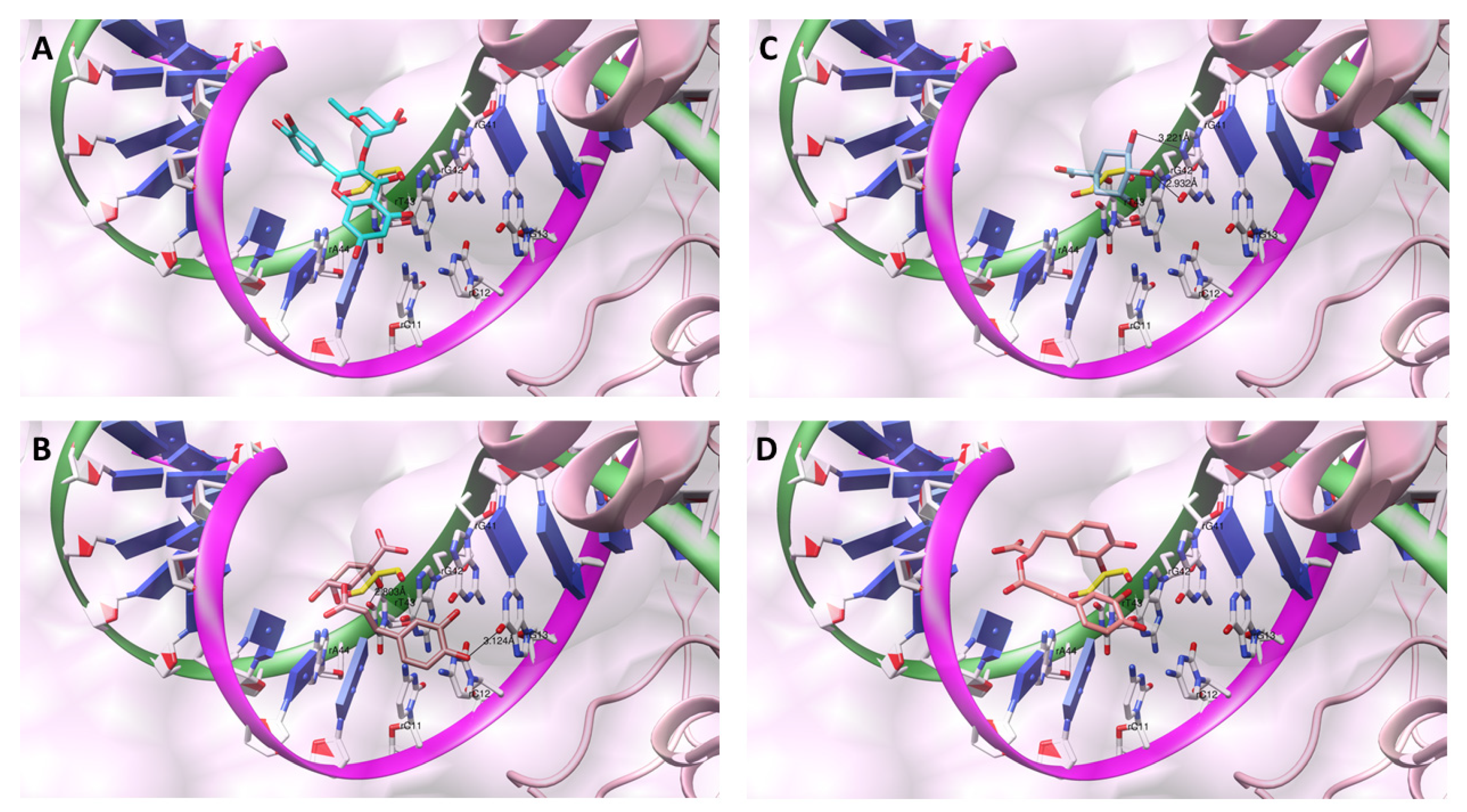{kind=link}
{kind=link}
{kind=link}
| Groups | rTP (g/L) | rSOD (U/mg) | rTBARS (nmol/mg) | rCAT (U/mg) | rGSH (mg/g) |
|---|---|---|---|---|---|
| I | 1 28.39 ± 0.14 | 5.38 ± 0.03 | 2.19 ± 0.02 | 131.33 ± 0.15 | 16.04 ± 0.12 |
| II | 27.07 ± 0.12 | 2.45 ± 0.05 * | 10.51 ± 0.14 * | 106.43 ± 0.3 * | 6.39 ± 0.16 * |
| III | 30.01 ± 0.17 | 4.05 ± 0.11 *† | 1.26 ± 0.08 † | 128.97 ± 0.08 † | 13.22 ± 0.20 *† |
| IV | 26.13 ± 0.23 | 4.29 ± 0.02 *† | 2.39 ± 0.09 † | 127.57 ± 0.10 † | 13.52 ± 0.12 *† |
| V | 25.90 ± 0.34 | 4.31 ± 0.06 *† | 1.33 ± 0.08 *† | 124.31 ± 0.15 *† | 14.79 ± 0.13 *† |
| VI | 27.61 ± 0.24 | 3.26 ± 0.06 *† | 2.24 ± 0.02 † | 113.07 ± 0.23 *† | 10.63 ± 0.14 *† |
| VII | 34.19 ± 0.17 | 3.26 ± 0.08 *† | 1.88 ± 0.03 † | 114.34 ± 0.22 *† | 11.44 ± 0.02 *† |
| VIII | 27.48 ± 0.41 | 3.28 ± 0.09 *† | 1.52 ± 0.06 *† | 120.93 ± 0.11 *† | 12.01 ± 0.18 *† |
| IX | 28.17 ± 0.28 | 3.41 ± 0.04 *† | 2.81 ± 0.09 *† | 113.65 ± 0.28 *† | 12.18 ± 0.08 *† |
| X | 25.23 ± 0.16 | 3.65 ± 0.02 *† | 1.82 ± 0.07 † | 126.40 ± 0.13 † | 13.26 ± 0.06 *† |
| XI | 28.88 ± 0.13 | 3.09 ± 0.07 *† | 2.93 ± 0.05 *† | 119.32 ± 0.19 *† | 12.18 ± 0.03 *† |
| XII | 27.78 ± 0.23 | 3.31 ± 0.08 *† | 4.31 ± 0.07 *† | 123.80 ± 0.16 *† | 12.79 ± 0.21 *† |
| XIII | 29.56 ± 0.16 | 3.11 ± 0.09 *† | 5.08 ± 0.10 *† | 115.25 ± 0.20 *† | 11.61 ± 0.13 *† |
| XIV | 27.70 ± 0.32 | 3.25 ± 0.01 *† | 4.99 ± 0.05 *† | 124.34 ± 0.14 *† | 12.18 ± 0.09 *† |
| Group/Compounds | Conc. (mg/kg bwt) | rTP (g/L) | rSOD (U/mg) | rTBARS (nmol/mg) | rCAT (U/mg) | rGSH (mg/g) |
|---|---|---|---|---|---|---|
| Control group | 1 27.43 ± 0.21 † | 5.26 ± 0.14 † | 2.15 ± 0.32 † | 134.17 ± 0.15 † | 17.32 ± 0.20 † | |
| CCl4 | 1 mL/kg | 25.43 ± 0.53 * | 2.53 ± 0.21 * | 10.78 ± 0.32 * | 107.56 ± 0.30 * | 6.42 ± 0.41 * |
| quinic acid | 50 | 22.43 ± 0.06 *† | 2.74 ± 0.12 * | 2.54 ± 0.21 † | 108.34 ± 0.32 *† | 8.04 ± 0.32 *† |
| 100 | 31.45 ± 0.21 *† | 2.81 ± 0.32 * | 2.10 ± 0.43 † | 109.17 ± 0.51 *† | 8.43 ± 0.27 *† | |
| 200 | 21.32 ± 0.45 *† | 2.91 ± 0.26 * | 1.98 ± 0.32 *† | 110.56 ± 0.32 *† | 9.21 ± 0.32 *† | |
| protocatechuic acid | 50 | 32.79 ± 0.46 *† | 2.66 ± 0.19 * | 2.95 ± 0.11 † | 110.25 ± 0.36 *† | 7.17 ± 0.63 *† |
| 100 | 29.18 ± 0.95 *† | 2.69 ± 0.48 * | 3.18 ± 0.17 *† | 110.65 ± 0.91 *† | 7.34 ± 0.64 *† | |
| 200 | 30.05 ± 0.09 *† | 2.75 ± 0.14 * | 3.20 ± 0.47 *† | 113.89 ± 0.56 *† | 7.95 ± 0.46 *† | |
| chlorogenic acid | 50 | 28.33 ± 0.47 *† | 2.88 ± 0.05 * | 2.43 ± 0.13 † | 110.11 ± 0.55 *† | 10.15 ± 0.28 *† |
| 100 | 26.18 ± 0.19 *† | 2.95 ± 0.18 * | 2.37 ± 0.53 † | 112.28 ± 0.32 *† | 11.92 ± 0.31 *† | |
| 200 | 25.48 ± 0.11 * | 2.98 ± 0.33 * | 2.25 ± 0.46 † | 112.78 ± 0.28 *† | 12.28 ± 0.25 *† | |
| p-hydroxybenzoic acid | 50 | 20.66 ± 0.16 *† | 2.59 ± 0.22 * | 3.99 ± 0.47 *† | 107.98 ± 0.79 * | 7.02 ± 0.17 *† |
| 100 | 23.17 ± 0.96 *† | 2.63 ± 0.24 * | 3.90 ± 0.79 *† | 108.11 ± 0.36 *† | 7.49 ± 0.87 *† | |
| 200 | 23.11 ± 0.36 *† | 2.90 ± 0.33 * | 3.69 ± 0.09 *† | 108.92 ± 0.46 *† | 7.51 ± 0.42 *† | |
| caffeic acid | 50 | 29.83 ± 0.45 *† | 2.64 ± 0.53 * | 4.18 ± 0.31 *† | 108.49 ± 0.25 *† | 6.81 ± 0.22 * |
| 100 | 27.55 ± 0.36† | 2.66 ± 0.34 * | 4.02 ± 0.07 *† | 110.71 ± 0.69 *† | 6.92 ± 0.13 * | |
| 200 | 27.19 ± 0.16† | 2.72 ± 0.47 * | 3.97 ± 0.71 *† | 111.31 ± 0.33 *† | 7.14 ± 0.74 *† | |
| syringic acid | 50 | 22.14 ± 0.32 *† | 2.78 ± 0.77 * | 2.23 ± 0.14 † | 115.28 ± 0.71 *† | 11.98 ± 0.49 *† |
| 100 | 28.18 ± 0.23 *† | 3.18 ± 0.65 *† | 2.20 ± 0.38 † | 116.22 ± 0.24 *† | 12.92 ± 0.28 *† | |
| 200 | 24.65 ± 0.45 *† | 3.55 ± 0.39 *† | 2.19 ± 0.24 † | 118.39 ± 0.44 *† | 13.45 ± 0.36 *† | |
| p-coumaric acid | 50 | 24.78 ± 0.15 *† | 2.74 ± 0.12 * | 2.72 ± 0.28 † | 112.28 ± 0.35 *† | 9.58 ± 0.27 *† |
| 100 | 23.38 ± 0.30 *† | 2.79 ± 0.18 * | 2.58 ± 0.16 † | 112.99 ± 0.76 *† | 10.81 ± 0.14 *† | |
| 200 | 25.47 ± 0.12 * | 2.92 ± 0.74 * | 2.49 ± 0.71 † | 113.85 ± 0.29 *† | 11.17 ± 0.47 *† | |
| ferulic acid | 50 | 28.91 ± 0.16 *† | 2.79 ± 0.61 * | 3.01 ± 0.19 *† | 113.11 ± 0.13 *† | 8.67 ± 0.37 *† |
| 100 | 25.71 ± 0.63 * | 2.85 ± 0.37 * | 2.87 ± 0.22 † | 114.26 ± 0.34 *† | 8.99 ± 0.47 *† | |
| 200 | 23.33 ± 0.24 *† | 2.91 ± 0.11 * | 2.64 ± 0.17 † | 116.96 ± 0.26 *† | 9.25 ± 0.66 *† | |
| luteolin-7-O-glucoside | 50 | 21.74 ± 0.29 *† | 2.85 ± 0.27 * | 2.98 ± 0.03 † | 109.45 ± 0.62 *† | 8.04 ± 0.72 *† |
| 100 | 26.41 ± 0.39 *† | 2.99 ± 0.39 * | 2.90 ± 0.15 † | 110.21 ± 0.32 *† | 8.19 ± 0.37 *† | |
| 200 | 25.77 ± 0.07 * | 3.10 ± 0.09 *† | 2.61 ± 0.13 † | 114.65 ± 0.16 *† | 8.41 ± 0.11 *† | |
| quercetin-3-O-glucoside | 50 | 1 27.43 ± 0.21† | 5.26 ± 0.14 † | 2.15 ± 0.32 † | 134.17 ± 0.15 † | 17.32 ± 0.20 † |
| 100 | 25.43 ± 0.53 * | 2.53 ± 0.21 * | 10.78 ± 0.32 * | 107.56 ± 0.30 * | 6.42 ± 0.41 * | |
| 200 | 24.57 ± 0.34 *† | 2.59 ± 0.13 * | 3.11 ± 0.16 *† | 110.22 ± 0.28 *† | 7.99 ± 0.24 *† | |
