
Impacts of Short-Term Grazing Intensity on the Plant Diversity and Ecosystem Function of Alpine Steppe on the Qinghai–Tibetan Plateau

 ,
,
Abstract
:1. Introduction
2. Results
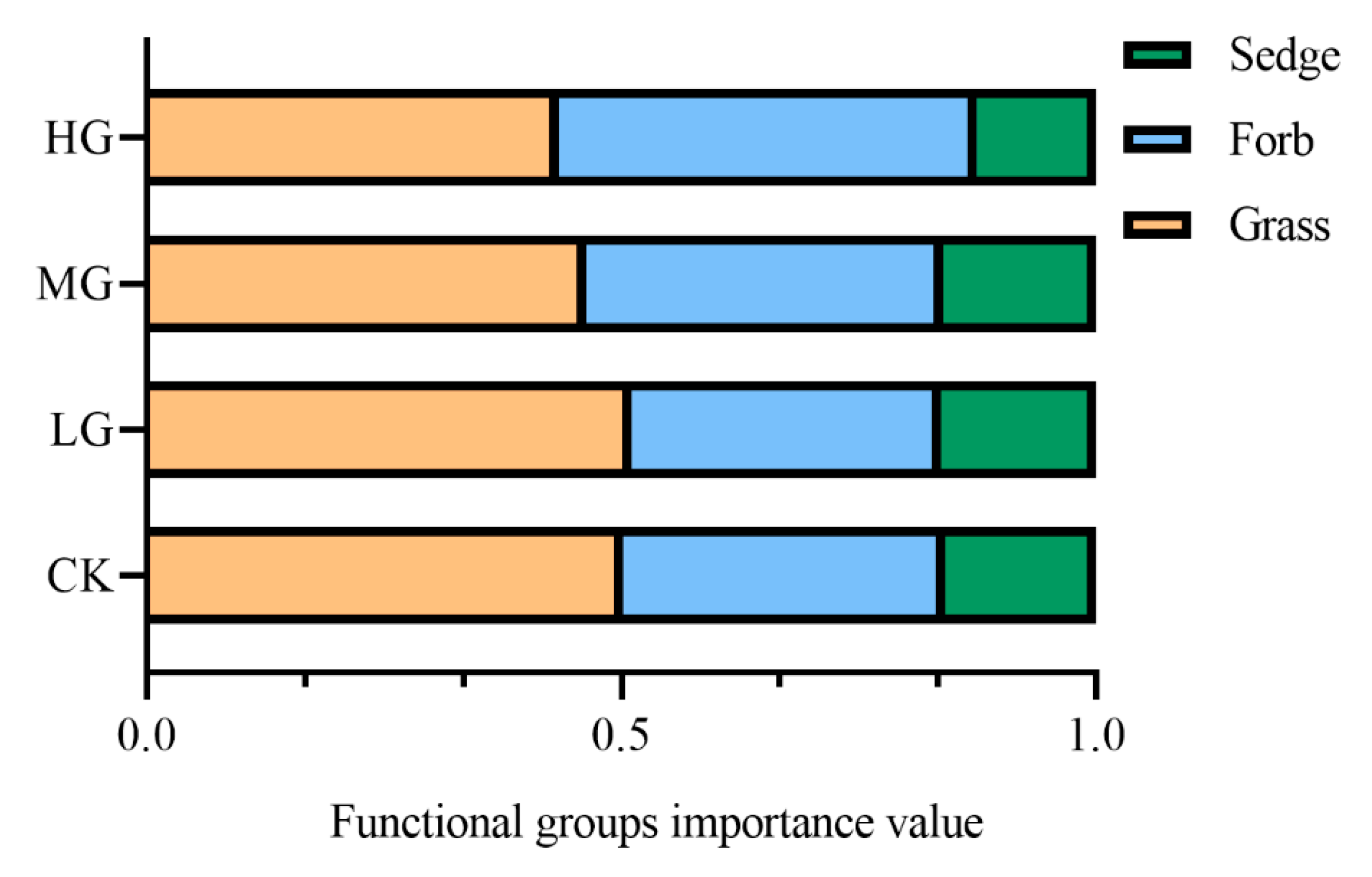
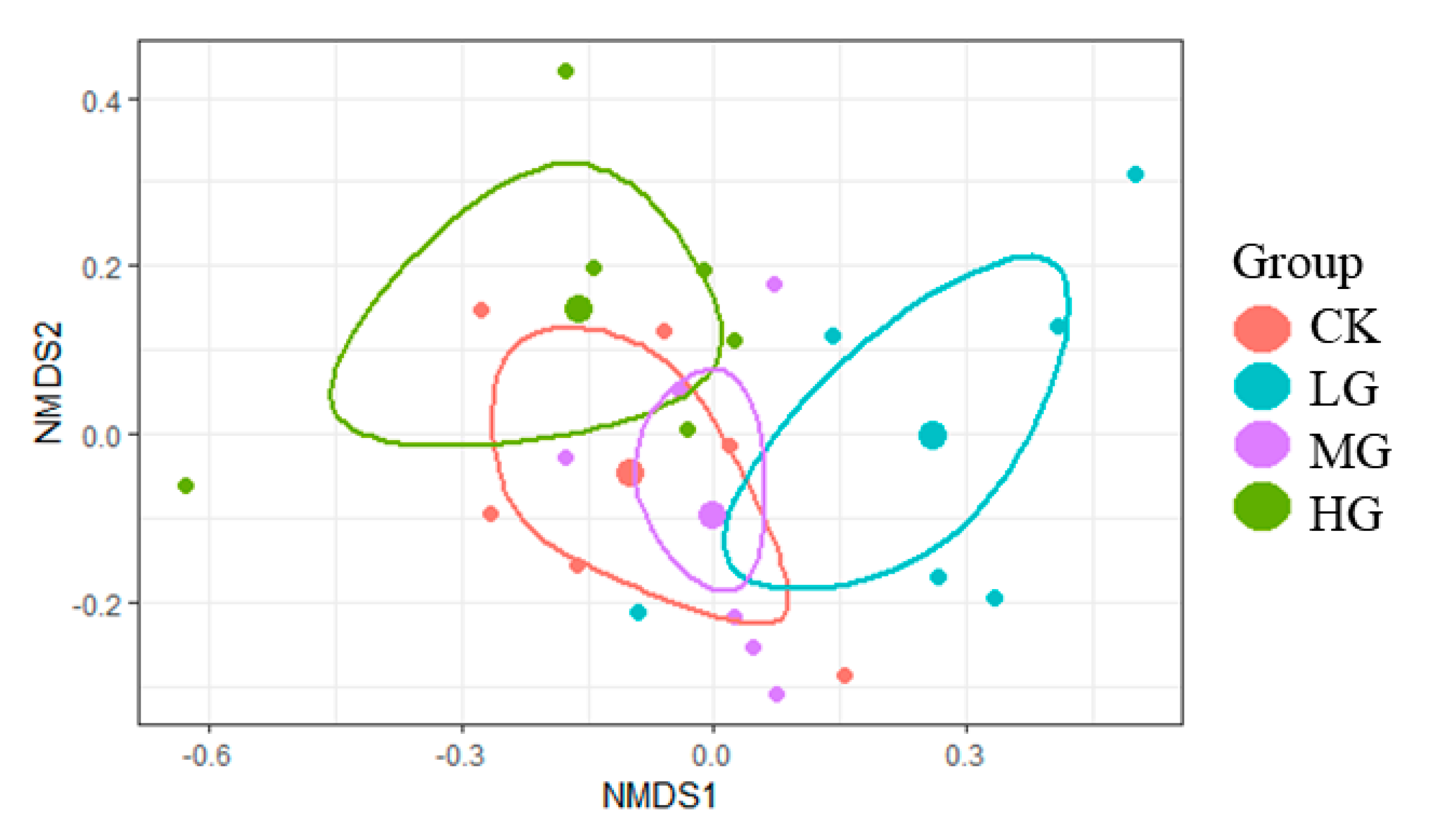
2.1. Effects of Grazing Intensity on Plant Species Composition of Alpine Steppe
2.2. Effects of Grazing Intensity on Plant Species Diversity of Alpine Steppe
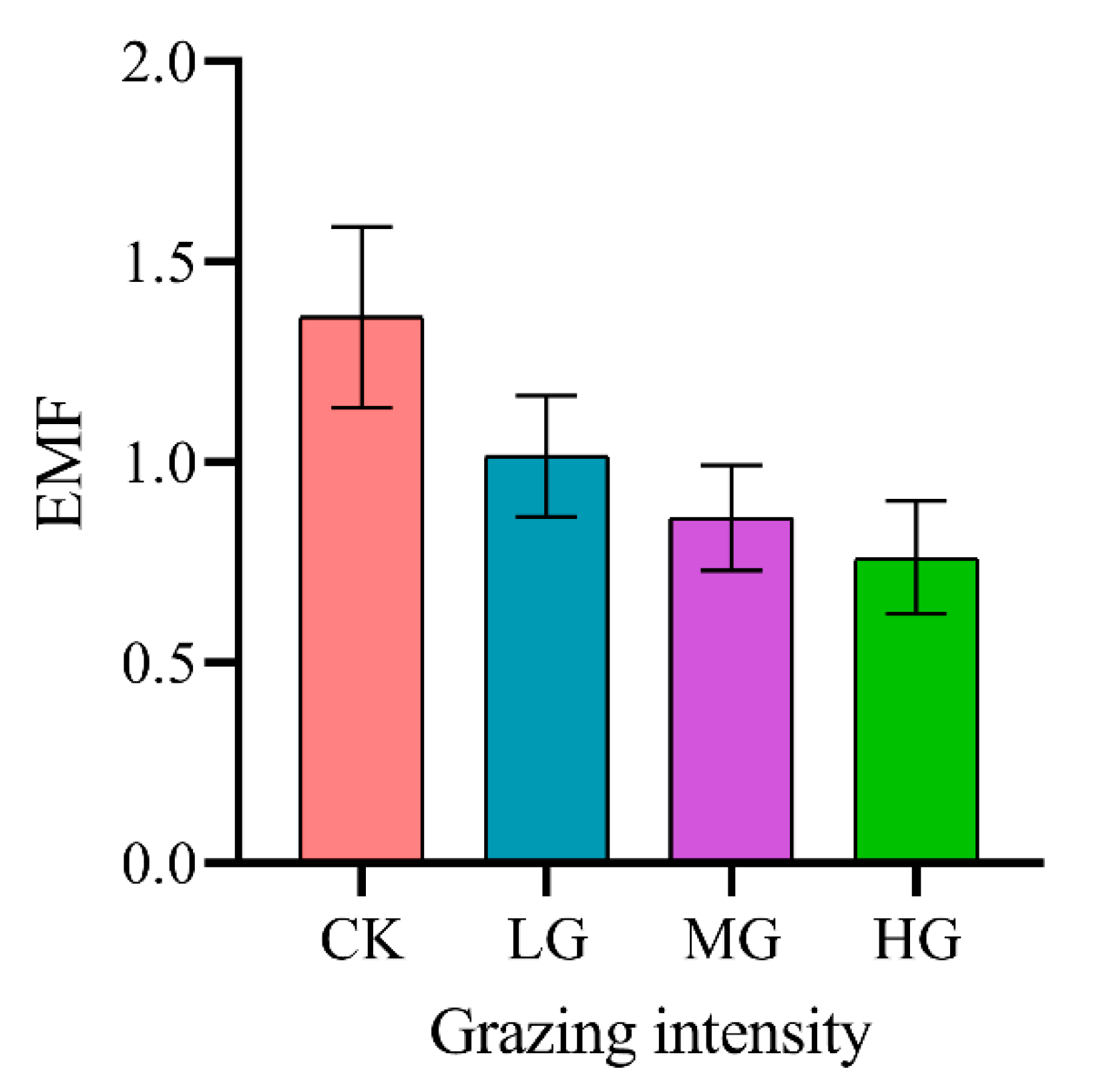
2.3. Effects of Grazing Intensity on Ecosystem Function Indicators and EMF
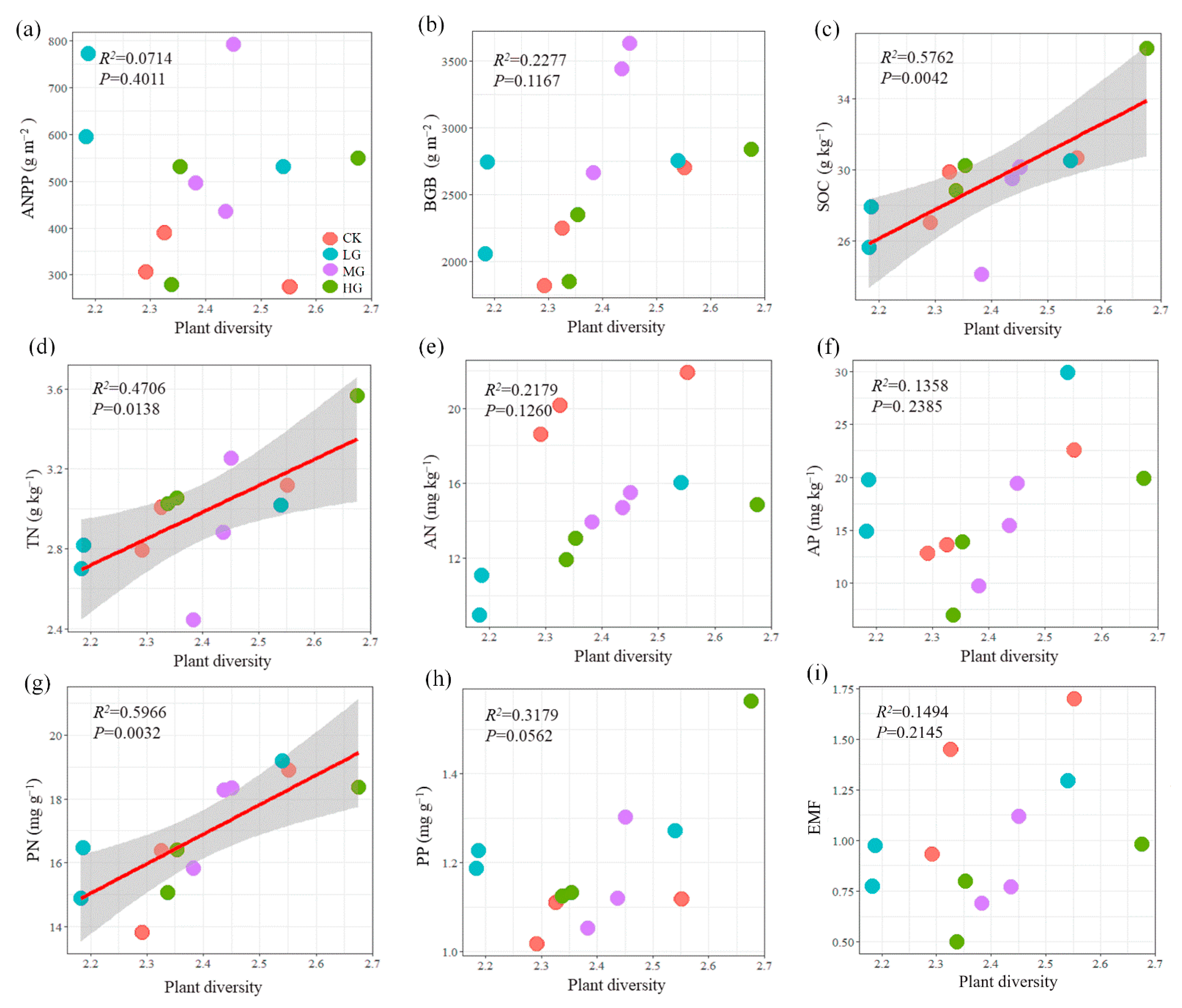
2.4. Relationships between Plant Diversity and Ecosystem Function Indicators and EMF under Grazing
3. Discussion
3.1. Effect of Grazing Intensity on Community Characteristics of Alpine Steppe
3.2. Effect of Grazing Intensity on Ecosystem Functions
3.3. Relationships between Plant Diversity and Ecosystem Functions
4. Materials and Methods
4.1. Study Sites
4.2. Experimental Design
4.3. Vegetation and Soil Sampling
4.4. Statistical Analysis
4.4.1. Species Importance Value and Species Diversity Index
4.4.2. Assessing Ecosystem Multifunctionality
4.4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, S.K.; Sherman, R. Enhancing the resilience of coupled human and natural systems of alpine rangelands on the Qinghai-Tibetan Plateau. Rangel. J. 2015, 37, i–iii. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.K.; Shang, Z.H.; Gao, J.X.; Boone, R. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2020, 287, 106684. [Google Scholar] [CrossRef]
- Dong, S.K.; Shang, Z.H.; Gao, J.X.; Boone, R. Enhancing the ecological services of the Qinghai-Tibetan Plateau’s grasslands through sustainable restoration and management in era of global change. Agric. Ecosyst. Environ. 2022, 326, 107756. [Google Scholar] [CrossRef]
- Jing, X.; He, J.S. Relationship between biodiversity, ecosystem multifunctionality and multiserviceability: Literature overview and research advances. Chin. J. Plant Ecol. 2021, 45, 1094–1111. [Google Scholar] [CrossRef]
- Li, Y.Y.; Dong, S.K.; Wen, L.; Wang, X.X.; Wu, Y. The effects of fencing on carbon stocks in the degraded alpine grasslands of the Qinghai-Tibetan Plateau. J. Environ. Manag. 2013, 128, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; Li, B.; Xu, X.C.; Zhang, L.X. Response of Vegetation and Soil Characteristics to Grazing Disturbance in Mountain Meadows and Temperate Typical Steppe in the Arid Regions of Central Asian, Xinjiang. Int. J. Environ. Res. Public Health 2020, 17, 4572. [Google Scholar] [CrossRef]
- Deng, L.; Sweeney, S.; Shangguan, Z.P. Grassland responses to grazing disturbance: Plant diversity changes with grazing intensity in a desert steppe. Grass Forage Sci. 2014, 69, 524–533. [Google Scholar] [CrossRef]
