
Isolation and Comprehensive in Silico Characterisation of a New 3-Hydroxy-3-Methylglutaryl-Coenzyme A Reductase 4 (HMGR4) Gene Promoter from Salvia miltiorrhiza: Comparative Analyses of Plant HMGR Promoters



Abstract
:1. Introduction
2. Results
2.1. In Silico Analysis of the S. miltiorrhiza HMGR4 Promoter
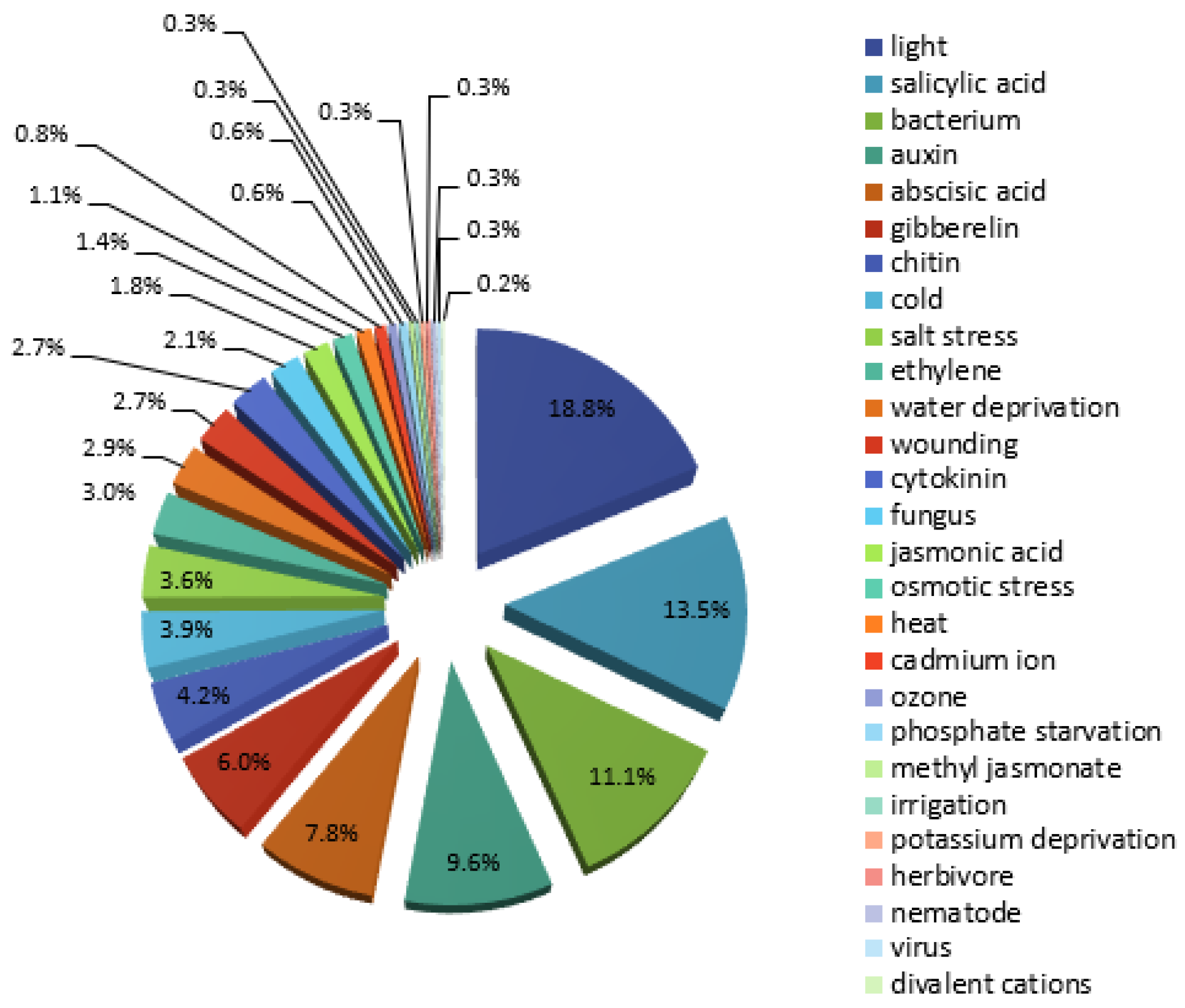
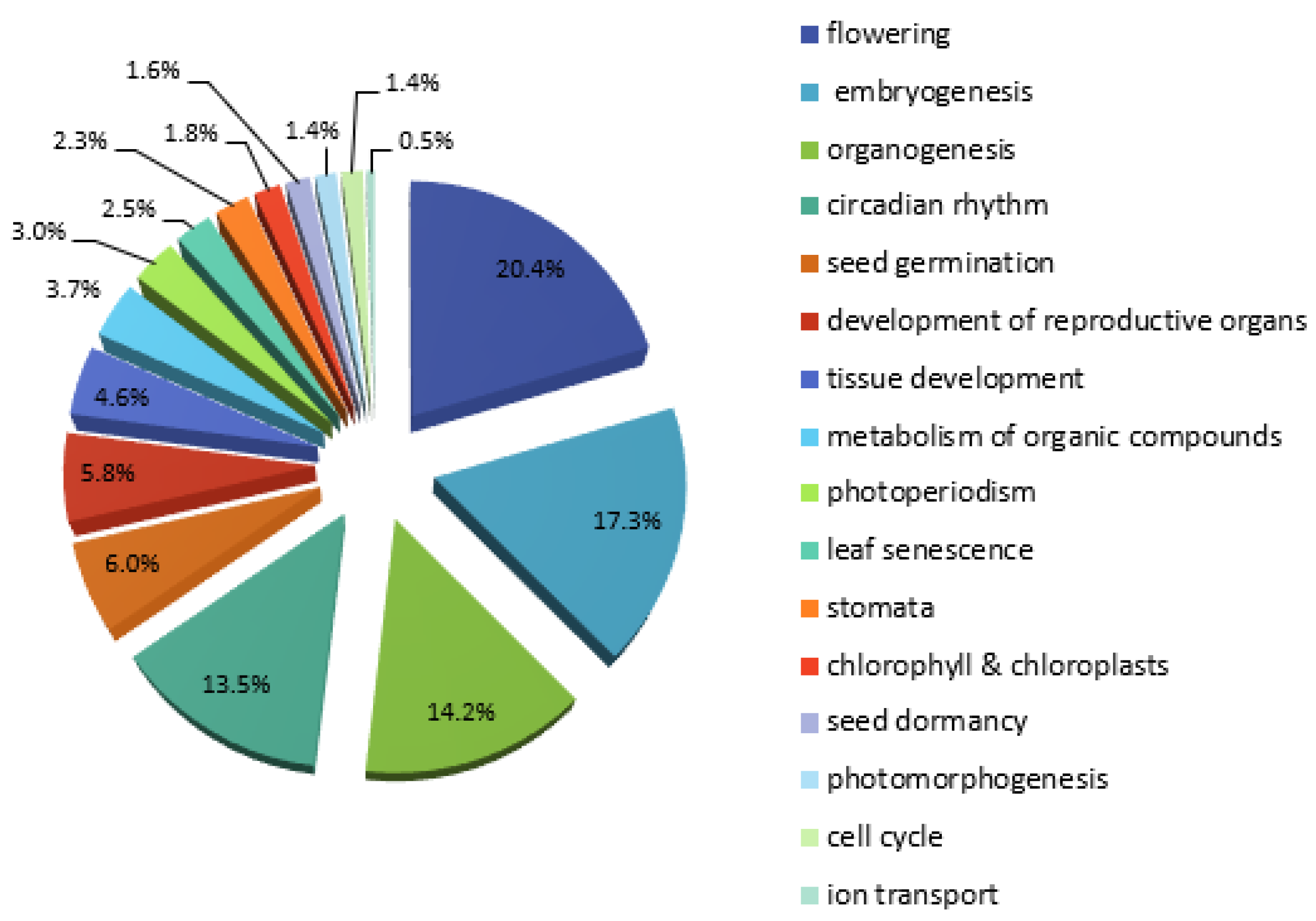
2.2. Microarray and Next-Generation Sequencing (NGS) Co-Expression Data Analysis
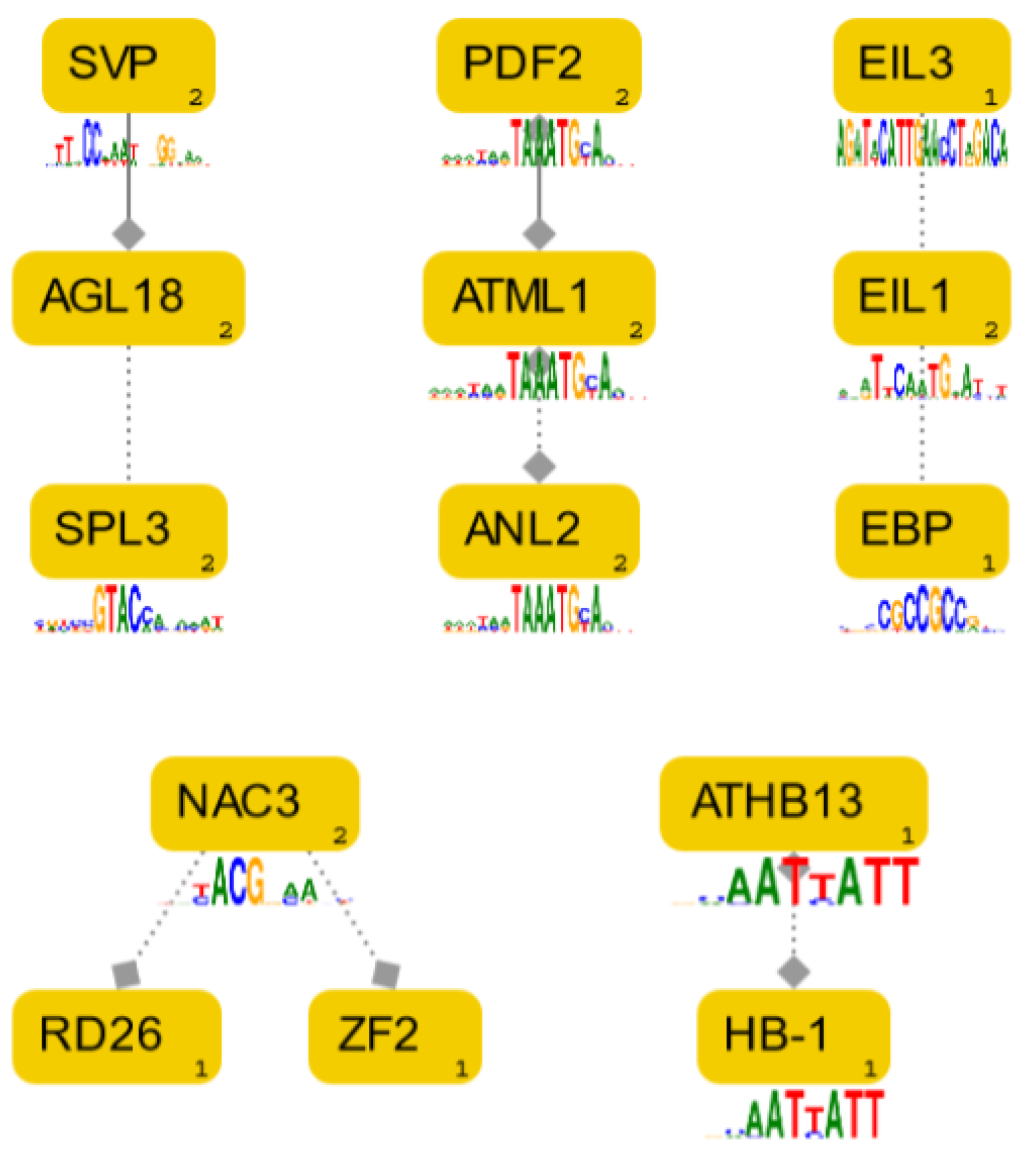
2.3. Comparison of the in Silico HMGR4 Analysis Results with Microarray Co-Expression Data
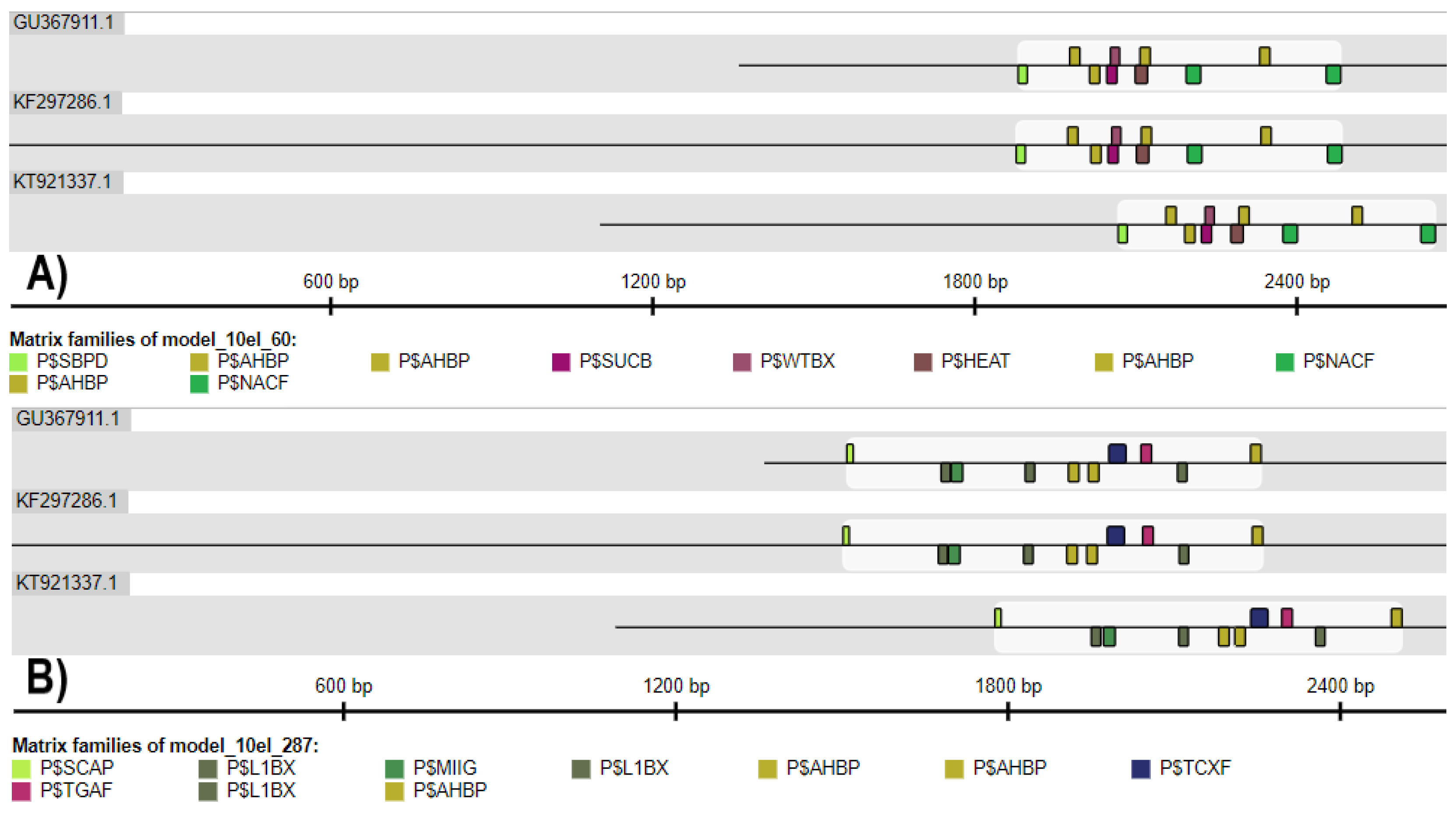
2.4. Comparison of S. miltiorrhiza HMGR Promoters
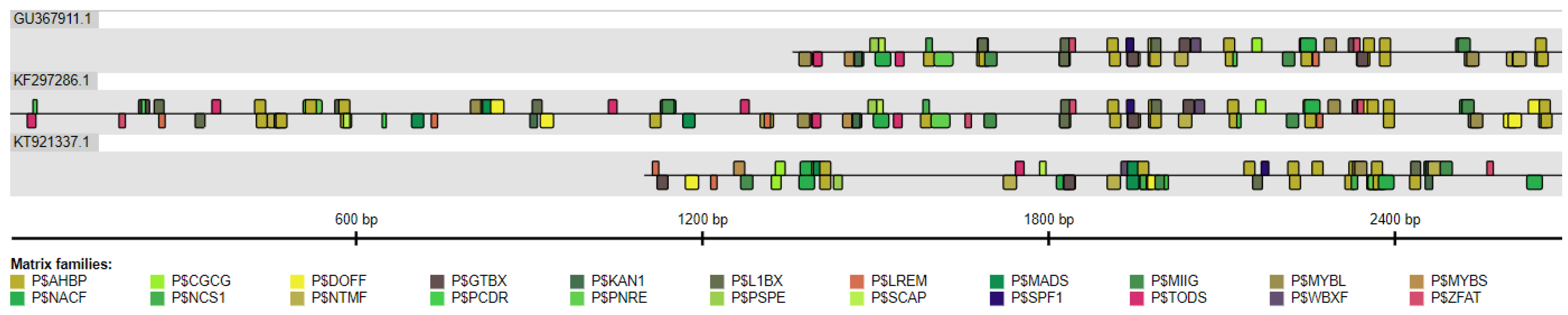
2.5. The Conservation of Plant HMGR Promoters
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Isolation of the S. miltiorrhiza HMGR4 Promoter Sequence
4.3. In Silico Analysis of the S. miltiorrhiza HMGR4 Promoter Sequence
4.4. Microarray and NGS Co-Expression Data Analysis
4.5. Comparison of S. miltiorrhiza HMGR Promoters
4.6. Assessment of Conservation of Plant HMGR Promoters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, Y.; Wan, R.; Lin, Y.; Yang, L.; Chen, Y.; Liu, C. Recent advance on research and application of Salvia miltiorrhiza. Asian J. Pharmacodyn. Pharmacokinet. 2007, 7, 99–130. [Google Scholar]
- Cheng, T.O. Cardiovascular effects of Danshen. Int. J. Cardiol. 2007, 121, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.S.; Wang, S.Q. Salvianolic acid B from Salvia miltiorrhiza inhibits tumor necrosis factor-alpha (TNF-alpha)-induced MMP-2 upregulation in human aortic smooth muscle cells via suppression of NAD(P)H oxidase-derived reactive oxygen species. J. Mol. Cell. Cardiol. 2006, 41, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.N.; An, C.N.; Zhang, H.N.; Pu, X.P. Protocatechuic acid inhibits neurotoxicity induced by MPTP in vivo. Neurosci. Lett. 2010, 474, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.K.; Ho, M.T.; Lin, H.Q.; Lau, K.F.; Rudd, J.A.; Chung, R.C.; Fung, K.P.; Shaw, P.C.; Wan, D.C. Cryptotanshinone, an acetylcholinesterase inhibitor from Salvia miltiorrhiza, ameliorates scopolamine-induced amnesia in Morris water maze task. Planta Med. 2010, 76, 228–234. [Google Scholar] [CrossRef]
- Kang, D.G.; Oh, H.; Sohn, E.J.; Hur, T.Y.; Lee, K.C.; Kim, K.J.; Kim, T.Y.; Lee, H.S. Lithospermic acid B isolated from Salvia miltiorrhiza ameliorates ischemia/reperfusion-induced renal injury in rats. Life Sci. 2004, 75, 1801–1816. [Google Scholar] [CrossRef]
- Wu, Z.M.; Wen, T.; Tan, Y.F.; Liu, Y.; Ren, F.; Wu, H. Effects of salvianolic acid a on oxidative stress and liver injury induced by carbon tetrachloride in rats. Basic Clin. Pharmacol. Toxicol. 2007, 100, 115–120. [Google Scholar] [CrossRef]
- Li, J.; Xu, M.; Fan, Q.; Xie, X.; Zhang, Y.; Mu, D.; Zhao, P.; Zhang, B.; Cao, F.; Wang, Y.; et al. Tanshinone IIA ameliorates seawater exposure-induced lung injury by inhibiting aquaporins (AQP) 1 and AQP5 expression in lung. Respir. Physiol. Neurobiol. 2011, 176, 39–49. [Google Scholar] [CrossRef]
- Che, X.H.; Park, E.J.; Zhao, Y.Z.; Kim, W.H.; Sohn, D.H. Tanshinone II A induces apoptosis and S phase cell cycle arrest in activated rat hepatic stellate cells. Basic Clin. Pharmacol. Toxicol. 2010, 106, 30–37. [Google Scholar] [CrossRef]
- Isacchi, B.; Fabbri, V.; Galeotti, N.; Bergonzi, M.C.; Karioti, A.; Ghelardini, C.; Vannucchi, M.G.; Bilia, A.R. Salvianolic acid B and its liposomal formulations: Anti-hyperalgesic activity in the treatment of neuropathic pain. Eur. J. Biochem. 2011, 44, 552–558. [Google Scholar] [CrossRef]
- Yang, X.Y.; Qiang, G.F.; Zhang, L.; Zhu, X.M.; Wang, S.B.; Sun, L.; Yang, H.G.; Du, G.H. Salvianolic acid A protects against vascular endothelial dysfunction in high-fat diet fed and streptozotocin-induced diabetic rats. J. Asian Nat. Prod. Res. 2011, 13, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Serra, S.; Vacca, G.; Tumatis, S.; Carrucciu, A.; Morazzoni, P.; Bombardelli, E.; Colombo, G.; Gessa, G.L.; Carai, M.A. Anti-relapse properties of IDN 5082, a standardized extract of Salvia miltiorrhiza, in alcohol-preferring rats. J. Ethnopharmacol. 2003, 88, 249–252. [Google Scholar] [CrossRef]
- Lu, S. The Salvia Miltiorrhiza Genome; Springer: Cham, Switzerland, 2019; pp. 129–139. [Google Scholar]
