
The Main Protease of SARS-CoV-2 as a Target for Phytochemicals against Coronavirus


Abstract
:1. Introduction
2. SARS-CoV-2 Structure and the Main Protease of SARS-CoV-2 as a Potential Protein Target
3. Standard Approaches That Could Be Applied in the Search for Mpro Potential Inhibitors
3.1. Virtual In Silico Screening: (The Data-Driven Approach)
3.2. The Classical Approach of High-Throughput Screening (HTS)
3.3. Antiviral Activity Cell-Based Assays
4. Phytochemicals as a Reservoir to Search for SARS-CoV-2 Mpro Potential Inhibitors
5. Perspectives of Large-Scale Synthesis of Anti-SARS-CoV-2 Compounds
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, P.I.; Hsueh, P.R. Emerging threats from zoonotic coronaviruses-from SARS and MERS to 2019-nCoV. J. Microbiol. Immunol. Infect. 2020, 53, 365–367. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Feng, Z. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus-Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.; Wang, W.; Song, Z.; Hu, Y.; Tao, Z.; Tian, J.; Pei, Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Morales, A.J.; MacGregor, K.; Kanagarajah, S.; Patel, D.; Schlagenhauf, P. Going global—Travel and the 2019 novel coronavirus. Travel Med. Infect. Dis. 2020, 33, 101578. [Google Scholar] [CrossRef] [PubMed]
- Silveira, M.M.; Moreira, G.M.S.G.; Mendonça, M. DNA vaccines against COVID-19: Perspectives and challenges. Life Sci. 2020, 267, 118919. [Google Scholar] [CrossRef] [PubMed]
- WHO. COVID-19 Dashboard. 2022. Available online: https://covid19.who.int/ (accessed on 5 May 2022).
- Chen, P.; Shi, X.; He, W.; Zhong, G.; Tang, Y.; Wang, H.; Zhang, P. mRNA vaccine-a desirable therapeutic strategy for surmounting COVID-19 pandemic. Hum. Vaccines Immunother. 2022, 18, 2040330. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Yang, H. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.Z.; Yasuo, N.; Sekijima, M. Screening for Inhibitors of Main Protease in SARS-CoV-2: In Silico and In Vitro Approach Avoiding Peptidyl Secondary Amides. J. Chem. Inf. Model. 2022, 62, 350–358. [Google Scholar] [CrossRef]
- Hossain, M.J.; Rabaan, A.A.; Mutair, A.A.; Alhumaid, S.; Emran, T.B.; Saikumar, G.; Dhama, K. Strategies to tackle SARS-CoV-2 Mu, a newly classified variant of interest likely to resist currently available COVID-19 vaccines. Hum. Vaccin. Immunother. 2022, 18, 2027197. [Google Scholar] [CrossRef]
- Konings, F.; Perkins, M.D.; Kuhn, J.H.; Pallen, M.J.; Alm, E.J.; Archer, B.N.; Van Kerkhove, M.D. SARS-CoV-2 Variants of Interest and Concern naming scheme conducive for global discourse. Nat. Microbiol. 2021, 6, 821–823. [Google Scholar] [CrossRef] [PubMed]
- Sharun, K.; Tiwari, R.; Dhama, K.; Emran, T.B.; Rabaan, A.A.; Al Mutair, A. Emerging SARS-CoV-2 variants: Impact on vaccine efficacy and neutralizing antibodies. Hum. Vaccin. Immunother. 2021, 17, 3491–3494. [Google Scholar] [CrossRef] [PubMed]
- Van Egeren, D.; Novokhodko, A.; Stoddard, M.; Tran, U.; Zetter, B.; Rogers, M.; Chakravarty, A. Risk of rapid evolutionary escape from biomedical interventions targeting SARS-CoV-2 spike protein. PLoS ONE 2021, 16, e0250780. [Google Scholar] [CrossRef] [PubMed]
- Brant, A.C.; Tian, W.; Majerciak, V.; Yang, W.; Zheng, Z.M. SARS-CoV-2: From its discovery to genome structure, transcription, and replication. Cell Biosci. 2021, 11, 136. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Zhang, B.; Jiang, X.M.; Su, H.; Li, J.; Zhao, Y.; Liu, H. Structure-based design of antiviral drug candidates targeting the SARS-CoV-2 main protease. Science 2020, 368, 1331–1335. [Google Scholar] [CrossRef] [Green Version]
- Ramajayam, R.; Tan, K.-P.; Liang, P.-H. Recent development of 3C and 3CL protease inhibitors for anti-coronavirus and anti-picornavirus drug discovery. Biochem. Soc. Trans. 2011, 39, 1371–1375. [Google Scholar] [CrossRef]
- Cao, Y.; Yang, R.; Lee, I.; Zhang, W.; Sun, J.; Wang, W.; Meng, X. Characterization of the SARS-CoV-2 E Protein: Sequence, Structure, Viroporin, and Inhibitors. Protein Sci. 2021, 30, 1114–1130. [Google Scholar] [CrossRef]
- Kim, D.; Lee, J.Y.; Yang, J.S.; Kim, J.W.; Kim, V.N.; Chang, H. The Architecture of SARS-CoV-2 Transcriptome. Cell 2020, 181, 914–921.e10. [Google Scholar] [CrossRef]
- Reiner, Ž.; Hatamipour, M.; Banach, M.; Pirro, M.; Al-Rasadi, K.; Jamialahmadi, T.; Radenkovic, D.; Montecucco, F.; Sahebkar, A. Statins and the COVID-19 main protease: In silico evidence on direct interaction. Arch. Med. Sci. 2020, 16, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, K.M.; Zhang, H.; Senthil, R.; Vijayakumar, K.K.; Sounderrajan, V.; Wei, Y.; Shakila, H. Structural basis for the inhibition of SARS-CoV2 main protease by Indian medicinal plant-derived antiviral compounds. J. Biomol. Struct. Dyn. 2022, 40, 1970–1978. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xie, W.; Xue, X.; Yang, K.; Ma, J.; Liang, W.; Rao, Z. Design of wide-spectrum inhibitors targeting coronavirus main proteases. PLoS Biol. 2005, 3, e324. [Google Scholar]
- Rocha, R.E.; Chaves, E.J.; Fischer, P.H.; Costa, L.S.; Grillo, I.B.; da Cruz, L.E.; de Lima, L.H. A higher flexibility at the SARS-CoV-2 main protease active site compared to SARS-CoV and its potentialities for new inhibitor virtual screening targeting multi-conformers. J. Biomol. Struct. Dyn. 2021, 10, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Delaunay-Moisan, A.; Timmis, K.; Danchin, A. SARS-CoV-2 biology and variants: Anticipation of viral evolution and what needs to be done. Environ. Microbiol. 2021, 23, 2339–2363. [Google Scholar] [CrossRef]
- Mótyán, J.A.; Mahdi, M.; Hoffka, G.; Tőzsér, J. Potential Resistance of SARS-CoV-2 Main Protease (Mpro) against Protease Inhibitors: Lessons Learned from HIV-1 Protease. Int. J. Mol. Sci. 2022, 23, 3507. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Rendine, N.; Mohammed MK, S.; Jeeva, A.; Ji, H.F.; Talluri, V.R. Structure-based virtual screening, in silico docking, ADME properties prediction and molecular dynamics studies for the identification of potential inhibitors against SARS-CoV-2 M(pro). Mol. Divers. 2021, 26, 1645–1661. [Google Scholar] [CrossRef]
