
Biodiversity and Ecosystem Function under Simulated Gradient Warming and Grazing


Abstract
:1. Introduction
2. Results
2.1. Effect of Simulated Gradient Warming and Grazing on Environmental Factors
2.2. Effects of Simulated Gradient Warming and Grazing on Ecosystem Structure and Function
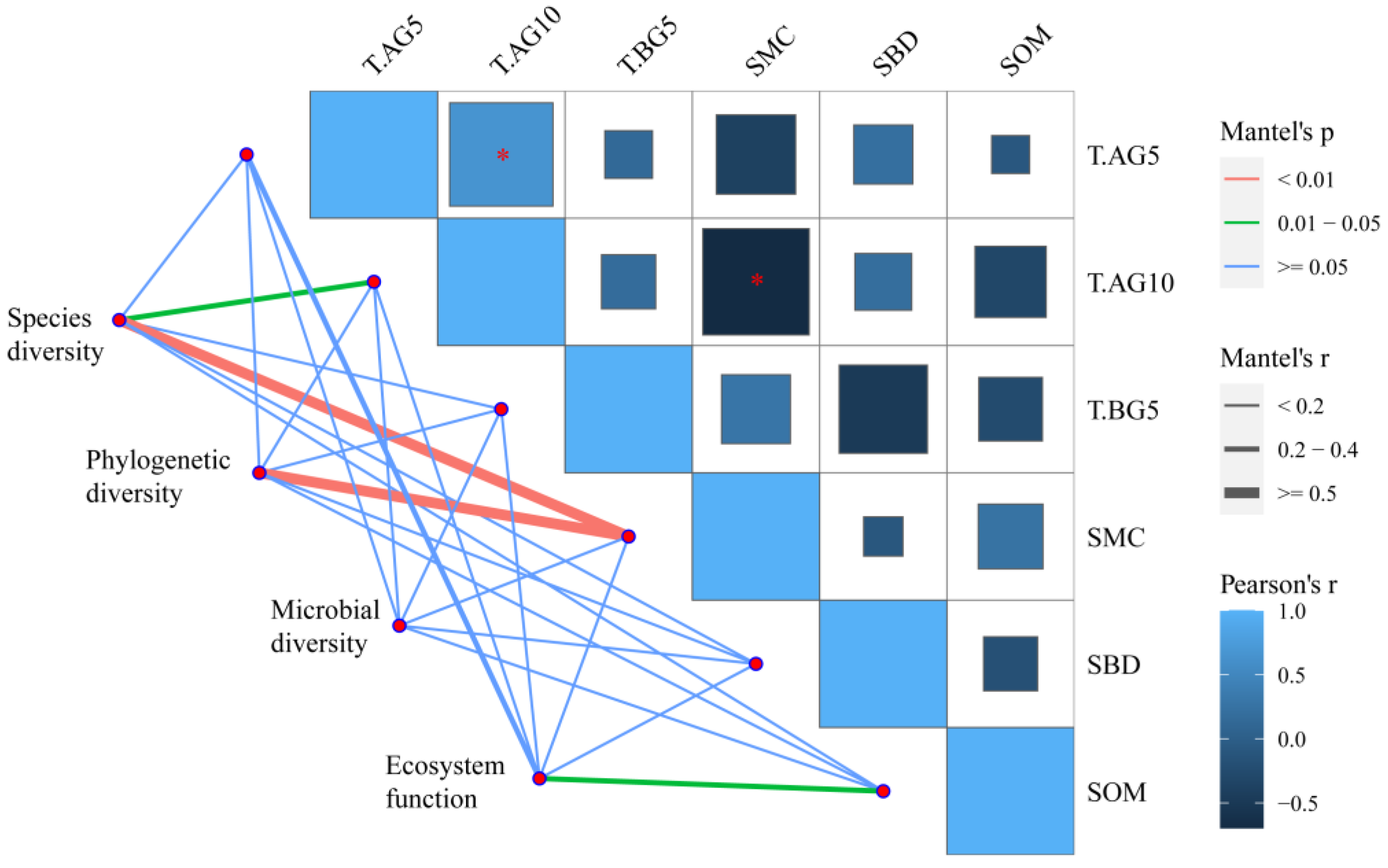
2.3. The Relationship between Biodiversity and Ecosystem Function
3. Discussion
4. Materials and Methods
4.1. Study Site
4.2. Experimental Design and Treatments
4.3. Sample Collection and Soil Physicochemical Property Analysis
4.4. Soil Microbial DNA Sequencing
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.A.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Ma, Z.Y.; Jing, X.; He, J.S. Biodiversity and ecosystem multifunctionality: Advances and perspectives. Biodivers. Sci. 2016, 24, 55–71. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Palacios, P.; Gross, N.; Gaitán, J.J.; Maestre, F.T. Climate mediates the biodiversity-ecosystem stability relationship globally. Proc. Natl. Acad. Sci. USA 2018, 15, 8400–8405. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Reich, P.B.; Knops, J.M.; Wedin, D.A.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [Green Version]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Caldeira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant diversity and productivity experiments in european grasslands. Science 1999, 286, 1123–1127. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Knops, J.; Tilman, D.; Craine, J.M.; Ellsworth, D.; Tjoelker, M.G.; Lee, T.D.; Wedin, D.A.; Naeem, S.; Bahauddin, D.; et al. Plant diversity enhances ecosystem responses to elevated CO2 and nitrogen deposition. Nature 2001, 410, 809–812. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.; Nakashizuka, T.; Raffaelli, D.G.; Schmid, B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Matulich, K.L.; Hooper, D.U.; Byrnes, J.E.; Duffy, J.; Gamfeldt, L.; Balvanera, P.; O’Connor, M.I.; Gonzalez, A. The functional role of producer diversity in ecosystems. Am. J. Bot. 2011, 98, 572–592. [Google Scholar] [CrossRef] [Green Version]
- Pennekamp, F.; Pontarp, M.; Tabi, A.; Altermatt, F.; Alther, R.; Choffat, Y.; Fronhofer, E.A.; Ganesanandamoorthy, P.; Garnier, A.; Griffiths, J.I.; et al. Biodiversity increases and decreases ecosystem stability. Nature 2018, 563, 109–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, W.; Huang, J.; Xu, H.; Zang, R.; Ding, Y.; Li, Y.; Lin, M.; Wang, J.; Zhang, C. Plant Functional Traits Are the Mediators in Regulating Effects of Abiotic Site Conditions on Aboveground Carbon Stock—Evidence from a 30 ha Tropical Forest Plot. Front. Plant Sci. 2019, 9, 1958. [Google Scholar] [CrossRef] [PubMed]