- Liao, P.; Hemmerlin, A.; Bach, T.J.; Chye, M.L. The potential of the mevalonate pathway for enhanced isoprenoid production. Biotechnol. Adv. 2016, 34, 697–713. [Google Scholar] [CrossRef] [PubMed]
- Hemmerlin, A. Post-translational events and modifications regulating plant enzymes involved in isoprenoid precursor biosynthesis. Plant Sci. 2013, 203–204, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Leivar, P.; Antolín-Llovera, M.; Ferrero, S.; Closa, M.; Arró, M.; Ferrer, A.; Boronat, A.; Campos, N. Multilevel control of Arabidopsis 3-hydroxy-3-methylglutaryl coenzyme A reductase by protein phosphatase 2A. Plant Cell 2011, 23, 1494–1511. [Google Scholar] [CrossRef] [Green Version]
- Liao, P.; Zhou, W.; Zhang, L.; Wang, J.; Yan, X.; Zhang, Y.; Zhang, R.; Li, L.; Zhou, G.; Kai, G. Molecular cloning, characterization and expression analysis of a new gene encoding 3-hydroxy-3-methylglutaryl coenzyme A reductase from Salvia miltiorrhiza. Acta Physiol. Plant. 2009, 31, 565–572. [Google Scholar] [CrossRef]
- Dai, Z.; Cui, G.; Zhou, S.F.; Zhang, X.; Huang, L. Cloning and characterization of a novel 3-hydroxy-3-methylglutaryl coenzyme A reductase gene from Salvia miltiorrhiza involved in diterpenoid tanshinone accumulation. J. Plant Physiol. 2011, 168, 148–157. [Google Scholar] [CrossRef]
- Ma, Y.; Yuan, L.; Wu, B.; Li, X.; Chen, S.; Lu, S. Genome-wide identification and characterization of novel genes involved in terpenoid biosynthesis in Salvia miltiorrhiza. J. Exp. Bot. 2012, 63, 2809–2823. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.P.; Lin, J.J.; Li, W.H. Positional distribution of transcription factor binding sites in Arabidopsis thaliana. Sci. Rep. 2016, 6, 25164. [Google Scholar] [CrossRef]
- Hussain, T.; Rehman, N.; Inam, S.; Ajmal, W.; Afroz, A.; Muhammad, A.; Zafar, Y.; Ali, G.M.; Khan, M.R. Homotypic clusters of transcription factor binding sites in the first large intron of AGL24 MADS-box transcription factor are recruited in the enhancement of floral expression. Plant Mol. Biol. Rep. 2019, 37, 24–40. [Google Scholar] [CrossRef]
- Allen, B.L.; Taatjes, D.J. The Mediator complex: A central integrator of transcription. Nat. Rev. Mol. Cell Biol. 2015, 16, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Khan, D.; Ziegler, D.J.; Kalichuk, J.L.; Hoi, V.; Huynh, N.; Hajihassani, A.; Parkin, I.A.P.; Robinson, S.J.; Belmonte, M.F. Gene expression profiling reveals transcription factor networks and subgenome bias during Brassica napus seed development. Plant J. 2022, 109, 477–489. [Google Scholar] [CrossRef]
- Engels, B.M.; Hutvagner, G. Principles and effects of microRNA-mediated post-transcriptional gene regulation. Oncogene 2006, 25, 6163–6169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, R.S.; Bhattacharyya, S.N.; Filipowicz, W. Repression of protein synthesis by miRNAs: How many mechanisms? Trends Cell Biol. 2007, 17, 118–126. [Google Scholar] [CrossRef]
- Yang, G.; Li, Y.; Wu, B.; Zhang, K.; Gao, L.; Zheng, C. MicroRNAs transcriptionally regulate promoter activity in Arabidopsis thaliana. J. Integr. Plant Biol. 2019, 61, 1128–1133. [Google Scholar] [CrossRef]
- Ye, R.; Wang, W.; Iki, T.; Liu, C.; Wu, Y.; Ishikawa, M.; Zhou, X.; Qi, Y. Cytoplasmic assembly and selective nuclear import of Arabidopsis Argonaute4/siRNA complexes. Mol. Cell 2012, 46, 859–870. [Google Scholar] [CrossRef] [Green Version]
- Place, R.F.; Li, L.C.; Pookot, D.; Noonan, E.J.; Dahiya, R. MicroRNA-373 induces expression of genes with complementary promoter sequences. Proc. Natl. Acad. Sci. USA 2008, 105, 1608–1613. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Saetrom, P.; Snøve, O., Jr.; Rossi, J.J. MicroRNA-directed transcriptional gene silencing in mammalian cells. Proc. Natl. Acad. Sci. USA 2008, 105, 16230–16235. [Google Scholar] [CrossRef] [Green Version]
- Holland, C.K.; Jez, J.M. Arabidopsis: The original plant chassis organism. Plant Cell Rep. 2018, 37, 1359–1366. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B. Biology in Bloom: A Primer on the Arabidopsis thaliana Model System. Genetics 2018, 208, 1337–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provart, N.J.; Alonso, J.; Assmann, S.M.; Bergmann, D.; Brady, S.M.; Brkljacic, J.; Browse, J.; Chapple, C.; Colot, V.; Cutler, S.; et al. 50 years of Arabidopsis research: Highlights and future directions. New Phytol. 2016, 209, 921–944. [Google Scholar] [CrossRef]