- Morris, G.M.; Lim-Wilby, M. Molecular docking. Methods Mol. Biol. 2008, 443, 365–382. [Google Scholar]
- Sari, S.; Avci, A.; Aslan, E.K. In silico Repurposing of Drugs for pan-HDAC and pan-SIRT Inhibitors: Consensus Structure-based Virtual Screening and Pharmacophore Modeling Investigations. Turk. J. Pharm. Sci. 2021, 18, 730–737. [Google Scholar] [CrossRef]
- Pinzi, L.; Rastelli, G. Molecular Docking: Shifting Paradigms in Drug Discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef] [Green Version]
- Bonanno, E.; Ebejer, J.-P. Applying Machine Learning to Ultrafast Shape Recognition in Ligand-Based Virtual Screening. Front. Pharmacol. 2020, 10, 1675. [Google Scholar] [CrossRef] [PubMed]
- Muegge, I.; Mukherjee, P. An overview of molecular fingerprint similarity search in virtual screening. Expert Opin. Drug Discov. 2015, 11, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Pinto, G.P.; Hendrikse, N.M.; Stourac, J.; Damborsky, J.; Bednar, D. Virtual screening of potential anticancer drugs based on microbial products. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2021. [Google Scholar]
- Pinto, G.P.; Vavra, O.; Filipovic, J.; Stourac, J.; Bednar, D.; Damborsky, J. Fast Screening of Inhibitor Binding/Unbinding Using Novel Software Tool CaverDock. Front. Chem. 2019, 7, 709. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Sarkar, C.; El-Kersh, D.M.; Jamaddar, S.; Uddin, S.J.; Shilpi, J.A.; Mubarak, M.S. Natural products and their derivatives against coronavirus: A review of the non-clinical and pre-clinical data. Phytother. Res. 2020, 34, 2471–2492. [Google Scholar] [CrossRef] [PubMed]
- Olubiyi, O.O.; Olagunju, M.; Keutmann, M.; Loschwitz, J.; Strodel, B. High Throughput Virtual Screening to Discover Inhibitors of the Main Protease of the Coronavirus SARS-CoV-2. Molecules 2020, 25, 3193. [Google Scholar] [CrossRef]
- Zhang, D.-H.; Wu, K.-L.; Zhang, X.; Deng, S.-Q.; Peng, B. In silico screening of Chinese herbal medicines with the potential to directly inhibit 2019 novel coronavirus. J. Integr. Med. 2020, 18, 152–158. [Google Scholar] [CrossRef]
- Joshi, R.S.; Jagdale, S.S.; Bansode, S.B.; Shankar, S.S.; Tellis, M.B.; Pandya, V.K.; Kulkarni, M.J. Discovery of potential multi-target-directed ligands by targeting host-specific SARS-CoV-2 structurally conserved main protease. J. Biomol. Struct. Dyn. 2021, 39, 3099–3114. [Google Scholar] [CrossRef] [Green Version]
- Tallei, T.E.; Tumilaar, S.G.; Niode, N.J.; Kepel, B.J.; Idroes, R.; Effendi, Y.; Emran, T.B. Potential of Plant Bioactive Compounds as SARS-CoV-2 Main Protease (Mpro) and Spike (S) Glycoprotein Inhibitors: A Molecular Docking Study. Scientifica 2020, 2020, 6307457. [Google Scholar] [CrossRef]
- Ul Qamar, M.T.; Alqahtani, S.M.; Alamri, M.A.; Chen, L.L. Structural basis of SARS-CoV-2 3CL(pro) and anti-COVID-19 drug discovery from medicinal plants. J. Pharm. Anal. 2020, 10, 313–319. [Google Scholar] [CrossRef]
- Khaerunnisa, S.; Kurniawan, H.; Awaluddin, R.; Suhartati, S.; Soetjipto, S. Potential Inhibitor of COVID-19 Main Protease (Mpro) from Several Medicinal Plant Compounds by Molecular Docking Study. Preprints 2020, 2020, 2020030226. [Google Scholar]
- da Silva JK, R.; Figueiredo PL, B.; Byler, K.G.; Setzer, W.N. Essential Oils as Antiviral Agents, Potential of Essential Oils to Treat SARS-CoV-2 Infection: An In-Silico Investigation. Int. J. Mol. Sci. 2020, 21, 3426. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.E.; Tawfeek, N.; Elbaramawi, S.S.; Fikry, E. Agathis robusta Bark Essential Oil Effectiveness against COVID-19: Chemical Composition, In Silico and In Vitro Approaches. Plants 2022, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- My, T.T.A.; Loan, H.T.P.; Hai, N.T.T.; Hieu, L.T.; Hoa, T.T.; Thuy, B.T.P.; Quang, D.T.; Triet, N.T.; Van Anh, T.T.; Dieu, N.T.X.; et al. Evaluation of the Inhibitory Activities of COVID-19 of Melaleuca cajuputi Oil Using Docking Simulation. ChemistrySelect 2020, 5, 6312–6320. [Google Scholar] [CrossRef] [PubMed]
- Attene-Ramos, M.S.; Austin, C.P.; Xia, M. High Throughput Screening. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; pp. 916–917. [Google Scholar]
- Ismail, F.M.; Nahar, L.; Sarker, S.D. High-Throughput Screening of Phytochemicals: Application of Computational Methods. Comput. Phytochem. 2018, 165–192. [Google Scholar]
- Lundblad, R.L. Drug Design. In Encyclopedia of Cell Biology; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Hajare, A.A.; Salunkhe, S.S.; Mali, S.S.; Gorde, S.S.; Nadaf, S.J.; Pishawikar, S.A. Review on: High-throughput screening is an approach to drug discovery. Am. J. Pharm. Tech. Res. 2013, 4, 112–129. [Google Scholar]
- Liu, J.; Li, K.; Cheng, L.; Shao, J.; Yang, S.; Zhang, W.; Zhou, G.; de Vries, A.A.; Yu, Z. A high-throughput drug screening strategy against coronaviruses. Int. J. Infect. Dis. 2020, 103, 300–304. [Google Scholar] [CrossRef]
- Xu, T.; Zheng, W.; Huang, R. High-throughput screening assays for SARS-CoV-2 drug development: Current status and future directions. Drug Discov. Today 2021, 26, 2439–2444. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Deng, C.L.; Liu, J.; Li, J.Q.; Zhang, H.Q.; Li, N.; Ye, H.Q. SARS-CoV-2 replicon for high-throughput antiviral screening. J. Gen. Virol. 2021, 102, 001583. [Google Scholar] [CrossRef]
- Zhao, Y.; Du, X.; Duan, Y.; Pan, X.; Sun, Y.; You, T.; Yang, H. High-throughput screening identifies established drugs as SARS-CoV-2 PLpro inhibitors. Protein Cell 2021, 12, 877–888. [Google Scholar] [CrossRef]
- Zhu, W.; Xu, M.; Chen, C.Z.; Guo, H.; Shen, M.; Hu, X.; Zheng, W. Identification of SARS-CoV-2 3CL Protease Inhibitors by a Quantitative High-Throughput Screening. ACS Pharmacol. Transl. Sci. 2020, 3, 1008–1016. [Google Scholar] [CrossRef]
- Froggatt, H.M.; Heaton, B.E.; Heaton, N.S. Development of a Fluorescence-Based, High-Throughput SARS-CoV-2 3CL(pro) Reporter Assay. J. Virol. 2020, 94, e01265-20. [Google Scholar] [CrossRef] [PubMed]