- Dickson, T.L.; Gross, K.L. Can the Results of Biodiversity-Ecosystem Productivity Studies Be Translated to Bioenergy Production? PLoS ONE 2015, 10, e0135253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmqvist, T.; Maltby, E.; Barker, T.; Mortimer, M.; Perrings, C.; Aronson, J.; De Groot, R.; Fitter, A.; Mace, G.; Norberg, J.; et al. Biodiversity, ecosystems and ecosystem services. In The Economics of Ecosystems and Biodiversity: Ecological and Economic Foundations, 1st ed.; Kumar, P., Ed.; Earthscan: Abingdon, UK, 2010; Volume 48, pp. 41–112. [Google Scholar]
- Harrison, P.A.; Berry, P.M.; Simpson, G.; Haslett, J.; Blicharska, M.; Bucur, M.M.; Dunford, R.W.; Egoh, B.N.; García-Llorente, M.; Geamănă, N.; et al. Linkages between biodiversity attributes and ecosystem services: A systematic review. Ecosyst. Serv. 2014, 9, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef]
- Breitkreuz, C.; Reitz, T.; Schulz, E.; Tarkka, M.T. Drought and Plant Community Composition Affect the Metabolic and Genotypic Diversity of Pseudomonas Strains in Grassland Soils. Microorganisms 2021, 9, 1677. [Google Scholar] [CrossRef]
- Raynaud, T.; Pivato, B.; Siol, M.; Spor, A.; Blouin, M. Soil microbes drive the effect of plant species and genotypic diversity interaction on productivity. Plant Soil 2021, 467, 165–180. [Google Scholar] [CrossRef]
- Hooper, D.U.; Vitousek, P.M. The Effects of Plant Composition and Diversity on Ecosystem Processes. Science 1997, 277, 1302–1305. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, R.K.; Raghubanshi, A.S.; Singh, J.S. Plant functional traits with particular reference to tropical deciduous forests: A review. J. Biosci. 2011, 36, 963–981. [Google Scholar] [CrossRef]
- MacArthur, R. Species packing and competitive equilibrium for many species. Theor. Popul. Biol. 1970, 1, 1–11. [Google Scholar] [CrossRef]
- Grace, J.; Michael Anderson, T.; Smith, M.D.; Seabloom, E.W.; Andelman, S.J.; Meche, G.; Weiher, E.; Allain, L.K.; Jutila, H.M.; Sankaran, M.; et al. Does species diversity limit productivity in natural grassland communities? Ecol. Lett. 2007, 10, 680–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, P.B.; Seabloom, E.W.; Borer, E.T.; Hillebrand, H.; Hautier, Y.; Hector, A.; Harpole, W.S.; O’Halloran, L.R.; Grace, J.; Anderson, T.M.; et al. Productivity Is a Poor Predictor of Plant Species Richness. Science 2011, 333, 1750–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardinale, B.J.; Hillebrand, H.; Harpole, W.S.; Gross, K.; Ptáčník, R. Separating the influence of resource ‘availability’ from resource ‘imbalance’ on productivity-diversity relationships. Ecol. Lett. 2009, 12, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Fridley, J.D. Resource availability dominates and alters the relationship between species diversity and ecosystem productivity in experimental plant communities. Oecologia 2002, 132, 271–277. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [Green Version]
