
Identification of Spring Wheat with Superior Agronomic Performance under Contrasting Nitrogen Managements Using Linear Phenotypic Selection Indices


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phenotyping
2.2. Data Analyses
2.3. Theory of Selection Indices and Selection Parameters
3. Results
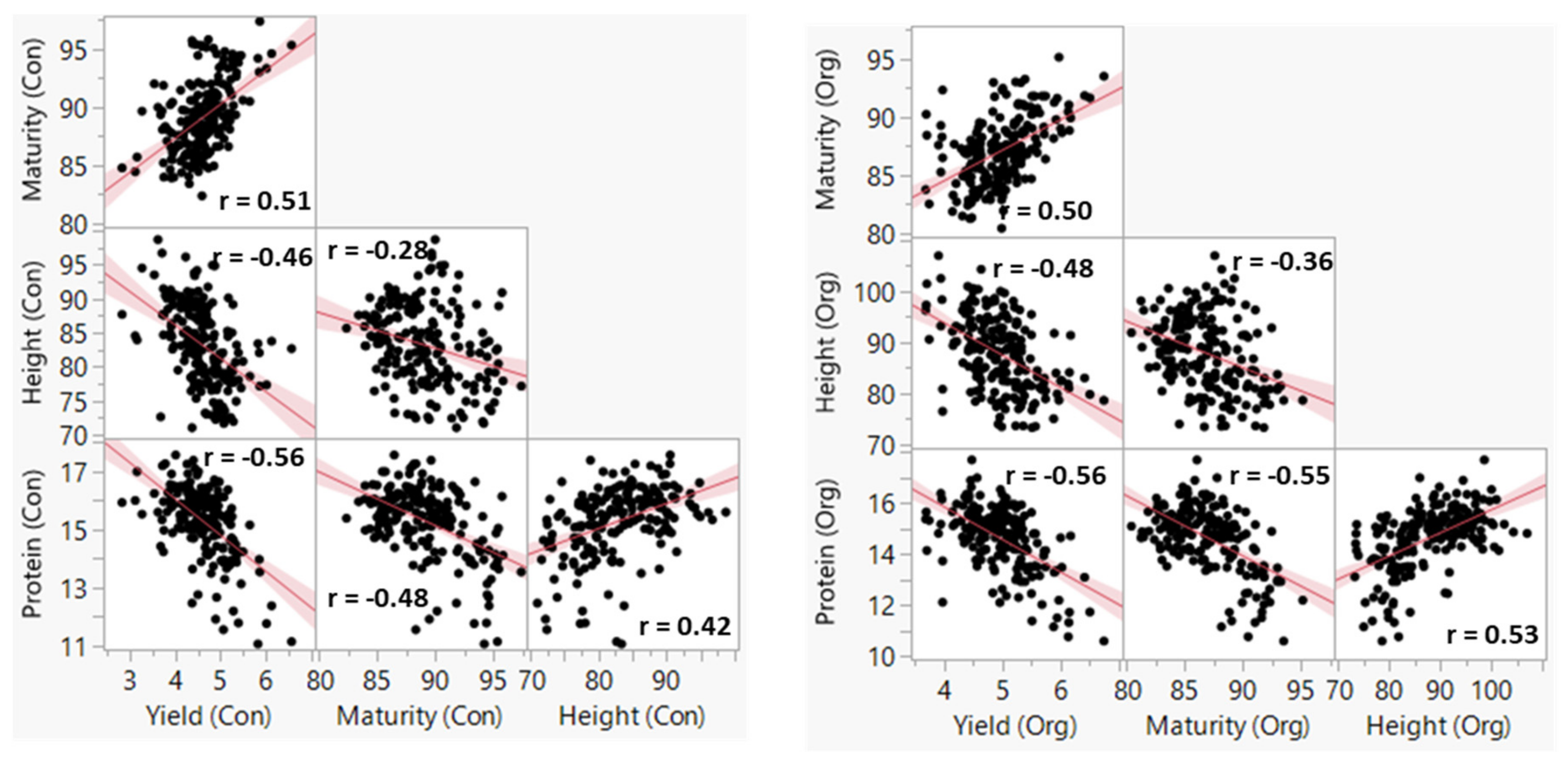
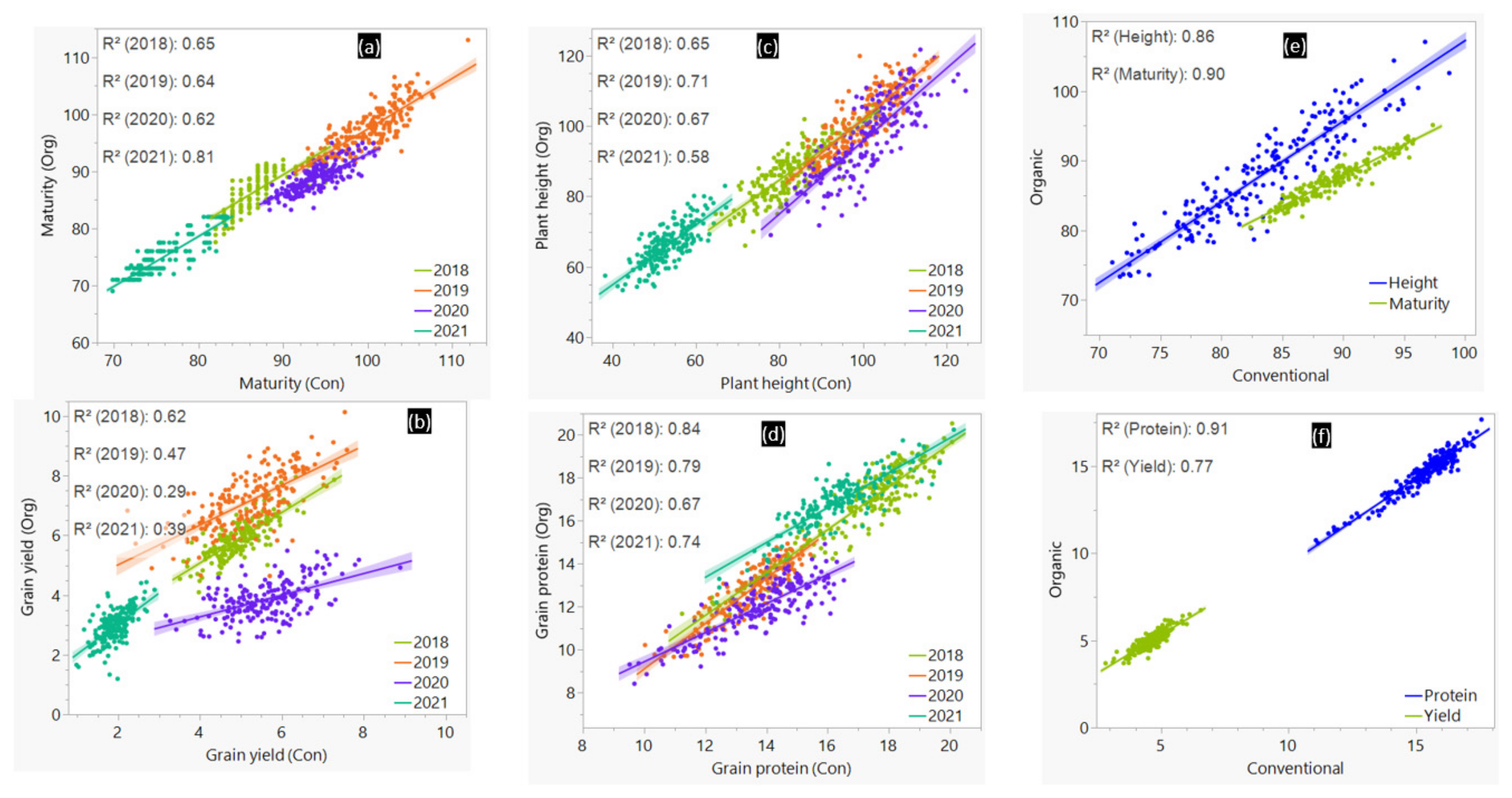
3.1. Phenotypic Variation
3.2. Single Trait Selection
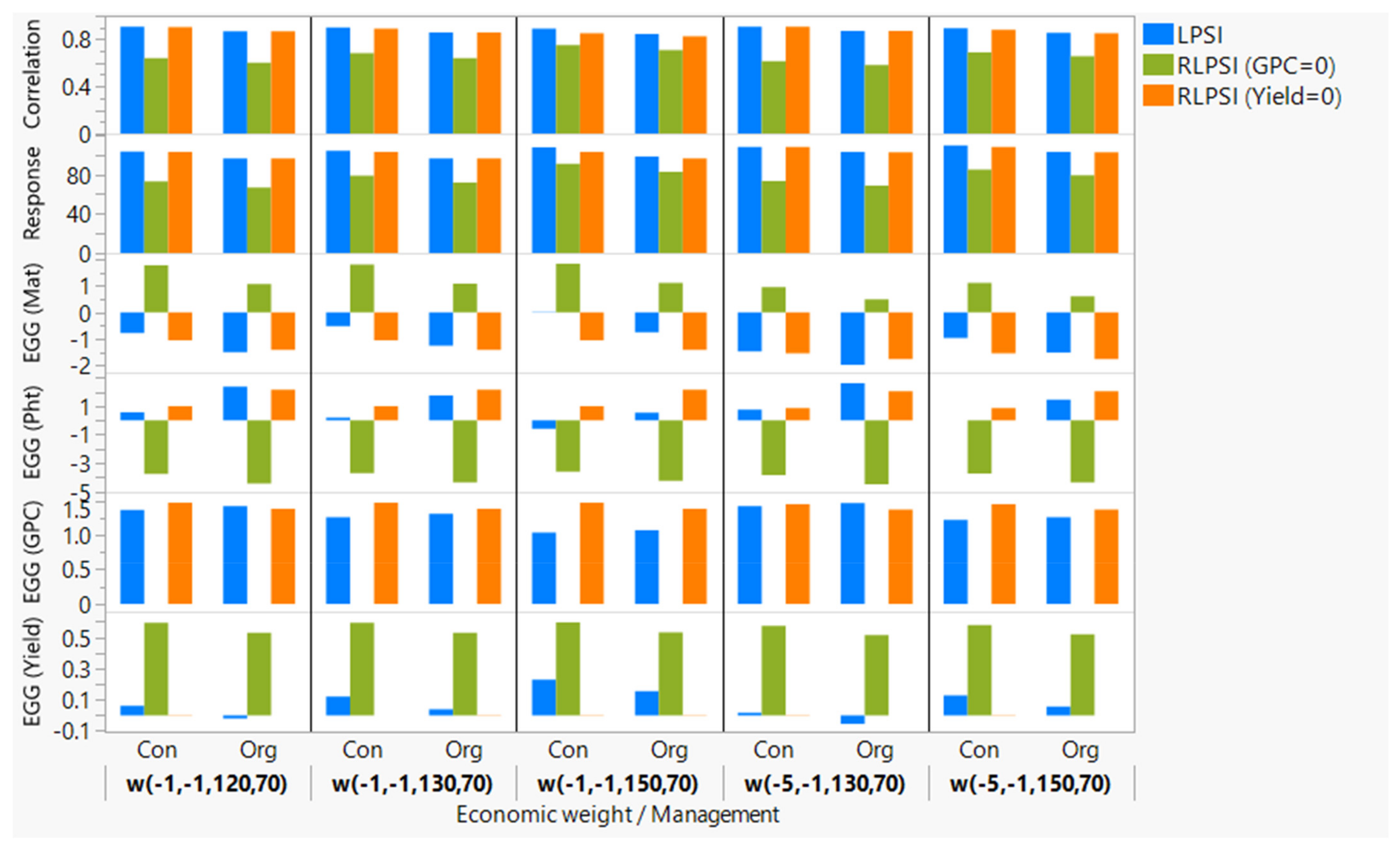
3.3. Comparisons of Economic Weights
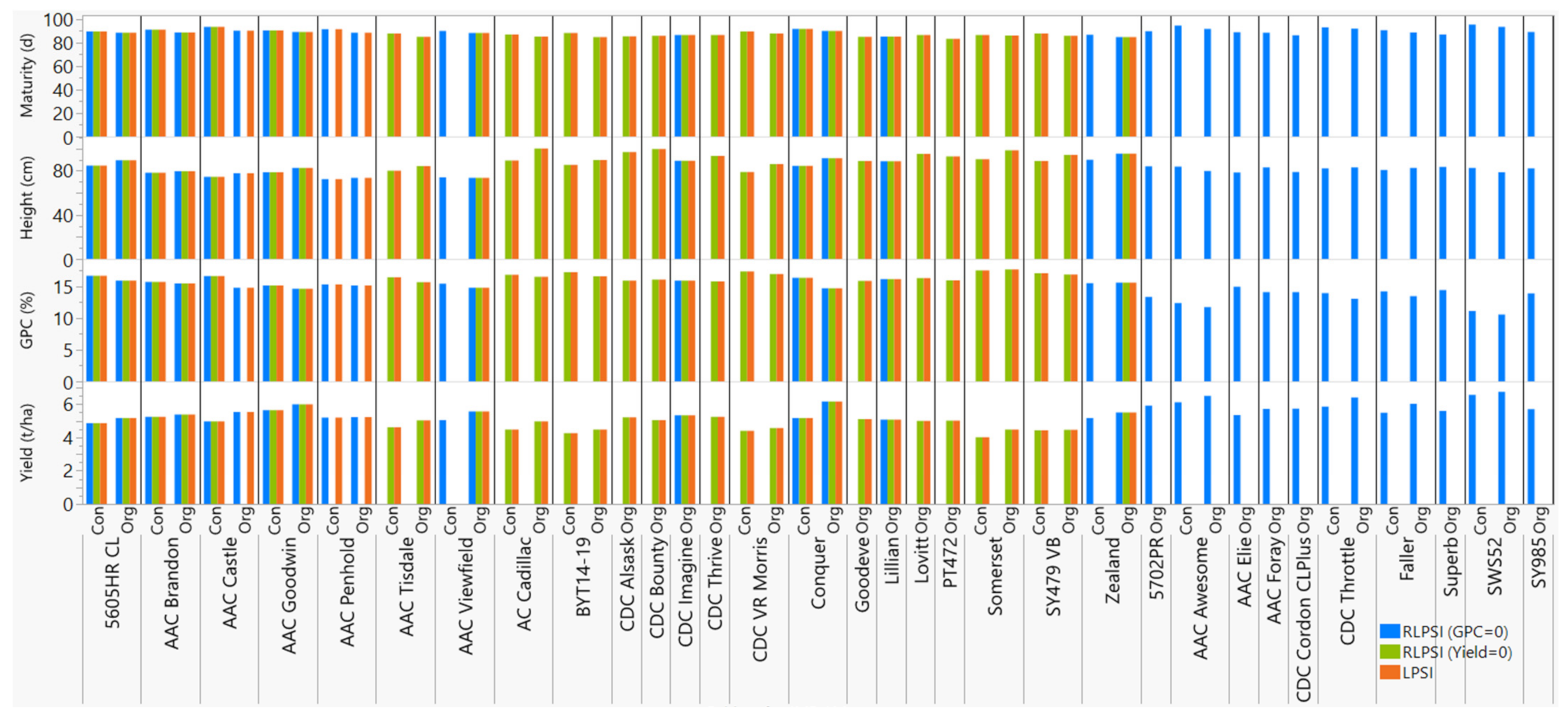
3.4. Multi-Trait Selection Using RLPSI by Restricting Grain Protein Content
3.5. Multi-Trait Selection Using RLPSI by Restricting Grain Yield
3.6. Multi-Trait Selection Using LPSI
3.7. Comparison of Economic Weights and Selection Indices
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Campion, A.; Oury, F.-X.; Heumez, E.; Rolland, B. Conventional versus organic farming systems: Dissecting comparisons to improve cereal organic breeding strategies. Org. Agric. 2020, 10, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Hawkesford, M.J. Reducing the reliance on nitrogen fertilizer for wheat production. J. Cereal Sci. 2014, 59, 276–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lammerts van Bueren, E.T.; Jones, S.S.; Tamm, L.; Murphy, K.M.; Myers, J.R.; Leifert, C.; Messmer, M.M. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. Wagening. J. Life Sci. 2011, 58, 193–205. [Google Scholar] [CrossRef]
- Wolfe, M.S.; Baresel, J.P.; Desclaux, D.; Goldringer, I.; Hoad, S.; Kovacs, G.; Löschenberger, F.; Miedaner, T.; Østergård, H.; Lammerts van Bueren, E.T. Developments in breeding cereals for organic agriculture. Euphytica 2008, 163, 323. [Google Scholar] [CrossRef] [Green Version]
- Osman, A.M.; Almekinders, C.J.M.; Struik, P.C.; Lammerts van Bueren, E.T. Adapting spring wheat breeding to the needs of the organic sector. Wagening. J. Life Sci. 2016, 76, 55–63. [Google Scholar] [CrossRef]
- Hoad, S.; Topp, C.; Davies, K. Selection of cereals for weed suppression in organic agriculture: A method based on cultivar sensitivity to weed growth. Euphytica 2008, 163, 355–366. [Google Scholar] [CrossRef]
- Zerner, M.C.; Gill, G.S.; Vandeleur, R.K. Effect of height on the competitive ability of wheat with oats. Agron. J. 2008, 100, 1729–1734. [Google Scholar] [CrossRef]
- Mason, H.; Navabi, A.; Frick, B.; O’Donovan, J.; Spaner, D. Cultivar and seeding rate effects on the competitive ability of spring cereals grown under organic production in Northern Canada. Agron. J. 2007, 99, 1199–1207. [Google Scholar] [CrossRef]
- Mason, H.E.; Navabi, A.; Frick, B.L.; O’Donovan, J.T.; Spaner, D.M. The weed-competitive ability of Canada western red spring wheat cultivars grown under organic management. Crop Sci. 2007, 47, 1167–1176. [Google Scholar] [CrossRef]
- Cudney, D.W.; Jordan, L.S.; Hall, A.E. Effect of wild oat (Avena fatua) infestations on light interception and growth-rate of wheat (Triticum aestivum). Weed Sci. 1991, 399, 175–179. [Google Scholar] [CrossRef]
- Watson, P.R.; Derksen, D.A.; Van Acker, R.C. The ability of 29 barley cultivars to compete and withstand competition. Weed Sci. 2006, 54, 783–792. [Google Scholar] [CrossRef]
- Huel, D.G.; Hucl, P. Genotypic variation for competitive ability in spring wheat. Plant Breed. 1996, 115, 325–329. [Google Scholar] [CrossRef]
- Lemerle, D.; Verbeek, B.; Cousens, R.D.; Coombes, N.E. The potential for selecting wheat varieties strongly competitive against weeds. Weed Res. 1996, 36, 505–513. [Google Scholar] [CrossRef]