- Fetzel, T.; Havlik, P.; Herrero, M.; Kaplan, J.O.; Kastner, T.; Kroisleitner, C.; Rolinski, S.; Searchinger, T.; Van Bodegom, P.M.; Wirsenius, S.; et al. Quantification of uncertainties in global grazing systems assessment. Glob. Biogeochem. Cycles 2017, 31, 1089–1102. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Hou, X.; Schellenberg, M.P.; Qin, Y.; Yun, X.; Wei, Z.; Jiang, C.; Wang, Y. Different responses of plant species to deferment of sheep grazing in a desert steppe of Inner Mongolia, China. Rangel. J. 2014, 36, 583–592. [Google Scholar] [CrossRef]
- Shen, H.; Dong, S.K.; Di Tommaso, A.; Xiao, J.N.; Zhi, Y.L. N deposition may accelerate grassland degradation succession from grasses- and sedges-dominated into forbs-dominated in overgrazed alpine grassland systems on Qinghai-Tibetan Plateau. Ecol. Indic. 2021, 129, 107898. [Google Scholar] [CrossRef]
- Zhan, T.Y.; Zhang, Z.C.; Sun, J.; Liu, M.; Zhang, X.B.; Peng, F.; Tsunekawa, A.; Zhou, H.K.; Gou, X.W.; Fu, S. Meta-analysis demonstrating that moderate grazing can improve the soil quality across China’s grassland ecosystems. Appl. Soil Ecol. 2020, 147, 103438. [Google Scholar] [CrossRef]
- Zhang, Y.; Ganjurjav, H.; Dong, S.K.; Gao, Q.Z. Excessive plant compensatory growth: A potential endogenous driver of meadow degradation on the Qinghai-Tibetan Plateau. Ecosyst. Health Sustain. 2020, 6, 1816500. [Google Scholar] [CrossRef]
- Dong, S.K.; Li, Y.; Ganjurjav, H.; Gao, Q.Z.; Gao, X.X.; Zhang, J.; Yan, Y.L.; Zhang, Y.; Liu, S.L.; Hu, G.Z.; et al. Grazing promoted soil microbial functional genes for regulating C and N cycling in alpine meadow of the Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2020, 303, 107111. [Google Scholar] [CrossRef]
- Li, W.; Cao, W.X.; Wang, J.L.; Li, X.L.; Xu, C.L.; Shi, S.L. Effects of grazing regime on vegetation structure, productivity, soil quality, carbon and nitrogen storage of alpine meadow on the Qinghai-Tibetan Plateau. Ecol. Eng. 2017, 98, 123–133. [Google Scholar] [CrossRef]
- Zhang, M.; Li, X.B.; Wang, H.; Huang, Q. Comprehensive analysis of grazing intensity impacts soil organic carbon: A case study in typical steppe of Inner Mongolia, China. Appl. Soil Ecol. 2018, 129, 1–12. [Google Scholar] [CrossRef]
- Chang, Q.; Wang, L.; Ding, S.W.; Xu, T.T.; Li, Z.Q.; Song, X.X.; Zhao, X.; Wang, D.L.; Pan, D.F. Grazer effects on soil carbon storage vary by herbivore assemblage in a semi-arid grassland. J. Appl. Ecol. 2018, 55, 2517–2526. [Google Scholar] [CrossRef]
