
Silicon Enhances Plant Vegetative Growth and Soil Water Retention of Soybean (Glycine max) Plants under Water-Limiting Conditions



Abstract
:1. Introduction
2. Results
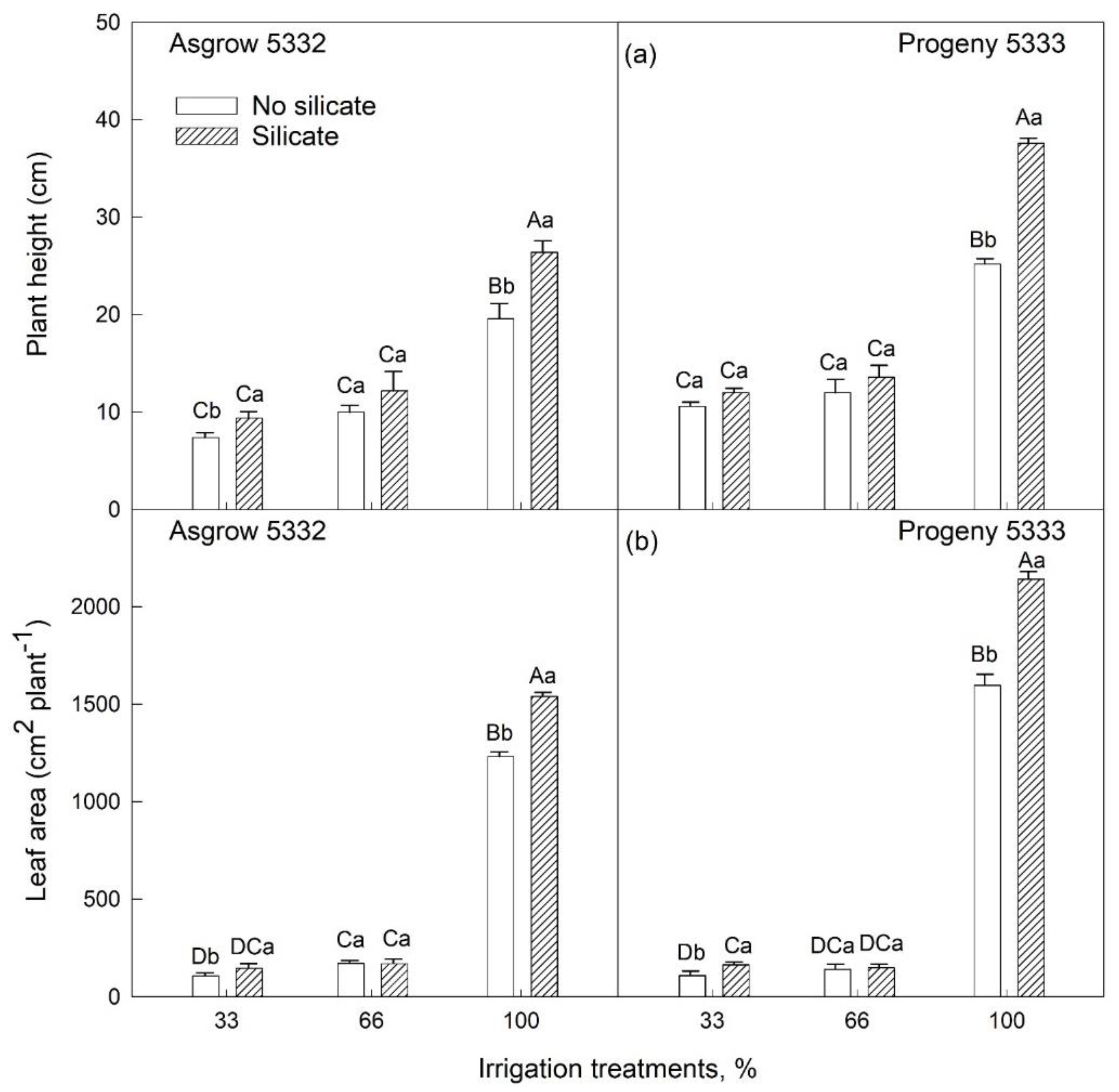
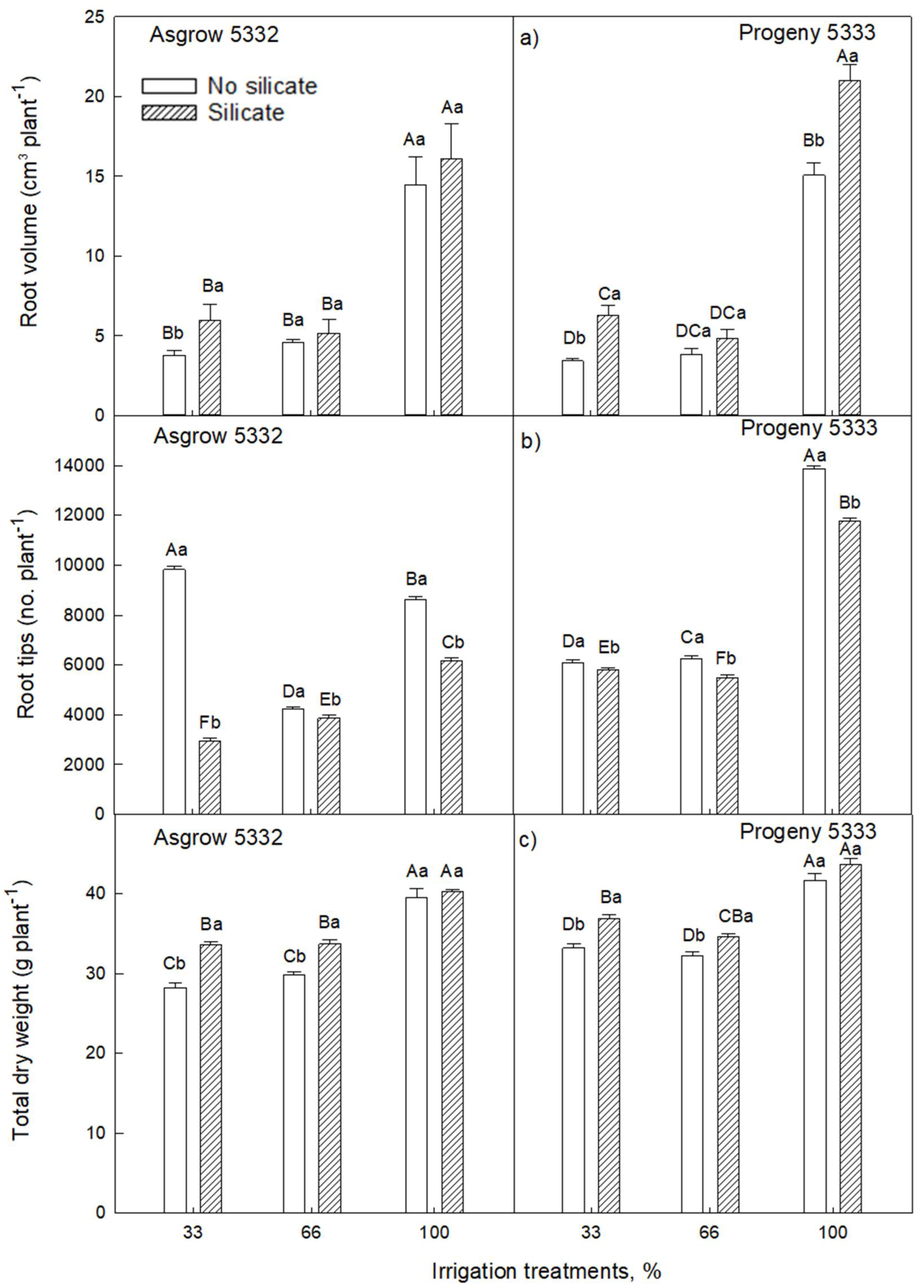
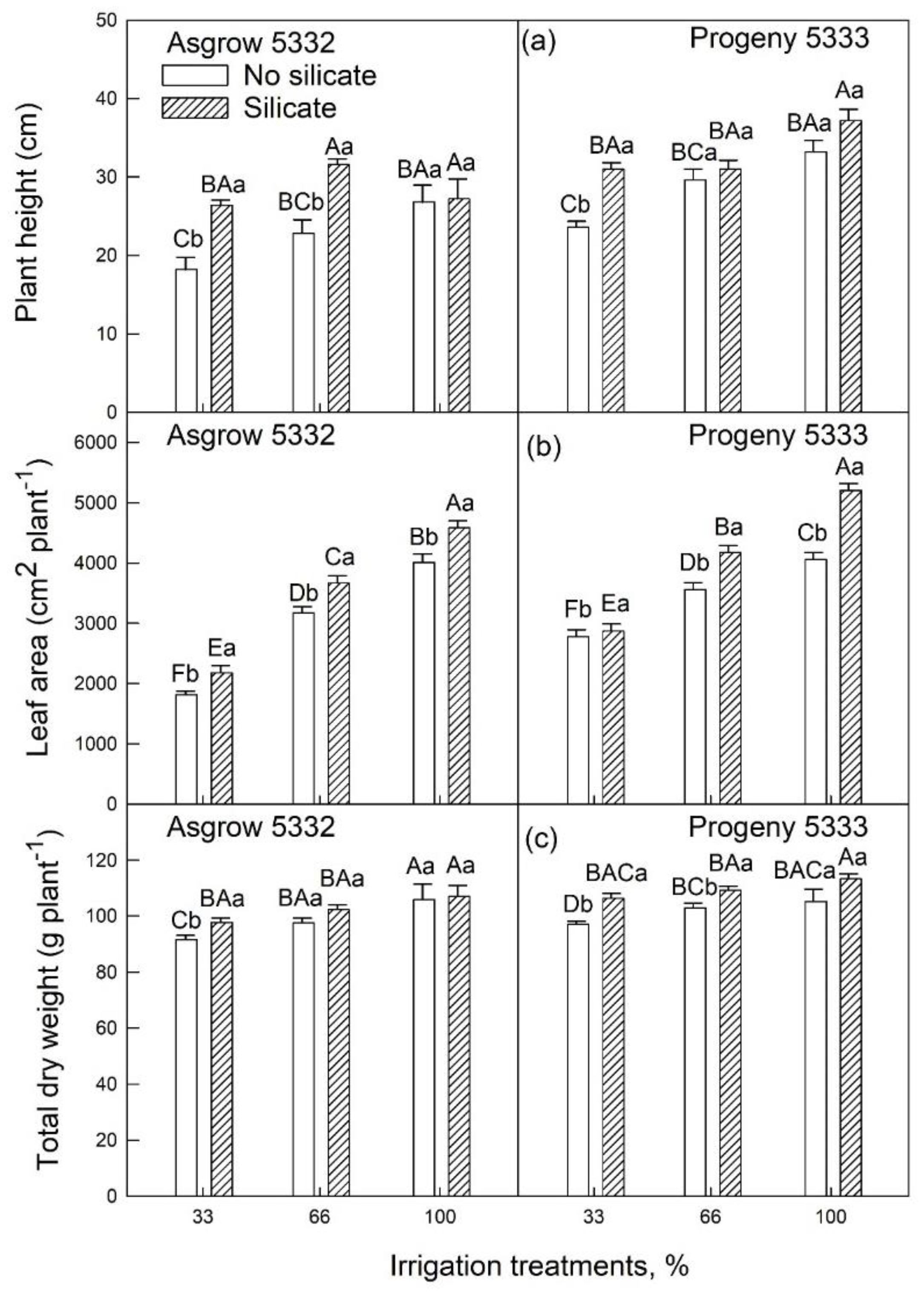
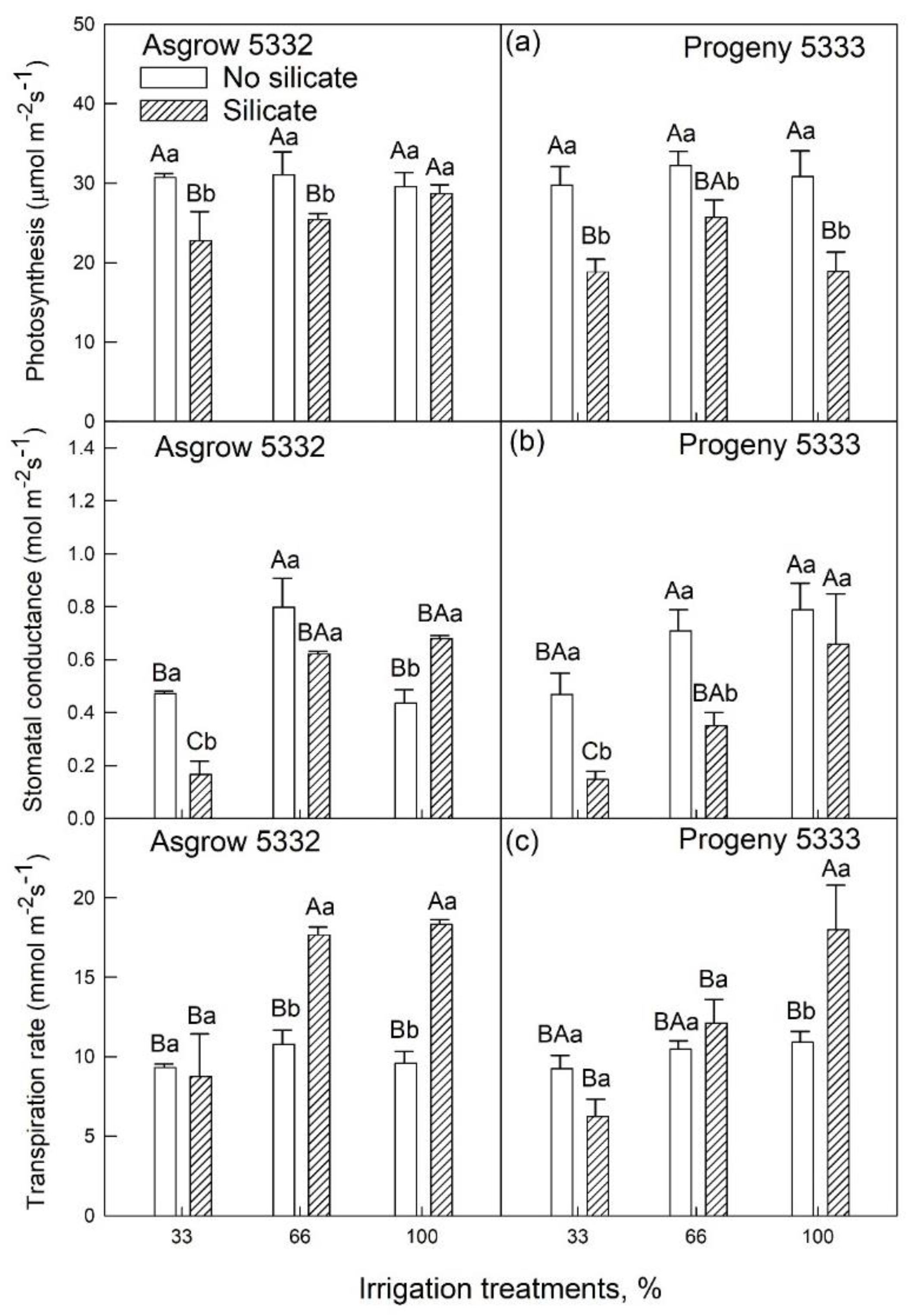
2.1. Silicon Application on Vegetative Growth of Soybeans
2.2. Effects of Silicon on Vegetative Growth of Soybeans (7–27 Days after Emergence) Grown under Water-Limiting Conditions