| quercetin-3-O-rutinoside | 50 | 22.18 ± 0.14 *† | 2.69 ± 0.08 * | 2.95 ± 0.39 † | 112.74 ± 0.36 *† | 8.10 ± 0.31 *† |
| 100 | 25.96 ± 0.19 * | 2.74 ± 0.16 * | 2.78 ± 0.44 † | 115.18 ± 0.58 *† | 8.24 ± 0.77 *† | |
| 200 | 27.72 ± 0.11 † | 2.71 ± 0.19 * | 3.14 ± 0.26 *† | 116.78 ± 0.07 *† | 8.01 ± 0.64 *† | |
| rosmarinic acid | 50 | 24.79 ± 0.46 *† | 2.75 ± 0.13 * | 3.08 ± 0.47 *† | 117.96 ± 0.45 *† | 8.09 ± 0.43 *† |
| 100 | 25.11 ± 0.35 * | 2.98 ± 0.47 * | 2.97 ± 0.58 † | 120.47 ± 0.67 *† | 8.20 ± 0.27 *† | |
| 200 | 22.13 ± 0.45 *† | 2.57 ± 0.03 * | 2.65 ± 0.18 † | 109.52 ± 0.14 *† | 10.25 ± 0.07 *† | |
| quercitrin | 50 | 21.18 ± 0.33 *† | 2.68 ± 0.09 * | 2.24 ± 0.08 † | 110.33 ± 0.23 *† | 11.78 ± 0.09 *† |
| 100 | 24.79 ± 0.26 *† | 2.99 ± 0.12 * | 2.22 ± 0.09 † | 110.98 ± 0.32 *† | 11.26 ± 0.36 *† | |
| 200 | 21.53 ± 0.53 *† | 2.86 ± 0.54 * | 2.05 ± 0.74 † | 112.56 ± 0.31 *† | 12.43 ± 0.84 *† | |
| kaempferol-3-O-glucoside | 50 | 28.43 ± 0.54 *† | 2.94 ± 0.23 * | 1.95 ± 0.51 *† | 114.52 ± 0.53 *† | 13.11 ± 0.31 *† |
| 100 | 25.54 ± 0.11 * | 3.72 ± 0.77 *† | 1.64 ± 0.32 *† | 117.32 ± 0.21 *† | 13.32 ± 0.43 *† | |
| 200 | 21.54 ± 0.18 *† | 2.96 ± 0.16 * | 3.01 ± 0.22 *† | 111.27 ± 0.21 *† | 11.98 ± 0.66 *† | |
| quercetin | 50 | 21.98 ± 0.47 *† | 3.14 ± 0.13 *† | 2.78 ± 0.61 † | 113.78 ± 0.36 *† | 13.56 ± 0.19 *† |
| 100 | 26.74 ± 0.07 *† | 3.25 ± 0.34 *† | 2.47 ± 0.07 † | 119.14 ± 0.25 *† | 14.76 ± 0.11 *† | |
| 200 | 22.74 ± 0.11 *† | 2.97 ± 0.17 * | 2.01 ± 0.26 † | 113.78 ± 0.51 *† | 14.27 ± 0.09 *† | |
| luteolin | 50 | 24.71 ± 0.24 *† | 3.15 ± 0.66 *† | 2.48 ± 0.41 † | 115.63 ± 0.14 *† | 14.98 ± 0.17 *† |
| 100 | 25.98 ± 0.64 * | 3.67 ± 0.47 *† | 2.22 ± 0.45 † | 118.39 ± 0.33 *† | 15.36 ± 0.33 *† | |
| 200 | 24.69 ± 0.37 *† | 2.57 ± 0.57 * | 3.28 ± 0.49 *† | 108.49 ± 0.34 *† | 7.58 ± 0.31 *† | |
| kaempferol | 50 | 23.15 ± 0.41 *† | 2.59 ± 0.64 * | 2.97 ± 0.34 † | 110.64 ± 0.97 *† | 8.25 ± 0.16 *† |
| 100 | 22.14 ± 0.12 *† | 2.74 ± 0.08 * | 2.68 ± 0.09 † | 111.93 ± 0.13 *† | 10.92 ± 0.06 *† | |
| 200 | 28.14 ± 0.05 *† | 2.78 ± 0.16 * | 3.15 ± 0.19 *† | 115.97 ± 0.14 *† | 10.44 ± 0.22 *† | |
| isorhamnetin | 50 | 26.81 ± 0.31 *† | 2.82 ± 0.64 * | 3.10 ± 0.24 *† | 119.36 ± 0.37 *† | 10.18 ± 0.20 *† |
| 100 | 26.93 ± 0.12 *† | 2.95 ± 0.47 * | 2.99 ± 0.11 † | 120.55 ± 0.96 *† | 12.93 ± 0.13 *† | |
| 200 | 25.49 ± 0.17 * | 2.99 ± 0.13 * | 3.01 ± 0.17 *† | 122.68 ± 0.11 *† | 9.16 ± 0.41 *† |
The Hepatocytes Cell Membrane Status
| Group | rAST (U/L) | rALT (U/L) | rALP (U/L) | rγ-GT (U/L) |
|---|---|---|---|---|
| I | 1 3.16 ± 0.02 | 20.50 ± 0.23 | 266.27 ± 0.21 | 4.10 ± 0.03 |
| II | 71.12 ± 0.08 * | 62.69 ± 0.30 * | 359.18 ± 0.18 * | 16.88 ± 0.15 * |
| III | 3.21 ± 0.02 † | 37.75 ± 0.09 *† | 269.44 ± 0.13 † | 4.18 ± 0.14 † |
| IV | 10.64 ± 0.25 *† | 36.49 ± 0.12 *† | 221.26 ± 0.19 *† | 7.37 ± 0.06 *† |
| V | 9.81 ± 0.09 *† | 30.71 ± 0.02 *† | 356.29 ± 0.25 * | 8.44 ± 0.08 *† |
| VI | 5.42 ± 0.01 *† | 61.59 ± 0.23 * | 297.82 ± 0.27 *† | 10.81 ± 0.08 *† |
| VII | 4.83 ± 0.04 † | 58.23 ± 0.18 *† | 284.34 ± 0.12 *† | 7.37 ± 0.31 *† |
| VIII | 3.78 ± 0.09 † | 39.04 ± 0.13 *† | 271.89 ± 0.14 *† | 5.02 ± 0.24 † |
| IX | 3.60 ± 0.04 † | 69.52 ± 0.14 *† | 349.44 ± 0.17 * | 11.55 ± 0.11 *† |
| X | 3.92 ± 0.03 † | 78.56 ± 0.28 *† | 324.30 ± 0.13 *† | 14.25 ± 0.05 *† |
| XI | 11.13 ± 0.12 *† | 39.98 ± 0.06 *† | 246.85 ± 0.09 *† | 28.00 ± 0.21 *† |
| XII | 2.29 ± 0.01 † | 22.86 ± 0.11 † | 329.11 ± 0.31 *† | 9.34 ± 0.09 *† |
| XIII | 3.46 ± 0.03 † | 28.13 ± 0.09 *† | 351.91 ± 0.36 * | 29.48 ± 0.01 *† |
| XIV | 11.87 ± 0.11 *† | 46.95 ± 0.14 *† | 456.12 ± 0.14 *† | 17.20 ± 0.04 * |
2.2.2. Hepatotoxic and Hepatoprotective Features of N. cataria Leaf Methanol Extract (LME)
The Hepatocytes Redox Status
The Hepatocytes Cell Membrane Status
2.2.3. Hepatotoxic and Hepatoprotective Features of N. cataria Stems Methanol Extract (SME)
The Hepatocytes’ Redox Status
The Hepatocytes’ Cell Membrane Status
| Group/Compounds | Conc. (mg/kg bwt) | rAST (U/L) | rALT (U/L) | rALP (U/L) | rγ-GT (U/L) |
|---|---|---|---|---|---|
| Control group | 1 4.08 ± 0.12 † | 21.37 ± 0.26 † | 272.94 ± 0.83 † | 4.01 ± 0.03 † | |
| CCl4 | 1 mL/kg | 69.64 ± 0.41 * | 64.72 ± 0.38 * | 371.14 ± 0.46 * | 15.78 ± 0.08 * |
| quinic acid | 50 | 5.58 ± 0.08 *† | 51.04 ± 0.62 *† | 289.59 ± 0.33 *† | 11.41 ± 0.03 *† |
| 100 | 5.12 ± 0.14 *† | 45.94 ± 0.13 *† | 285.17 ± 0.28 *† | 9.23 ± 0.11 *† | |
| 200 | 4.96 ± 0.07 † | 44.19 ± 0.27 *† | 279.86 ± 0.19 † | 8.40 ± 0.26 *† | |
| protocatechuic acid | 50 | 9.11 ± 0.18 *† | 59.67 ± 0.10 *† | 323.16 ± 0.85 *† | 12.53 ± 0.32 *† |
| 100 | 7.56 ± 0.06 *† | 54.46 ± 0.34 *† | 300.54 ± 0.62 *† | 11.70 ± 0.10 *† | |
| 200 | 7.01 ± 0.20 *† | 51.74 ± 0.22 *† | 298.44 ± 0.30 *† | 9.29 ± 0.13 *† | |
| chlorogenic acid | 50 | 7.34 ± 0.05 *† | 50.11 ± 0.41 *† | 302.51 ± 0.49 *† | 10.21 ± 0.15 *† |
| 100 | 6.03 ± 0.10 *† | 42.67 ± 0.30 *† | 284.53 ± 0.32 *† | 9.28 ± 0.50 *† | |