- Baenziger, P.S.; Salah, I.; Little, R.S.; Santra, D.K.; Regassa, T.; Wang, M.Y. Structuring an efficient organic wheat breeding program. Sustainability 2011, 3, 1190–1205. [Google Scholar] [CrossRef] [Green Version]
- Kirk, A.P.; Fox, S.L.; Entz, M.H. Comparison of organic and conventional selection environments for spring wheat. Plant Breed. 2012, 131, 687–694. [Google Scholar] [CrossRef]
- Reid, T.A.; Yang, R.C.; Salmon, D.F.; Navabi, A.; Spaner, D. Realized gains from selection for spring wheat grain yield are different in conventional and organically managed systems. Euphytica 2011, 177, 253–266. [Google Scholar] [CrossRef]
- Kaut, E.; Mason, E.; Navabi, A.; O’Donovan, T.; Spaner, D. Organic and conventional management of mixtures of wheat and spring cereals. Agron. Sustain. Dev. 2008, 28, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Reid, T.A.; Yang, R.C.; Salmon, D.F.; Spaner, D. Should spring wheat breeding for organically managed systems be conducted on organically managed land? Euphytica 2009, 169, 239–252. [Google Scholar] [CrossRef]
- Kaut, A.H.E.E.; Mason, H.E.; Navabi, A.; O’Donovan, J.T.; Spaner, D. Performance and stability of performance of spring wheat variety mixtures in organic and conventional management systems in western Canada. J. Agric. Sci. 2009, 147, 141–153. [Google Scholar] [CrossRef]
- Chen, H.; Nguyen, K.; Iqbal, M.; Beres, B.L.; Hucl, P.J.; Spaner, D. The performance of spring wheat cultivar mixtures under conventional and organic management in Western Canada. Agric. Geosci. Environ. 2020, 3, e20003. [Google Scholar] [CrossRef] [Green Version]
- Kubota, H.; Quideau, S.A.; Hucl, P.J.; Spaner, D.M. The effect of weeds on soil arbuscular mycorrhizal fungi and agronomic traits in spring wheat (Triticum aestivum L.) under organic management in Canada. Can. J. Plant Sci. 2015, 95, 615–627. [Google Scholar] [CrossRef]
- Mason, H.; Navabi, A.; Frick, B.; O’Donovan, J.; Niziol, D.; Spaner, D. Does growing Canadian Western Hard Red Spring wheat under organic management alter its breadmaking quality? Renew. Agric. Food Syst. 2007, 22, 157–167. [Google Scholar] [CrossRef]
- Zou, J.; Semagn, K.; Iqbal, M.; N’Diaye, A.; Chen, H.; Asif, M.; Navabi, A.; Perez-Lara, E.; Pozniak, C.; Yang, R.C.; et al. Mapping QTLs controlling agronomic traits in the Attila x CDC Go spring wheat population under organic management using 90K SNP array. Crop Sci. 2017, 57, 365–377. [Google Scholar] [CrossRef]
- Xiang, R.; Semagn, K.; Iqbal, M.; Chen, H.; Yang, R.-C.; Spaner, D. Phenotypic performance and associated QTL of ‘Peace’ × ‘CDC Stanley’ mapping population under conventional and organic management systems. Crop Sci. 2021, 61, 3469–3483. [Google Scholar] [CrossRef]
- Semagn, K.; Iqbal, M.; Chen, H.; Perez-Lara, E.; Bemister, D.H.; Xiang, R.; Zou, J.; Asif, M.; Kamran, A.; N’Diaye, A.; et al. Physical mapping of QTL associated with agronomic and end-use quality traits in spring wheat under conventional and organic management systems. Theor. Appl. Genet. 2021, 134, 3699–3719. [Google Scholar] [CrossRef]
- Semagn, K.; Iqbal, M.; Chen, H.; Perez-Lara, E.; Bemister, D.H.; Xiang, R.; Zou, J.; Asif, M.; Kamran, A.; N’Diaye, A.; et al. Physical mapping of QTL in four spring wheat populations under conventional and organic management systems. I. Earliness. Plants 2021, 10, 853. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Iqbal, M.; N’Diaye, A.; Pozniak, C.; Ciechanowska, I.; Barbu, S.-P.; Spaner, D. Genome-wide association mapping of agronomic traits and grain characteristics in spring wheat under conventional and organic management systems. Crop Sci. 2022, 62, 1069–1087. [Google Scholar] [CrossRef]
