
Stimulation of Germination of Freshly Collected and Cold-Stored Seeds of Ambrosia artemisiifolia L.


Abstract
:1. Introduction
2. Results
2.1. Laboratory Experiments
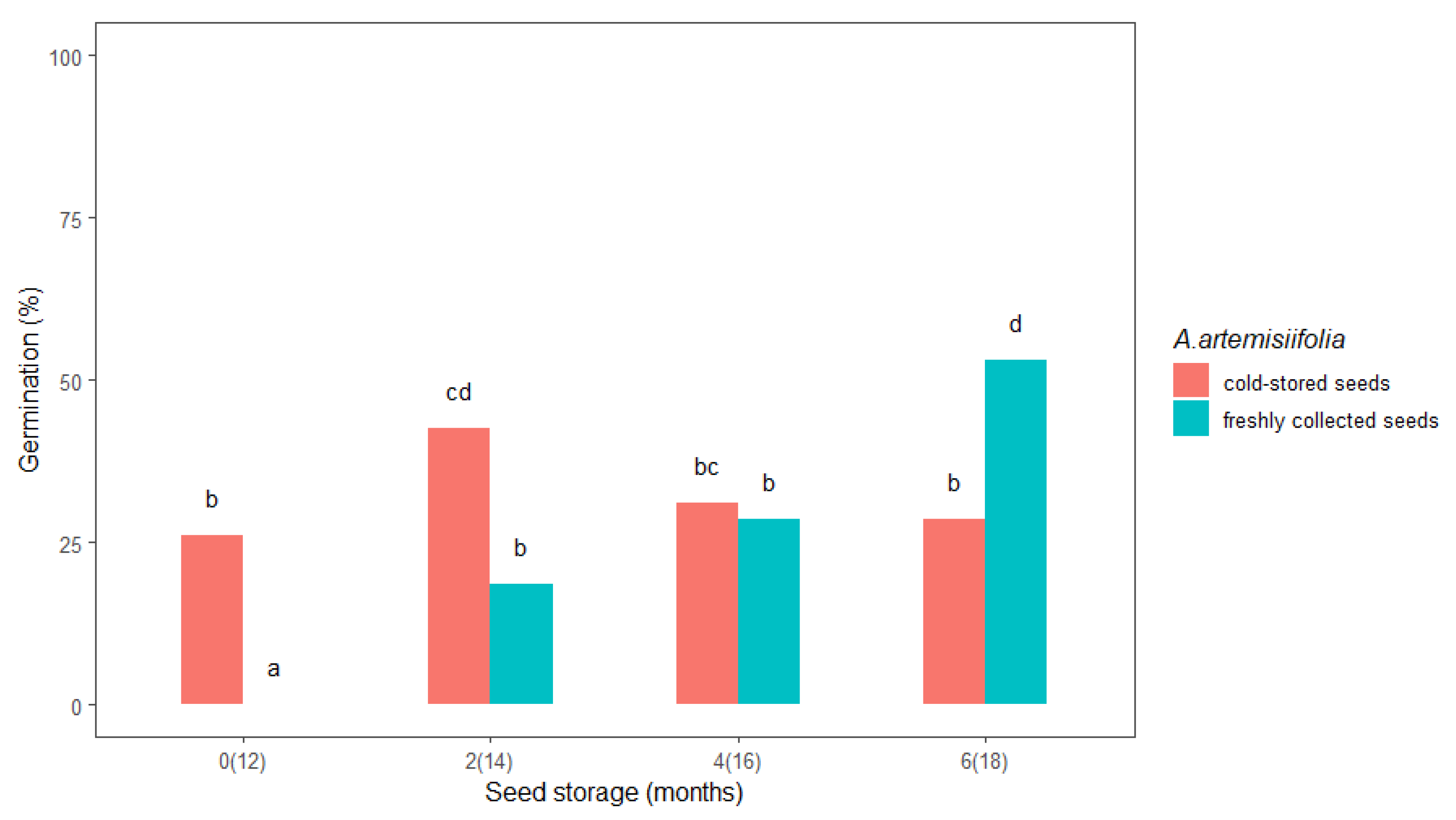
2.1.1. Seed Storage
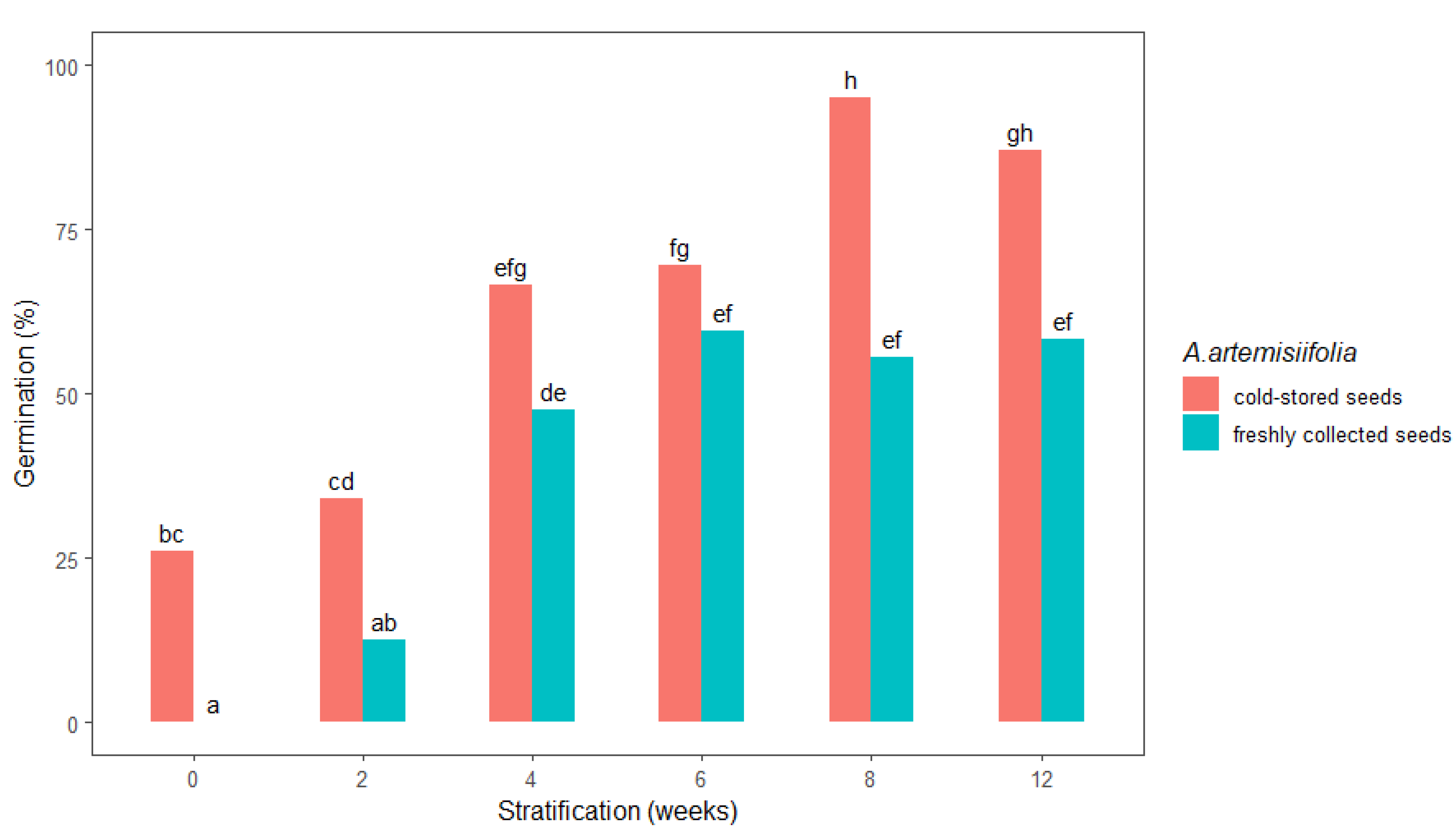
2.1.2. Cold Stratification
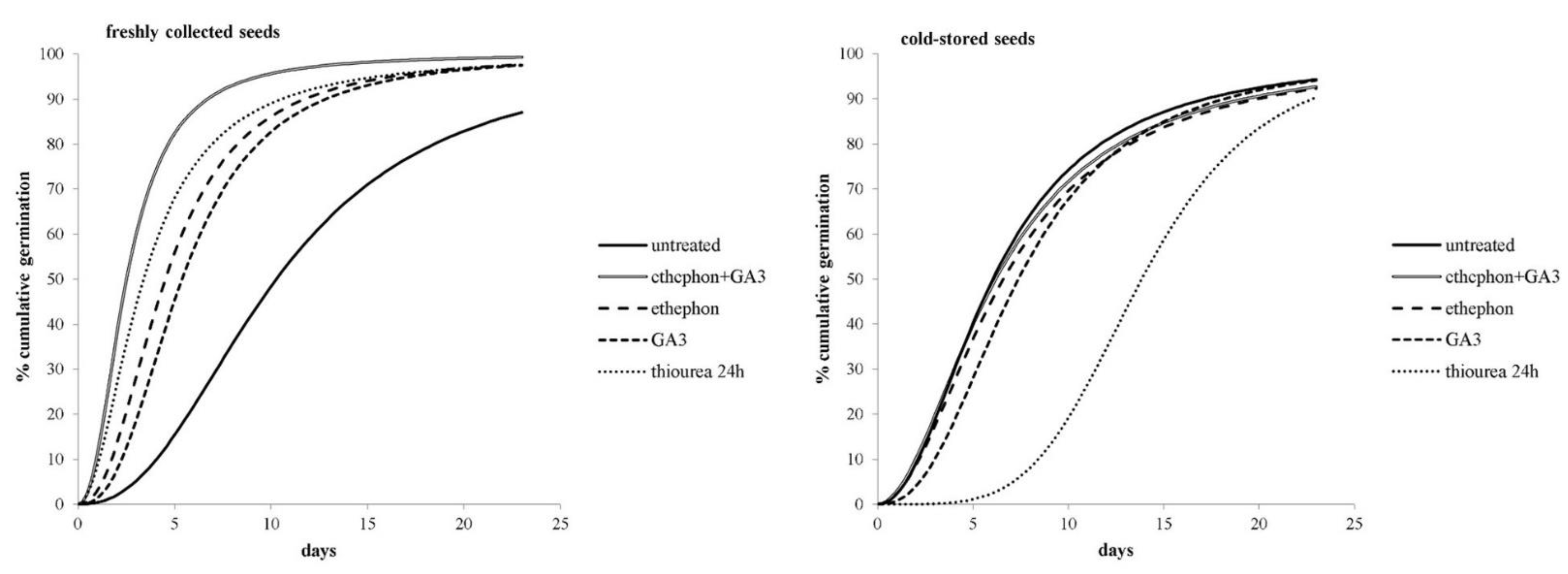
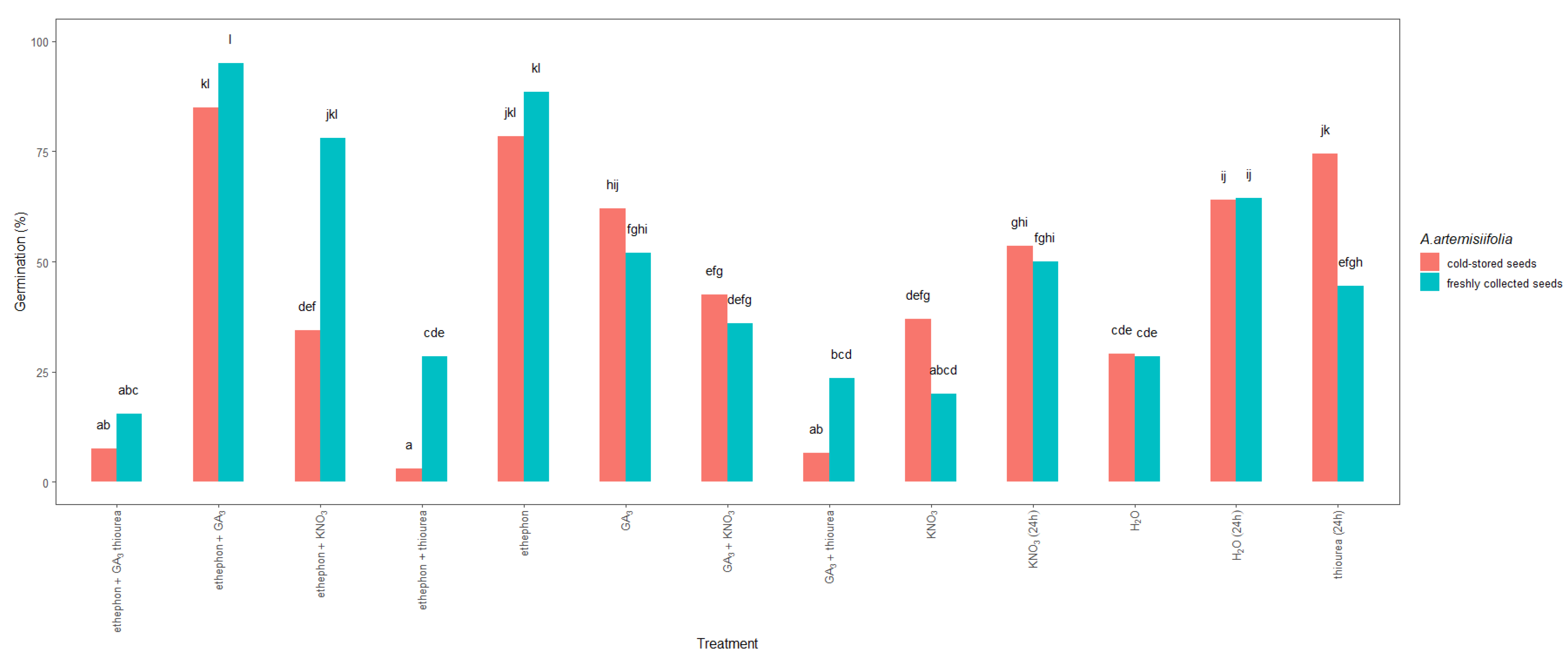