- Cantó-Pastor, A.; Mason, G.A.; Brady, S.M.; Provart, N.J. Arabidopsis bioinformatics: Tools and strategies. Plant J. 2021, 108, 1585–1596. [Google Scholar] [CrossRef] [PubMed]
- Dey, N.; Sarkar, S.; Acharya, S.; Maiti, I.B. Synthetic promoters in planta. Planta 2015, 242, 1077–1094. [Google Scholar] [CrossRef]
- Shi, M.; Wang, Y.; Wang, X.; Deng, C.; Cao, W.; Hua, Q.; Kai, G. Simultaneous promotion of tanshinone and phenolic acid biosynthesis in Salvia miltiorrhiza hairy roots by overexpressing Arabidopsis MYC2. Ind. Crops Prod. 2020, 155, 112826. [Google Scholar] [CrossRef]
- Deng, C.; Hao, X.; Shi, M.; Fu, R.; Wang, Y.; Zhang, Y.; Zhou, W.; Feng, Y.; Makunga, N.P.; Kai, G. Tanshinone production could be increased by the expression of SmWRKY2 in Salvia miltiorrhiza hairy roots. Plant Sci. 2019, 284, 1–8. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Z.; Ji, A.; Luo, H.; Song, J. Genomic survey of bZIP transcription factor genes related to tanshinone biosynthesis in Salvia miltiorrhiza. Acta Pharm. Sin. B 2018, 8, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Luo, H.; Xu, Z.; Zhu, Y.; Ji, A.; Song, J.; Chen, S. Genome-wide characterisation and analysis of bHLH transcription factors related to tanshinone biosynthesis in Salvia miltiorrhiza. Sci. Rep. 2015, 5, 11244. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, Y.; Guo, J.; Yang, J.; Zhang, X.; Wang, Z.; Cheng, Y.; Du, Z.; Qi, Z.; Huang, Y.; et al. Integrated Transcriptomics and Proteomics to Reveal Regulation Mechanism and Evolution of SmWRKY61 on Tanshinone Biosynthesis in Salvia miltiorrhiza and Salvia castanea. Front. Plant Sci. 2021, 12, 820582. [Google Scholar] [CrossRef]
- Shahmuradov, I.A.; Umarov, R.K.; Solovyev, V.V. TSSPlant: A new tool for prediction of plant Pol II promoters. Nucleic Acids Res. 2017, 45, e65. [Google Scholar] [CrossRef]
- Molina, C.; Grotewold, E. Genome wide analysis of Arabidopsis core promoters. BMC Genom. 2005, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Szymczyk, P.; Grąbkowska, R.; Skała, E.; Żebrowska, M.; Balcerczak, E.; Jeleń, A. Isolation and characterization of a 3-hydroxy-3-methylglutaryl coenzyme A reductase 2 promoter from Salvia miltiorrhiza. J. Plant Biochem. Biotechnol. 2018, 27, 223–236. [Google Scholar] [CrossRef]
- Tirosh, I.; Weinberger, A.; Carmi, M.; Barkai, N. A genetic signature of interspecies variations in gene expression. Nat. Genet. 2006, 38, 830–834. [Google Scholar] [CrossRef]
- Yang, C.; Bolotin, E.; Jiang, T.; Sladek, F.M.; Martinez, E. Prevalence of the initiator over the TATA box in human and yeast genes and identification of DNA motifs enriched in human TATA-less core promoters. Gene 2007, 389, 52–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basehoar, A.D.; Zanton, S.J.; Pugh, B.F. Identification and distinct regulation of yeast TATA box-containing genes. Cell 2004, 116, 699–709. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.H.; Han, H.W.; Moon, J. Functional analysis of the molecular interactions of TATA box-containing genes and essential genes. PLoS ONE 2015, 10, e0120848. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, W.J.; O’Duibhir, E.; Lijnzaad, P.; van Leenen, D.; Groot Koerkamp, M.J.; Kemmeren, P.; Holstege, F.C. Molecular mechanisms that distinguish TFIID housekeeping from regulatable SAGA promoters. EMBO J. 2017, 36, 274–290. [Google Scholar] [CrossRef] [PubMed]
- Cliften, P.; Sudarsanam, P.; Desikan, A.; Fulton, L.; Fulton, B.; Majors, J.; Waterston, R.; Cohen, B.A.; Johnston, M. Finding functional features in Saccharomyces genomes by phylogenetic footprinting. Science 2003, 301, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinces, M.D.; Legendre, M.; Caldara, M.; Hagihara, M.; Verstrepen, K.J. Unstable tandem repeats in promoters confer transcriptional evolvability. Science 2009, 324, 1213–1216. [Google Scholar] [CrossRef] [Green Version]
- Davey, C.; Pennings, S.; Allan, J. CpG methylation remodels chromatin structure in vitro. J. Mol. Biol. 1997, 267, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.C.; Felsenfeld, G. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature 2000, 405, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Henderson, I.R.; Jacobsen, S.E. Epigenetic inheritance in plants. Nature 2007, 447, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Saulière, J.; Sureau, A.; Expert-Bezançon, A.; Marie, J. The polypyrimidine tract binding protein (PTB) represses splicing of exon 6B from the beta-tropomyosin pre-mRNA by directly interfering with the binding of the U2AF65 subunit. Mol. Cell. Biol. 2006, 26, 8755–8769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirosh, I.; Barkai, N. Evolution of gene sequence and gene expression are not correlated in yeast. Trends Genet. 2008, 24, 109–113. [Google Scholar] [CrossRef]