- Zhou, G.Y.; Zhou, X.H.; He, Y.H.; Shao, J.J.; Hu, Z.H.; Liu, R.Q.; Zhou, H.M.; Hosseinibai, S. Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: A meta-analysis. Glob. Chang. Biol. 2017, 23, 1167–1179. [Google Scholar] [CrossRef]
- Liang, Y.; Li, Y.; Wang, L.; Zhang, X.D.; Wang, J.Z.; Wu, H.D.; Yan, Z.Q.; Zhang, K.R.; Kang, X.M. Grazing significantly increases root shoot ratio but decreases soil organic carbon in Qinghai-Tibetan Plateau grasslands: A hierarchical meta-analysis. Land Degrad. Dev. 2020, 31, 2369–2378. [Google Scholar] [CrossRef]
- Dao, R.N.; Song, Y.T.; Wu, Y.N.; Huo, G.W.; Wang, X.M.; Xu, Z.C. Response of plant leaf traits to grazing intensity in Stipa krylovii steppe. Chin. J. Appl. Ecol. 2016, 27, 2231–2238. [Google Scholar] [CrossRef]
- Chai, J.L.; Yu, X.J.; Xu, C.L.; Xiao, H.; Zhang, J.W.; Yang, H.L.; Pan, T.T. Effects of yak and Tibetan sheep trampling on soil properties in the northeastern Qinghai-Tibetan Plateau. Appl. Soil Ecol. 2019, 144, 147–154. [Google Scholar] [CrossRef]
- Ganjurjav, H.; Duan, M.J.; Wan, Y.F.; Zhang, W.N.; Gao, Q.Z.; Li, Y.; Jiang, C.; Wang, Z.; Danjiu, L.B.; Guo, H.B. Effects of grazing by large herbivores on plant diversity and productivity of semi-arid alpine steppe on the Qinghai-Tibetan Plateau. Rangel. J. 2015, 37, 389–397. [Google Scholar] [CrossRef]
- Han, G.D.; Hao, X.Y.; Zhao, M.L.; Wang, M.J.; Ellert, B.H.; Walter, W.; Wang, M.J. Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia. Agric. Ecosyst. Environ. 2008, 125, 21–32. [Google Scholar] [CrossRef]
- Maestre, F.T.; Castillo-Monroy, A.P.; Bowker, M.A.; Ochoa-Hueso, R. Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. J. Ecol. 2012, 100, 317–330. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Wang, Z.W.; Niu, S.L.; Tian, D.S.; Wu, Q.; Gao, X.F.; Schellenberg, M.P.; Han, G.D. Diversity of plant and soil microbes mediates the response of ecosystem multifunctionality to grazing disturbance. Sci. Total Environ. 2021, 776, 145730. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, F.Y.; Wang, Y.; Liu, X.; Cheng, J.; Zhang, J.; Baoyin, T.; Bardgett, R.D. High ecosystem multifunctionality under moderate grazing is associated with high plant but low bacterial diversity in a semi-arid steppe grassland. Plant Soil 2020, 448, 265–276. [Google Scholar] [CrossRef]
- Yang, W.S.; Liu, Y.; Zhao, J.X.; Chang, X.F.; Wiesmeier, M.; Sun, J.; Lopez-Vicente, M.; Garcia-Ruiz, R.; Gomez, J.A.; Zhou, H.K.; et al. SOC changes were more sensitive in alpine grasslands than in temperate grasslands during grassland transformation in China: A meta-analysis. J. Clean. Prod. 2021, 308, 127430. [Google Scholar] [CrossRef]