- Chen, H.; Bemister, D.H.; Iqbal, M.; Strelkov, S.E.; Spaner, D.M. Mapping genomic regions controlling agronomic traits in spring wheat under conventional and organic managements. Crop Sci. 2020, 60, 2038–2052. [Google Scholar] [CrossRef]
- Asif, M.; Yang, R.C.; Navabi, A.; Iqbal, M.; Kamran, A.; Lara, E.P.; Randhawa, H.; Pozniak, C.; Spaner, D. Mapping QTL, selection differentials, and the effect of Rht-B1 under organic and conventionally managed systems in the Attila × CDC Go spring wheat mapping population. Crop Sci. 2015, 55, 1129–1142. [Google Scholar] [CrossRef]
- Semagn, K.; Iqbal, M.; Crossa, J.; Jarquin, D.; Howard, R.; Chen, H.; Bemister, D.H.; Beres, B.L.; Randhawa, H.; N’Diaye, A.; et al. Genome-based prediction of agronomic traits in spring wheat under conventional and organic management systems. Theor. Appl. Genet. 2022, 135, 537–552. [Google Scholar] [CrossRef]
- Semagn, K.; Crossa, J.; Cuevas, J.; Iqbal, M.; Ciechanowska, I.; Henriquez, M.H.; Randhawa, H.; Beres, B.L.; Aboukhaddour, R.; McCallum, B.D.; et al. Comparison of single-trait and multi-trait genomic predictions on agronomic and disease resistance traits in spring wheat. Theor. Appl. Genet. 2022; in press. [Google Scholar] [CrossRef]
- Gelli, M.; Mitchell, S.E.; Liu, K.; Clemente, T.E.; Weeks, D.P.; Zhang, C.; Holding, D.R.; Dweikat, I.M. Mapping QTLs and association of differentially expressed gene transcripts for multiple agronomic traits under different nitrogen levels in sorghum. BMC Plant Biol. 2016, 16, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Liu, C.; Wang, Y.; He, T.; Gao, R.; Xu, H.; Guo, G.; Li, Y.; Zhou, L.; Lu, R.; et al. Expression analysis of nitrogen metabolism-related genes reveals differences in adaptation to low-nitrogen stress between two different barley cultivars at seedling stage. Int. J. Genom. 2018, 2018, 8152860. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Y.; Zhao, Y.; Zhang, Y.; Zhang, J.; Ma, H.; Han, Y. Transcriptome analysis reveals nitrogen deficiency induced alterations in leaf and root of three cultivars of potato (Solanum tuberosum L.). PLoS ONE 2020, 15, e0240662. [Google Scholar] [CrossRef] [PubMed]
- Allier, A.; Lehermeier, C.; Charcosset, A.; Moreau, L.; Teyssèdre, S. Improving short- and long-term genetic gain by accounting for within-family variance in optimal cross-selection. Front. Genet. 2019, 10, 1006. [Google Scholar] [CrossRef] [Green Version]
- Branlard, G.; Pierre, J.; Rousset, M. Selection indices for quality evaluation in wheat breeding. Theor. Appl. Genet. 1992, 84, 57–64. [Google Scholar] [CrossRef]
- Ullah, H.; Subthain, H.; Khalil, I.H.; Khan, W.U.; Jamal, Y.; Alam, M. Stress selection indices an acceptable tool to screen superior wheat genotypes under irrigated and rain-fed conditions. Pak. J. Bot. 2014, 46, 627–638. [Google Scholar]
- Malav, A.K.; Monpara, I.B.A.; Raghuwanshi, S.S. Selection indices for yield improvement in bread wheat (Triticum aestivum L.). J. Pure Appl. Microbiol. 2016, 10, 2801–2805. [Google Scholar] [CrossRef]
- Shah, S.; Mehta, D.R.; Raval, L. Selection indices in bread wheat [Triticum aestivum L.]. Electron. J. Plant Breed. 2016, 7, 459–463. [Google Scholar] [CrossRef]
- Fellahi, Z.E.A.; Hannachi, A.; Bouzerzour, H. Expected genetic gains from mono trait and indexbased selection in advanced bread wheat (Triticum aestivum L.) populations. Rev. Fac. Nac. 2020, 73, 9131–9141. [Google Scholar] [CrossRef]