- Chen, I.J.; Lee, M.S.; Lin, M.K.; Ko, C.Y.; Chang, W.T. Blue light decreases tanshinone IIA content in Salvia miltiorrhiza hairy roots via genes regulation. J. Photochem. Photobiol. B 2018, 183, 164–171. [Google Scholar] [CrossRef]
- Wang, C.H.; Zheng, L.P.; Tian, H.; Wang, J.W. Synergistic effects of ultraviolet-B and methyl jasmonate on tanshinone biosynthesis in Salvia miltiorrhiza hairy roots. J. Photochem. Photobiol. B 2016, 159, 93–100. [Google Scholar] [CrossRef]
- Hao, X.; Shi, M.; Cui, L.; Xu, C.; Zhang, Y.; Kai, G. Effects of methyl jasmonate and salicylic acid on tanshinone production and biosynthetic gene expression in transgenic Salvia miltiorrhiza hairy roots. Biotechnol. Appl. Biochem. 2015, 62, 24–31. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, S.; Zhang, J.; Ma, P.; Duan, J.; Liang, Z. Effect and mechanism of endophytic bacteria on growth and secondary metabolite synthesis in Salvia miltiorrhiza hairy roots. Acta Physiol. Plant. 2014, 36, 1095–1105. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, S.; Yang, D.; Zhang, J.; Liang, Z. Effects of Streptomyces pactum Act12 on Salvia miltiorrhiza hairy root growth and tanshinone synthesis and its mechanisms. Appl. Biochem. Biotechnol. 2014, 173, 883–893. [Google Scholar] [CrossRef]
- Yang, D.; Ma, P.; Liang, X.; Wei, Z.; Liang, Z.; Liu, Y.; Liu, F. PEG and ABA trigger methyl jasmonate accumulation to induce the MEP pathway and increase tanshinone production in Salvia miltiorrhiza hairy roots. Physiol. Plant. 2012, 146, 173–183. [Google Scholar] [CrossRef]
- Yu, W.; Yu, Y.; Wang, C.; Zhang, Z.; Xue, Z. Mechanism by which salt stress induces physiological responses and regulates tanshinone synthesis. Plant Physiol. Biochem. 2021, 164, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, J.; Islam, F.; Najeeb, U.; Pan, J.; Hou, Z.; Shou, J.; Qin, J.; Xu, L. 5-Aminolevulinic acid could enhance the salinity tolerance by alleviating oxidative damages in Salvia miltiorrhiza. Food Sci. Technol. 2021, 42, 1–9. [Google Scholar] [CrossRef]
- Setlow, J.K. Genetic Engineering. Principles and Methods, 1st ed.; Springer Science Business Media LLC: New York, NY, USA, 1997; pp. 15–47. [Google Scholar]
- Reeve, J.N. Archaeal chromatin and transcription. Mol. Microbiol. 2003, 48, 587–598. [Google Scholar] [CrossRef] [Green Version]
- Matthews, J.M. Protein Dimerization and Oligomerization in Biology; Springer Science+Business Media: New York, NY, USA, 2012; pp. 1–170. [Google Scholar]
- Amoutzias, G.D.; Robertson, D.L.; Van de Peer, Y.; Oliver, S.G. Choose your partners: Dimerization in eukaryotic transcription factors. Trends Biochem. Sci. 2008, 33, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef]
- San-Bento, R.; Farcot, E.; Galletti, R.; Creff, A.; Ingram, G. Epidermal identity is maintained by cell-cell communication via a universally active feedback loop in Arabidopsis thaliana. Plant J. 2014, 77, 46–58. [Google Scholar] [CrossRef]
- Rombolá-Caldentey, B.; Rueda-Romero, P.; Iglesias-Fernández, R.; Carbonero, P.; Oñate-Sánchez, L. Arabidopsis DELLA and two HD-ZIP transcription factors regulate GA signaling in the epidermis through the L1 box cis-element. Plant Cell 2014, 26, 2905–2919. [Google Scholar] [CrossRef] [Green Version]
- Abe, M.; Takahashi, T.; Komeda, Y. Identification of a cis-regulatory element for L1 layer-specific gene expression, which is targeted by an L1-specific homeodomain protein. Plant J. 2001, 26, 487–494. [Google Scholar] [CrossRef]