| 200 | 4.28 ± 0.07 † | 37.12 ± 0.28 *† | 280.22 ± 0.41 *† | 6.80 ± 0.77 *† | |
| p-hydroxybenzoic acid | 50 | 19.71 ± 0.08 *† | 55.16 ± 0.52 *† | 354.79 ± 0.97 *† | 11.83 ± 0.65 *† |
| 100 | 14.28 ± 0.26 *† | 52.11 ± 0.40 *† | 331.60 ± 0.54 *† | 10.91 ± 0.38 *† | |
| 200 | 10.46 ± 0.37 *† | 48.73 ± 0.13 *† | 310.93 ± 0.19 *† | 9.04 ± 0.50 *† | |
| caffeic acid | 50 | 27.53 ± 0.41 *† | 63.61 ± 0.55 *† | 360.38 ± 0.59 *† | 13.91 ± 0.05 *† |
| 100 | 20.14 ± 0.52 *† | 62.11 ± 0.43 *† | 322.62 ± 0.74 *† | 10.16 ± 0.37 *† | |
| 200 | 19.97 ± 0.77 *† | 48.23 ± 0.71 *† | 307.08 ± 0.62 *† | 9.10 ± 0.52 *† | |
| syringic acid | 50 | 16.46 ± 0.06 *† | 62.39 ± 0.30 *† | 382.63 ± 0.95 *† | 13.67 ± 0.08 *† |
| 100 | 15.17 ± 0.17 *† | 53.65 ± 0.44 *† | 366.19 ± 0.48 *† | 11.49 ± 0.23 *† | |
| 200 | 12.39 ± 0.53 *† | 41.25 ± 0.31 *† | 350.02 ± 0.36 *† | 10.42 ± 0.64 *† | |
| p-coumaric acid | 50 | 24.73 ± 0.46 *† | 60.18 ± 0.68 *† | 314.09 ± 0.43 *† | 11.44 ± 0.13 *† |
| 100 | 20.16 ± 0.38 *† | 54.03 ± 0.34 *† | 296.91 ± 0.84 *† | 9.97 ± 0.08 *† | |
| 200 | 15.69 ± 0.18 *† | 40.82 ± 0.08 *† | 289.92 ± 0.38 *† | 8.05 ± 0.61 *† | |
| ferulic acid | 50 | 8.28 ± 0.61 *† | 52.10 ± 0.24 *† | 312.14 ± 0.92 *† | 11.77 ± 0.19 *† |
| 100 | 7.39 ± 0.05 *† | 47.22 ± 0.56 *† | 297.96 ± 0.43 *† | 9.26 ± 0.46 *† | |
| 200 | 5.94 ± 0.23 *† | 41.69 ± 0.10 *† | 288.71 ± 0.55 *† | 8.10 ± 0.49 *† | |
| luteolin-7-O-glucoside | 50 | 23.17 ± 0.46 *† | 59.71 ± 0.26 *† | 319.37 ± 0.53 *† | 13.78 ± 0.10 *† |
| 100 | 21.55 ± 0.83 *† | 55.96 ± 0.09 *† | 302.74 ± 0.33 *† | 10.62 ± 0.41 *† | |
| 200 | 17.49 ± 0.09 *† | 39.28 ± 0.17 *† | 281.93 ± 0.42 *† | 9.11 ± 0.72 *† | |
| quercetin-3-O-glucoside | 50 | 7.82 ± 0.36 *† | 51.69 ± 0.44 *† | 304.03 ± 0.20 *† | 10.58 ± 0.04 *† |
| 100 | 6.99 ± 0.42 *† | 46.02 ± 0.13 *† | 296.37 ± 0.17 *† | 9.35 ± 0.50 *† | |
| 200 | 5.17 ± 0.07 *† | 40.33 ± 0.76 *† | 284.46 ± 0.66 *† | 6.43 ± 0.21 *† | |
| quercetin-3-O-rutinoside | 50 | 9.26 ± 0.18 *† | 53.10 ± 0.22 *† | 300.23 ± 0.44 *† | 12.77 ± 0.16 *† |
| 100 | 7.33 ± 0.17 *† | 48.47 ± 0.07 *† | 294.16 ± 0.62 *† | 9.16 ± 0.41 *† | |
| 200 | 5.70 ± 0.30 *† | 43.64 ± 0.39 *† | 280.15 ± 0.38 *† | 7.73 ± 0.06 *† | |
| rosmarinic acid | 50 | 5.71 ± 0.18 *† | 43.01 ± 0.30 *† | 297.55 ± 0.14 *† | 10.40 ± 0.31 *† |
| 100 | 4.42 ± 0.20 † | 35.12 ± 0.22 *† | 281.96 ± 0.35 *† | 8.04 ± 0.13 *† | |
| 200 | 4.13 ± 0.02 † | 32.64 ± 0.40 *† | 273.58 ± 0.31 † | 5.61 ± 0.23 *† | |
| quercitrin | 50 | 4.72 ± 0.31 † | 47.66 ± 0.38 *† | 299.48 ± 0.21 *† | 9.56 ± 0.29 *† |
| 100 | 4.25 ± 0.10 † | 40.19 ± 0.09 *† | 293.13 ± 0.84 *† | 6.28 ± 0.08 *† | |
| 200 | 4.10 ± 0.27 † | 38.62 ± 0.67 *† | 274.22 ± 0.34 † | 4.40 ± 0.40 † | |
| quercetin-3-O-glucoside | 50 | 7.82 ± 0.36 *† | 51.69 ± 0.44 *† | 304.03 ± 0.20 *† | 10.58 ± 0.04 *† |
| 100 | 6.99 ± 0.42 *† | 46.02 ± 0.13 *† | 296.37 ± 0.17 *† | 9.35 ± 0.50 *† | |
| 200 | 5.17 ± 0.07 *† | 40.33 ± 0.76 *† | 284.46 ± 0.66 *† | 6.43 ± 0.21 *† | |
| kaempferol-3-O-glucoside | 50 | 15.82 ± 0.09 *† | 61.22 ± 0.08 *† | 313.08 ± 0.54 *† | 11.60 ± 0.08 *† |
| 100 | 13.95 ± 0.20 *† | 53.18 ± 0.16 *† | 302.60 ± 0.97 *† | 10.35 ± 0.40 *† | |
| 200 | 10.61 ± 0.15 *† | 50.29 ± 0.23 *† | 290.53 ± 0.72 *† | 9.94 ± 0.31 *† | |
| quercetin | 50 | 6.28 ± 0.17 *† | 50.52 ± 0.24 *† | 317.37 ± 0.55 *† | 11.64 ± 0.30 *† |
| 100 | 5.14 ± 0.62 *† | 45.26 ± 0.61 *† | 300.56 ± 0.31 *† | 9.92 ± 0.07 *† | |
| 200 | 4.49 ± 0.43 † | 41.40 ± 0.38 *† | 291.74 ± 0.97 *† | 7.50 ± 0.46 *† | |
| luteolin | 50 | 25.18 ± 0.63 *† | 62.83 ± 0.49 *† | 325.74 ± 0.30 *† | 12.75 ± 0.32 *† |
| 100 | 22.79 ± 0.08 *† | 57.01 ± 0.37 *† | 299.41 ± 0.67 *† | 10.06 ± 0.42 *† | |
| 200 | 18.95 ± 0.84 *† | 49.46 ± 0.26 *† | 292.16 ± 0.11 *† | 9.14 ± 0.58 *† | |
| kaempferol | 50 | 15.82 ± 0.09 *† | 61.22 ± 0.08 *† | 313.08 ± 0.54 *† | 11.60 ± 0.08 *† |
| 100 | 13.95 ± 0.20 *† | 53.18 ± 0.16 *† | 302.60 ± 0.97 *† | 10.35 ± 0.40 *† | |
| 200 | 10.61 ± 0.15 *† | 50.29 ± 0.23 *† | 290.53 ± 0.72 *† | 9.94 ± 0.31 *† | |
| isorhamnetin | 50 | 14.74 ± 0.44 *† | 59.25 ± 0.67 *† | 294.30 ± 0.26 *† | 12.01 ± 0.18 *† |
| 100 | 11.62 ± 0.19 *† | 52.11 ± 0.91 *† | 291.62 ± 0.45 *† | 9.93 ± 0.06 *† | |
| 200 | 7.30 ± 0.07 *† | 44.08 ± 0.33 *† | 282.06 ± 0.36 *† | 8.01 ± 0.32 *† |
2.3. N. cataria Extracts Impact the Genome In Vivo
2.3.1. Genotoxic and Antigenotoxic Activities of N. cataria Flower Methanol Extract (FME)
| Groups | rtail Moment | % rDNA in Tail | rtail Length |
|---|---|---|---|
| I | 1.44 ± 0.32 a† | 3.08 ± 0.40 † | 2.60 ± 0.73 † |
| II | 16.51 ± 1.23 * | 10.80 ± 1.50 * | 24.65 ± 1.03 * |
| III | 1.64 ± 0.63 † | 3.72 ± 0.94 † | 2.90 ± 0.86 † |
| IV | 2.99 ± 0.6 *† | 4.22 ± 0.54 *† | 3.17 ± 0.63 † |
| V | 4.72 ± 0.3 *† | 7.62 ± 0.34 *† | 5.6 ± 0.82 *† |
| Groups | Comet class | Total Score 1 | % R | ||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |||
| I | 73.90 ± 1.02 | 26.10 ± 0.82 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 26.10 ± 0.82 † | / |