- Heijden, M.V.; Klironomos, J.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Wilson, R.M.; Hough, M.; Verbeke, B.A.; Hodgkins, S.B.; Chanton, J.P.; Saleska, S.D.; Rich, V.I.; Tfaily, M.M.; Tyson, G.W.; Sullivan, M.B.; et al. Plant organic matter inputs exert a strong control on soil organic matter decomposition in a thawing permafrost peatland. Sci. Total Environ. 2022, 820, 152757. [Google Scholar] [CrossRef]
- Richardson, S.J.; Hartley, S.E.; Press, M.C. Climate warming experiments: Are tens a potential barrier to interpretation? Ecol. Entomol. 2000, 25, 367–370. [Google Scholar] [CrossRef]
- Rui, Y.C.; Wang, S.P.; Xu, Z.; Wang, Y.; Chen, C.; Zhou, X.; Kang, X.; Lu, S.; Hu, Y.; Lin, Q.; et al. Warming and grazing affect soil labile carbon and nitrogen pools differently in an alpine meadow of the Qinghai–Tibet Plateau in China. J. Soils Sediments 2011, 11, 903–914. [Google Scholar] [CrossRef]
- Rakkar, M.K.; Blanco-Canqui, H. Grazing of crop residues: Impacts on soils and crop production. Agric. Ecosyst. Environ. 2018, 258, 71–90. [Google Scholar] [CrossRef]
- Willis, K.J.; Whittaker, R.J. Species diversity—Scale matters. Science 2002, 295, 1245–1248. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Zhong, M.Y.; Wu, R.; Dong, Q.; Wang, K.; Shao, X.Q. Response of plant functional traits to grazing for three dominant species in alpine steppe habitat of the Qinghai–Tibet Plateau, China. Ecol. Res. 2016, 31, 515–524. [Google Scholar] [CrossRef]
- Shi, G.X.; Yao, B.Q.; Liu, Y.J.; Pan, J.B.; Jiang, S.J.; Wang, Y.B.; Wang, Z.B.; Feng, H.; Zhou, H.K. The effects of long-term warming on arbuscular mycorrhizal fungal communities depend on habitat type on the Qinghai–Tibet Plateau. Appl. Soil Ecol. 2021, 167, 104030. [Google Scholar] [CrossRef]
- Bu, H.Y.; Du, G.Z.; Chen, X.L.; Xu, X.L.; Liu, K.; Wen, S.J. Community-wide germination strategies in an alpine meadow on the eastern Qinghai–Tibet plateau: Phylogenetic and life-history correlates. Plant Ecol. 2008, 195, 87–98. [Google Scholar] [CrossRef]
- Liu, S.L.; Zhao, H.D.; Su, X.K.; Deng, L.; Dong, S.K.; Zhang, X. Spatio-temporal variability in rangeland conditions associated with climate change in the Altun Mountain National Nature Reserve on the Qinghai–Tibet Plateau over the past 15 years. Rangel. J. 2015, 37, 67–75. [Google Scholar] [CrossRef]
- Chen, B.X.; Zhang, X.Z.; Tao, J.; Wu, J.S.; Wang, J.S.; Shi, P.L.; Zhang, Y.J.; Yu, C.Q. The impact of climate change and anthropogenic activities on alpine grassland over the Qinghai–Tibet Plateau. Agric. For. Meteorol. 2014, 189, 11–18. [Google Scholar] [CrossRef]
- Chen, J.H.; Yan, F.; Lu, Q. Spatiotemporal variation of vegetation on the Qinghai–Tibet Plateau and the influence of climatic factors and human activities on vegetation trend (2000–2019). Remote Sens. 2020, 12, 3150. [Google Scholar] [CrossRef]