- Cheng, X.; Zhao, Y.; Jiang, Q.; Yang, J.; Zhao, W.; Taylor, I.A.; Peng, Y.L.; Wang, D.; Liu, J. Structural basis of dimerization and dual W-box DNA recognition by rice WRKY domain. Nucleic Acids Res. 2019, 47, 4308–4318. [Google Scholar] [CrossRef] [Green Version]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Ciolkowski, I.; Wanke, D.; Birkenbihl, R.P.; Somssich, I.E. Studies on DNA-binding selectivity of WRKY transcription factors lend structural clues into WRKY-domain function. Plant Mol. Biol. 2008, 68, 81–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Q.H.; Saijo, Y.; Mauch, S.; Biskup, C.; Bieri, S.; Keller, B.; Seki, H.; Ulker, B.; Somssich, I.E.; Schulze-Lefert, P. Nuclear activity of MLA immune receptors links isolate-specific and basal disease-resistance responses. Science 2007, 315, 1098–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagisawa, S. Dof DNA-binding domains of plant transcription factors contribute to multiple protein-protein interactions. Eur. J. Biochem. 1997, 250, 403–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Jiang, Q.; Ma, X.; Ying, Q.; Shen, B.; Qian, Y.; Song, H.; Wang, H. Deep sequencing identifies tissue-specific microRNAs and their target genes involving in the biosynthesis of tanshinones in Salvia miltiorrhiza. PLoS ONE 2014, 9, e111679. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Qureshi, M.I.; Kamaluddin; Alam, T.; Abdin, M.Z. Protocol for isolation of genomic DNA from dry and fresh roots of medicinal plants suitable for RAPD and restriction digestion. Afr. J. Biotechnol. 2007, 6, 175–178. [Google Scholar] [CrossRef]
- Majewska, M.; Wysokińska, H.; Kuźma, Ł.; Szymczyk, P. Eukaryotic and prokaryotic promoter databases as valuable tools in exploring the regulation of gene transcription: A comprehensive overview. Gene 2018, 644, 38–48. [Google Scholar] [CrossRef]
- Shahmuradov, I.A.; Gammerman, A.J.; Hancock, J.M.; Bramley, P.M.; Solovyev, V.V. PlantProm: A database of plant promoter sequences. Nucleic Acids Res. 2003, 31, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Chow, C.N.; Zheng, H.Q.; Wu, N.Y.; Chien, C.H.; Huang, H.D.; Lee, T.Y.; Chiang-Hsieh, Y.F.; Hou, P.F.; Yang, T.Y.; Chang, W.C. PlantPAN 2.0: An update of plant promoter analysis navigator for reconstructing transcriptional regulatory networks in plants. Nucleic Acids Res. 2016, 44, D1154–D1160. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Toufighi, K.; Brady, S.M.; Austin, R.; Ly, E.; Provart, N.J. The Botany Array Resource: E-Northerns, Expression Angling, and promoter analyses. Plant J. 2005, 43, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, F.; Yu, Y.; Feng, L.; Jia, J.; Liu, B.; Li, B.; Guo, H.; Zhai, J. A Comprehensive Online Database for Exploring ~20,000 Public Arabidopsis RNA-Seq Libraries. Mol. Plant 2020, 13, 1231–1233. [Google Scholar] [CrossRef] [PubMed]
- UniProt, C. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Oughtred, R.; Rust, J.; Chang, C.; Breitkreutz, B.J.; Stark, C.; Willems, A.; Boucher, L.; Leung, G.; Kolas, N.; Zhang, F.; et al. The BioGRID database: A comprehensive biomedical resource of curated protein, genetic, and chemical interactions. Protein Sci. 2021, 30, 187–200. [Google Scholar] [CrossRef]
- Chow, C.N.; Lee, T.Y.; Hung, Y.C.; Li, G.Z.; Tseng, K.C.; Liu, Y.H.; Kuo, P.L.; Zheng, H.Q.; Chang, W.C. PlantPAN3.0: A new and updated resource for reconstructing transcriptional regulatory networks from ChIP-seq experiments in plants. Nucleic Acids Res. 2019, 47, D1155–D1163. [Google Scholar] [CrossRef] [Green Version]
- Luján, M.A.; Soria-García, Á.; Claver, A.; Lorente, P.; Rubio, M.C.; Picorel, R.; Alfonso, M. Different Cis-Regulatory Elements Control the Tissue-Specific Contribution of Plastid ω-3 Desaturases to Wounding and Hormone Responses. Front. Plant Sci. 2021, 12, 727292. [Google Scholar] [CrossRef]
- Yuan, L.; Perry, S.E. Plant Transcription Factors, 1st ed.; Humana Press: Totowa, NJ, USA, 2011; pp. 45–66. [Google Scholar]
- Zhang, X.; Ge, F.; Deng, B.; Shah, T.; Huang, Z.; Liu, D.; Chen, C. Molecular Cloning and Characterization of PnbHLH1 Transcription Factor in Panax notoginseng. Molecules 2017, 22, 1268. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Cao, Y.; Wang, W.; Li, Y.; Wang, D.; Wang, S.; Cao, X. Transcription factor SmSPL7 promotes anthocyanin accumulation and negatively regulates phenolic acid biosynthesis in Salvia miltiorrhiza. Plant Sci. 2021, 310, 110993. [Google Scholar] [CrossRef]
- Chen, X.; Bhadauria, V.; Ma, B. ChIP-Seq: A Powerful Tool for Studying Protein-DNA Interactions in Plants. Curr. Issues Mol. Biol. 2018, 27, 171–180. [Google Scholar] [CrossRef]
- Chen, H.; Yang, Q.; Fu, H.; Chen, K.; Zhao, S.; Zhang, C.; Cai, T.; Wang, L.; Lu, W.; Dang, H.; et al. Identification of Key Gene Networks and Deciphering Transcriptional Regulators Associated With Peanut Embryo Abortion Mediated by Calcium Deficiency. Front. Plant Sci. 2022, 13, 814015. [Google Scholar] [CrossRef]