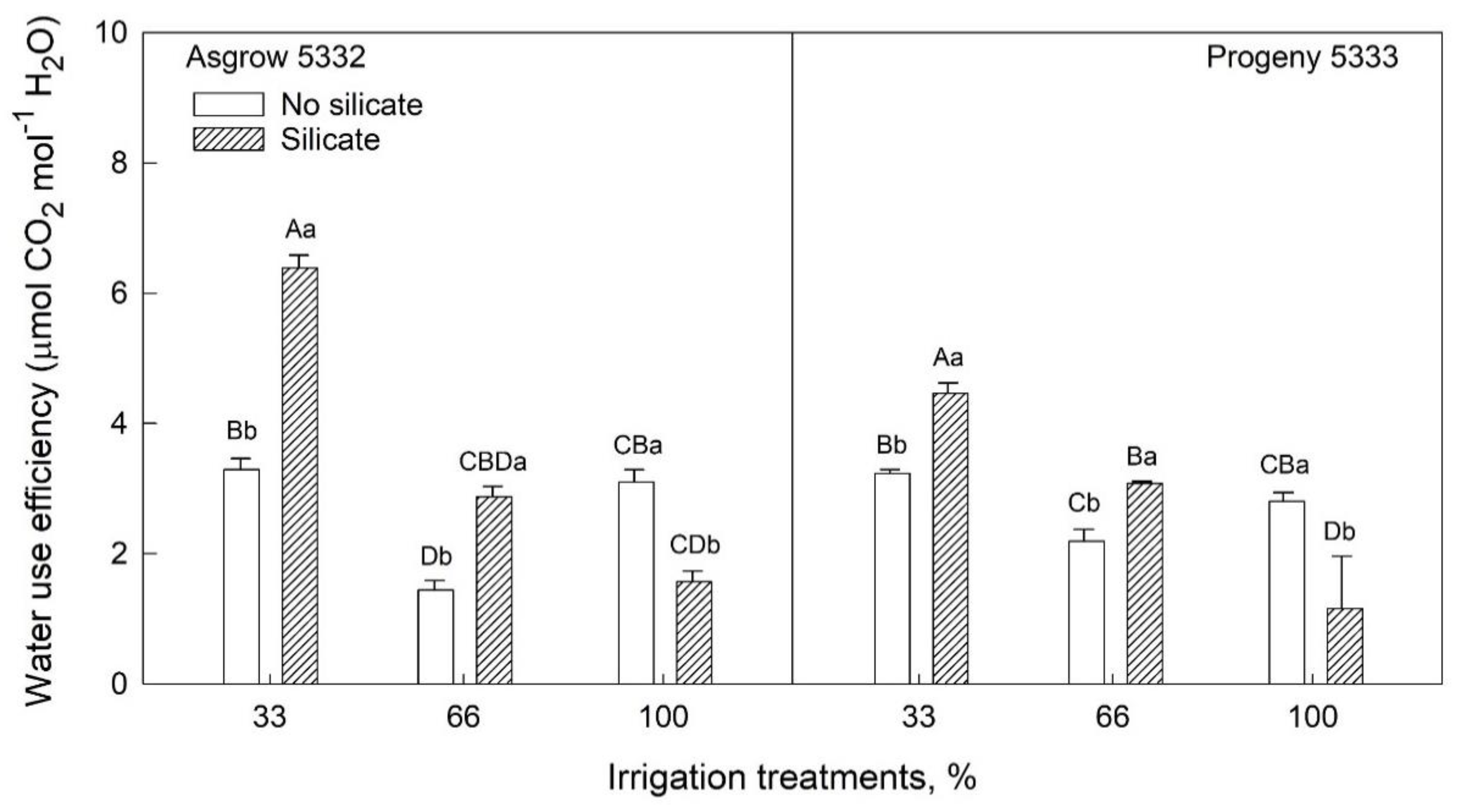
2.3. Effects of Silicon on Growth and Physiology of Soybeans (25–45 Days after Emergence) Grown under Water-Limiting Conditions
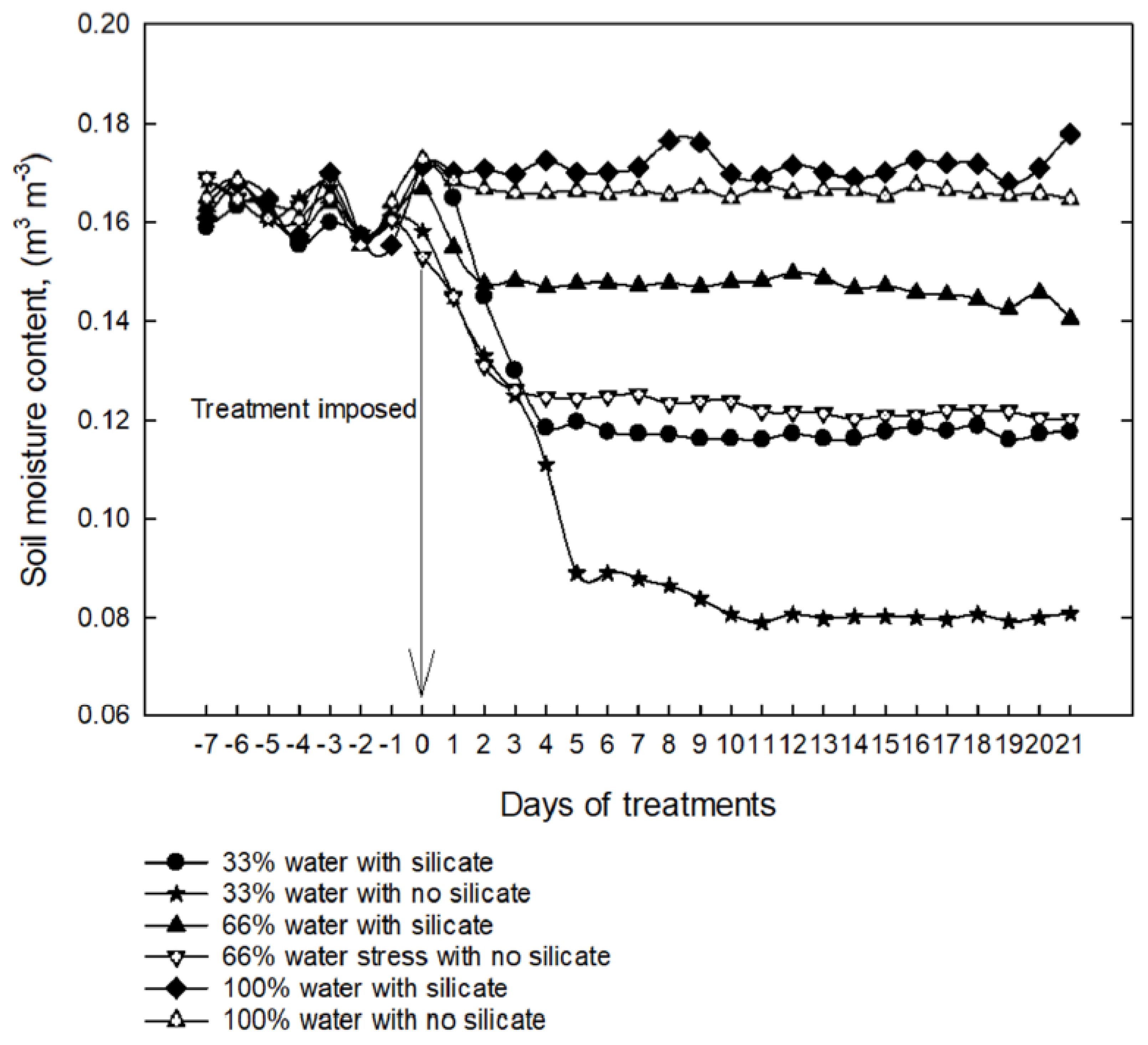
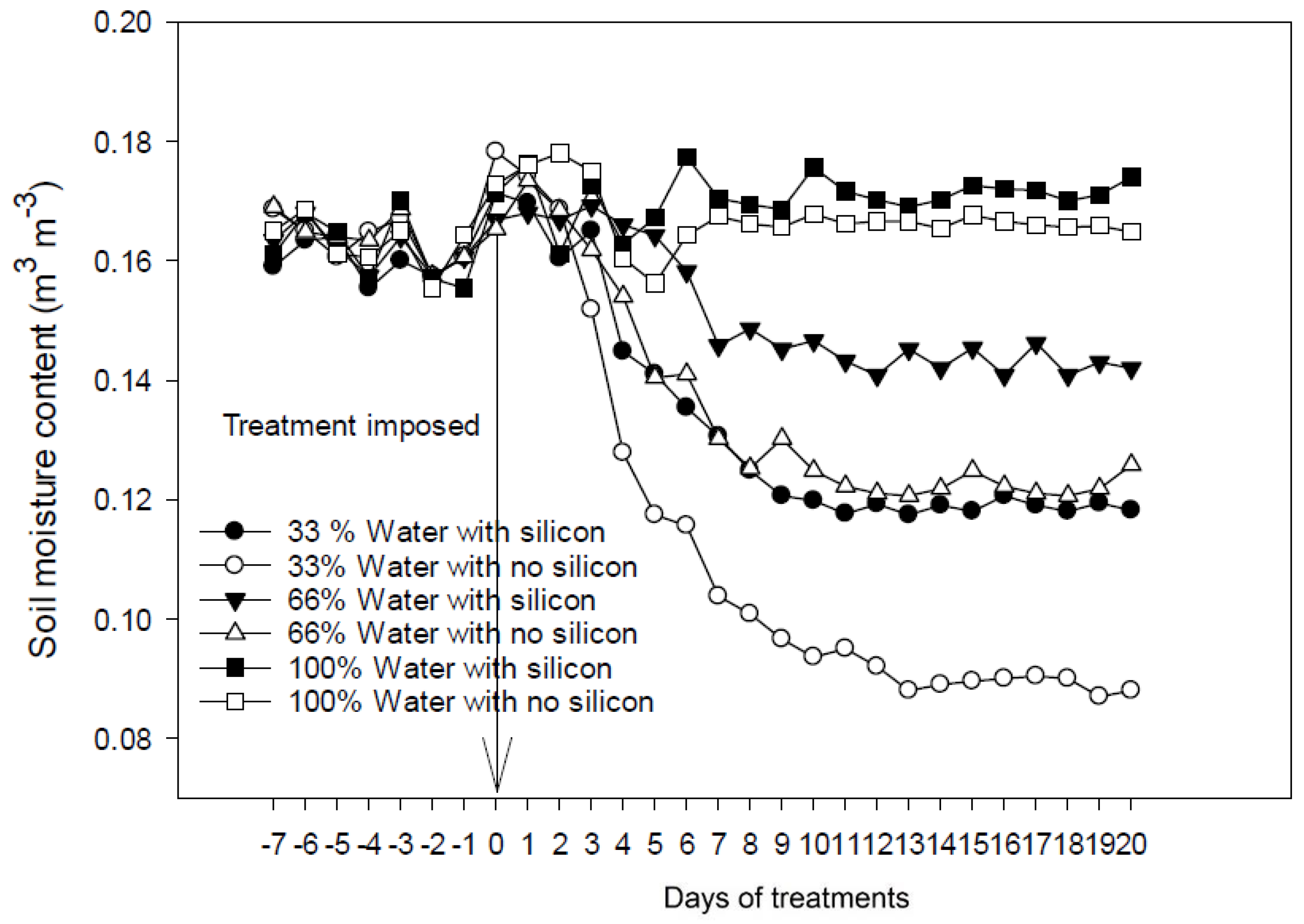
2.4. Silicon Application on Soil Moisture Content under Water-Limiting Conditions
3. Discussion
4. Materials and Methods
4.1. Experimental Facility and Seed Material
4.2. Silicate and Soil Moisture Treatments
4.3. Seedling Growth and Biomass Yield
4.4. Root Growth Parameters