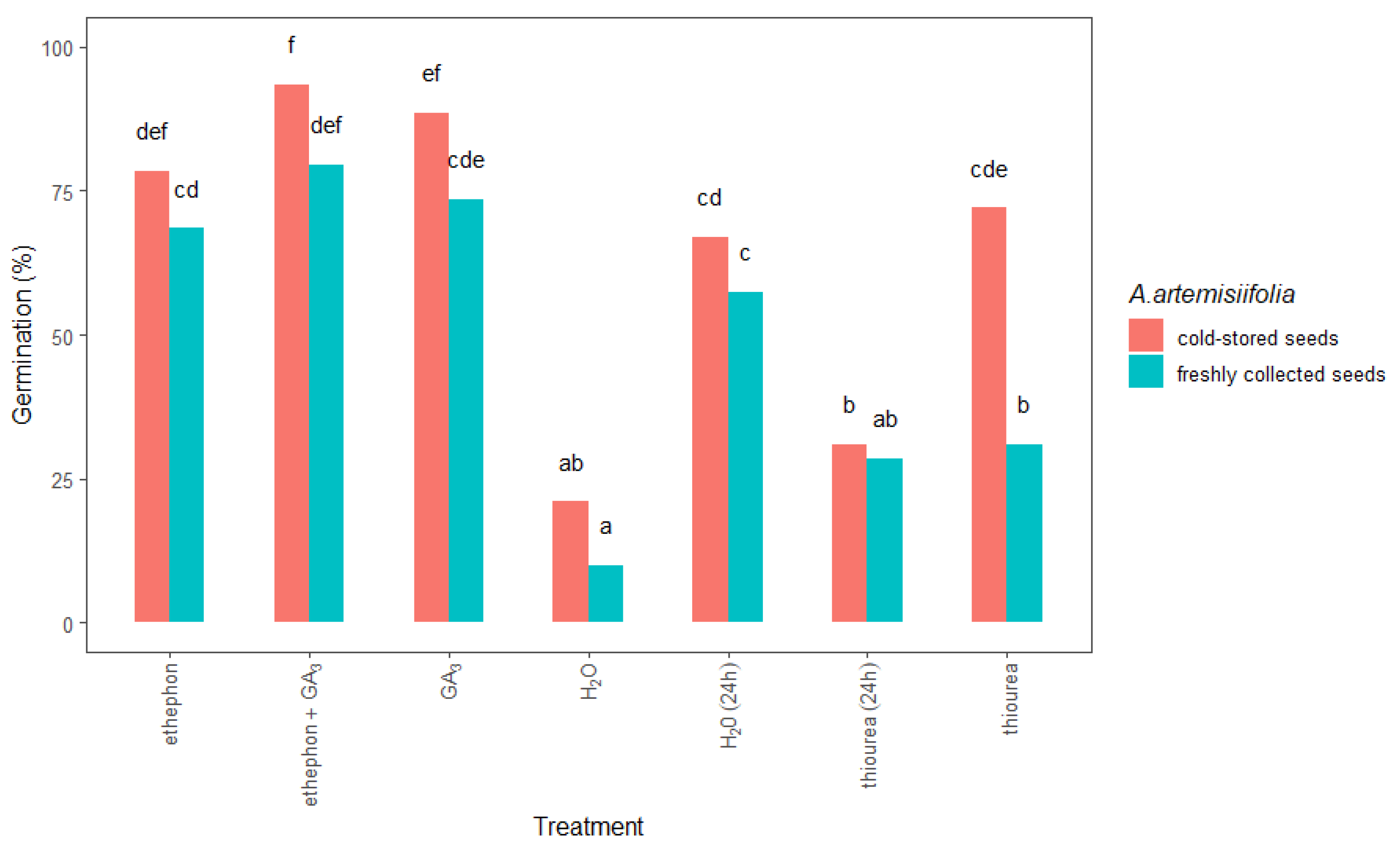
2.1.3. Chemical Treatments
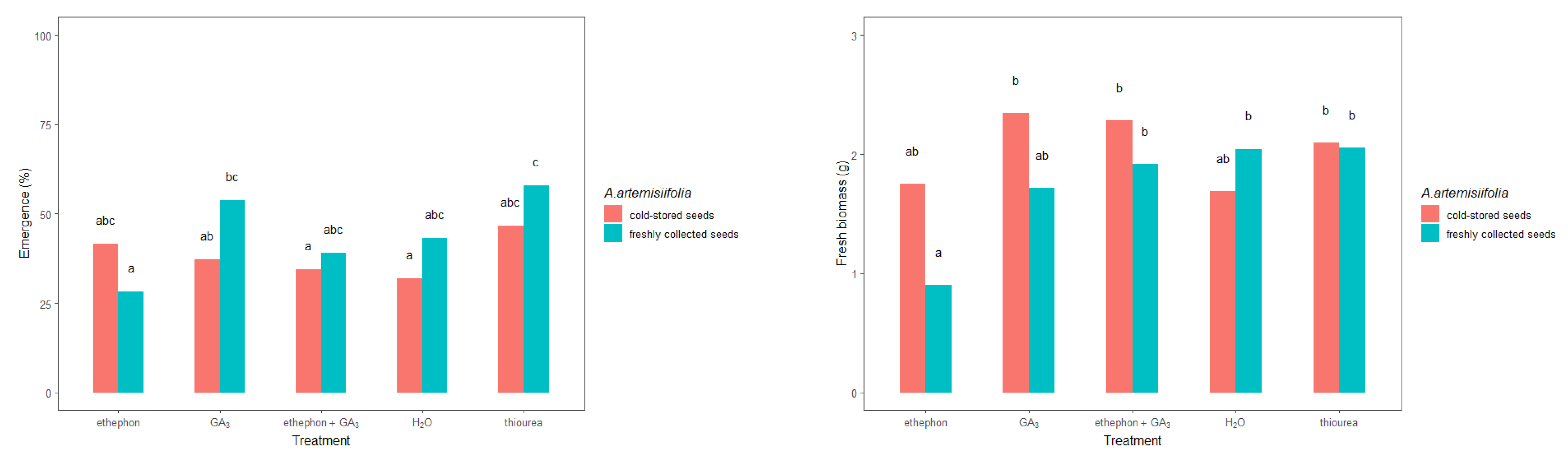
2.2. Greenhouse Experiment
3. Discussion
4. Materials and Methods
4.1. Seed Collection and Storage
4.2. Laboratory Experiments
4.2.1. Cold Stratification
4.2.2. Chemical Treatments
4.2.3. Germination Tests
4.3. Greenhouse Experiment