- Lei, X.; Liu, Z.; Xie, Q.; Fang, J.; Wang, C.; Li, J.; Wang, C.; Gao, C. Construction of two regulatory networks related to salt stress and lignocellulosic synthesis under salt stress based on a Populus davidiana × P. bolleana transcriptome analysis. Plant Mol. Biol. 2022. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcription Factor (TF) Family Name | Fragment of Promoter Sequence with Underlined TFBSs a and Potentially Interacting TF Pairs |
|---|---|
| ˗ | AGAAAAATGGAATAAGAAGA two salicylic acid (SA)-responsive TCA elements (AGAAAA and AGAAGA) spaced by eight nucleotides |
| ˗ | ATCTCCAATCT two LREs (ATCT) spaced by three nucleotides |
| HD-ZIP | atTTAATgtaTTCATAAATata ATML1/HDG1 and PDF2/ATML1 spaced by four nucleotides |
| WRKY | AGTCATAACAATGTCAA WRKY2/WRKY14/WRKY45/WRKY57/WRKY69 and WRKY2/WRKY14/WRKY45/WRKY57/WRKY69 spaced by six nucleotides |
| Dof | AAAGAAAAAAGA DOF5.4 and DOF5.4 spaced by two nucleotides |
| miRNA Name and Source | miRNA Sequence | Sequence Alignment Position Start/End | Strand | e-Value |
|---|---|---|---|---|
| HMGR4Promoter | ||||
| miR1128 Saccharum sp. | UACUACUCCCUCCGUCCCAAA | 350/368 405/423 | + − | 0.75 4.2 |
| miR6462c-5p Populus trichocarpa | AAGGGACAAAAAUGGCAUAAGA | 259/279 | − | 3.5 |
| miR1128 Triticum aestivum | UACUACUCCCUCCGUCCGAAA | 350/368 | + | 4.2 |
| miR1436 Oryza sativa and Hordeum vulgare | ACAUUAUGGGACGGAGGGAGU | 354/368 | − | 6.2 |
| miR5205a Medicago truncatula | CAUACAAUUUGGGACGGAGGGAG | 355/374 | − | 9.1 |
| miR8740 Gossypium raimondii | UAAUGAUGUGGCACAAUAUUA | 634/653 | − | 9.1 |
| miR11573a and miR11573b Picea abies | UUGGGGAGCGUAUUGUAGAUU | 197/216 | − | 9.1 |
| 5′UTR of HMGR4 | ||||
| miR477 Gossypium raimondii | CGAAGUCUUGGAAGAGAGUAA | 59/75 | − | 3.2 |
| miR6180 Hordeum vulgare | AGGGUGGAAGAAAGAGGGCG | 55/69 | − | 3.9 |
| miR4993 Glycine max | GAGCGGCGGCGGUGGAGGAUG | 13/30 | − | 6.9 |
| miR12107-5p Citrus sinensis | CUGAUGAGAGAGCGAAUGAUA | 51/66 | − | 8.4 |
| TF Family Name | TF Gene Name and Locus | Processes in Which TF is Involved a | r-Value b | TFBS Motif and Localisation c,d | |
|---|---|---|---|---|---|
| Homeodomain; HD-ZIP | ATHB-13; At1g69780 | cotyledon and leaf morphogenesis; primary root development; sucrose-signalling pathway | 0.594 | ATAAT 310; 309 | AATAA 308; 307 |
| ATHB-16; At4g40060 | regulation of timing of transition from vegetative to reproductive phase; repression of cell expansion during plant development; response to blue light | 0.562 | ATAAT 309 | ||
| HDG1; At3g61150 | maintenance of floral organ identity | 0.507 | ATTAA 161 | TTAAT 1218; 1331; 1332 | |
| ANL2; At4g00730 | regulation of tissue-specific accumulation of anthocyanins; cellular organisation of primary root; cuticle hydrocarbon biosynthetic process; plant-type cell wall modification; root hair cell differentiation | 0.524 | TTAAT 1218 | ATTAA 1161 | |
| ATML1; At4g21750 | cotyledon development; epidermal cell differentiation; seed dormancy and germination | 0.583 | TTAAT 1332 ATTTA 1057; 1282 | TAAAT 987; 1272; 1346 | |
| PDF2; At4g04890 | cotyledon development; epidermal cell differentiation; seed dormancy and germination; maintenance of floral organ identity | 0.587 | ATTTA 1057; 1282 | TAAAT 987; 1272; 1346 | |
| Homeodomain; bZIP; HD-ZIP | HAT5; At3g01470 | leaf morphogenesis; response to blue light and salt stress | 0.521 | ATAAT 307; 310 | AATAA 308; 307 |
| bZIP | BZIP25; At3g54620 | positive regulation of seed maturation | 0.666 | CCACG 822 TACGT 46; 720 ACGTA 47; 721 | AACGT 290;1188 ACGTT 291; 1189 |
| WRKY | WRKY2; At5g56270 | regulation of basal cell division patterns during early embryogenesis; establishment of cell polarity; longitudinal axis specification; pollen development | 0.575/ 0.557 | TGACT 5; 832 AGTCA 111; 1240 | TTGAC 831 GTCAA 913; 1147; 1251 |
| WRKY14; At1g30650 | ˗ | 0.576 | |||
| WRKY57; At1g69310 | response to osmotic stress, salt stress and water deprivation | 0.504 | |||
| WRKY45; At3g01970 | phosphate ion transport | 0.515 | TTGAC 830; 831 TGACT 5; 832 | AGTCA 111; 1240 GTCAA 913; 1147; 1251 | |
| WRKY69; At3g58710 | ˗ | 0.564 | |||
| Myb/SANT; ARR-B | ARR2; At4g16110 | His-to-Asp phosphorelay signal transduction system; expression of nuclear genes for components of mitochondrial complex I; ethylene- and cytokinin-activated signalling pathways; promotion of cytokinin-mediated leaf longevity; root meristem growth; seed growth; stomatal movement | 0.552/0.611 | AATCT 15; 197; 919 AGATT 1405 | AATCC 179; 204; 548 |