- Gorgulla, C.; Das KM, P.; Leigh, K.E.; Cespugli, M.; Fischer, P.D.; Wang, Z.F.; Arthanari, H. A multi-pronged approach targeting SARS-CoV-2 proteins using ultra-large virtual screening. iScience 2021, 24, 102021. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Mitchison, T.J.; Bender, A.; Young, D.W.; Tallarico, J.A. Multi-parameter phenotypic profiling: Using cellular effects to characterize small-molecule compounds. Nat. Rev. Drug Discov. 2009, 8, 567–578. [Google Scholar] [CrossRef]
- Nierode, G.; Kwon, P.S.; Dordick, J.S.; Kwon, S.-J. Cell-Based Assay Design for High-Content Screening of Drug Candidates. J. Microbiol. Biotechnol. 2016, 26, 213–225. [Google Scholar] [CrossRef] [Green Version]
- Zaman, G.J. Cell-based screening. Comb. Chem. High Throughput Screen 2008, 11, 494. [Google Scholar] [CrossRef] [PubMed]
- Rumlová, M.; Ruml, T. In vitro methods for testing antiviral drugs. Biotechnol. Adv. 2017, 36, 557–576. [Google Scholar] [CrossRef]
- Green, N.; Ott, R.D.; Isaacs, R.J.; Fang, H. Cell-based assays to identify inhibitors of viral disease. Expert Opin. Drug Discov. 2008, 3, 671–676. [Google Scholar] [CrossRef] [Green Version]
- McLaren, C.; Ellis, M.; Hunter, G. A colorimetric assay for the measurement of the sensitivity of herpes simplex viruses to antiviral agents. Antivir. Res. 1983, 3, 223–234. [Google Scholar] [CrossRef]
- Severson, W.E.; Shindo, N.; Sosa, M.; Fletcher, I.T.; White, E.L.; Ananthan, S.; Jonsson, C.B. Development and Validation of a High-Throughput Screen for Inhibitors of SARS-CoV and Its Application in Screening of a 100,000-Compound Library. SLAS Discov. Adv. Sci. Drug Discov. 2007, 12, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Westby, M.; Nakayama, G.; Butler, S.; Blair, W. Cell-based and biochemical screening approaches for the discovery of novel HIV-1 inhibitors. Antivir. Res. 2005, 67, 121–140. [Google Scholar] [CrossRef]
- Lindsten, K.; Uhlíková, T.; Konvalinka, J.; Masucci, M.; Dantuma, N.P. Cell-Based Fluorescence Assay for Human Immunodeficiency Virus Type 1 Protease Activity. Antimicrob. Agents Chemother. 2001, 45, 2616–2622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.-S.E.; Lo, K.-H.; Hsu, H.-H.; Shao, Y.-M.; Yang, W.-B.; Lin, C.-H.; Wong, C.-H. Screening for HIV protease inhibitors by protection against activity-mediated cytotoxicity in Escherichia coli. J. Virol. Methods 2006, 137, 82–87. [Google Scholar] [CrossRef]
- Bertolin, A.P.; Weissmann, F.; Zeng, J.; Posse, V.; Milligan, J.C.; Canal, B.; Diffley, J.F. Identifying SARS-CoV-2 antiviral compounds by screening for small molecule inhibitors of nsp12/7/8 RNA-dependent RNA polymerase. Biochem. J. 2021, 478, 2425–2443. [Google Scholar] [CrossRef] [PubMed]
- Jan, J.T.; Cheng TJ, R.; Juang, Y.P.; Ma, H.H.; Wu, Y.T.; Yang, W.B.; Wong, C.H. Identification of existing pharmaceuticals and herbal medicines as inhibitors of SARS-CoV-2 infection. Proc. Natl. Acad. Sci. USA 2021, 118, e2021579118. [Google Scholar] [CrossRef]
- Van Damme, E.; De Meyer, S.; Bojkova, D.; Ciesek, S.; Cinatl, J.; De Jonghe, S.; Van Loock, M. In vitro activity of itraconazole against SARS-CoV-2. J. Med. Virol. 2021, 93, 4454–4460. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, S.; Yi, D.; Li, Q.; Ma, L.; Zhang, Y.; Cen, S. A cell-based assay to discover inhibitors of SARS-CoV-2 RNA dependent RNA polymerase. Antiviral Res. 2021, 190, 105078. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Li, Y.S.; Zeng, R.; Liu, F.L.; Luo, R.H.; Huang, C.; Yang, S. SARS-CoV-2 M(pro) inhibitors with antiviral activity in a transgenic mouse model. Science 2021, 371, 1374–1378. [Google Scholar] [CrossRef] [PubMed]
- Fabricant, D.S.; Farnsworth, N.R. The value of plants used in traditional medicine for drug discovery. Environ. Health Perspect. 2001, 109 (Suppl. S1), 69–75. [Google Scholar]
- Mandal, A.; Jha, A.K.; Hazra, B. Plant Products as Inhibitors of Coronavirus 3CL Protease. Front. Pharmacol. 2021, 12, 583387. [Google Scholar] [CrossRef]
- Solecki, R.S. Shanidar IV, a Neanderthal Flower Burial in Northern Iraq. Science 1975, 190, 880–881. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Natural product drug discovery in the next millennium. Pharm. Biol. 2001, 39 (Suppl. S1), 8–17. [Google Scholar] [PubMed] [Green Version]
- Denaro, M.; Smeriglio, A.; Barreca, D.; De Francesco, C.; Occhiuto, C.; Milano, G.; Trombetta, D. Antiviral activity of plants and their isolated bioactive compounds: An update. Phytother. Res. 2019, 34, 742–768. [Google Scholar] [CrossRef]
- Khursheed, A.; Jain, V.; Rasool, A.; Rather, M.A.; Malik, N.A.; Shalla, A.H. Molecular scaffolds from mother nature as possible lead compounds in drug design and discovery against coronaviruses: A landscape analysis of published literature and molecular docking studies. Microb. Pathog. 2021, 157, 104933. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-T.; Hsu, W.-C.; Lin, C.-C. Antiviral Natural Products and Herbal Medicines. J. Tradit. Complement. Med. 2014, 4, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuraskin, C.A.; Marlina; Idroes, R.; Soraya, C. Djufri Activities inhibition methanol extract Laban Leaf (Vitex pinnata) on growth of bacteria S. mutans Atcc 31987. IOP Conf. Series Mater. Sci. Eng. 2019, 523, 012008. [Google Scholar] [CrossRef] [Green Version]
- Pourkhosravani, E.; Nayeri, F.D.; Bazargani, M.M. Decoding antibacterial and antibiofilm properties of cinnamon and cardamom essential oils: A combined molecular docking and experimental study. AMB Express 2021, 11, 143. [Google Scholar] [CrossRef]
- Rahmad, R.; Earlia, N.; Nabila, C.; Inayati, I.; Amin, M.; Prakoeswa, C.R.S.; Khairan, K.; Idroes, R. Antibacterial cream formulation of ethanolic Pliek U extracts and ethanolic residue hexane Pliek U extracts against Staphylococcus aureus. IOP Conf. Ser. Mater. Sci. Eng. 2019, 523, 012011. [Google Scholar] [CrossRef]