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Tommaso, A. Germination behavior of common ragweed (Ambrosia artemisiifolia) populations across a range of salinities. Weed Sci. 2004, 52, 1002–1009. [Google Scholar] [CrossRef]
- Pickett, S.T.; Baskin, J.M. The role of temperature and light in the germination behavior of Ambrosia artemisiifolia. Bull. Torrey Bot. Club 1973, 100, 165–170. [Google Scholar] [CrossRef]
- Sang, W.; Liu, X.; Axmacher, J.C. Germination and emergence of Ambrosia artemisiifolia L. under changing environmental conditions in China. Plant Species Biol. 2011, 26, 125–133. [Google Scholar] [CrossRef]
- Brandes, D.; Nitzsche, J. Biology, introduction, dispersal, and distribution of common ragweed (Ambrosia artemisiifolia L.) with special regard to Germany. Nachr. Deut Pflanzenschutzd 2006, 58, 286–291. [Google Scholar]
- Šarić, T.; Ostojić, Z.; Stefanović, L.; Deneva Milanova, S.; Kazinczi, G.; Tyšer, L. The changes of the composition of weed flora in south eastern and central Europe as affected by cropping practices. Herbologia 2011, 12, 8–12. [Google Scholar]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2021. Available online: www.weedsicence.org (accessed on 21 January 2021).