| ARR14; At2g01760 | His-to-Asp phosphorelay signal transduction system; activation of some type-A response regulators in response to cytokinins | 0.509/ 0.530 | |||
| Myb/SANT; MYB | MYB6; At4g09460 | response to ethylene, abscisic acid (ABA), indole-3-acetic acid, and Pseudomonas syringae pv. phaseolica | 0.599 | ACCTA 886 | |
| MYB-related | RVE1; At5g17300 | morning-phased TF integrating circadian clock and auxin pathways; regulation of free indole-3-acetic acid level in time-of-day specific manner; negative regulation of freezing tolerance | 0.501 | ATATC 1166 | |
| RVE4; At5g02840 | regulation of circadian rhythm | 0.545/ 0.573 | ATATC 1166 | GATAT 1215 | |
| EIN3; EIL | EIL1; At2g27050 | positive regulation of ethylene response pathway; cellular response to iron ion; defence response to bacterium | 0.554 | TGTAT 374; 759 | ATACA 391 |
| EIL3; At1g73730 | ethylene response pathway; sulphur metabolic process; cellular response to iron ion | 0.525 | ATGTA 757 | ||
| MADS box; MIKC | AGL18; At3g57390 | negative regulation of flowering and short-day photoperiodism; pollen development | 0.567 | TTTCC 804; 801 TTTTG 805 CAAAA 1396 | AGAAA 293; 1402 GGAAA 1364; 1362 TTTTT 77; 78; 79, 80; 81; 82; 83; 84 |
| SVP; At2g22540 | inhibition of floral transition in autonomous flowering pathway; identity of floral meristem; response to temperature stimulus | 0.537 | TTTCC 801 | GGAAA 1362 | |
| NAC; NAM | NAC055; At3g15500 | jasmonic acid-mediated signalling pathway; response to water deprivation | 0.594 | TACGT 44; 718 CGTA 720 ACAT 644 | TTGAC 829 ACGTA 44; 718; 720 |
| NAC072; At4g27410 | activator in ABA-mediated dehydration response | 0.543 | CGTA 720 TTGAC 829 | ACAT 644 | |
| NF-YB | NFYB5; At2g47810 | protein heterodimerization activity | 0.558 | CTAAT 42 ATCGG 102; 131 CCCAT 139 CCAAG 149; 213 CCAAT 195; 202 GTTGG 389 ATGGG 959 ATTGC 1039 CGAAT 1115 ATTAG 1389 CCTAT 890 TTTGG 811 | TCAAT 13; 1149; 1253 AATGG 462; 847 CCACT 144; 380; 1364 CCATT 169; 266; 701 CCAAC 530 AGTGG 592 ATTGA 766; 1099 ACAAT 460; 800; 1247 ATTGT 599; 637; 1145; 1152 CCAAA 176; 534; 1261; 1396 CAAAT 177; 469; 897; 1107; 1161; 1262; 1313 |
| NF-YC | NFYC10; At5g38140 | 0.523 | |||
| TBP | TBP2; At1g55520 | required for basal transcription (facilitating the recruitment of TFIID to the promoter, forming a preinitiation complex with RNA polymerase) | 0.645 | ATATA 746; 739 TTTTA 1022; 1078 TAAAA 541; 1465; 1024 | TATAT 677; 1305; 743; 736 ATAAA 1025; 1466; 1344; 298; 439; 1024; 1465 TTTAT 669; 1016; 1072; 1297; 1223; 1303; 674; 1077; 1302 |
| TCP | TCP21; At5g08330 | positive regulation of circadian clock | 0.546 | CCCAC 818 | |
| AP2; ERF | RAP2-3; At3g16770 | cell death; heat acclimation; ethylene-activated signalling pathway; response to cytokinin, jasmonic acid and other organism | 0.502 | TAAGA 494 | |
| C2H2 | AZF2; At3g19580 | inhibition of plant growth under abiotic stress conditions; negative regulation of ABA signalling during seed germination; positive regulation of leaf senescence; jasmonate early signalling response; response to chitin and water deprivation; plants overexpressing AZF2 have increased sensitivity to salt stress and barely survive under high salt conditions | 0.503 | ACACT 29; 1462 | |
| Dof | DOF5.4; At5g60850 | metal ion binding; binding of OBF TFs to OCS elements | 0.727 | CGTTA 685 AACGT 286; 1184 ACGTT 289; 1187 GCCTT 79 CCTTT 80; 807 AAAGT 109; 982 AAGGG 1031 AAGGA 1372 | ACCTT 158; 184; 605; 612 GCTTT 369 AAAGC 400; 571 AAAGA 939; 1413; 1420 AAAGG 1030; 1289; 1371 TCTTT 252; 328; 586; 874 TCCTT 275; 338; 806; 1156 ACTTT 382; 509; 881; 1021; 1291 |
| SBP | SPL3; At2g33810 | promotion of vegetative phase change and flowering; vegetative- to reproductive-phase transition of meristem | 0.596 | AGTAC 51; 972 GTACA 578; 973 TGTAC 577 GTACT 52 ATACG 43 CGTAG 46 | CTTAC 275; 427; 625 CGTCC 360 GGACG 405 CGAAC 524 CTACG 717 CGTAA 720 |
| TF Matrix Family | Processes in Which TF Is Involved a |
|---|---|
| Arabidopsis homeobox proteins (P$AHBP) | root, leaf and anther development; seed maturation; meristem initiation and growth; xylem and phloem pattern formation; cell differentiation; determination of bilateral symmetry; transition from vegetative to reproductive phase; glucosinolate metabolic process; response to: auxin, gibberellin, ABA, water deprivation, blue light and salt stress |