- Tang, L.; Dong, S.K.; Sherman, R.; Liu, S.L.; Liu, Q.R.; Wang, X.X.; Su, X.K.; Zhang, Y.; Li, Y.Y.; Wu, Y.; et al. Changes in vegetation composition and plant diversity with rangeland degradation in the alpine region of Qinghai-Tibet Plateau. Rangel. J. 2015, 37, 107–115. [Google Scholar] [CrossRef]
- Yang, Z.A.; Xiong, W.; Xu, Y.Y.; Jiang, L.; Zhu, E.X.; Zhan, W.; He, Y.X.; Zhu, D.; Zhu, Q.A.; Peng, C.H.; et al. Soil properties and species composition under different grazing intensity in an alpine meadow on the eastern Tibetan Plateau, China. Environ. Monit. Assess. 2016, 188, 678. [Google Scholar] [CrossRef]
- Li, G.; Zhang, Z.; Shi, L.L.; Zhou, Y.; Yang, M.; Cao, J.X.; Wu, S.H.; Lei, G.C. Effects of Different Grazing Intensities on Soil C, N, and P in an Alpine Meadow on the QinghaiTibetan Plateau, China. Int. J. Environ. Res. Public Health 2018, 15, 2584. [Google Scholar] [CrossRef] [Green Version]
- Eldridge, D.J.; Poore, A.G.B.; Ruiz-Colmenero, M.; Letnic, M.; Soliveres, S. Ecosystem structure, function, and composition in rangelands are negatively affected by livestock grazing. Ecol. Appl. 2016, 26, 1273–1283. [Google Scholar] [CrossRef] [Green Version]
- Li, W.H.; Xu, F.W.; Zheng, S.X.; Taube, F.; Bai, Y.F. Patterns and thresholds of grazing-induced changes in community structure and ecosystem functioning: Species-level responses and the critical role of species traits. J. Appl. Ecol. 2017, 54, 963–975. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Y.J.; Xu, M.J.; Zhu, J.T.; Wimberly, M.C.; Yu, G.R.; Niu, S.L.; Xi, Y.; Zhang, X.Z.; Wang, J.S. Light-intensity grazing improves alpine meadow productivity and adaption to climate change on the Tibetan Plateau. Sci. Rep. 2015, 5, 15949. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.Y.; Wang, Z.W.; Han, G.D.; Schellenberg, M.P.; Wu, Q.; Gu, C. Grazing induced changes in plant diversity is a critical factor controlling grassland productivity in the Desert Steppe, Northern China. Agric. Ecosyst. Environ. 2018, 265, 73–83. [Google Scholar] [CrossRef]
- Herrero-Jáuregui, C.; Oesterheld, M. Effects of grazing intensity on plant richness and diversity: A meta-analysis. Oikos 2018, 127, 757–766. [Google Scholar] [CrossRef]
- Liu, J.; Feng, C.; Wang, D.L.; Wang, L.; Wilsey, B.J.; Zhong, Z.W. Impacts of grazing by different large herbivores in grassland depend on plant species diversity. J. Appl. Ecol. 2015, 52, 1053–1062. [Google Scholar] [CrossRef]
- Wang, C.; Tang, Y.J. A global meta-analyses of the response of multi-taxa diversity to grazing intensity in grasslands. Environ. Res. Lett. 2019, 14, 114003. [Google Scholar] [CrossRef] [Green Version]