- Sardi, J.C.O.; Scorzoni, L.; Bernardi, T.; Fusco-Almeida, A.M.; Mendes Giannini, M.J.S. Candida species: Current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options. J. Med. Microbiol. 2013, 62, 10–24. [Google Scholar] [CrossRef]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; Wasef, L.G.; Elewa, Y.H.; Al-Sagan, A.A.; El-Hack, A.; Taha, M.E.; Abd-Elhakim, Y.M.; Prasad Devkota, H. Chemical Constituents and Pharmacological Activities of Garlic (Allium sativum L.): A Review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef] [Green Version]
- Liana, D.; Phanumartwiwath, A. Leveraging knowledge of Asian herbal medicine and its active compounds as COVID-19 treatment and prevention. J. Nat. Med. 2021, 76, 20–37. [Google Scholar] [CrossRef]
- Grigore, A.; Cord, D.; Tanase, C.; Albulescu, R. Herbal medicine, a reliable support in COVID therapy. J. Immunoass. Immunochem. 2020, 41, 976–999. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.Q.; Xu, J.; Chen, S.D. In Silico Screening of Potential Chinese Herbal Medicine Against COVID-19 by Targeting SARS-CoV-2 3CLpro and Angiotensin Converting Enzyme II Using Molecular Docking. Chin. J. Integr. Med. 2020, 26, 527–532. [Google Scholar] [CrossRef]
- Shree, P.; Mishra, P.; Selvaraj, C.; Singh, S.K.; Chaube, R.; Garg, N.; Tripathi, Y.B. Targeting COVID-19 (SARS-CoV-2) main protease through active phytochemicals of ayurvedic medicinal plants—Withania somnifera (Ashwagandha), Tinospora cordifolia (Giloy) and Ocimum sanctum (Tulsi)—A molecular docking study. J. Biomol. Struct. Dyn. 2022, 40, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Shaik, F.B.; Swarnalatha, K.; Mohan, M.; Thomas, A.; Chikati, R.; Sandeep, G.; Maddu, N. Novel antiviral effects of chloroquine, hydroxychloroquine, and green tea catechins against SARS-CoV-2 main protease (Mpro) and 3C-like protease for COVID-19 treatment. Clin. Nutr. Open Sci. 2022, 42, 62–72. [Google Scholar] [CrossRef]
- Sudeep, H.V.; Gouthamchandra, K.; Shyamprasad, K. Molecular docking analysis of Withaferin A from Withania somnifera with the Glucose regulated protein 78 (GRP78) receptor and the SARS-CoV-2 main protease. Bioinformation 2020, 16, 411–417. [Google Scholar] [CrossRef]
- Chojnacka, K.; Witek-Krowiak, A.; Skrzypczak, D.; Mikula, K.; Młynarz, P. Phytochemicals containing biologically active polyphenols as an effective agent against COVID-19-inducing coronavirus. J. Funct. Foods 2020, 73, 104146. [Google Scholar] [CrossRef] [PubMed]
- Nguyen TT, H.; Jung, J.H.; Kim, M.K.; Lim, S.; Choi, J.M.; Chung, B.; Kim, D. The Inhibitory Effects of Plant Derivate Polyphenols on the Main Protease of SARS Coronavirus 2 and Their Structure-Activity Relationshi. Molecules 2021, 26, 1924. [Google Scholar] [CrossRef]
- Khwaza, V.; Oyedeji, O.O.; Aderibigbe, B.A. Antiviral Activities of Oleanolic Acid and Its Analogues. Molecules 2018, 23, 2300. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, R.; Chakraborty, A.; Biswas, A.; Chowdhuri, S. Depicting the inhibitory potential of polyphenols from Isatis indigotica root against the main protease of SARS-CoV-2 using computational approaches. J. Biomol. Struct. Dyn. 2020, 40, 4110–4121. [Google Scholar] [CrossRef]
- Hu, X.; Cai, X.; Song, X.; Li, C.; Zhao, J.; Luo, W.; Zhang, Q.; Ekumi, I.O.; He, Z. Possible SARS-coronavirus 2 inhibitor revealed by simulated molecular docking to viral main protease and host toll-like receptor. Futur. Virol. 2020, 15, 359–368. [Google Scholar] [CrossRef]
- Zhu, Y.; Scholle, F.; Kisthardt, S.C.; Xie, D.Y. Flavonols and dihydroflavonols inhibit the main protease activity of SARS-CoV-2 and the replication of human coronavirus 229E. Virology 2022, 571, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Naringenin as a possible candidate against SARS-CoV-2 infection and in the pathogenesis of COVID-19. Nat. Prod. Commun. 2021, 16, 1934578X211066723. [Google Scholar] [CrossRef]
- Mishra, A.; Rath, S.C.; Baitharu, I.; Bag, B.P. Millet Derived Flavonoids as Potential SARS-CoV-2 Main Protease Inhibitors: A Computational Approach; Cambridge Open Engage: Cambridge, UK, 2020. [Google Scholar]
- Su, H.X.; Yao, S.; Zhao, W.F.; Li, M.J.; Liu, J.; Shang, W.J.; Xu, Y.C. Anti-SARS-CoV-2 activities in vitro of Shuanghuanglian preparations and bioactive ingredients. Acta Pharmacol. Sin. 2020, 41, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Adem, Ş.; Eyupoglu, V.; Sarfraz, I.; Rasul, A.; Zahoor, A.F.; Ali, M.; Elfiky, A.A. Caffeic acid derivatives (CAFDs) as inhibitors of SARS-CoV-2: CAFDs-based functional foods as a potential alternative approach to combat COVID-19. Phytomedicine 2021, 85, 153310. [Google Scholar] [CrossRef] [PubMed]
- Bahun, M.; Jukić, M.; Oblak, D.; Kranjc, L.; Bajc, G.; Butala, M.; Ulrih, N.P. Inhibition of the SARS-CoV-2 3CLpro main protease by plant polyphenols. Food Chem. 2022, 373, 131594. [Google Scholar] [CrossRef] [PubMed]
- Shahab, S.; Kaviani, S.; Sheikhi, M.; Almodarresiyeh, H.A.; Al Saud, S. Dft calculations and in silico study of chlorogenic, ellagic and quisqualic acids as potential inhibitors of SARS-CoV-2 main protease mpro. Biointerface Res. Appl. Chem. 2022, 12, 61–73. [Google Scholar]
- Ngwe Tun, M.M.; Luvai, E.; Nwe, K.M.; Toume, K.; Mizukami, S.; Hirayama, K.; Morita, K. Anti-SARS-CoV-2 activity of various PET-bottled Japanese green teas and tea compounds in vitro. Arch. Virol. 2022, 167, 1547–1557. [Google Scholar] [CrossRef]
- Khan, A.; Heng, W.; Wang, Y.; Qiu, J.; Wei, X.; Peng, S.; Wei, D.Q. In silico and in vitro evaluation of kaempferol as a potential inhibitor of the SARS-CoV-2 main protease (3CLpro). Phytother. Res. 2021, 35, 2841–2845. [Google Scholar] [CrossRef]
- Owis, A.I.; El-Hawary, M.S.; El Amir, D.; Aly, O.M.; Abdelmohsen, U.R.; Kamel, M.S. Molecular docking reveals the potential of Salvadora persica flavonoids to inhibit COVID-19 virus main protease. RSC Adv. 2020, 10, 19570–19575. [Google Scholar] [CrossRef]
- Shahhamzehei, N.; Abdelfatah, S.; Efferth, T. In Silico and In Vitro Identification of Pan-Coronaviral Main Protease Inhibitors from a Large Natural Product Library. Pharmaceuticals 2022, 15, 308. [Google Scholar] [CrossRef]