- Karthikeya Reddy, S.G.P.; Babariya, C.A. Selection indices for yield improvement in bread wheat (Triticum aestivum L.). Electron. J. Plant Breed. 2020, 11, 314–317. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.F. A discriminant function for plant selection. Ann. Eugenic. 1936, 7, 240–250. [Google Scholar] [CrossRef]
- Kempthorne, O.; Nordskog, A.W. Restricted selection indices. Biometrics 1959, 15, 10–19. [Google Scholar] [CrossRef]
- Cerón-Rojas, J.J.; Crossa, J.; Sahagún-Castellanos, J.; Castillo-González, F.; Santacruz-Varela, A. A selection index method based on eigenanalysis. Crop Sci. 2006, 46, 1711–1721. [Google Scholar] [CrossRef]
- Cerón-Rojas, J.J.; Sahagún-Castellanos, J.; Castillo-González, F.; Santacruz-Varela, A.; Crossa, J. A restricted selection index method based on eigenanalysis. J. Agric. Biol. Environ. Stat. 2008, 13, 440–457. [Google Scholar] [CrossRef]
- Cerón-Rojas, J.J.; Crossa, J. Efficiency of a constrained linear genomic selection index to predict the net genetic merit in plants. G3 (Bethesda) 2019, 9, 3981–3994. [Google Scholar] [CrossRef] [Green Version]
- Cerón-Rojas, J.J.; Crossa, J. The statistical theory of linear selection indices from phenotypic to genomic selection. Crop Sci. 2022, 62, 537–563. [Google Scholar] [CrossRef]
- Cerón-Rojas, J.J.; Crossa, J. Expectation and variance of the estimator of the maximized selection response of linear selection indices with normal distribution. Theor. Appl. Genet. 2020, 133, 2743–2758. [Google Scholar] [CrossRef]
- Cerón-Rojas, J.J.; Crossa, J. Linear Selection Indices in Modern Plant Breeding; Springer: Cham, Switzerland, 2018; pp. 1–256. [Google Scholar]
- Céron-Rojas, J.J.; Crossa, J. (Eds.) The linear phenotypic selection index theory. In Linear Selection Indices in Modern Plant Breeding; Springer: Cham, Switzerland, 2018; pp. 15–42. [Google Scholar]
- Céron-Rojas, J.J.; Crossa, J. (Eds.) Constrained linear phenotypic selection indices. In Linear Selection Indices in Modern Plant Breeding; Springer: Cham, Switzerland, 2018; pp. 43–69. [Google Scholar]
- Cerón-Rojas, J.J.; Crossa, J.; Sahagún-Castellanos, J. Statistical sampling properties of the coefficients of three phenotypic selection indices. Crop Sci. 2016, 56, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Cerón-Rojas, J.J.; Toledo, F.H.; Crossa, J. Optimum and decorrelated constrained multistage linear phenotypic selection indices theory. Crop Sci. 2019, 59, 2585–2600. [Google Scholar] [CrossRef] [Green Version]
- Cerón-Rojas, J.J.; Toledo, F.H.; Crossa, J. The relative efficiency of two multistage linear phenotypic selection indices to predict the net genetic merit. Crop Sci. 2019, 59, 1037–1051. [Google Scholar] [CrossRef] [Green Version]
- Hazel, L.N.; Dickerson, G.E.; Freeman, A.E. The selection index—Then, now, and for the future. J. Dairy Sci. 1994, 77, 3236–3251. [Google Scholar] [CrossRef]
- Perez-Elizalde, S.; Cerón-Rojas, J.J.; Crossa, J.; Fleury, D.; Alvarado, G. Rindsel: An R package for phenotypic and molecular selection indices used in Plant Breeding. Methods Mol. Biol. 2014, 1145, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Gebre-Mariam, H.; Larter, E.N. Genetic response to index selection for grain yield, kernel weight and per cent protein in four wheat crosses. Plant Breed. 1996, 115, 459–464. [Google Scholar] [CrossRef]
- Maich, R.H.; Chaves, A.G.; Coraglio, M.C.; Costero, B.; Torres, L.E. Agronomic performance of bread wheat (Triticum aestivum L.) and hexaploid triticale (X Triticosecale Wittmack) based on the use of a selection index. Cereal Res. Commun. 2006, 34, 1123–1127. [Google Scholar] [CrossRef]