- Huang, K.; Zhang, Y.J.; Zhu, J.T.; Liu, Y.J.; Zu, J.X.; Zhang, J. The influences of climate change and human activities on vegetation dynamics in the Qinghai–Tibet Plateau. Remote Sens. 2016, 8, 876. [Google Scholar] [CrossRef] [Green Version]
- Xi, N.; Chen, D.; Bahn, M.; Wu, H.; Chu, C.; Cadotte, M.W.; Bloor, J.M. Drought soil legacy alters drivers of plant diversity-productivity relationships in oldfield systems. Sci. Adv. 2022, 8, eabn3368. [Google Scholar] [CrossRef]
- Reich, P.B.; Sendall, K.M.; Stefanski, A.; Rich, R.L.; Hobbie, S.E.; Montgomery, R.A. Effects of climate warming on photosynthesis in boreal tree species depend on soil moisture. Nature 2018, 562, 263–267. [Google Scholar] [CrossRef]
- Liancourt, P.; Sharkhuu, A.; Ariuntsetseg, L.; Boldgiv, B.; Helliker, B.R.; Plante, A.F.; Petraitis, P.S.; Casper, B.B. Temporal and spatial variation in how vegetation alters the soil moisture response to climate manipulation. Plant Soil 2011, 351, 249–261. [Google Scholar] [CrossRef]
- Kohyani, P.T.; Bossuyt, B.; Bonte, D.; Hoffmann, M. Grazing impact on plant spatial distribution and community composition. Plant Ecol. Evol. 2011, 144, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Milchunas, D.G.; Lauenroth, W.K.; Burke, I.C. Livestock grazing: Animal and plant biodiversity of shortgrass steppe and the relationship to ecosystem function. Oikos 1998, 83, 65–74. [Google Scholar] [CrossRef]
- Chaneton, E.J.; Facelli, J.M. Disturbance effects on plant community diversity: Spatial scales and dominance hierarchies. Vegetatio 1991, 93, 143–155. [Google Scholar] [CrossRef]
- Múgica, L.; Canals, R.M.; San Emeterio, L.; Peralta, F.J. Decoupling of traditional burnings and grazing regimes alters plant diversity and dominant species competition in high-mountain grasslands. Sci. Total Environ. 2021, 790, 147917. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Liu, L.; Wang, J.; Zhang, Y.S.; Xiao, C.W. Effects of warming on the bacterial community and its function in a temperate steppe. Sci. Total Environ. 2021, 792, 148409. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.R.; Schindlbacher, A.; Wang, J.X.; Yang, Y.J.; Song, Z.C.; You, Y.M.; Shi, Z.M.; Li, Z.Y.; Chen, L.; et al. Experimental warming reduced topsoil carbon content and increased soil bacterial diversity in a subtropical planted forest. Soil Biol. Biochem. 2019, 133, 155–164. [Google Scholar] [CrossRef]
- Bråthen, K.A.; Jahiri, X.; Jusdado, J.G.H.; Soininen, E.M.; Jensen, J.B. Fungal endophyte diversity in tundra grasses increases by grazing. Fungal Ecol. 2015, 17, 41–51. [Google Scholar] [CrossRef]