- Waters, C.M.; Orgill, S.E.; Melville, G.J.; Toole, I.D.; Smith, W.J. Management of Grazing Intensity in the Semi-Arid Rangelands of Southern Australia: Effects on Soil and Biodiversity. Land Degrad. Dev. 2016, 28, 1363–1375. [Google Scholar] [CrossRef]
- Wu, G.L.; Du, G.Z.; Liu, Z.H.; Thirgood, S. Effect of fencing and grazing on a Kobresia-dominated. Plant Soil 2009, 319, 115–126. [Google Scholar] [CrossRef]
- Luo, F.L.; Zhang, F.W.; Zhang, G.R.; Wang, C.Y.; Zhu, J.B.; Yang, Y.S.; Li, Y.N. Effects of grazing intensity on community characteristics and vegetation living states in alpine meadows. Pratacult. Sci. 2021, 38, 2097–2105. [Google Scholar] [CrossRef]
- Song, J.; Wang, F.G.; Wen, L.; Wang, L.X.; Li, J.L.; Wu, S.N.; Xu, Z.C. Effects of grazing on plant diverity and soil nutrients in typical temperate steppe. Acat Agrestia Sin. 2019, 27, 1694–1701. [Google Scholar] [CrossRef]
- Zhu, J.B.; Li, H.Q.; He, H.D. Analysis of the variation characteristics and controlling factors of soil rapidly available nitrogen in Haibei alpine meadow under grazing pressure gradient. J. Glaciol. Geocryol. 2015, 37, 1660–1669. [Google Scholar] [CrossRef]
- Liu, Y.Z.; Liu, W.T.; Yang, X.X.; Li, C.D.; Feng, B.; Yu, Y.; Zhang, C.P.; Dong, Q.M. Effects of grazing on the C:N:P stoichiometry in global grassland ecosystems: A Meta analysis. Chin. J. Appl. Ecol. 2022, 33, 1251–1259. [Google Scholar] [CrossRef]
- Knops, J.M.H.; Bradley, K.L.; Wedin, D.A. Mechanisms of plant species impacts on ecosystem nitrogen cycling. Ecol. Lett. 2010, 5, 454–466. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.P.; Zhang, W.P.; Yu, Y.C.; Yu, S.B.; Lambers, H.; Li, L. Linking shifts in species composition induced by grazing with root traits for phosphorus acquisition in a typical steppe in Inner Mongolia. Sci. Total Environ. 2020, 712, 136495. [Google Scholar] [CrossRef] [PubMed]
- Loeser, M.R.; Crews, T.E.; Sisk, T.D. Defoliation increased above-ground productivity in a semi-arid grassland. J. Range Manag. 2004, 57, 442–447. [Google Scholar] [CrossRef]
- Sun, J.; Zhan, T.; Liu, M.; Zhang, Z.; Wang, Y.; Liu, S.; Wu, G.L.; Liu, G.; Tsunekawa, A. Verification of the biomass transfer hypothesis under moderate grazing across the Tibetan plateau: A meta-analysis. Plant Soil 2019, 458, 139–150. [Google Scholar] [CrossRef]
- Wang, L.; Delgado-Baquerizo, M.; Wang, D.L.; Isbell, F.; Liu, J.; Feng, C.; Liu, J.S.; Zhong, Z.W.; Zhu, H.; Yuan, X.; et al. Diversifying livestock promotes multidiversity and multifunctionality in managed grasslands. Proc. Natl. Acad. Sci. USA 2019, 116, 6187–6192. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gao, Q.Z.; Dong, S.K.; Liu, S.L.; Wang, X.X.; Su, X.K.; Li, Y.Y.; Tang, L.; Wu, X.Y.; Zhao, H.D. Effects of grazing and climate warming on plant diversity, productivity and living state in the alpine rangelands and cultivated grasslands of the Qinghai-Tibetan Plateau. Rangel. J. 2015, 37, 57–65. [Google Scholar] [CrossRef]