4.5. Photosynthesis Rate, Stomatal Conductance, Transpiration Rate, and Chlorophyll Fluorescence
4.6. Soil Moisture Content
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liang, X.-Z.; Wu, Y.; Chambers, R.G.; Schmoldt, D.L.; Gao, W.; Liu, C.; Liu, Y.-A.; Sun, C.; Kennedy, J.A. Determining climate effects on US total agricultural productivity. Proc. Natl. Acad. Sci. USA 2017, 114, E2285–E2292. [Google Scholar] [CrossRef] [Green Version]
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops that feed the World 2. Soybean—worldwide production, use, and constraints caused by pathogens and pests. Food Secur. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- Dwevedi, A.; Kayastha, A.M. Soybean: A Multifaceted Legume with Enormous Economic Capabilities, Soybean-Biochemistry, Chemistry and Physiology, Prof; Tzi-Bun, N., Ed.; InTech: London, UK, 2011; ISBN 978-953-307-219-7. Available online: http://www.intechopen.com/books/soybean-biochemistry-chemistry-andphysiology/soybean-a-multifaceted-legume-with-enormous-economic-capabilities (accessed on 13 May 2022).
- Ash, M.; Knisley, S. United States Department of Agriculture. Economic Research Services. 2016. Available online: http://www.ers.usda.gov/topics/crops/soybeans-oil-crops/related-data-statistics.aspx (accessed on 13 May 2022).
- Tubana, B.S.; Babu, T.; Datnoff, L.E. A Review of Silicon in Soils and Plants and Its Role in US Agriculture: History and Future Perspectives. Soil Sci. 2016, 181, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Epstein, E. SILICON. Annu. Rev. Plant Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef]