- Yvon-Durocher, G.; Montoya, J.M.; Trimmer, M.; Woodward, G. Warming alters the size spectrum and shifts the distribution of biomass in freshwater ecosystems. Glob. Chang. Biol. 2011, 17, 1681–1694. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.H.; Willis, C.G.; Klein, J.A.; Ma, Z.; Li, J.; Zhou, H.K.; Zhao, X.Q. Recovery of plant species diversity during long-term experimental warming of a species-rich alpine meadow community on the Qinghai–Tibet plateau. Biol. Conserv. 2017, 213, 218–224. [Google Scholar] [CrossRef]
- Shi, G.X.; Yao, B.Q.; Liu, Y.J.; Jiang, S.J.; Wang, W.Y.; Pan, J.B.; Zhao, X.Q.; Feng, H.Y.; Zhou, H.K. The phylogenetic structure of AMF communities shifts in response to gradient warming with and without winter grazing on the Qinghai–Tibet Plateau. Appl. Soil Ecol. 2017, 121, 31–40. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE v5 enables improved estimates of phylogenetic tree confidence by ensemble bootstrapping. bioRxiv 2021. [Google Scholar] [CrossRef]
- Rosauer, D.F.; Laffan, S.W.; Crisp, M.D.; Donnellan, S.C.; Cook, L.G. Phylogenetic endemism: A new approach for identifying geographical concentrations of evolutionary history. Mol. Ecol. 2009, 18, 4061–4072. [Google Scholar] [CrossRef] [PubMed]
- Helmus, M.R.; Bland, T.J.; Williams, C.K.; Ives, A.R. Phylogenetic measures of biodiversity. Am. Nat. 2007, 169, E68–E83. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment * | Average Degree | Average Weighted Degree | Modularity | Statistical Inference | Graph Density | Nodes | Edges |
|---|---|---|---|---|---|---|---|
| CK + NG | 0.577 | 0.577 | 0.507 | 226.254 | 0.011 | 52 | 15 |
| CK + G | 0.706 | 0.627 | 0.736 | 230.432 | 0.014 | 51 | 18 |
| A + NG | 0.375 | 0.375 | 0.519 | 190.919 | 0.008 | 48 | 9 |
| A + G | 0.851 | 0.766 | 0.847 | 215.927 | 0.019 | 47 | 20 |
| B + NG | 0.356 | 0.356 | 0.688 | 175.426 | 0.008 | 45 | 8 |
| B + G | 0.304 | 0.130 | 0.333 | 176.360 | 0.007 | 46 | 7 |
| C + NG | 0.600 | 0.440 | 0.975 | 218.481 | 0.012 | 50 | 15 |
| C + G | 0.809 | 0.723 | 0.268 | 213.521 | 0.018 | 47 | 19 |
| D + NG | 0.682 | 0.591 | 0.814 | 190.962 | 0.016 | 44 | 15 |
| D + G | 0.732 | 0.634 | 0.423 | 177.897 | 0.018 | 41 | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Ma, L.; Yang, X.; Zhang, Q.; She, Y.; Chang, T.; Su, H.; Sun, J.; Shao, X.; Zhou, H.; et al. Biodiversity and Ecosystem Function under Simulated Gradient Warming and Grazing. Plants 2022, 11, 1428. https://doi.org/10.3390/plants11111428
Zhang Z, Ma L, Yang X, Zhang Q, She Y, Chang T, Su H, Sun J, Shao X, Zhou H, et al. Biodiversity and Ecosystem Function under Simulated Gradient Warming and Grazing. Plants. 2022; 11(11):1428. https://doi.org/10.3390/plants11111428
Chicago/Turabian StyleZhang, Zhonghua, Li Ma, Xiaoyuan Yang, Qian Zhang, Yandi She, Tao Chang, Hongye Su, Jian Sun, Xinqing Shao, Huakun Zhou, and et al. 2022. "Biodiversity and Ecosystem Function under Simulated Gradient Warming and Grazing" Plants 11, no. 11: 1428. https://doi.org/10.3390/plants11111428