- Choudhry, N.; Zhao, X.; Xu, D.; Zanin, M.; Chen, W.; Yang, Z.; Chen, J. Chinese therapeutic strategy for fighting COVID-19 and potential small-molecule inhibitors against severe acute respiratory syndrome Coronavirus 2 (SARS-CoV-2). J. Med. Chem. 2020, 63, 13205–13227. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzyme Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Q.; Chen, Z.; Tao, Y.; Zhang, B.; Wu, X.; Yang, L.; Wang, Z. An integrated method for optimized identification of effective natural inhibitors against SARS-CoV-2 3CLpro. Sci. Rep. 2021, 11, 22796. [Google Scholar] [CrossRef] [PubMed]
- Hariono, M.; Hariyono, P.; Dwiastuti, R.; Setyani, W.; Yusuf, M.; Salin, N.; Wahab, H. Potential SARS-CoV-2 3CLpro Inhibitors from Chromene, Flavonoid and Hydroxamic Acid Compound based on FRET Assay, Docking and Pharmacophore Studies. Results Chem. 2021, 3, 100195. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Chakraborty, A.; Biswas, A.; Chowdhuri, S. Evaluation of green tea polyphenols as novel corona virus (SARS-CoV-2) main protease (Mpro) inhibitors—An in silico docking and molecular dynamics simulation study. J. Biomol. Struct. Dyn. 2021, 39, 4362–4374. [Google Scholar] [CrossRef]
- Rehman MF, U.; Akhter, S.; Batool, A.I.; Selamoglu, Z.; Sevindik, M.; Eman, R.; Aslam, M. Effectiveness of Natural Antioxidants against SARS-CoV-2? Insights from the In-Silico World. Antibiotics 2021, 10, 1011. [Google Scholar] [CrossRef]
- Alrasheid, A.A.; Babiker, M.Y.; Awad, T.A. Evaluation of certain medicinal plants compounds as new potential inhibitors of novel corona virus (COVID-19) using molecular docking analysis. Silico Pharmacol. 2021, 9, 10. [Google Scholar] [CrossRef]
- Emon, N.U.; Alam, M.M.; Akter, I.; Akhter, S.; Sneha, A.A.; Irtiza, M.; Hossain, S. Virtual screenings of the bioactive constituents of tea, prickly chaff, catechu, lemon, black pepper, and synthetic compounds with the main protease (Mpro) and human angiotensin-converting enzyme 2 (ACE 2) of SARS-CoV-2. Futur. J. Pharm. Sci. 2021, 7, 121. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Singh, P.; Sharma, S.; Singh, T.P.; Ethayathulla, A.S.; Kaur, P. Identification of bioactive molecule from Withania somnifera (Ashwagandha) as SARS-CoV-2 main protease inhibitor. J. Biomol. Struct. Dyn. 2021, 39, 5668–5681. [Google Scholar] [CrossRef]
- Zígolo, M.A.; Goytia, M.R.; Poma, H.R.; Rajal, V.B.; Irazusta, V.P. Virtual screening of plant-derived compounds against SARS-CoV-2 viral proteins using computational tools. Sci. Total Environ. 2021, 781, 146400. [Google Scholar] [CrossRef]
- van de Sand, L.; Bormann, M.; Alt, M.; Schipper, L.; Heilingloh, C.S.; Steinmann, E.; Krawczyk, A. Glycyrrhizin Effectively Inhibits SARS-CoV-2 Replication by Inhibiting the Viral Main Protease. Viruses 2021, 13, 609. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulou, I.; Sapountzaki, E.; Rova, U.; Christakopoulos, P. Inhibition of the main protease of SARS-CoV-2 (Mpro) by repurposing/designing drug-like substances and utilizing nature’s toolbox of bioactive compounds. Comput. Struct. Biotechnol. J. 2022, 20, 1306–1344. [Google Scholar] [CrossRef] [PubMed]
- Veerasamy, R.; Karunakaran, R. Molecular docking unveils the potential of andrographolide derivatives against COVID-19: An in silico approach. J. Genet. Eng. Biotechnol. 2022, 20, 58. [Google Scholar] [CrossRef] [PubMed]
- Shady, N.H.; Youssif, K.A.; Sayed, A.M.; Belbahri, L.; Oszako, T.; Hassan, H.M.; Abdelmohsen, U.R. Sterols and Triterpenes: Antiviral Potential Supported by In-Silico Analysis. Plants 2020, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Falade, V.A.; Adelusi, T.I.; Adedotun, I.O.; Abdul-Hammed, M.; Lawal, T.A.; Agboluaje, S.A. In silico investigation of saponins and tannins as potential inhibitors of SARS-CoV-2 main protease (Mpro). Silico Pharmacol. 2021, 9, 1–15. [Google Scholar] [CrossRef]
- Wang, S.C.; Chen, Y.; Wang, Y.C.; Wang, W.J.; Yang, C.S.; Tsai, C.L.; Hung, M.C. Tannic acid suppresses SARS-CoV-2 as a dual inhibitor of the viral main protease and the cellular TMPRSS2 protease. Am. J. Cancer Res. 2020, 10, 4538–4546. [Google Scholar]
- Sisakht, M.; Mahmoodzadeh, A.; Darabian, M. Plant-derived chemicals as potential inhibitors of SARS-CoV-2 main protease (6LU7), a virtual screening study. Phytother. Res. 2021, 35, 3262–3274. [Google Scholar] [CrossRef]
- Kaul, R.; Paul, P.; Kumar, S.; Büsselberg, D.; Dwivedi, V.D.; Chaari, A. Promising antiviral activities of natural flavonoids against SARS-CoV-2 targets: Systematic review. Int. J. Mol. Sci. 2021, 22, 11069. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [Green Version]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kang, O.-J.; Gweon, O.-C. Comparison of phenolic acids and flavonoids in black garlic at different thermal processing steps. J. Funct. Foods 2013, 5, 80–86. [Google Scholar] [CrossRef]
- Li, F.; Song, X.; Su, G.; Wang, Y.; Wang, Z.; Jia, J.; Qing, S.; Huang, L.; Wang, Y.; Zheng, K.; et al. Amentoflavone Inhibits HSV-1 and ACV-Resistant Strain Infection by Suppressing Viral Early Infection. Viruses 2019, 11, 466. [Google Scholar] [CrossRef] [Green Version]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaihatsu, K.; Yamabe, M.; Ebara, Y. Antiviral Mechanism of Action of Epigallocatechin-3-O-gallate and Its Fatty Acid Esters. Molecules 2018, 23, 2475. [Google Scholar] [CrossRef] [Green Version]
- Lingwan, M.; Shagun, S.; Pahwa, F.; Kumar, A.; Verma, D.K.; Pant, Y.; Masakapalli, S.K. Phytochemical rich Himalayan Rhododendron arboreum petals inhibit SARS-CoV-2 infection in vitro. J. Biomol. Struct. Dyn. 2021, 28, 1–11. [Google Scholar] [CrossRef]
- Zhu, Y.; Xie, D.-Y. Docking Characterization and in vitro Inhibitory Activity of Flavan-3-ols and Dimeric Proanthocyanidins against the Main Protease Activity of SARS-CoV-2. Front. Plant Sci. 2020, 11, 601316. [Google Scholar] [CrossRef] [PubMed]