| L1 box (P$L1BX) | cotyledon development; seed germination and dormancy; epidermal cell differentiation; maintenance of floral organ identity |
| MYB IIG-type binding sites (P$MIIG) | root, seed, stamen and xylem development; cellular cadmium ion homeostasis; gibberellin and flavonol biosynthesis; defence response to fungi; response to: ABA, chitin, salt stress, cold, water deprivation, phosphate starvation, potassium ion and light |
| DNA binding with one finger (P$DOFF) | secondary shoot, cotyledon and seed development; cell wall modification; cell cycle; gibberellin biosynthesis; response to: SA, auxin, chitin, red light and cold |
| GT box elements (P$GTBX) | shoot system and stomatal complex development; trichome morphogenesis; seed maturation and germination; cell size and growth; response to: auxin and water deprivation |
| MADS box proteins (P$MADS) | flower, ovule and seed coat development; seed maturation; meristem structural organisation; transition from vegetative to reproductive phase; short-day photoperiodism; circadian rhythm; response to auxin |
| MYB-like proteins (P$MYBL) | integument, anther and pollen development; leaf morphogenesis; seed growth and dormancy; endothelial cell proliferation; vacuole organisation; wax biosynthesis; long-day photoperiodism; defence response to bacteria and fungi; response to: SA, brassinosteroid, gibberellin, ABA, jasmonic acid, chitin, salt, water deprivation and cold |
| MYB proteins with single DNA binding repeat (P$MYBS) | leaf and lateral root development; leaf senescence; circadian rhythm; peroxidase activity; auxin and sulphate ion homeostasis; response to: ABA, phosphate starvation, absence of light and high light intensity |
| NAC factors with transmembrane motif (P$NTMF) | leaf and trichome morphogenesis; xylem development; seed germination; photoperiodism; membrane protein proteolysis; response to: gibberellin, salt stress |
| plant specific NAC proteins (P$NACF) | leaf and secondary shoot development; primary shoot apical meristem specification; formation of organ boundary; regulation of timing of organ formation; response to water deprivation |
| transcription repressor KANADI (P$KAN1) | phenylpropanoid metabolic process |
| W box family (P$WBXF) | induced systemic resistance; JA-mediated signalling pathway; phosphate ion transport; defence response to: bacteria, fungi and viruses; response to: SA, chitin and wounding |
| time-of-day-specific regulatory elements (P$TODS) | circadian rhythm; red or far-red light signalling pathway; response to temperature |
| nodulin consensus sequence 1 (P$NCS1) | nodule-specific expression |
| zinc finger proteins (P$ZFAT) | regulation of root development; phosphate ion homeostasis |
| light-responsive elements (P$LREM) | response to hypoxia |
| protein secretory pathway elements (P$PSPE) | SA induction of secretion-related genes via NPR1 |
| CGCG box binding proteins (P$CGCG) | leaf senescence; defence response to: bacteria, fungi and insects; response to: cold, auxins and water deprivation |
| proteins involved in programmed cell death response (P$PCDR) | regulation of expression of vacuolar processing enzyme |
| plant nitrate-responsive elements (P$PNRE) | nitrate assimilation; stomatal movement; response to: nitrate and water deprivation |
| stomatal carpenter (P$SCAP) | stomatal movement |
| sweet potato DNA-binding factor with two WRKY domains (P$SPF1) | ˗ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majewska, M.; Kuźma, Ł.; Szymczyk, P. Isolation and Comprehensive in Silico Characterisation of a New 3-Hydroxy-3-Methylglutaryl-Coenzyme A Reductase 4 (HMGR4) Gene Promoter from Salvia miltiorrhiza: Comparative Analyses of Plant HMGR Promoters. Plants 2022, 11, 1861. https://doi.org/10.3390/plants11141861
Majewska M, Kuźma Ł, Szymczyk P. Isolation and Comprehensive in Silico Characterisation of a New 3-Hydroxy-3-Methylglutaryl-Coenzyme A Reductase 4 (HMGR4) Gene Promoter from Salvia miltiorrhiza: Comparative Analyses of Plant HMGR Promoters. Plants. 2022; 11(14):1861. https://doi.org/10.3390/plants11141861
Chicago/Turabian StyleMajewska, Małgorzata, Łukasz Kuźma, and Piotr Szymczyk. 2022. "Isolation and Comprehensive in Silico Characterisation of a New 3-Hydroxy-3-Methylglutaryl-Coenzyme A Reductase 4 (HMGR4) Gene Promoter from Salvia miltiorrhiza: Comparative Analyses of Plant HMGR Promoters" Plants 11, no. 14: 1861. https://doi.org/10.3390/plants11141861