| II | 19.33 ± 0.34 | 59.70 ± 1.02 | 16.13 ± 0.30 | 4.84 ± 1.70 | 0.00 ± 0.00 | 106.50 ± 1.04 * | / |
| VI | 38.60 ± 0.33 | 59.10 ± 1.40 | 2.30 ± 0.73 | 0.00 ± 0.00 | 0.00 ± 0.00 | 63.70 ± 0.34 *† | 53.23 |
| VII | 44.86 ± 0.23 | 53.10 ± 0.20 | 2.04 ± 0.03 | 0.00 ± 0.00 | 0.00 ± 0.00 | 57.20 ± 0.30 *† | 61.32 |
| VIII | 53.85 ± 0.40 | 44.61 ± 0.32 | 1.54 ± 0.80 | 0.00 ± 0.00 | 0.00 ± 0.00 | 47.70 ± 1.54 *† | 73.13 |
| IX | 48.00 ± 0.23 | 46.00 ± 0.64 | 6.00 ± 0.50 | 0.00 ± 0.00 | 0.00 ± 0.00 | 58.00 ± 1.70 *† | 60.32 |
| X | 49.95 ± 1.02 | 45.70 ± 0.54 | 4.35 ± 1.60 | 0.00 ± 0.00 | 0.00 ± 0.00 | 54.40 ± 1.02 *† | 64.80 |
| XI | 40.00 ± 0.80 | 60.00 ± 0.30 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 60.00 ± 0.12 *† | 57.80 |
| XII | 62.20 ± 0.20 | 32.40 ± 0.92 | 4.05 ± 0.90 | 1.35 ± 0.52 | 0.00 ± 0.00 | 44.55 ± 0.54 *† | 77.05 |
| XIII | 43.80 ± 0.82 | 47.90 ± 0.32 | 8.30 ± 0.54 | 0.00 ± 0.00 | 0.00 ± 0.00 | 64.50 ± 0.81 *† | 52.24 |
| XIV | 47.90 ± 0.41 | 37.50 ± 0.90 | 12.50 ± 0.32 | 2.10 ± 0.30 | 0.00 ± 0.00 | 68.80 ± 0.46 *† | 46.90 |
| Group/Compounds | Conc. (mg/kg bwt) | Comet Class | Total Score 1 | % R | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | ||||
| Control group | 75.59 ± 1.30 | 24.41 ± 0.52 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 24.41 ± 0.52 † | / | |
| CCl4 | 1 mL/kg | 19.09 ± 0.57 | 58.57 ± 0.44 | 17.42 ± 0.29 | 4.92 ± 0.69 | 0.00 ± 0.00 | 108.17 ± 0.31 * | / |
| quinic acid | 50 | 35.90 ± 0.67 | 60.08 ± 1.03 | 4.02 ± 0.50 | 0.00 ± 0.00 | 0.00 ± 0.00 | 68.12 ± 0.81 *† | 47.82 |
| 100 | 46.48 ± 0.93 | 50.67 ± 0.82 | 2.85 ± 0.43 | 0.00 ± 0.00 | 0.00 ± 0.00 | 56.37 ± 0.94 *† | 61.84 | |
| 200 | 52.70 ± 0.03 | 45.05 ± 0.14 | 2.25 ± 0.52 | 0.00 ± 0.00 | 0.00 ± 0.00 | 49.55 ± 0.61 *† | 69.99 | |
| protocatechuic acid | 50 | 26.95 ± 1.01 | 58.36 ± 0.29 | 11.14 ± 0.37 | 3.55 ± 0.18 | 0.00 ± 0.00 | 91.29 ± 0.28 *† | 20.15 |
| 100 | 36.10 ± 0.57 | 47.56 ± 0.68 | 12.20 ± 0.66 | 4.14 ± 1.06 | 0.00 ± 0.00 | 84.38 ± 1.09 *† | 28.40 | |
| 200 | 42.30 ± 0.22 | 38.70 ± 0.54 | 15.27 ± 0.58 | 3.73 ± 0.27 | 0.00 ± 0.00 | 80.43 ± 1.14 *† | 33.12 | |
| chlorogenic acid | 50 | 46.10 ± 0.91 | 48.65 ± 0.30 | 5.25 ± 0.37 | 0.00 ± 0.00 | 0.00 ± 0.00 | 59.15 ± 0.84 *† | 58.52 |
| 100 | 45.67 ± 0.69 | 48.20 ± 0.48 | 6.13 ± 0.79 | 0.00 ± 0.00 | 0.00 ± 0.00 | 60.46 ± 0.37 *† | 56.96 | |
| 200 | 48.25 ± 0.63 | 41.13 ± 0.54 | 10.62 ± 0.40 | 0.00 ± 0.00 | 0.00 ± 0.00 | 62.37 ± 1.15 *† | 54.68 | |
| p-hydroxybenzoic acid | 50 | 31.82 ± 0.59 | 51.81 ± 0.33 | 13.98 ± 0.52 | 2.39 ± 0.38 | 0.00 ± 0.00 | 86.94 ± 0.33 *† | 25.35 |
| 100 | 33.48 ± 0.50 | 55.00 ± 1.42 | 6.85 ± 0.02 | 4.67 ± 0.50 | 0.00 ± 0.00 | 82.71 ± 0.84 *† | 30.40 | |
| 200 | 29.09 ± 1.06 | 64.23 ± 1.11 | 4.63 ± 0.83 | 2.05 ± 0.44 | 0.00 ± 0.00 | 79.64 ± 1.25 *† | 34.06 | |
| caffeic acid | 50 | 44.42 ± 0.97 | 46.29 ± 0.24 | 9.29 ± 0.62 | 0.00 ± 0.00 | 0.00 ± 0.00 | 64.87 ± 0.34 *† | 51.70 |
| 100 | 46.80 ± 0.39 | 46.69 ± 0.52 | 6.51 ± 0.94 | 0.00 ± 0.00 | 0.00 ± 0.00 | 59.71 ± 0.99 *† | 57.86 | |
| 200 | 49.12 ± 0.94 | 44.33 ± 0.56 | 5.49 ± 0.26 | 1.06 ± 0.31 | 0.00 ± 0.00 | 58.49 ± 0.26 *† | 59.31 | |
| syringic acid | 50 | 39.40 ± 0.23 | 43.76 ± 0.17 | 12.88 ± 0.19 | 3.96 ± 0.00 | 0.00 ± 0.00 | 81.40 ± 1.48 *† | 31.96 |
| 100 | 37.44 ± 0.33 | 50.05 ± 0.59 | 11.46 ± 0.27 | 1.05 ± 0.00 | 0.00 ± 0.00 | 76.12 ± 1.02 *† | 38.26 | |
| 200 | 33.18 ± 0.61 | 59.61 ± 1.28 | 7.21 ± 0.03 | 0.00 ± 0.00 | 0.00 ± 0.00 | 74.03 ± 0.95 *† | 40.76 | |
| p-coumaric acid | 50 | 31.49 ± 0.52 | 61.47 ± 0.55 | 5.40 ± 0.68 | 1.64 ± 0.24 | 0.00 ± 0.00 | 77.19 ± 0.54 *† | 36.99 |
| 100 | 39.57 ± 0.49 | 51.01 ± 0.91 | 6.79 ± 0.32 | 2.63 ± 0.81 | 0.00 ± 0.00 | 72.48 ± 0.29 *† | 42.61 | |
| 200 | 29.62 ± 0.22 | 69.10 ± 0.68 | 1.28 ± 0.08 | 0.00 ± 0.00 | 0.00 ± 0.00 | 71.66 ± 0.33 *† | 43.59 | |
| ferulic acid | 50 | 30.65 ± 0.34 | 59.23 ± 0.67 | 10.12 ± 0.49 | 0.00 ± 0.00 | 0.00 ± 0.00 | 79.47 ± 0.48 *† | 34.26 |
| 100 | 29.32 ± 0.50 | 64.20 ± 1.23 | 6.48 ± 0.33 | 0.00 ± 0.00 | 0.00 ± 0.00 | 77.16 ± 0.97 *† | 37.02 | |
| 200 | 35.89 ± 1.02 | 58.14 ± 1.40 | 5.97 ± 0.25 | 0.00 ± 0.00 | 0.00 ± 0.00 | 70.08 ± 0.90 *† | 45.48 | |
| luteolin-7-O-glucoside | 50 | 29.96 ± 0.46 | 63.83 ± 0.30 | 6.21 ± 0.96 | 0.00 ± 0.00 | 0.00 ± 0.00 | 76.25 ± 0.82 *† | 38.11 |
| 100 | 32.59 ± 0.81 | 60.43 ± 0.26 | 6.98 ± 0.20 | 0.00 ± 0.00 | 0.00 ± 0.00 | 74.39 ± 0.39 *† | 40.33 | |
| 200 | 31.62 ± 1.22 | 63.61 ± 1.00 | 4.77 ± 0.19 | 0.00 ± 0.00 | 0.00 ± 0.00 | 73.15 ± 0.08 *† | 41.81 | |
| quercetin-3-O-glucoside | 50 | 41.76 ± 0.16 | 50.32 ± 0.90 | 7.92 ± 0.05 | 0.00 ± 0.00 | 0.00 ± 0.00 | 66.16 ± 0.67 *† | 50.16 |
| 100 | 44.55 ± 1.06 | 51.77 ± 0.38 | 3.68 ± 0.40 | 0.00 ± 0.00 | 0.00 ± 0.00 | 59.13 ± 0.47 *† | 58.55 | |
| 200 | 56.77 ± 1.31 | 40.77 ± 1.23 | 2.46 ± 0.14 | 0.00 ± 0.00 | 0.00 ± 0.00 | 45.69 ± 0.49 *† | 74.59 | |