- Dong, Q.M.; Zhao, X.Q.; Ma, Y.S. Effects of Grazing Intensity on Soil Nutrition in the Alpine Mixed- sown Grassland. Chin. J. Soil Sci. 2008, 39, 553–557. [Google Scholar] [CrossRef]
- Katsalirou, E.; Deng, S.; Gerakis, A.; Nofziger, D.L. Long-term management effects on soil P, microbial biomass P, and phosphatase activities in prairie soils. Eur. J. Soil Biol. 2016, 76, 61–69. [Google Scholar] [CrossRef]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; van Ruijven, J.; et al. High plant diversity is needed to maintain ecosystem services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.P.; Wang, W.T.; Xu, W.T.; Wang, Y.; Wan, H.W.; Chen, D.M.; Tang, Z.Y.; Tang, X.L.; Zhou, G.Y.; Xie, Z.Q.; et al. Plant diversity enhances productivity and soil carbon storage. Proc. Natl. Acad. Sci. USA 2018, 115, 4027–4032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Ran, J.; Dong, L.; Du, Q.; Ji, M.; Yao, S.; Sun, Y.; Gong, C.; Hou, Q.; Gong, H.; et al. Aridity-driven shift in biodiversity-soil multifunctionality relationships. Nat. Commun. 2021, 12, 5350. [Google Scholar] [CrossRef] [PubMed]
- Maestre, F.T.; Quero, J.L.; Gotelli, N.J.; Escudero, A.; Ochoa, V.; Delgado-Baquerizo, M.; García-Gómez, M.; Bowker, M.A.; Soliveres, S.; Escolar, C.; et al. Plant Species Richness and Ecosystem Multifunctionality in Global Drylands. Science 2012, 335, 214–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Dong, S.K.; Sherman, R.; Liu, Q.; Liu, S.; Li, Y.; Wu, Y. A comparison of biodiversity–ecosystem function relationships in alpine grasslands across a degradation gradient on the Qinghai–Tibetan Plateau. Rangel. J. 2015, 37, 45–55. [Google Scholar] [CrossRef]
- Jochum, M.; Fischer, M.; Isbell, F.; Roscher, C.; Van der Plas, F.; Boch, S.; Boenisch, G.; Buchmann, N.; Catford, J.A.; Cavender-Bares, J.; et al. The results of biodiversity-ecosystem functioning experiments are realistic. Nat. Ecol. Evol. 2020, 4, 1485–1494. [Google Scholar] [CrossRef]
- Ren, J.Z. Research Method of Prataculture Science; China Agriculture Press: Beijing, China, 1998; pp. 200–210. [Google Scholar]
- Shannon, C.E. A mathematical theory of communications. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 168, 668. [Google Scholar] [CrossRef]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Pielou, E.C. Population and Community Ecology: Principles and Methods; Gordon and Breach: Philadelphia, PA, USA, 1974. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | CK | LG | MG | HG |