- Lopez-Cruz, M.; Olson, E.; Rovere, G.; Crossa, J.; Dreisigacker, S.; Mondal, S.; Singh, R.; Campos, G.d.l. Regularized selection indices for breeding value prediction using hyper-spectral image data. Sci. Rep. 2020, 10, 8195. [Google Scholar] [CrossRef]
- Juliana, P.; He, X.; Poland, J.; Roy, K.K.; Malaker, P.K.; Mishra, V.K.; Chand, R.; Shrestha, S.; Kumar, U.; Roy, C.; et al. Genomic selection for spot blotch in bread wheat breeding panels, full-sibs and half-sibs and index-based selection for spot blotch, heading and plant height. Theor. Appl. Genet. 2022, 135, 1965–1983. [Google Scholar] [CrossRef]
- Magnussen, S. Selection index: Economic weights for maximum simultaneous genetic gain. Theor. Appl. Genet. 1990, 79, 289–293. [Google Scholar] [CrossRef]
- Ramos Guimarães, P.H.; Guimarães Santos Melo, P.; Centeno Cordeiro, A.C.; Pereira Torga, P.; Nakano Rangel, P.H.; Pereira de Castro, A. Index selection can improve the selection efficiency in a rice recurrent selection population. Euphytica 2021, 217, 95. [Google Scholar] [CrossRef]
- Sabouri, H.; Rabiei, B.; Fazlalipour, M. Use of selection indices based on multivariate analysis for improving grain yield in rice. Rice Sci. 2008, 15, 303–310. [Google Scholar] [CrossRef]
- Mistele, M.; Zeddies, J.; Urz, H.F.; Melchinger, A.E. Economic aspects of breeding for yield and quality traits in forage maize: I. Determination of economic weights. Plant Breed. 1994, 112, 102–109. [Google Scholar] [CrossRef]
- Famula, T.R. A comparison of restricted selection index and linear programming in sire selection. Theor. Appl. Genet. 1992, 84, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Xu, S. Restricted multistage selection indices. Genet. Sel. Evol. 1997, 29, 193–203. [Google Scholar] [CrossRef]
- Pešek, J.; Baker, R.J. Desired improvement in relation to selection indices. Can. J. Plant Sci. 1969, 49, 803–804. [Google Scholar] [CrossRef] [Green Version]
- Semagn, K.; Iqbal, M.; Alachiotis, N.; N’Diaye, A.; Pozniak, C.; Spaner, D. Genetic diversity and selective sweeps in historical and modern Canadian spring wheat cultivars using the 90K SNP array. Sci. Rep. 2021, 11, 23773. [Google Scholar] [CrossRef]
- Semagn, K.; Iqbal, M.; Jarquin, D.; Crossa, J.; Howard, R.; Ciechanowska, I.; Henriquez, M.H.; Randhawa, H.; Aboukhaddour, R.; McCallum, B.D.; et al. Genomic predictions for common bunt, FHB, stripe rust, leaf rust, and leaf spotting resistance in spring wheat. Genes 2022, 13, 565. [Google Scholar] [CrossRef]
- Yan, W. Estimation of the optimal number of replicates in crop variety trials. Front. Plant Sci. 2021, 11, 590762. [Google Scholar] [CrossRef]
- Yan, W.; Tinker, N.A.; Bekele, W.A.; Mitchell-Fetch, J.; Fregeau-Reid, J. Theoretical unification and practical integration of conventional methods and genomic selection in Plant Breeding. Crop Breed. Genet. Genom. 2019, 1, e190003. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, G.; Rodríguez, F.M.; Pacheco, A.; Burgueño, J.; Crossa, J.; Vargas, M.; Pérez-Rodríguez, P.; Lopez-Cruz, M.A. META-R: A software to analyze data from multi-environment Plant Breeding trials. Crop J. 2020, 8, 745–756. [Google Scholar] [CrossRef]
- Laidig, F.; Piepho, H.-P.; Rentel, D.; Drobek, T.; Meyer, U.; Huesken, A. Breeding progress, environmental variation and correlation of winter wheat yield and quality traits in German official variety trials and on-farm during 1983–2014. Theor. Appl. Genet. 2017, 130, 223–245. [Google Scholar] [CrossRef] [Green Version]