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Fogliatto, S.; Milan, M.; De Palo, F.; Vidotto, F. The effect of various after-ripening temperature regimens on the germination behaviour of Ambrosia artemisiifolia. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2019, 154, 165–172. [Google Scholar] [CrossRef]
- Willemsen, R.W.; Rice, E.L. Mechanism of seed dormancy in Ambrosia artemisiifolia. Am. J. Bot. 1972, 59, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Kelly, R.J. Abscisic acid and gibberellic acid regulation of seed germination and dormancy. Biologist 1969, 51, 91–99. [Google Scholar]
- Ame, R.D. A model of seed dormancy. Bot. Rev. 1968, 34, 1–31. [Google Scholar]
- Corbineau, F.; Xia, Q.W.; Bailly, C.; El-Maarouf-Bouteau, H. Ethylene, a key factor in the regulation of seed dormancy. Front. Plant Sci. 2014, 5, 539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matilla, A.J. Ethylene in seed formation and germination. Seed Sci. Res. 2000, 10, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Ghassemian, M.; Nambara, E.; Cutler, S.; Kawaide, H.; Kamiya, Y.; McCourt, P. Regulation of abscisic acid signaling by the ethylene response pathway in arabidopsis. Plant Cell 2000, 12, 1117–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samimy, C.; Khan, A.A. Secondary dormancy, growth-regulator effects, and embryo growth potential in curly dock (Rumex crispus) seeds. Weed Sci. 1983, 31, 153–158. [Google Scholar] [CrossRef]
- Harre, N.T.; Weller, S.C.; Young, B.G. An improved method to shorten physiological dormancy of giant ragweed (Ambrosia trifida) seed. Weed Sci. 2019, 67, 205–213. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Breaking seed dormancy during dry storage: A useful tool of major problem for successful restoration via direct seeding? Plants 2020, 9, 636. [Google Scholar] [CrossRef]
- Page, E.R.; Nurse, R.E. Comparing physical, chemical, and cold stratification methods for alleviating dormancy of giant ragweed (Ambrosia trifida) seeds. Weed Technol. 2015, 29, 311–317. [Google Scholar] [CrossRef]
- Kazinczi, G.; Biro, K.; Beres, I.; Ferger, B. Intraspecific differences in the germination of common ragweed (Ambrosia artemisiifolia L.). (Fajon belüli), (intraspecifikus) különbségek az ürömlevelű parlagfű (Ambrosia artemisiifolia L.) csírázásában). Növényvédelem 2006, 42, 477–481. [Google Scholar]
- Ciappetta, S.; Ghiani, A.; Gilardelli, F.; Bonini, M.; Citterio, S.; Gentili, R. Invasion of Ambrosia artemisiifolia in Italy: Assessment via analysis of genetic variability and herbarium data. Flora Morphol. Distrib. Funct. Ecol. Plants 2016, 223, 106–113. [Google Scholar] [CrossRef]
- Dinelli, G.; Marotti, I.; Catizone, P.; Bosi, S.; Tanveer, A.; Abbas, R.N.; Pavlovic, D. Germination ecology of Ambrosia artemisiifolia L. and Ambrosia trifida L. biotypes suspected of glyphosate resistance. Cent. Eur. J. Biol. 2013, 8, 286–296. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Secondary dormancy in the seeds of the common ragweed Ambrosia artemisiifolia. Bull. Torrey Bot. Club 1970, 97, 302–305. [Google Scholar] [CrossRef]
- Šoštarčić, V.; Košćak, L.; Culjak, A.; Jerić, I.; Šćepanović, M. Utjecaj hladnog skladištenja na klijanje korovne vrste Ambrosia artemisiifolia L. Fragm. Phytomedica 2020, 34, 16–26. [Google Scholar]