- Park, R.; Jang, M.; Park, Y.-I.; Park, Y.; Jung, W.; Park, J.; Park, J. Epigallocatechin Gallate (EGCG), a Green Tea Polyphenol, Reduces Coronavirus Replication in a Mouse Model. Viruses 2021, 13, 2533. [Google Scholar] [CrossRef]
- Zhao, Z.; Feng, M.; Wan, J.; Zheng, X.; Teng, C.; Xie, X.; Pan, W.; Hu, B.; Huang, J.-A.; Liu, Z.-H.; et al. Research progress of epigallocatechin-3-gallate (EGCG) on anti-pathogenic microbes and immune regulation activities. Food Funct. 2021, 12, 9607–9619. [Google Scholar] [CrossRef]
- Maiti, S.; Banerjee, A.; Kanwar, M. Effects of theaflavin-gallate in-silico binding with different proteins of SARS-CoV-2 and host inflammation and vasoregulations referring an experimental rat-lung injury. Phytomed. Plus 2022, 2, 100237. [Google Scholar] [CrossRef]
- Chung, K.-T.; Wong, T.Y.; Wei, C.-I.; Huang, Y.-W.; Lin, Y. Tannins and Human Health: A Review. Crit. Rev. Food Sci. Nutr. 1998, 38, 421–464. [Google Scholar] [CrossRef]
- Chen, C.N.; Lin, C.P.; Huang, K.K.; Chen, W.C.; Hsieh, H.P.; Liang, P.H.; Hsu, J.T.A. Inhibition of SARS-CoV 3C-like Protease Activity by Theaflavin-3,3′-digallate (TF3). Evid. Based Complement. Alternat. Med. 2005, 2, 209–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaraj, J.; Rajan, L.; Selvaraj, D.; Palanisamy, D.; Pk, K.N.; Mohankumar, S.K. Identification of (2R,3R)-2-(3,4-dihydroxyphenyl)chroman-3-yl-3,4,5-trihydroxy benzoate as multiple inhibitors of SARS-CoV-2 targets; a systematic molecular modelling approach. RSC Adv. 2021, 11, 13051–13060. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hao, L.; Liu, L.; Chen, G.; Jiang, W.; Xu, W.; Zhu, C.; Yao, G.; Fang, S. Cinnamtannin B-1 Prevents Ovariectomy-Induced Osteoporosis via Attenuating Osteoclastogenesis and ROS Generation. Front. Pharmacol. 2020, 11, 1023. [Google Scholar] [CrossRef]
- Abookleesh, F.L.; Al-Anzi, B.S.; Ullah, A. Potential Antiviral Action of Alkaloids. Molecules 2022, 27, 903. [Google Scholar] [CrossRef]
- Reyes-Escogido, M.D.L.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and Pharmacological Aspects of Capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhaykumar, K.J.W.S.N. Phytochemical studies on Achyranthes aspera. World Sci. News 2018, 100, 16–34. [Google Scholar]
- Bergman, M.E.; Davis, B.; Phillips, M.A. Medically Useful Plant Terpenoids: Biosynthesis, Occurrence, and Mechanism of Action. Molecules 2019, 24, 3961. [Google Scholar] [CrossRef] [Green Version]
- Venkatesan, N.; Punithavathi, D.; Babu, M. Protection from Acute and Chronic Lung Diseases By Curcumin. Mol. Targets Ther. Uses Curcumin Health Dis. 2007, 595, 379–405. [Google Scholar]
- Lelli, D.; Sahebkar, A.; Johnston, T.P.; Pedone, C. Curcumin use in pulmonary diseases: State of the art and future perspectives. Pharmacol. Res. 2016, 115, 133–148. [Google Scholar] [CrossRef]
- Thimmulappa, R.K.; Mudnakudu-Nagaraju, K.K.; Shivamallu, C.; Subramaniam, K.; Radhakrishnan, A.; Bhojraj, S.; Kuppusamy, G. Antiviral and immunomodulatory activity of curcumin: A case for prophylactic therapy for COVID-19. Heliyon 2021, 7, e06350. [Google Scholar] [CrossRef]
- Salehi, B.; Upadhyay, S.; Erdogan Orhan, I.; Kumar Jugran, A.; LDJayaweera, S.; ADias, D.; Sharifi-Rad, J. Therapeutic Potential of α- and β-Pinene: A Miracle Gift of Nature. Biomolecules 2019, 9, 738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alviano, D.S.A.A.C.S.; Alviano, C.S. Plant Extracts: Search for New Alternatives to Treat Microbial Diseases. Curr. Pharm. Biotechnol. 2009, 10, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Koudou, J.; Abena, A.; Ngaissona, P.; Bessière, J. Chemical composition and pharmacological activity of essential oil of Canarium schweinfurthii. Fitoterapia 2005, 76, 700–703. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Saab, A.; Tundis, R.; Statti, G.A.; Lampronti, I.; Menichini, F.; Gambari, R.; Cinatl, J.; Doerr, H.W. Phytochemical analysis and in vitro evaluation of the biological activity against herpes simplex virus type 1 (HSV-1) of Cedrus libani A. Rich. Phytomedicine 2008, 15, 79–83. [Google Scholar] [CrossRef]
- da Silva Rivas, A.C.; Lopes, P.M.; de Azevedo Barros, M.M.; Costa Machado, D.C.; Alviano, C.S.; Alviano, D.S. Biological Activities of a-Pinene and β-Pinene Enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar]
- Enmozhi, S.K.; Raja, K.; Sebastine, I.; Joseph, J. Andrographolide as a potential inhibitor of SARS-CoV-2 main protease: An in silico approach. J. Biomol. Struct. Dyn. 2021, 39, 3092–3098. [Google Scholar] [CrossRef]
- Alsaffar, D.F.; Yaseen, A. In Silico Molecular Docking Studies of Medicinal Arabic Plant-Based Bioactive Compounds as a Promising Drug Candidate against COVID-19. Int. J. Innov. Sci. Res. Technol. 2020, 5, 876–896. [Google Scholar]
- Chaudhuri, S.K.; Fullas, F.; Brown, D.M.; Wani, M.C.; Wall, M.E.; Cai, L.; Mar, W.; Lee, S.K.; Luo, Y.; Zaw, K.; et al. Isolation and Structural Elucidation of Pentacyclic Triterpenoids from Maprounea africana. J. Nat. Prod. 1995, 58, 1–9. [Google Scholar] [CrossRef]
- Kai, G.; Xu, H.; Zhou, C.; Liao, P.; Xiao, J.; Luo, X.; You, L.; Zhang, L. Metabolic engineering tanshinone biosynthetic pathway in Salvia miltiorrhiza hairy root cultures. Metab. Eng. 2011, 13, 319–327. [Google Scholar] [CrossRef]
- Deng, C.; Hao, X.; Shi, M.; Fu, R.; Wang, Y.; Zhang, Y.; Zhou, W.; Feng, Y.; Makunga, N.P.; Kai, G. Tanshinone production could be increased by the expression of SmWRKY2 in Salvia miltiorrhiza hairy roots. Plant Sci. 2019, 284, 1–8. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, S.; Shen, Y.; Liu, Y.; Maoz, I.; Gao, X.; Chen, C.; Liu, T.; Wang, C.; Kai, G. Overexpression of SmSCR1 Promotes Tanshinone Accumulation and Hairy Root Growth in Salvia miltiorrhiza. Front. Plant Sci. 2022, 13, 475. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Ding, M.; Yin, X.; Wang, G.; Zhang, B.; Chen, L.; Dong, J. MAPKK2/4/5/7-MAPK3-JAZs modulate phenolic acid biosynthesis in Salvia miltiorrhiza. Phytochemistry 2022, 199, 113177. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, S.; Yu, O. Metabolic engineering of flavonoids in plants and microorganisms. Appl. Microbiol. Biotechnol. 2011, 91, 949–956. [Google Scholar] [CrossRef]