- Longin, C.F.H.; Gowda, M.; Mühleisen, J.; Ebmeyer, E.; Kazman, E.; Schachschneider, R.; Schacht, J.; Kirchhoff, M.; Zhao, Y.; Reif, J.C. Hybrid wheat: Quantitative genetic parameters and consequences for the design of breeding programs. Theor. Appl. Genet. 2013, 126, 2791–2801. [Google Scholar] [CrossRef]
- Michel, S.; Löschenberger, F.; Ametz, C.; Pachler, B.; Sparry, E.; Bürstmayr, H. Simultaneous selection for grain yield and protein content in genomics-assisted wheat breeding. Theor. Appl. Genet. 2019, 132, 1745–1760. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; Sall, J. JMP statistical discovery software. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 188–194. [Google Scholar] [CrossRef]
- Mallard, J. La theorie et le calcul des index de selection avec restriction: Synthese critique. Biometrics 1972, 28, 713–735. [Google Scholar] [CrossRef]
- Pesek, J.; Baker, R.J. An application of index selection to the improvement of self-pollinated species. Can. J. Plant Sci. 1970, 50, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Batista, L.G.; Gaynor, R.C.; Margarido, G.R.A.; Byrne, T.; Amer, P.; Gorjanc, G.; Hickey, J.M. Long-term comparison between index selection and optimal independent culling in Plant Breeding programs with genomic prediction. PLoS ONE 2021, 16, e0235554. [Google Scholar] [CrossRef]
- Paprstein, F.; Blazek, J. A selection index for evaluating sweet cherry genetic resources and choosing parents for crosses. Acta Hortic. 1996, 410, 405–412. [Google Scholar] [CrossRef]
- Elgin, J.H., Jr.; Hill, R.R., Jr.; Zeiders, K.E. Comparison of four methods of multiple trait selection for five traits in alfalfa. Crop Sci. 1970, 10, 190–193. [Google Scholar] [CrossRef]
- Sharma, R.C.; Duveiller, E. Selection index for improving helminthosporium leaf blight resistance, maturity, and kernel weight in spring wheat. Crop Sci. 2003, 43, 2031–2036. [Google Scholar] [CrossRef]
- Marulanda, J.J.; Mi, X.; Utz, H.F.; Melchinger, A.E.; Würschum, T.; Longin, C.F.H. Optimum breeding strategies using genomic and phenotypic selection for the simultaneous improvement of two traits. Theor. Appl. Genet. 2021, 134, 4025–4042. [Google Scholar] [CrossRef]
- Dolan, D.J.; Stuthman, D.D.; Kolb, F.L.; Hewings, A.D. Multiple trait selection in a recurrent selection population in oat (Avena sativa L.). Crop Sci. 1996, 36, 1207–1211. [Google Scholar] [CrossRef]
- Cuthbert, R.D.; DePauw, R.M.; Knox, R.E.; Singh, A.K.; McCaig, T.N.; McCallum, B.; Fetch, T. AAC Brandon hard red spring wheat. Can. J. Plant Sci. 2016, 97, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, R.D.; DePauw, R.M.; Knox, R.E.; Singh, A.K.; McCallum, B.; Fetch, T. AAC Viewfield hard red spring wheat. Can. J. Plant Sci. 2018, 99, 102–110. [Google Scholar] [CrossRef]
- DePauw, R.M.; Thomas, J.B.; Knox, R.E.; Clarke, J.M.; Fernandez, M.R.; McCaig, T.N.; McLeod, J.G. AC Cadillac hard red spring wheat. Can. J. Plant Sci. 1998, 78, 459–462. [Google Scholar] [CrossRef]
- DePauw, R.M.; Knox, R.E.; Thomas, J.B.; Smith, M.; Clarke, J.M.; Clarke, F.R.; McCaig, T.N.; Fernandez, M.R. Goodeve hard red spring wheat. Can. J. Plant Sci. 2009, 89, 937–944. [Google Scholar] [CrossRef]
- DePauw, R.M.; Knox, R.E.; Clarke, J.M.; McCaig, T.N.; Clarke, F.R.; Fernandez, M.R. Lovitt hard red spring wheat. Can. J. Plant Sci. 2004, 84, 811–814. [Google Scholar] [CrossRef] [Green Version]
- Burt, A.J.; Humphreys, D.G.; Fetch, J.M.; Green, D.; Fetch, T.G.; McCallum, B.D.; Menzies, J.; Aboukhaddour, R.; Henriquez, M.A.; Kumar, S. AAC redstar hard red spring wheat. Can. J. Plant Sci. 2021, 101, 274–283. [Google Scholar] [CrossRef]