|---|---|---|---|---|---|
| Poaceae | Leymus secalinus (Georgi) Tzvelev. | 0.207 | 0.128 | 0.168 | 0.179 |
| Poa pratensis L. | 0.092 | 0.081 | 0.079 | 0.085 | |
| Agropyron cristatum (L.) Gaertn. | 0.049 | 0.067 | 0.048 | 0.048 | |
| koeleria cristata (Linn.) Pers. | 0.072 | 0.144 | 0.099 | 0.059 | |
| Stipa capillata L. | 0.083 | 0.088 | 0.068 | 0.054 | |
| Elymus nutans Griseb. | — | — | — | 0.008 | |
| Cyperaceae | Carex parvula O. Yano | 0.151 | 0.179 | 0.161 | 0.125 |
| Carex melanantha C. A. Mey. | 0.008 | — | — | — | |
| Fabaceae | Thermopsis lupinoides (L.) Link | 0.009 | — | — | 0.007 |
| Medicago ruthenica (L.) Trautv. | 0.002 | — | — | 0.005 | |
| Astragalus membranaceus (Fisch.) Bunge. | 0.008 | 0.010 | 0.041 | 0.019 | |
| Rosaceae | Potentilla bifurca L. | 0.032 | — | 0.008 | 0.028 |
| Sibbaldia adpressa Bge. | 0.011 | — | — | 0.006 | |
| Potentilla multifida L. | 0.022 | 0.017 | 0.039 | 0.089 | |
| Compositae | Aster tataricus L. f. | 0.077 | 0.117 | 0.081 | 0.086 |
| Taraxacum mongolicum Hand.-Mazz. | 0.031 | 0.005 | 0.008 | 0.010 | |
| Artemisia frigida Willd. | 0.020 | 0.021 | 0.007 | 0.011 | |
| Artemisia scoparia Waldst. et Kit. | 0.027 | 0.025 | 0.020 | 0.015 | |
| Iridaceae | Irisensata thumb. | 0.037 | 0.063 | 0.052 | 0.039 |
| Gentianaceae | Gentiana macrophylla Pall. | — | — | — | 0.005 |
| Comastoma pulmonarium (Turcz.) Toyok. | — | — | 0.003 | 0.002 | |
| Gentiana scabra Bunge. | 0.002 | 0.003 | 0.001 | 0.003 | |
| Scrophulariaceae | Lancea tibetica Hook. f. et Thoms. | 0.018 | 0.017 | 0.001 | 0.029 |
| Pedicularis qinghaiensis T. Yamaz. | — | 0.002 | 0.0145 | 0.002 | |
| Labiatae | Dracocephalum heterophyllum Benth. | 0.022 | 0.012 | 0.054 | 0.049 |
| Thymelaeaceae | Stellera chamaejasme L. | 0.005 | — | 0.011 | — |
| Umbelliferae | Bupleurum chinensis DC. | 0.012 | 0.018 | 0.026 | 0.008 |
| Plantaginaceae | Plantago asiatica L. | — | 0.004 | — | 0.012 |
| Ranunculaceae | Aconitum gymnandrum Maxim. | — | — | — | 0.008 |
| Caryophyllaceae | Silene firma Siebold & Zucc. | 0.002 | — | 0.009 | 0.004 |
| Orchidaceae | Herminium monorchis (Linn.) R. Br. | — | — | — | 0.001 |
| Grazing Intensity | Shannon-Wiener Index (H’) | Simpson Index (D) | Pielou Index (J) | Margalef Index (Dmg) |
|---|---|---|---|---|
| CK | 2.340 (0.082) a | 0.882 (0.014) a | 0.870 (0.022) a | 2.360 (0.136) a |
| LG | 2.303 (0.118) a | 0.880 (0.012) a | 0.861 (0.027) a | 2.271 (0.163) a |
| MG | 2.423 (0.021) a | 0.886 (0.004) a | 0.889 (0.019) a | 2.382 (0.096) a |
| HG | 2.456 (0.110) a | 0.892 (0.016) a | 0.890 (0.017) a | 2.312 (0.254) a |