- Rai, A.; Smita, S.S.; Singh, A.K.; Shanker, K.; Nagegowda, D.A. Heteromeric and Homomeric Geranyl Diphosphate Synthases from Catharanthus roseus and Their Role in Monoterpene Indole Alkaloid Biosynthesis. Mol. Plant 2013, 6, 1531–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duran, N.; Polat, M.F.; Aktas, D.A.; Alagoz, M.A.; Ay, E.; Cimen, F.; Tek, E.; Anil, B.; Burmaoglu, S.; Algul, O. New chalcone derivatives as effective against SARS-CoV-2 agent. Int. J. Clin. Pract. 2021, 75, e14846. [Google Scholar] [CrossRef] [PubMed]


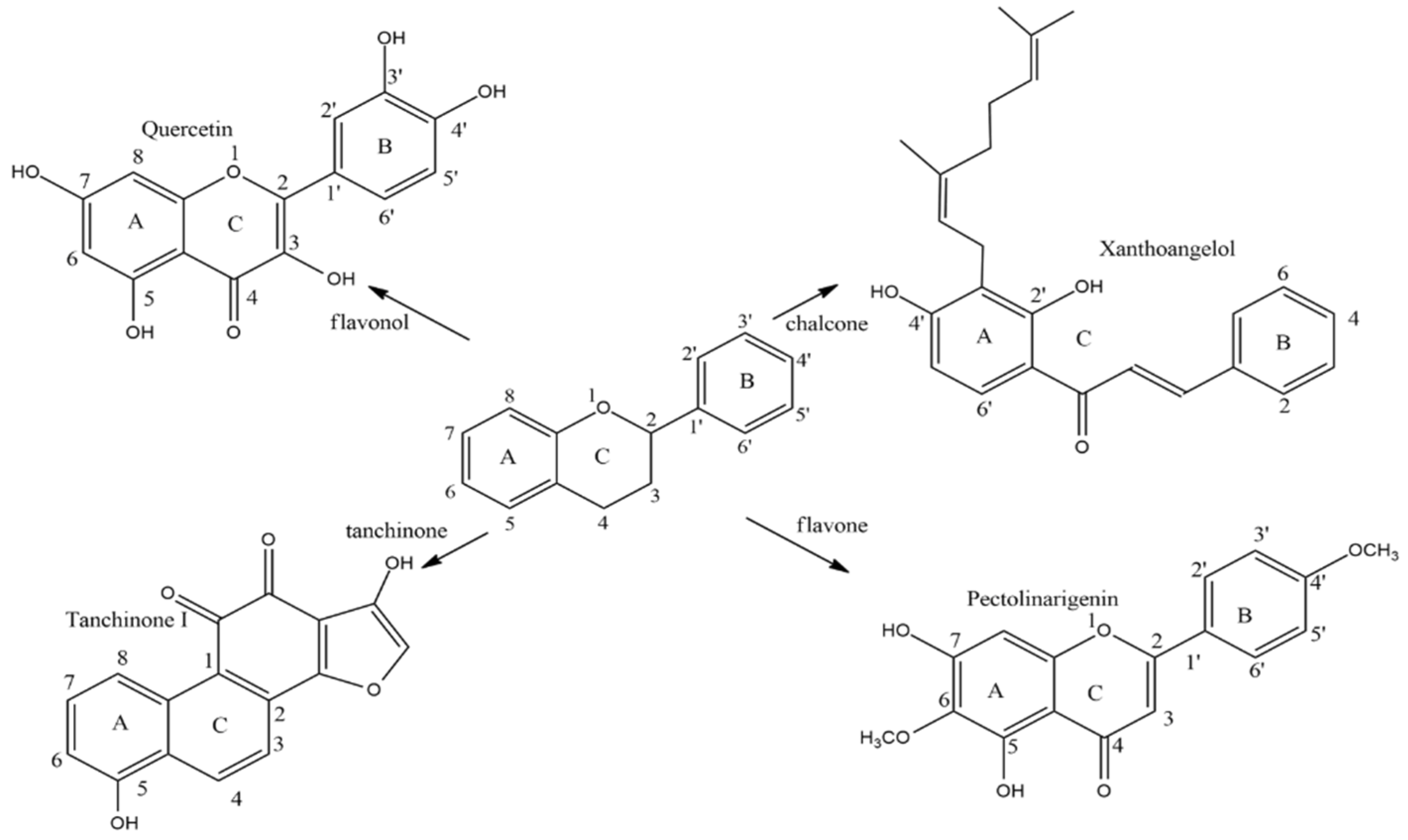{kind=link}
| Class of Compound | Compound | Distributed in | Including Food Plants and Spices * | Inhibition (%), with IC50 Value, μM | Binding Energy ** (kcal/mol) | Ref. |
|---|---|---|---|---|---|---|
| Isoflavone | Daidzein | Streptomyces, and predominantly in Fabaceae plant family | Vigna radiata, Glycine max | 56 | −6.5 | [93] |
| Isoflavone | Puerarin | Predominantly in Fabaceae plant family | Cicer arietinum, Glycine max, Glycyrrhiza glabra, Phaseolus vulgaris, Pisum sativum, Vigna radiata | 42 ± 2 | −6.63 | [94] |
| Flavonol | Myricetin | Widely distributed in various plant families | Buchanania lanzan, Mangifera indica, Asparagus officinalis, Davidsonia pruriens, Hippophae rhamnoides, Vicia faba, Salvia hispanica, Thymus capitatus, Punica granatum, Hibiscus sabdariffa L., Moringa oleifera, Eugenia jambolana, Pimenta dioica, Plinia pinnata, Syzygium aromaticum, Syzygium cumini, Syzygium samarangense, Diploknema butyracea, Ampelopsis grossedentata, Morella rubra | 43 ± 1 | −22.13 | [41,91] |
| Flavonol | Quercetin | Mucor hiemalis, and widely distributed in various plant families | Allium cepa, Allium Sativum, Allium ascalonicum, Mangifera indica, Annona muricata, Asparagus officinalis, Capparis spinosa, Carica papaya, Garcinia cowa, Garcinia dulcis, Brassica oleracea var. gongylodes, Raphanus sativus, Momordica charantia, Ceratonia siliqua, Vicia faba, Crocus sativus, Punica granatum, Toona sinensis, Moringa stenopetala, Musa acuminata, Psidium guajava, Phyllanthus emblica, Zea mays, Nigella sativa, Eriobotrya japonica, Prunus avium, Kadsura heteroclita, Capsicum annuum, Zingiber officinale | 93 ± 5 | −7.6 | [23,91] |
| Flavonol | Quercetagetin (Quercetagenin) | Asteraceae, Eriocaulaceae, Fabaceae plant families | Acacia catechu, Leucaena glauca | 145 ± 6 | −15.2 | [91] |
| Flavanonol | Ampelopsin (dihydromyricetin) | Widely distributed in various plant families | Asparagus officinalis, Punica granatum L., Pimenta dioica, Zea mays, Syzygium cumini, Capsicum annuum, Vitis rotundifolia, Manilkara zapota, | 128 ± 5 | −7.5 | [91,95] |
| Flavanonol | Ampelopsin-4′-O-α-d-glucopyranoside | Widely distributed in various plant families | - | 195 ± 5 | 7.4 | [91] |
| Flavanone | Naringenin | Widely distributed in various plant families | Camellia sinensis, Prunus cerasus, Prunus persica, Citrus grandi, etc. | 150 ± 10 | −7.7 | [91,96] |
| Flavan-3-ol | Epigallocatechin gallate (EGCG) | Widely distributed in various plant families | Vitis vinifera | 171 ± 5 | −7.6 −8.2 | [88,91,93] |
| Flavone | Vitexin | Widely distributed in various plant families | Pisum sativum | 180 ± 6 | −7.6 | [97] |
| Hydrocinnamic acid | Chlorogenic acid | Widely distributed in various plant families | Lactuca sativa | 39.48 ± 5.51 | −12.98 | [91,98] |
| Dihydroxycinnamic acid | Caffeic acid | Widely distributed in various plant families | Actinidia deliciosa, Allium Sativum, Mangifera indica, Ilex paraguariensis, Carica papaya, Beta vulgaris, Terminalia catappa, Terminalia chebula, Brassica oleracea var. gongylodes, Raphanus sativus, Momordica charantia, Arachis hypogaea, Cicer arietinum, Glycine max, Phaseolus vulgaris, Pisum sativum, Tetrapleura tetraptera, Ocimum basilicum, Rosmarinus officinalis, Thymus capitatus, Punica granatum, Triticum aestivum, Zea mays, Crataegus pinnatifida, Prunus avium, Coffea arabica, Citrus limon, Citrus sinensis, Solanum lycopersicum, Solanum phureja, Solanum pimpinellifollium, Solanum tuberosum, Curcuma longa, Bergera koenigii | 197 ± 1 | −12.4985 | [91,99] |
| Polyphenol | Ellagic acid | Widely distributed in various plant families | Mangifera indica, Terminalia chebula, Punica granatum, Moringa oleifera, Moringa peregrine, Moringa stenopetala, Eugenia jambolana, Myrciaria cauliflora, Syzygium aromaticum, Syzygium cumini, Emblica officinalis, Rubus idaeus L., | 11.8 ± 5.7 | −15.955 | [100,101] |
| Phenylpropanoid | Chicoric acid | Alliaceae, Asteraceae, and Labiatae plant families | Lactuca sativa, Ocimum basilicum, Cichorium intybus | - | −8.2 | [93] |
| Polyphenol | Gallocatechin gallate (GCG) | Cistaceae, Elaeagnaceae, Ericaceae, Polygonaceae, Theaceae, and Vitaceae plant families | Hippophae rhamnoides, Camellia sinensis, Vitis vinifera | 5.774 ± 0.805 | −9 | [93] |
| Flavan-3-ol | Epicatechin gallate (ECG) | Cistaceae, Elaeagnaceae, Ericaceae, Polygonaceae, Theaceae, and Vitaceae plant families | Hippophae rhamnoides, Camellia sinensis, Vitis vinifera, Rheum sp. | 12.5 | −8.2 | [93,102] |
| Flavonoids | Kaempferol glycosides | Widely distributed in various plant families | Prunus avium, Allium cepa | 125.00 | −7.4, −8.1 | [103,104] |
| Flavonoids | Isorhamnetin glycosides | Widely distributed in various plant families | Brassica oleracea, Allium ascalonicum | 13.13 | −6.6, −8.2 | [104,105] |
| Flavonoids | Pectolinarin | Labiatae, Plantaginaceae, and Verbenaceae plant families | 37.7 | −8.2 | [40,106,107] | |
| Flavonoid | Herbacetin | Asteraceae, Chenopodiaceae, Crassulaceae, Ephedraceae, Equisetaceae, Linaceae, Malvaceae, Papaveraceae, Phrymaceae, Primulaceae, Rosaceae, Rutaceae, and Taxaceae plant families | Linum usitatissimum, Citrus limon | 33.1 | −7.2 | [40,106] |