| quercetin-3-O-rutinoside | 50 | 47.52 ± 0.96 | 35.05 ± 1.14 | 15.28 ± 1.31 | 2.15 ± 0.01 | 0.00 ± 0.00 | 72.06 ± 0.16 *† | 43.11 |
| 100 | 43.51 ± 0.59 | 44.64 ± 1.30 | 9.57 ± 0.66 | 2.28 ± 0.61 | 0.00 ± 0.00 | 70.62 ± 0.56 *† | 44.83 | |
| 200 | 54.61 ± 0.16 | 32.64 ± 0.27 | 11.69 ± 0.35 | 1.06 ± 0.09 | 0.00 ± 0.00 | 59.20 ± 0.22 *† | 58.46 | |
| rosmarinic acid | 50 | 53.66 ± 1.12 | 38.05 ± 0.93 | 8.29 ± 0.81 | 0.00 ± 0.00 | 0.00 ± 0.00 | 54.63 ± 1.41 *† | 63.92 |
| 100 | 48.16 ± 0.82 | 51.84 ± 0.55 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 51.84 ± 0.55 *† | 67.25 | |
| 200 | 53.71 ± 0.35 | 42.44 ± 0.19 | 3.85 ± 0.47 | 0.00 ± 0.00 | 0.00 ± 0.00 | 50.14 ± 0.68 *† | 69.28 | |
| quercitrin | 50 | 39.30 ± 0.70 | 58.56 ± 1.05 | 2.14 ± 0.64 | 0.00 ± 0.00 | 0.00 ± 0.00 | 62.84 ± 0.46 *† | 54.12 |
| 100 | 45.25 ± 0.24 | 50.76 ± 0.16 | 1.99 ± 0.11 | 0.00 ± 0.00 | 0.00 ± 0.00 | 54.74 ± 0.07 *† | 63.79 | |
| 200 | 55.00 ± 0.61 | 43.88 ± 0.78 | 1.12 ± 0.45 | 0.00 ± 0.00 | 0.00 ± 0.00 | 46.12 ± 0.98 *† | 74.08 | |
| kaempferol-3-O-glucoside | 50 | 39.77 ± 0.33 | 50.75 ± 1.52 | 9.48 ± 0.22 | 0.00 ± 0.00 | 0.00 ± 0.00 | 69.71 ± 0.62 *† | 45.92 |
| 100 | 39.52 ± 0.84 | 57.27 ± 1.03 | 1.96 ± 0.40 | 1.25 ± 0.03 | 0.00 ± 0.00 | 64.94 ± 1.25 *† | 51.61 | |
| 200 | 56.90 ± 0.19 | 35.15 ± 0.65 | 7.95 ± 0.16 | 0.00 ± 0.00 | 0.00 ± 0.00 | 51.05 ± 0.51 *† | 68.19 | |
| quercetin | 50 | 51.28 ± 1.20 | 35.47 ± 0.37 | 10.62 ± 1.46 | 2.63 ± 0.16 | 0.00 ± 0.00 | 64.60 ± 0.68 *† | 52.02 |
| 100 | 57.02 ± 0.62 | 27.99 ± 0.60 | 14.99 ± 0.24 | 0.00 ± 0.00 | 0.00 ± 0.00 | 57.97 ± 0.66 *† | 59.93 | |
| 200 | 63.54 ± 1.48 | 25.47 ± 0.61 | 9.40 ± 0.33 | 1.59 ± 0.08 | 0.00 ± 0.00 | 49.04 ± 0.42 *† | 70.59 | |
| luteolin | 50 | 36.39 ± 0.53 | 55.09 ± 0.95 | 6.49 ± 0.37 | 2.03 ± 0.08 | 0.00 ± 0.00 | 74.16 ± 0.60 *† | 40.60 |
| 100 | 52.45 ± 0.42 | 37.14 ± 0.22 | 9.26 ± 0.14 | 1.15 ± 0.12 | 0.00 ± 0.00 | 59.11 ± 0.93 *† | 58.57 | |
| 200 | 58.75 ± 1.05 | 30.54 ± 0.80 | 10.71 ± 1.07 | 0.00 ± 0.00 | 0.00 ± 0.00 | 51.96 ± 0.30 *† | 67.11 | |
| kaempferol | 50 | 39.43 ± 1.15 | 55.12 ± 0.64 | 5.45 ± 0.16 | 0.00 ± 0.00 | 0.00 ± 0.00 | 66.02 ± 0.92 *† | 50.32 |
| 100 | 50.67 ± 0.50 | 39.23 ± 0.99 | 10.10 ± 0.94 | 0.00 ± 0.00 | 0.00 ± 0.00 | 59.43 ± 0.83 *† | 58.19 | |
| 200 | 55.50 ± 0.72 | 31.32 ± 0.80 | 13.18 ± 0.37 | 0.00 ± 0.00 | 0.00 ± 0.00 | 57.68 ± 0.29 *† | 60.28 | |
| isorhamnetin | 50 | 33.93 ± 0.33 | 56.04 ± 1.20 | 10.03 ± 0.93 | 0.00 ± 0.00 | 0.00 ± 0.00 | 76.10 ± 0.31 *† | 38.29 |
| 100 | 55.12 ± 0.59 | 25.52 ± 0.60 | 16.28 ± 1.17 | 3.08 ± 0.13 | 0.00 ± 0.00 | 67.32 ± 0.10 *† | 48.77 | |
| 200 | 58.30 ± 0.28 | 30.18 ± 0.56 | 11.52 ± 1.09 | 0.00 ± 0.00 | 0.00 ± 0.00 | 53.22 ± 0.87 *† | 65.60 | |
2.3.2. Genotoxic and Antigenotoxic Activities of N. cataria Leaf Methanol Extract (LME)
2.3.3. Genotoxic and Antigenotoxic Activities of N. cataria Stem Methanol Extract (SME)
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material
3.3. Preparation of the Extracts
3.4. Liquid Chromatography-ESI-MS/MS Tandem Mass Spectrometry Analysis
3.5. N. cataria Secondary Metabolites Retrieval
3.6. Animals and Study Design
3.7. Determination of Hepatoprotective Activity
3.7.1. Measurement of Serum Toxicity Markers
3.7.2. Measurement of Liver Homogenate Antioxidant Markers
3.8. Determination of Genotoxic Activity
3.8.1. Detection of DNA Damage
3.8.2. Data Scoring and Photomicrographs
3.9. Statistical Analysis
3.10. Structure-Based Studies
3.10.1. Ligands Modeling
3.10.2. N. cataria Secondary Metabolites Bioactive Conformations Determination within the rCATCmpd I
3.10.3. N. cataria Secondary Metabolites Bioactive Conformations Determination within the rTopIIα
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jamzad, Z.; Grayer, R.J.; Kite, G.C.; Simmonds, M.S.J.; Ingrouille, M.; Jalili, A. Leaf surface flavonoids in Iranian species of Nepeta (Lamiaceae) and some related genera. Biochem. Syst. Ecol. 2003, 31, 587–600. [Google Scholar] [CrossRef]
- Bourrel, C.; Perineau, F.; Michel, G.; Bessiere, J.M. Catnip (Nepeta cataria L.) essential oil: Analysis of chemical constituents, bacteriostatic and fungistatic properties. J. Essent. Oil Res. 1993, 5, 159–167. [Google Scholar] [CrossRef]
- Sharma, A.; Cooper, R.; Bhardwaj, G.; Cannoo, D.S. The genus Nepeta: Traditional uses, phytochemicals and pharmacological properties. J. Ethnopharmacol. 2021, 268, 113679. [Google Scholar] [CrossRef] [PubMed]
- Formisano, C.; Rigano, D.; Senatore, F. Chemical constituents and biological activities of Nepeta species. Chem. Biodivers. 2011, 8, 1783–1818. [Google Scholar] [CrossRef]
- Salehi, P.; Sonboli, A.; Allahyari, L. Antibacterial and antioxidant properties of the essential oil and various extracts of Nepeta ispahanica from Iran. J. Essent. Oil Bear. Plants 2007, 10, 324–331. [Google Scholar] [CrossRef]