- Boddy, L.G.; Bradford, K.J.; Fischer, A.J. Stratification requirements for seed dormancy alleviation in a wetland weed. PLoS ONE 2013, 8, e71457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tieng, M.T. Germination Inhibition and Promotion of Ragweed (Ambrosia trifida L.) and Smartweed (Polygonum pensylvanicum L.). Ph.D. Thesis, Purdue University, West Lafayette, IN, USA, 1962. [Google Scholar]
- Šoštarčić, V.; Masin, R.; Turčinov, M.; Carin, N.; Šćepanović, M. Intrapopulation variability in morphological and functional properties of Ambrosia artemisiifolia L. seeds. J. Cent. Eur. Agric. 2020, 21, 366–378. [Google Scholar] [CrossRef]
- Vink, J.P.; Soltani, N.; Robinson, D.E.; Tardif, F.J.; Lawton, M.B.; Sikkema, P.H. Occurrence and distribution of glyphosate-resistant giant ragweed (Ambrosia trifida L.) in southwestern Ontario. Can. J. Plant Sci. 2012, 92, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Westhoven, A.M.; Davis, V.M.; Gibson, K.D.; Weller, S.C.; Johnson, W.G. Field presence of glyphosate-resistant horseweed (Conyza canadensis), common lambsquarters (Chenopodium album), and giant ragweed (Ambrosia trifida) biotypes with elevated tolerance to glyphosate. Weed Technol. 2008, 22, 544–548. [Google Scholar] [CrossRef]
- Moretti, M.L.; Van Horn, C.R.; Robertson, R.R.; Segobye, K.; Weller, S.C.; Young, B.G.; Johnson, W.G.; Sammons, R.D.; Wang, D.; Ge, X.; et al. Glyphosate resistance in Ambrosia trifida. Part 2. Rapid response physiology and non-target site resistance. Pest Manag. Sci. 2018, 74, 1079–1088. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Jha, P.; Steckel, L.E.; Scott, R.C. Confirmation and control of glyphosate-resistant giant ragweed (Ambrosia trifida) in Tennessee. Weed Technol. 2010, 24, 64–70. [Google Scholar] [CrossRef]
- Ballard, T.O.; Foley, M.E.; Bauman, T.T. Germination, viability, and protein changes during cold stratification of giant ragweed (Ambrosia trifida L.) seed. J. Plant Physiol. 1996, 149, 229–232. [Google Scholar] [CrossRef]
- Vange, V.; Heuch, I.; Vandvik, V. Do seed mass and family affect germination and juvenile performance in Knautia arvensis? A study using failure-time methods. Acta Oecol. 2004, 25, 169–178. [Google Scholar] [CrossRef]
- Burgos, N.R.; Tranel, P.J.; Streibig, J.C.; Davis, V.M.; Shaner, D.; Norsworthy, J.K.; Ritz, C. Review: Confirmation of resistance to herbicides and evaluation of resistance levels. Weed Sci. 2013, 61, 4–20. [Google Scholar] [CrossRef]
- Schutte, B.J.; Regnier, E.E.; Harrison, S.K. Seed dormancy and adaptive seedling emergence timing in giant ragweed (Ambrosia trifida). Weed Sci. 2012, 60, 19–26. [Google Scholar] [CrossRef]
- Fenner, M. The effects of the parent environment on seed germinability. Seed Sci. Res. 1991, 1, 75–84. [Google Scholar] [CrossRef]
- Šoštarčić, V.; Masin, R.; Loddo, D.; Brijačak, E.; Šćepanović, M. Germination Parameters of Selected Summer Weeds: Transferring of the AlertInf Model to Other Geographical Regions. Agronomy 2021, 11, 292. [Google Scholar] [CrossRef]
- Dahlquist, R.; Prather, T.; Stapleton, J. Time and temperature requirements for weed seed thermal death. Weed Sci. 2007, 55, 619–625. [Google Scholar] [CrossRef]