- Fox, S.L.; Townley-Smith, T.F.; Humphreys, D.G.; McCallum, B.D.; Fetch, T.G.; Gaudet, D.A.; Gilbert, J.A.; Menzies, J.G.; Noll, J.S.; Howes, N.K. Somerset hard red spring wheat. Can. J. Plant Sci. 2006, 86, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Spaner, D.; Iqbal, M.; Navabi, A.; Strenzke, K.; Beres, B. Zealand hard red spring wheat. Can. J. Plant Sci. 2018, 98, 1409–1415. [Google Scholar] [CrossRef]
- Brown, P.D.; Randhawa, H.S.; Fetch, J.M.; Meiklejohn, M.; Fox, S.L.; Humphreys, D.G.; Green, D.; Wise, I.; Fetch, T.; Gilbert, J.; et al. Conquer red spring wheat. Can. J. Plant Sci. 2016, 97, 147–152. [Google Scholar] [CrossRef]
- DePauw, R.M.; Townley-Smith, T.F.; Humphreys, G.; Knox, R.E.; Clarke, F.R.; Clarke, J.M. Lillian hard red spring wheat. Can. J. Plant Sci. 2005, 85, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, H.S.; Brown, P.D.; Fetch, J.M.; Fetch, T.; McCallum, B.; Henriquez, M.A.; Menzies, J. AAC Castle red spring wheat. Can. J. Plant Sci. 2022, 102, 278–284. [Google Scholar] [CrossRef]
- Cuthbert, R.D.; DePauw, R.M.; Knox, R.E.; Singh, A.K.; McCaig, T.N.; McCallum, B.; Fetch, T.; Beres, B.L. AAC penhold Canada prairie spring red wheat. Can. J. Plant Sci. 2017, 98, 207–214. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Economic Weight | Scenarios | EGG (Mat) | EGG (Pht) | EGG (Yld) | EGG (GPC) | Response | Correlation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Con | Org | Con | Org | Con | Org | Con | Org | Con | Org | Con | Org | ||
| Heritability | 0.51 | 0.38 | 0.43 | 0.58 | 0.48 | 0.32 | 0.80 | 0.60 | |||||
| Wt (−1, −1, 120, 70) | LPSI | −0.78 | −1.49 | 0.58 | 2.37 | 0.06 | −0.02 | 1.37 | 1.43 | 103.41 | 96.35 | 0.91 | 0.87 |
| RLPSI (GPC = 0) | 1.77 | 1.07 | −3.76 | −4.43 | 0.59 | 0.53 | 0.00 | 0.00 | 73.10 | 66.74 | 0.64 | 0.60 | |
| RLPSI (Yield = 0) | −1.05 | −1.40 | 0.99 | 2.16 | 0.00 | 0.00 | 1.47 | 1.39 | 103.08 | 96.31 | 0.91 | 0.87 | |
| Wt (−1, −1, 130, 70) | LPSI | −0.52 | −1.24 | 0.18 | 1.76 | 0.12 | 0.04 | 1.26 | 1.31 | 104.32 | 96.44 | 0.90 | 0.86 |
| RLPSI (GPC = 0) | 1.79 | 1.08 | −3.70 | −4.36 | 0.59 | 0.53 | 0.00 | 0.00 | 79.03 | 72.02 | 0.68 | 0.64 | |
| RLPSI (Yield = 0) | −1.05 | −1.40 | 0.99 | 2.16 | 0.00 | 0.00 | 1.47 | 1.39 | 103.08 | 96.31 | 0.89 | 0.86 | |
| Wt (−1, −1, 150, 70) | LPSI | −0.01 | −0.75 | −0.58 | 0.54 | 0.23 | 0.15 | 1.04 | 1.07 | 107.82 | 98.37 | 0.89 | 0.84 |
| RLPSI (GPC = 0) | 1.82 | 1.11 | −3.61 | −4.24 | 0.59 | 0.53 | 0.00 | 0.00 | 90.90 | 82.61 | 0.75 | 0.71 | |
| RLPSI (Yield = 0) | −1.05 | −1.40 | 0.99 | 2.16 | 0.00 | 0.00 | 1.47 | 1.39 | 103.08 | 96.31 | 0.85 | 0.83 | |
| Wt (−5, −1, 130, 70) | LPSI | −1.46 | −1.96 | 0.76 | 2.61 | 0.02 | −0.06 | 1.42 | 1.47 | 108.30 | 102.90 | 0.91 | 0.87 |
| RLPSI (GPC = 0) | 0.96 | 0.49 | −3.83 | −4.49 | 0.57 | 0.51 | 0.00 | 0.00 | 73.47 | 68.84 | 0.62 | 0.58 | |
| RLPSI (Yield = 0) | −1.54 | −1.75 | 0.87 | 2.05 | 0.00 | 0.00 | 1.45 | 1.37 | 108.27 | 102.63 | 0.91 | 0.87 | |
| Wt (−5, −1, 150, 70) | LPSI | −0.96 | −1.51 | 0.00 | 1.47 | 0.13 | 0.06 | 1.23 | 1.26 | 109.76 | 102.92 | 0.89 | 0.86 |
| RLPSI (GPC = 0) | 1.11 | 0.60 | −3.73 | −4.36 | 0.58 | 0.52 | 0.00 | 0.00 | 84.98 | 79.17 | 0.69 | 0.66 | |
| RLPSI (Yield = 0) | −1.54 | −1.75 | 0.87 | 2.05 | 0.00 | 0.00 | 1.45 | 1.37 | 108.27 | 102.63 | 0.88 | 0.85 | |
| Cultivar (Line) | Pedigree | Wheat Class * | Registration Year | Maturity (d) | Plant Height (cm) | Yield (t/ha) | GPC (%) ** | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Con | Org | Con | Org | Con | Org | Con | Org | ||||