- Zomorodian, K.; Saharkhiz, M.J.; Shariati, S.; Pakshir, K.; Rahimi, M.J.; Khashei, R. Chemical composition and antimicrobial activities of essential oils from Nepeta cataria L. against common causes of food-borne infections. ISRN Pharm. 2012, 2012, 591953. [Google Scholar] [CrossRef]
- Suntar, I.; Nabavi, S.M.; Barreca, D.; Fischer, N.; Efferth, T. Pharmacological and chemical features of Nepeta L. genus: Its importance as a therapeutic agent. Phytother. Res. 2017, 32, 185–198. [Google Scholar] [CrossRef]
- Mišić, D.; Šiler, B.; Gašić, U.; Avramov, S.; ŽivkoviĆ, S.; Živković, J.N.; Milutinović, M.; Tešić, Ž. Simultaneous UHPLC/DAD/(+/-)HESI-MS/MS Analysis of phenolic acids and nepetalactones in methanol extracts of Nepeta species: A possible application in chemotaxonomic studies. Phytochem. Anal. 2015, 26, 72–85. [Google Scholar] [CrossRef]
- Lichman, B.R.; Godden, G.T.; Hamilton, J.P.; Palmer, L.; Kamileen, M.O.; Zhao, D.; Vaillancourt, B.; Wood, J.C.; Sun, M.; Kinser, T.J.; et al. The evolutionary origins of the cat attractant nepetalactone in catnip. Sci. Adv. 2020, 6, eaba0721. [Google Scholar] [CrossRef]
- Tagawa, M.; Murai, F. A new iridoid glucoside, nepetolglucosylester from Nepeta cataria. Planta Med. 1980, 39, 144–147. [Google Scholar] [CrossRef]
- Xie, S.; Uesato, S.; Inouye, H.; Fujita, T.; Murai, F.; Tagawa, M.; Shingu, T. Absolute structure of nepetaside, a new iridoid glucoside from Nepeta cataria. Phytochemistry 1988, 27, 469–472. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: An overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef] [PubMed]
- Eid, H.H.; Labib, R.M.; Hamid, N.S.A.; Hamed, M.A.; Ross, S.A. Hepatoprotective and antioxidant polyphenols from a standardized methanolic extract of the leaves of Liquidambar styraciflua L. Bull. Fac. Pharm. Cairo Univ. 2015, 53, 117–127. [Google Scholar] [CrossRef]
- Bojić, M.; Maleš, Ž.; Antolić, A.; Babić, I.; Tomičić, M. Antithrombotic activity of flavonoids and polyphenols rich plant species. Acta Pharm. 2019, 69, 483–495. [Google Scholar] [CrossRef] [PubMed]
- de Araujo, F.F.; de Paulo Farias, D.; Neri-Numa, I.A.; Pastore, G.M. Polyphenols and their applications: An approach in food chemistry and innovation potential. Food Chem. 2021, 338, 127535. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and human health: The role of bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Sharma, A.; Nayik, G.A.; Cannoo, D.S. Pharmacology and Toxicology of Nepeta cataria (Catmint) Species of Genus Nepeta: A Review. In Plant and Human Health; Ozturk, M., Hakeem, K., Eds.; Springer: Cham, Switzerland, 2018; Volume 3. [Google Scholar] [CrossRef]
- Smitherman, L.C.; Janisse, J.; Mathur, A. The use of folk remedies among children in an urban black community: Remedies for fever, colic, and teething. Pediatrics 2005, 115, e297–e304. [Google Scholar] [CrossRef]
- Hatch, R.C. Effect of drugs on catnip (Nepeta cataria)-induced pleasure behavior in cats. Am. J. Vet. Res. 1972, 33, 143–155. [Google Scholar]
- Jackson, B.; Reed, A. Catnip and the alteration of consciousness. J. Am. Med. Assoc. 1969, 207, 1349–1350. [Google Scholar] [CrossRef]
- Barcelos, G.R.M.; Grotto, D.; Angeli, J.P.F.; Serpeloni, J.M.; Rocha, B.A.; Bastos, J.K.; Barbosa, F. Evaluation of antigenotoxic effects of plant flavonoids quercetin and rutin on HepG2 cells. Phytother. Res. 2011, 25, 1381–1388. [Google Scholar] [CrossRef]
- Lee, S.; Lee, J.; Lee, H.; Sung, J. Relative protective activities of quercetin, quercetin-3-glucoside, and rutin in alcohol-induced liver injury. J. Food Biochem. 2019, 43, e13002. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Sharma, A.; Talukder, G. Plant extracts as modulators of genotoxic effects. Bot. Rev. 1996, 62, 275–300. [Google Scholar] [CrossRef]
- Modnicki, D.; Tokar, M.; Klimek, B. Flavonoids and phenolic acids of Nepeta cataria L. var. citriodora (Becker) BALB. (Lamiaceae). Acta Pol. Pharm. 2007, 64, 247–252. [Google Scholar] [PubMed]
- Del Carmen García-Rodríguez, M.; Carvente-Juarez, M.M.; Altamirano-Lozano, M.A. Antigenotoxic and apoptotic activity of green tea polyphenol extracts on hexavalent chromium-induced DNA damage in peripheral blood of CD-1 mice: Analysis with differential acridine orange/ethidium bromide staining. Oxid. Med. Cell. Longev. 2013, 2013, 486419. [Google Scholar] [CrossRef]
- Kaur, P.; Walia, A.; Kumar, S.; Kaur, S. Antigenotoxic activity of polyphenolic rich extracts from Aegle marmelos (L.) Correa in human blood lymphocytes and E. coli PQ 37. Rec. Nat. Prod. 2009, 3, 68–75. [Google Scholar]
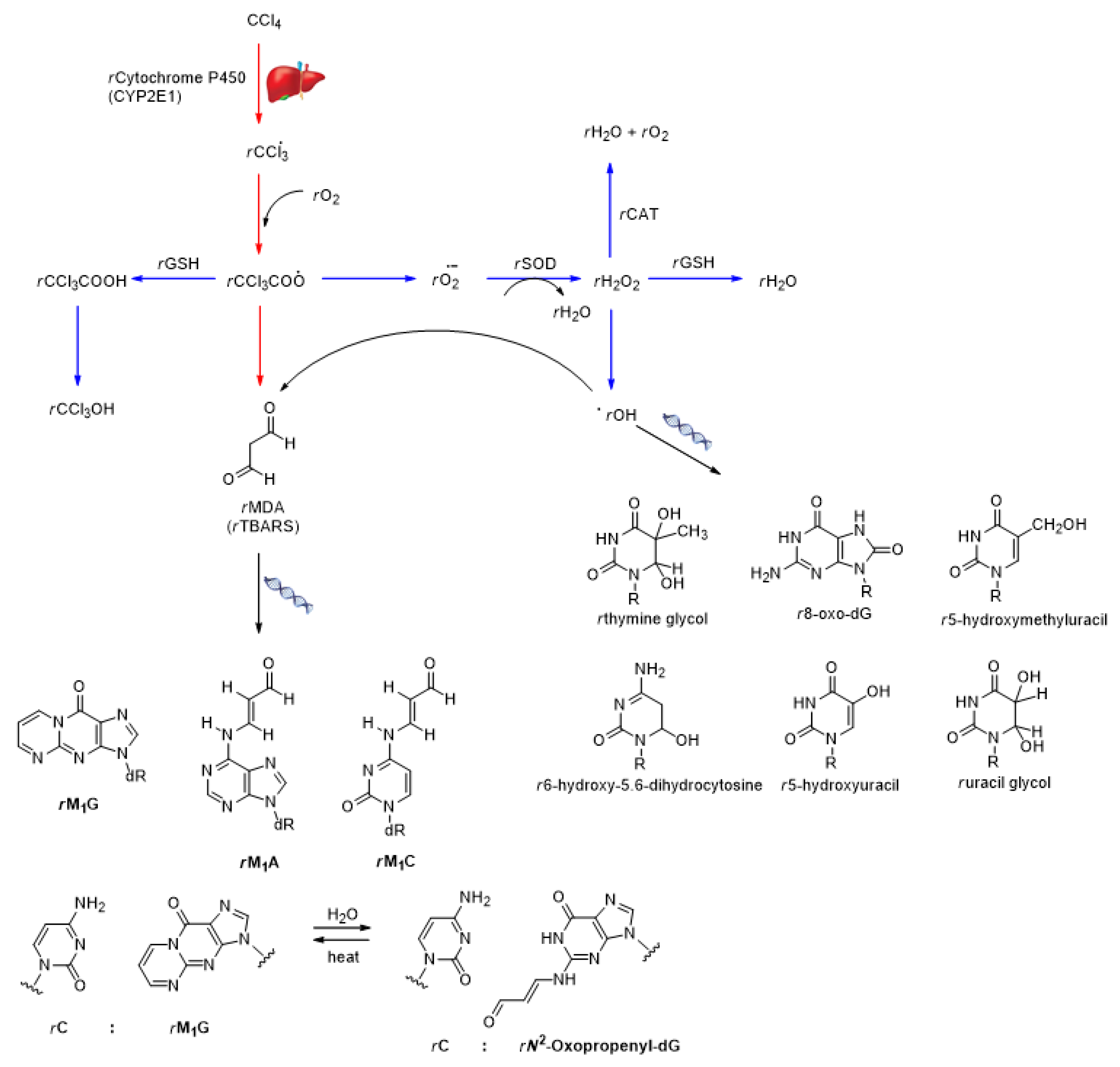
- Marnett, L.J. Oxyradicals and DNA damage. Carcinogenesis 2000, 21, 361–370. [Google Scholar] [CrossRef]
- Higuchi, H.; Gores, G.J. Mechanisms of liver injury: An overview. Curr. Mol. Med. 2003, 3, 483–490. [Google Scholar] [CrossRef]
- Hensley, K.; Robinson, K.; Gabbita, S.P.; Salsman, S.; Floyd, R.A. Reactive oxygen species, cell signaling, and cell injury. Free Radic. Biol. Med. 2000, 28, 1456–1462. [Google Scholar] [CrossRef]