| Flavonoid | Rhoifolin (apigenin-7-O-rhamnoglucoside) | Acanthaceae, Anacardiaceae, Apocynaceae, Fabaceae, Lythraceae, Oleaceae, Rutaceae, and Caprifoliaceae plant families | Vicia faba, Hordeum vulgare L | 27.4 | −8.2 | [40,106] |
| Flavonoid metabolite | Vicenin | Rare compound | Trigonella foenum-graecum | 38.856 | −8.97 | [87,108] |
| Flavone | Isorientin 4′-O-glucoside 2″-O-p-hydroxybenzoate | Gentianaceae and Lamiaceae | Ocimum sanctum | - | −8.55 | [87] |
| Biflavonoid | Amentoflavone | Various plant species: yew, juniper, oak, and willow | Garcinia brasiliensis, Garcinia dulcis, Garcinia hombroniana, Garcinia indica, Garcinia intermedia, Garcinia livingstonei, Garcinia madruno, Garcinia mangostana, Garcinia morella, Garcinia wightii, Garcinia xanthochymus, | - | −10.0 | [23] |
| Flavonoid | Silymarin (silibinin) | Asteraceae | Silybum marianum | 46.88 | −7.6 | [23] |
| Isoflavone | Torvanol A | Solanaceae | - | −7.5 | [23] | |
| Flavone | Scutellarein | Asphodelaceae, Asteraceae, Fabaceae, Bignoniaceae, Labiatae, Plantaginaceae, Polygonaceae, and Verbenaceae plant families | Oroxylum indicum, Garcinia andamanica Origanum majorana | - | −7.4 | [23] |
| Flavone | Apigenin | Widely distributed in various plant families, mainly in Labiatae | Artemisia diffusa., Ocimum americanum var. pilosum Ocimum basilicum, Ocimum x citriodorum, Rosmarinus officinalis, Salvia officinalis, Thymus piperella, Passiflora foetida, Piper peepuloides, Kaempferia parviflora | 925 | −7.1 | [23,109] |
| Isoflavone | 5,7,3′,4′-Tetrahydroxy-2′-(3,3-dimethylallyl) isoflavone | Fabaceae | - | −29.57 | [41] | |
| Polyphenol | Methyl rosmarinate | Labiatae | Rosmarinus officinalis | 21.32 | −20.62 | [41,110] |
| Flavonoid | Baicalin | Mainly in Labiatae | Oroxylum indicum | 6.41 ± 0.95 μM in vitro 27.87 ± 0.04 μM in cells | −8.85 | [99,111] |
| Flavonoid | Baicalein | Mainly in Labiatae | Oroxylum indicum | 0.94 ± 0.20 μM in vitro 2.94 ± 1.19 μM in cells | [98] | |
| Alkaloid | Capsaicin | Solanaceae | Capsicum annuum | - | −13.90 | [112] |
| Alkaloid | Psychotrine | Mainly in Rubiaceae | - | −13.5 | [112] | |
| Alkaloid | Achyranthine | Amaranthaceae | - | 4.1 | [113] | |
| Terpenoid | Withanoside V | Solanaceae | 5.774 ± 0.805 | −10.32 | [114] | |
| Triterpenoid | Ursolic acid | Apocynaceae, Asteraceae, Boraginaceae, Dryopteridaceae, Ericaceae, Gesneriaceae, Labiatae, Lamiaceae, Lythraceae, Moraceae, Myricaceae, Nothofagaceae, Oleaceae, Rosaceae, Rubiaceae, Solanaceae, Stilbaceae, and Ulmaceae plant families | Vaccinium macrocarpon, Punica granatum, Olea europaea, Prunus avium, Pyrus spp. | 12.6 | −8.2 | [114] |
| Triterpenoid | Glycyrrhizic acid | Asteraceae and Fabaceae | Stevia rebaudiana, Glycyrrhiza glabra | - | −8.03 | [115,116] |
| Pentacyclic triterpenoid | Torvoside H | Solanaceae | - | - | −8.4 | [23] |
| Pentacyclic triterpenoid | Lupeol | In Coprinaceae, and widely distributed in various plant families | Cichorium intybus, Zanthoxylum armatum, Olea europaea, Myrica rubra, Morus alba, Ficus carica, Carica papaya | - | −7.6 | [116] |
| Diterpene | Scopadulcic acid B | Plantaginaceae (Scoparia dulcis) | - | - | −8.5 | [23] |
| Diterpene | Ovatodiolide | Lamiaceae | - | - | −6.9 | [23] |
| Terpene | Curcumin | Mainly in Zingiberaceae | Curcuma longa, Curcuma mangga, Zingiber officinale | 11.9 | −6.5 | [23,117] |
| Terpene | Parthenolide | Asteraceae, Magnoliaceae, and Celastraceae plant families | - | - | −6.0 | [23] |
| Meroterpenoid | Illicinone A | Illiaceae | Illiciumverum | - | −5.0 | [23] |
| Meroterpenoid | Piperitenone | Labiatae and Poaceae | Mentha spp. | - | −4.3 | [23] |
| Cyclic monoterpene | Limonene | Widely distributed in various plant families | Citrus aurantium, Citrus aurantifolia, Citrus bergamia, Citrus grandis, Citrus junos, Citrus latifolia, Citrus limettioides, Citrus limon, Citrus medica, Citrus paradisi, Citrus reticulata, Citrus sinensis, Zanthoxylum armatum, Allium sativum, Anacardium occidentale, Mangifera indica, Monodora myristica, Xylopia aethiopica, Cuminum cyminum. Foeniculum vulgare, Petroselinum crispum, Porophyllum ruderale, Beta vulgaris, Ocimum basilicum, Thymus piperella, Acca sellowiana, Psidium guajava, Averrhoa carambola, Piper nigrum Prunus avium, Coffea arabica, Coffea canephora, Citrus aurantifolia, Curcuma amada, Curcuma longa | - | −5.2 | [113] |
| Cyclic monoterpene | Sabinene | Labiatae, Cupressaceae, Myristicaceae, and Pinaceae plant families | - | - | −4.8 | [113] |
| Bicyclic monoterpene | Pinene | Widely distributed in various plant families | Piper nigrum, Allium sativum, Anacardium occidentale, Mangifera indica, Pistacia vera, Monodora myristica, Cuminum cyminum, Foeniculum vulgare, Petroselinum crispum, Ocimum basilicum, Origanum vulgare, Rosmarinus officinalis, Myristica fragrans, Eriobotrya japonica, Fragaria vesca, Citrofortunella mitis, Citrus aurantifolia, Citrus spp., Curcuma mangga, Curcuma amada, Aframomum melegueta, Solanum lycopersicum, Zanthoxylum armatum, | - | −4.6 | [113] |
| Labdane diterpenoid | Andrographolide | Acanthaceae | - | - | −6.6 | [118] |
| Triterpene | 1β-hydroxyaleuritolic acid 3-p-hydroxybenzoate | Euphorbiaceae | - | - | −8.5 | [119] |
| Steroidal lactone | Withaferin A | Solanaceae | - | - | −9.83 | [89] |
| Tannin | Tannic acid | Ephedraceae and Geraniaceae | - | 13.4 | −7.5 | [120,121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Issa, S.S.; Sokornova, S.V.; Zhidkin, R.R.; Matveeva, T.V. The Main Protease of SARS-CoV-2 as a Target for Phytochemicals against Coronavirus. Plants 2022, 11, 1862. https://doi.org/10.3390/plants11141862
Issa SS, Sokornova SV, Zhidkin RR, Matveeva TV. The Main Protease of SARS-CoV-2 as a Target for Phytochemicals against Coronavirus. Plants. 2022; 11(14):1862. https://doi.org/10.3390/plants11141862
Chicago/Turabian StyleIssa, Shaza S., Sofia V. Sokornova, Roman R. Zhidkin, and Tatiana V. Matveeva. 2022. "The Main Protease of SARS-CoV-2 as a Target for Phytochemicals against Coronavirus" Plants 11, no. 14: 1862. https://doi.org/10.3390/plants11141862