| 5605HR CL | 99S2232-10/99S3228-4 | CNHR | 2013 | 89.6 | 88.4 | 84.8 | 89.6 | 4.9 | 5.2 | 16.7 | 15.9 |
| AAC Brandon | Superb/CDC Osler//ND744 | CWRS | 2013 | 91.0 | 88.7 | 78.1 | 79.6 | 5.2 | 5.4 | 15.7 | 15.5 |
| AAC Penhold | 5700PR/HY644-BE//HY469 | CPSR | 2014 | 91.4 | 88.5 | 72.4 | 73.7 | 5.2 | 5.2 | 15.3 | 15.2 |
| AAC Viewfield | Stettler/Glenn | CWRS | 2016 | 90.1 | 88.2 | 74.1 | 73.6 | 5.1 | 5.6 | 15.4 | 14.8 |
| AC Cadillac | BW90 * 3/BW553 | CWRS | 1996 | 86.8 | 85.2 | 89.4 | 100.1 | 4.5 | 5.0 | 16.9 | 16.5 |
| AAC Goodwin | Carberry/AC Cadillac | CPSR | 2017 | 90.5 | 89.0 | 78.6 | 82.5 | 5.6 | 6.0 | 15.2 | 14.6 |
| Zealand | Alvena/IAS64/ALDAN//URES/3/TNMU/4/TNMU) | CWRS | 2016 | 86.6 | 84.7 | 90.1 | 95.5 | 5.2 | 5.5 | 15.5 | 15.6 |
| BYT14-19 | Carberry/PT764//CDC Stanley | CWRS | - | 88.1 | 84.9 | 85.4 | 89.9 | 4.3 | 4.5 | 17.2 | 16.6 |
| CDC Alsask | AC Elsa/AC Cora | CWRS | 2005 | 87.7 | 85.3 | 88.1 | 96.9 | 4.6 | 5.2 | 16.4 | 15.9 |
| CDC Bounty | Katepwa/W-82624//Kenyon | CWRS | 2000 | 86.1 | 85.9 | 90.0 | 99.7 | 4.3 | 5.0 | 16.5 | 16.1 |
| CDC Imagine | CDC Teal * 4/Fidel(FS2) | CWRS | 2002 | 88.4 | 86.4 | 84.2 | 89.2 | 4.7 | 5.3 | 16.3 | 15.9 |
| CDC Thrive | CDC Bounty/CDC Imagine | CWRS | 2010 | 88.4 | 86.5 | 89.4 | 93.5 | 4.8 | 5.2 | 16.0 | 15.8 |
| CDC VR Morris | CDC Bounty/FHB9 | CWRS | 2012 | 89.6 | 87.8 | 78.8 | 86.1 | 4.4 | 4.6 | 17.4 | 17.0 |
| Conquer | HY639/99 EPWA-Mdg 61 | CNHR | 2010 | 91.7 | 89.9 | 84.5 | 91.4 | 5.2 | 6.2 | 16.4 | 14.7 |
| Goodeve | 94B43-BLW4/AC Intrepid | CWRS | 2007 | 85.9 | 85.0 | 80.5 | 88.9 | 4.5 | 5.1 | 16.0 | 15.9 |
| AAC Castle | Conquer/CDN Bison//5701PR | CPSR | 2018 | 93.4 | 90.3 | 74.6 | 77.7 | 5.0 | 5.5 | 16.6 | 14.8 |
| Lillian | BW621 * 3/90B07-AU2B | CNHR | 2003 | 86.8 | 85.3 | 84.4 | 88.6 | 4.3 | 5.1 | 16.2 | 16.2 |
| Lovitt | 8405-JC3C * 2/BW152 | CWRS | 2002 | 86.8 | 86.5 | 87.2 | 95.3 | 4.1 | 5.0 | 16.2 | 16.3 |
| AAC Tisdale | Somerset/BW865//Waskada | CWRS | 2017 | 87.9 | 85.0 | 80.0 | 84.2 | 4.6 | 5.0 | 16.4 | 15.7 |
| PT472 | PT425/Helios//PT435 | CWRS | - | 84.8 | 83.2 | 86.7 | 93.0 | 4.7 | 5.0 | 16.0 | 16.0 |
| Somerset | 90B01-AD4D/Pasqua | CWRS | 2005 | 86.5 | 86.1 | 90.5 | 98.7 | 4.0 | 4.5 | 17.6 | 17.7 |
| SY479 VB | 01S2004-2-13/Glenn | CWRS | 2016 | 87.8 | 85.9 | 88.8 | 94.5 | 4.4 | 4.5 | 17.1 | 16.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iqbal, M.; Semagn, K.; Céron-Rojas, J.J.; Crossa, J.; Jarquin, D.; Howard, R.; Beres, B.L.; Strenzke, K.; Ciechanowska, I.; Spaner, D. Identification of Spring Wheat with Superior Agronomic Performance under Contrasting Nitrogen Managements Using Linear Phenotypic Selection Indices. Plants 2022, 11, 1887. https://doi.org/10.3390/plants11141887
Iqbal M, Semagn K, Céron-Rojas JJ, Crossa J, Jarquin D, Howard R, Beres BL, Strenzke K, Ciechanowska I, Spaner D. Identification of Spring Wheat with Superior Agronomic Performance under Contrasting Nitrogen Managements Using Linear Phenotypic Selection Indices. Plants. 2022; 11(14):1887. https://doi.org/10.3390/plants11141887
Chicago/Turabian StyleIqbal, Muhammad, Kassa Semagn, J. Jesus Céron-Rojas, José Crossa, Diego Jarquin, Reka Howard, Brian L. Beres, Klaus Strenzke, Izabela Ciechanowska, and Dean Spaner. 2022. "Identification of Spring Wheat with Superior Agronomic Performance under Contrasting Nitrogen Managements Using Linear Phenotypic Selection Indices" Plants 11, no. 14: 1887. https://doi.org/10.3390/plants11141887