- Nandita, S.; Vasudeva, K.; Narasimhamurthy, K.; Rajini, P.S. Protective effect of potato peel extracts against carbon tetrachloride-induced liver injury in rats. Environ. Toxicol. Pharmacol. 2008, 26, 241–246. [Google Scholar] [CrossRef]
- Hubert, D.J.; Dawe, A.; Florence, N.T.; Gilbert, K.D.; Angele, T.N.; Buonocore, D.; Finzi, P.V.; Vidari, G.; Bonaventure, N.T.; Marzatico, F.; et al. In vitro hepatoprotective and antioxidant activities of crude extract and isolated compounds from Ficus gnaphalocarpa. Inflammopharmacol. 2011, 19, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Esmat, A.Y.; Said, M.M.; Soliman, A.A.; El-Masry, K.S.; Badiea, E.A. Bioactive compounds, antioxidant potential, and hepatoprotective activity of sea cucumber (Holothuria atra) against thioacetamide intoxication in rats. Nutrition 2013, 29, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Hong, C.O.; Lee, G.P.; Kim, C.T.; Lee, K.W. The hepatoprotection of caffeic acid and rosmarinic acid, major compounds of Perilla frutescens, against t-BHP-induced oxidative liver damage. Food Chem. Toxicol. 2013, 55, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Szende, B.; Timár, F.; Hargitai, B. Olive oil decreases liver damage in rats caused by carbon tetrachloride (CCl4). Exp. Toxicol. Pathol. 1994, 46, 355–359. [Google Scholar] [CrossRef]
- Abdel-Wahhab, K.G.E.D.; El-Shamy, K.A.; El-Beih, N.A.E.Z.; Morcy, F.A.; Mannaa, F.A.E. Protective effect of a natural herb (Rosmarinus officinalis) against hepatotoxicity in male albino rats. Comun. Sci. 2011, 2, 9–17. [Google Scholar] [CrossRef]
- Carini, R.; Chiarpotto, E.; Biasi, F.; Leonarduzzi, G.; Comoglio, A.; Carpi, C.; Poli, G. Relation between liver necrosis and intrahepatic cholestasis in rats poisoned with CCl4. Boll. Soc. Ital. Biol. Sper. 1987, 63, 273–280. [Google Scholar]
- Milani, L.; Galindo, C.M.; de Oliveira, N.M.T.; Corso, C.R.; Adami, E.R.; Stipp, M.C.; Beltrame, O.C.; Acco, A. The GLP-1 analog liraglutide attenuates acute liver injury in mice. Ann. Hepatol. 2019, 18, 918–928. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the auto-oxidation of epinephrine and simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Goth, L.A. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–152. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Stickel, F.; Seitz, H.K.; Hahn, E.G.; Schuppan, D. Liver toxicity of drugs of plant origin. Z. Gastroenterol. 2001, 39, 225–232, 234–237. [Google Scholar] [CrossRef] [PubMed]
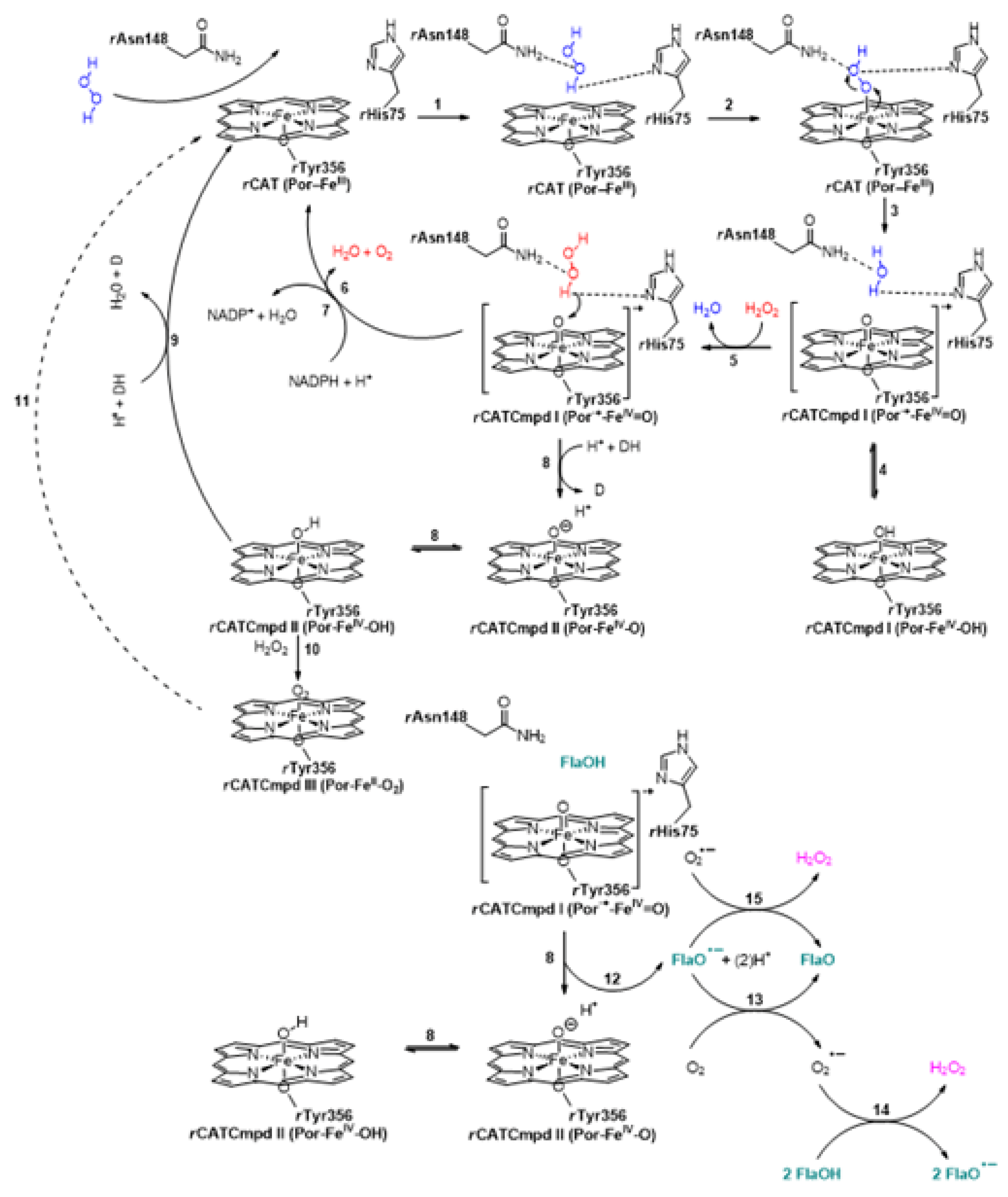
- Putnam, C.D.; Arvai, A.S.; Bourne, Y.; Tainer, J.A. Active and inhibited human catalase structures: Ligand and NADPH binding and catalytic mechanism. J. Mol. Biol. 2000, 296, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Kirkman, H.N.; Rolfo, M.; Ferraris, A.M.; Gaetani, G.F. Mechanisms of protection of catalase by NADPH. Kinetics and stoichiometry. J. Biol. Chem. 1999, 274, 13908–13914. [Google Scholar] [CrossRef] [PubMed]
- Switala, J.; Loewen, P.C. Diversity of properties among catalases. Arch. Biochem. Biophys. 2002, 401, 145–154. [Google Scholar] [CrossRef]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry, 6th ed.; W.H. freeman: New York, NY, USA, 2007. [Google Scholar]
- Alfonso-Prieto, M.; Vidossich, P.; Rovira, C. The reaction mechanisms of heme catalases: An atomistic view by ab initio molecular dynamics. Arch. Biochem. Biophys. 2012, 525, 121–130. [Google Scholar] [CrossRef]
- Diaz, A.; Loewen, P.C.; Fita, I.; Carpena, X. Thirty years of heme catalases structural biology. Arch. Biochem. Biophys. 2012, 525, 102–110. [Google Scholar] [CrossRef]
- Nicholls, P. Classical catalase: Ancient and modern. Arch. Biochem. Biophys. 2012, 525, 95–101. [Google Scholar] [CrossRef]
- Zamocky, M.; Koller, F. Understanding the structure and function of catalases: Clues from molecular evolution and in vitro mutagenesis. Prog. Biophys. Mol. Biol. 1999, 72, 19–66. [Google Scholar] [CrossRef]
- Alfonso-Prieto, M.; Vidossich, P.; Rodríguez-Fortea, A.; Carpena, X.; Fita, I.; Loewen, P.C.; Rovira, C. Electronic state of the molecular oxygen released by catalase. J. Phys. Chem. A 2008, 112, 12842–12848. [Google Scholar] [CrossRef]
- Rovira, C. Structure, protonation state and dynamics of catalase compound II. ChemPhysChem. 2005, 6, 1820–1826. [Google Scholar] [CrossRef] [PubMed]
- Vidossich, P.; Alfonso-Prieto, M.; Carpena, X.; Loewen, P.C.; Fita, I.; Rovira, C. Versatility of the electronic structure of compound I in catalase-peroxidases. J. Am. Chem. Soc. 2007, 129, 13436–13446. [Google Scholar] [CrossRef]
- Krych, J.; Gebicki, J.L.; Gebicka, L. Flavonoid-induced conversion of catalase to its inactive form—Compound II. Free Radic. Res. 2014, 48, 1334–1341. [Google Scholar] [CrossRef] [PubMed]
- Lala, V.; Goyal, A.; Bansal, P.; Minter, D.A. StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482489/ (accessed on 30 June 2021).