- Guillemin, J.P.; Chauvel, B. Effects of the seed weight and burial depth on the seed behavior of common ragweed (Ambrosia artemisiifolia). Weed Biol. Manag. 2011, 11, 217–223. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Df | SumSq | MeanSq | F-Value | Sig. | ||
|---|---|---|---|---|---|---|---|
| Seed storage | S | 1 | 784 | 784.00 | 10.220 | ** | |
| T | 3 | 6334 | 2111.33 | 27.522 | *** | ||
| S × T | 3 | 6650 | 2216.67 | 28.895 | *** | ||
| Error | 56 | 4296 | 76.71 | ||||
| Stratification | S | 1 | 14,292 | 14,291.5 | 88.6847 | *** | |
| T | 5 | 55,441 | 11,088.3 | 68.8073 | *** | ||
| S × T | 5 | 1969 | 393.8 | 2.4435 | * | ||
| Error | 83 | 13,375 | 161.1 | ||||
| Chemical treatment | Direct addition | S | 1 | 680 | 679.7 | 7.5818 | ** |
| T | 12 | 125,307 | 10,442.2 | 116.4800 | *** | ||
| S × T | 12 | 17,078 | 1423.2 | 15.8753 | *** | ||
| Error | 182 | 16,316 | 89.6 | ||||
| Conditioning | S | 1 | 6062 | 6062.3 | 47.6656 | *** | |
| T | 6 | 67,856 | 11,309.3 | 88.9209 | *** | ||
| S × T | 6 | 3613 | 602.6 | 4.7382 | *** | ||
| Error | 98 | 12,464 | 127.2 | ||||
| Variables | Df | SumSq | MeanSq | F-Value | Sig. | |
|---|---|---|---|---|---|---|
| Emergence (%) | S | 1 | 1464 | 1464.10 | 5.2093 | * |
| T | 4 | 6808 | 1702.08 | 6.0561 | *** | |
| S × T | 4 | 4359 | 1089.80 | 3.8776 | ** | |
| Error | 150 | 42,158 | 281.05 | |||
| Hypocotyl length (cm) | S | 1 | 1.3504 | 1.35038 | 8.7901 | ** |
| T | 4 | 1.0876 | 0.27190 | 1.7699 | ns | |
| S × T | 4 | 1.3802 | 0.34505 | 2.2460 | ns | |
| Error | 149 | 22.8902 | 0.15363 | |||
| Fresh biomass (g) | S | 1 | 3.721 | 3.7210 | 5.7545 | * |
| T | 4 | 13.298 | 3.3246 | 5.1414 | *** | |
| S × T | 4 | 7.320 | 1.8301 | 2.8302 | * | |
| Error | 150 | 96.994 | 0.6466 |
| Treatment | Concentration (mM) | Method | ||
|---|---|---|---|---|
| Imbibition (24 h) | Direct Addition | Conditioning (48 h) | ||
| Distilled water d | - | + | + | + |
| Ethephon a,d | 1 | - | + | + |
| Gibberellic acid (GA3) b,d | 1 | - | + | + |
| Thiourea c,d | 263 | + | + | + |
| KNO3 | 198 | + | + | - |
| Ethephon + GA3 d | 1 + 1 | - | + | + |
| GA3 + thiourea | 1 + 263 | - | + | - |
| Ethephon + thiourea | 1 + 263 | - | + | - |
| Ethephon + KNO3 | 1 + 198 | - | + | - |
| GA3 + KNO3 | 1 + 198 | - | + | - |
| Thiourea + KNO3 | 263 + 198 | - | + | - |
| Ethephon + GA3 + thiourea | 1 + 1 + 263 | - | + | - |
| GA3 + thiourea + ethephon + KNO3 | 1 + 263 + 1 + 198 | - | + | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šćepanović, M.; Košćak, L.; Pismarović, L.; Šoštarčić, V. Stimulation of Germination of Freshly Collected and Cold-Stored Seeds of Ambrosia artemisiifolia L. Plants 2022, 11, 1888. https://doi.org/10.3390/plants11141888
Šćepanović M, Košćak L, Pismarović L, Šoštarčić V. Stimulation of Germination of Freshly Collected and Cold-Stored Seeds of Ambrosia artemisiifolia L. Plants. 2022; 11(14):1888. https://doi.org/10.3390/plants11141888
Chicago/Turabian StyleŠćepanović, Maja, Laura Košćak, Laura Pismarović, and Valentina Šoštarčić. 2022. "Stimulation of Germination of Freshly Collected and Cold-Stored Seeds of Ambrosia artemisiifolia L." Plants 11, no. 14: 1888. https://doi.org/10.3390/plants11141888