| Grazing Intensity | Ecosystem Functions Indicators | |||||||
|---|---|---|---|---|---|---|---|---|
| ANPP (g m−2) | BGB (g m−2) | SOC (g kg−1) | TN (g kg−1) | AP (mg kg−1) | AN (mg kg−1) | PN (mg g−1) | PP (mg g−1) | |
| CK | 323.63 (34.31) a | 1185.08 (181.18) a | 29.19 (1.12) a | 2.98 (0.10) a | 16.34 (3.12) a | 20.26 (0.95) a | 18.80 (0.52) a | 1.25 (0.07) ab |
| LG | 633.07 (72.09) a | 1111.56 (178.41) a | 28.01 (1.42) a | 2.85 (0.09) a | 21.56 (4.42) a | 12.03 (2.09) b | 19.96 (0.66) a | 1.37 (0.07) a |
| MG | 574.93 (110.17) a | 1228.22 (189.37) a | 27.92 (1.92) a | 2.86 (0.23) a | 14.88 (2.80) a | 14.73 (0.45) ab | 17.05 (0.88) ab | 1.15 (0.03) ab |
| HG | 453.59 (87.34) a | 829.26 (150.06) a | 31.97 (2.47) a | 3.22 (0.18) a | 13.58 (3.72) a | 13.29 (0.86) b | 15.02 (0.64) b | 1.07 (0.02) b |
| Ecosystem Functional Indicators | Importance |
|---|---|
| Aboveground net primary productivity | Primary production function, a key ecosystem process that supports ecosystem belowground functionality. |
| Belowground biomass | |
| Soil organic carbon | Soil carbon sequestration function, and build-up of nutrient pools for plants and microorganisms. |
| Soil total nitrogen | |
| Soil available phosphorus | Nutrient cycling function. Soil available phosphorus and soil available nitrogen are important nutrients sources for both microorganisms and plants. |
| Soil available nitrogen | |
| Plant nitrogen content | Sustain human welfare, plant nitrogen and phosphorus content embody the nutrient utilization of plants, and involve the chemical cycling of nutrients in ecosystems. |
| Plant phosphorus content |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, X.; Yang, J.; Dong, S.; Shen, H.; He, F.; Zhi, Y.; Kwaku, E.A.; Tu, D.; Dou, S.; Zhou, X.; et al. Impacts of Short-Term Grazing Intensity on the Plant Diversity and Ecosystem Function of Alpine Steppe on the Qinghai–Tibetan Plateau. Plants 2022, 11, 1889. https://doi.org/10.3390/plants11141889
Hao X, Yang J, Dong S, Shen H, He F, Zhi Y, Kwaku EA, Tu D, Dou S, Zhou X, et al. Impacts of Short-Term Grazing Intensity on the Plant Diversity and Ecosystem Function of Alpine Steppe on the Qinghai–Tibetan Plateau. Plants. 2022; 11(14):1889. https://doi.org/10.3390/plants11141889
Chicago/Turabian StyleHao, Xinghai, Juejie Yang, Shikui Dong, Hao Shen, Fengcai He, Yangliu Zhi, Emmanuella A. Kwaku, Danjia Tu, Shengyun Dou, Xueli Zhou, and et al. 2022. "Impacts of Short-Term Grazing Intensity on the Plant Diversity and Ecosystem Function of Alpine Steppe on the Qinghai–Tibetan Plateau" Plants 11, no. 14: 1889. https://doi.org/10.3390/plants11141889