- Giannini, E.G.; Testa, R.; Savarino, V. Liver enzyme alteration: A guide for clinicians. CMAJ 2005, 172, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; Mc Coy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1998, 175, 184–191. [Google Scholar] [CrossRef]
- Cadet, J.; Delatour, T.; Douki, T.; Gasparutto, D.; Pouget, J.-P.; Ravanat, J.-L.; Sauvaigo, S. Hydroxyl radicals and DNA base damage. Mut. Res. 1999, 424, 9–21. [Google Scholar] [CrossRef]
- Mladenović, M.; Stanković, N.; Matić, S.; Stanić, S.; Mihailović, M.; Mihailović, V.; Katanić, J.; Boroja, T.; Vuković, N. Newly discovered chroman-2,4-diones neutralize the in vivo DNA damage induced by alkylation through the inhibition of Topoisomerase Iiα: A story behind the molecular modeling approach. Biochem. Pharmacol. 2015, 98, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Orcic, D.; Franciskovic, M.; Bekvalac, K.; Svircev, E.; Beara, I.; Lesjak, M.; Mimica-Dukic, N. Quantitative determination of plant phenolics in Urtica dioica extracts by high-performance liquid chromatography coupled with tandem mass spectrometric detection. Food Chem. 2014, 143, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Vukić, M.D.; Vuković, N.L.; Djelić, G.T.; Obradović, A.; Kacaniova, M.M.; Marković, S.; Popović, S.; Baskić, D. Phytochemical analysis, antioxidant, antibacterial and cytotoxic activity of different plant organs of Eryngium serbicum L. Ind. Crops Prod. 2018, 115, 88–97. [Google Scholar] [CrossRef]
- Quick, A.J.; Stanley-Brown, M.; Bancroft, F.W. A study of the coagulation defect in hemophilia and in jaundice. Am. J. Med. Sci. 1935, 190, 501–511. [Google Scholar] [CrossRef]
- Bergmeyer, H.U.; Bowers, G.N.; Horder, M.; Moss, D.W. Provisional recommendations on IFCC Methods for the measurement of catalytic concentrations of enzymes. Part 2. IFCC method for aspartat aminotransferase. Clin. Chim. Acta 1976, 70, 19–42. [Google Scholar] [CrossRef]
- Bergmeyer, H.U.; Horder, M. IFCC methods for measurement of catalytic concentrations of enzymes. Part 3. IFCC. Method for alanine aminotransferase (l-alanine 2 -oxoglutarate aminotransferase, ec 2.6.1.2). Clin. Chim. Acta 1980, 105, F145–F172. [Google Scholar]
- Walters, M.I.; Gerarde, H.W. An ultramicromethod for the determination of conjugated and total bilirubin in serum or plasma. Microchem. J. 1970, 15, 231–243. [Google Scholar] [CrossRef]
- Schumann, G.; Bonora, R.; Ceriotti, F.; Ferard, G.; Ferrero, C.A.; Franck, P.F.; Gella, F.J.; Hoelzel, W.; Jorgensen, P.J.; Kanno, T.; et al. IFCC Primary reference procedures for the measurement of catalytic activity concentrations of enzymes at 37 °C. Part 6. Reference procedure for the measurement of catalytic concentration of γ-glutamyltransferase. Clin. Chem. Lab. Med. 2005, 40, 734–738. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Hartmann, A.; Speit, G. The contribution of cytotoxicity to DNA effects in the single cell gel test (Comet assay). Toxicol. Lett. 1997, 90, 183–188. [Google Scholar] [CrossRef]
- Collins, A.R.; Oscoz, A.A.; Brunborg, G.; Gaivao, I.; Giovannelli, L.; Kruszewski, M.; Smith, C.C.; Stetina, R. The comet assay: Topical issues. Mutagenesis 2008, 23, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, K.; Banerjee, M.R. β-Carotene reduces sister chromatid exchange induce chemical carcinogens in mouse mammary cells in organ culture. Cell Biol. Int. Rep. 1985, 9, 783–789. [Google Scholar] [CrossRef]
- Waters, M.D.; Brady, A.L.; Stack, H.F.; Brockman, H.E. Antimutagenicity profiles for some models compounds. Mutat. Res. 1990, 238, 57–85. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef]
- Marvin Beans; 15.4.27.0; ChemAxon: Budapest, Hungary, 2015.
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.A.; Olson, J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Case, D.A.; Darden, T.A.; Cheatham III, T.E.; Simmerling, C.L.; Wang, J.; Duke, R.E.; Luo, R.; Walker, R.C.; Zhang, W.; Merz, K.M.; et al. AMBER 12; University of California: San Francisco, CA, USA, 2012. [Google Scholar]





| Concentration (μg/g) 1 | |||||
|---|---|---|---|---|---|
| No | Compounds | tR 2 | FME 3 | LME 4 | SME 5 |
| 1 | quinic acid | 0.49 | 536.11 ± 6.12 6 | 196.33 ± 2.11 | 113.16 ± 2.03 |
| 2 | protocatechuic acid | 0.79 | 100.51 ± 0.35 | 261.42 ± 0.38 | 71.64 ± 0.34 |
| 3 | chlorogenic acid | 0.80 | 1647.32 ± 8.12 | 648.52 ± 6.15 | 1408.43 ± 8.01 |
| 4 | p-hydroxybenzoic acid | 1.08 | 138.21 ± 2.15 | 363.32 ± 5.32 | 42.24 ± 2.99 |
| 5 | caffeic acid | 1.18 | 80.13 ± 2.13 | 137.03 ± 2.69 | 42.01 ± 2.01 |
| 6 | syringic acid | 1.31 | 15.93 ± 0.87 | 36.34 ± 1.29 | 30.32 ± 1.19 |
| 7 | p-coumaric acid | 1.69 | 72.84 ± 2.45 | 138.22 ± 2.87 | 38.64 ± 1.99 |
| 8 | ferulic acid | 1.90 | 129.23 ± 1.54 | 155.64 ± 1.56 | 55.84 ± 1.12 |
| 9 | luteolin-7-O-glucoside | 2.13 | 7.64 ± 0.36 | 5.43 ± 0.34 | 5.96 ± 0.34 |
| 10 | quercetin-3-O-glucoside | 2.25 | 72.97 ± 4.23 | 354.33 ± 6.59 | 87.97 ± 4.55 |
| 11 | quercetin-3-O-rutinoside | 2.33 | 86.55 ± 1.99 | 98.71 ± 2.11 | 114.01 ± 2.16 |
| 12 | rosmarinic acid | 2.42 | 326.15 ± 1.19 | 1056.14 ± 2.69 | 250.54 ± 1.18 |
| 13 | quercitrin | 2.75 | 8406.31 ± 12.59 | 26.21 ± 0.45 | 1.02 ± 0.03 |
| 14 | kaempferol-3-O-glucoside | 2.80 | 409.21 ± 15.33 | 374.13 ± 15.11 | 168.54 ± 9.23 |
| 15 | quercetin | 3.74 | 88.97 ± 3.25 | 78.33 ± 3.19 | 94.54 ± 3.26 |
| 16 | luteolin | 4.03 | 6.12 ± 0.45 | 4.13 ± 0.36 | 3.96 ± 0.39 |
| 17 | kaempferol | 4.55 | 90.03 ± 1.59 | 82.87 ± 1.54 | 67.25 ± 1.49 |
| 18 | isorhamnetin | 4.59 | 107.53 ± 1.11 | 175.34 ± 1.14 | 117.24 ± 1.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vukić, M.D.; Vuković, N.L.; Mladenović, M.; Tomašević, N.; Matić, S.; Stanić, S.; Sapienza, F.; Ragno, R.; Božović, M.; Kačániová, M. Chemical Composition of Various Nepeta cataria Plant Organs’ Methanol Extracts Associated with In Vivo Hepatoprotective and Antigenotoxic Features as well as Molecular Modeling Investigations. Plants 2022, 11, 2114. https://doi.org/10.3390/plants11162114
Vukić MD, Vuković NL, Mladenović M, Tomašević N, Matić S, Stanić S, Sapienza F, Ragno R, Božović M, Kačániová M. Chemical Composition of Various Nepeta cataria Plant Organs’ Methanol Extracts Associated with In Vivo Hepatoprotective and Antigenotoxic Features as well as Molecular Modeling Investigations. Plants. 2022; 11(16):2114. https://doi.org/10.3390/plants11162114
Chicago/Turabian StyleVukić, Milena D., Nenad L. Vuković, Milan Mladenović, Nevena Tomašević, Sanja Matić, Snežana Stanić, Filippo Sapienza, Rino Ragno, Mijat Božović, and Miroslava Kačániová. 2022. "Chemical Composition of Various Nepeta cataria Plant Organs’ Methanol Extracts Associated with In Vivo Hepatoprotective and Antigenotoxic Features as well as Molecular Modeling Investigations" Plants 11, no. 16: 2114. https://doi.org/10.3390/plants11162114