- E Richmond, K.; Sussman, M. Got silicon? The non-essential beneficial plant nutrient. Curr. Opin. Plant Biol. 2003, 6, 268–272. [Google Scholar] [CrossRef]
- Liang, Y.C.; Ma, T.S.; Li, F.J.; Feng, Y.J. Silicon availability and response of rice and wheat to silicon in calcareous soils. Commun. Soil Sci. Plant Anal. 1994, 25, 2285–2297. [Google Scholar] [CrossRef]
- Reynolds, O.L.; Padula, M.P.; Zeng, R.; Gurr, G.M. Silicon: Potential to Promote Direct and Indirect Effects on Plant Defense Against Arthropod Pests in Agriculture. Front. Plant Sci. 2016, 7, 744. [Google Scholar] [CrossRef] [Green Version]
- Epstein, E. The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.; Handreck, K. Silica In Soils, Plants, and Animals. Adv. Agron. 1967, 19, 107–149. [Google Scholar] [CrossRef]
- Raven, J.A. The Transport and Function of Silicon in Plants. Biol. Rev. 1983, 58, 179–207. [Google Scholar] [CrossRef]
- Gong, H.; Chen, K.; Chen, G.; Wang, S.-M.; Zhang, C. Effects of Silicon on Growth of Wheat Under Drought. J. Plant Nutr. 2003, 26, 1055–1063. [Google Scholar] [CrossRef]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2013, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Guntzer, F.; Keller, C.; Meunier, J.-D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon in Agriculture: From Theory to Practice; Springer: Berlin, Germany, 2015. [Google Scholar]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon Alleviates Drought Stress of Rice Plants by Improving Plant Water Status, Photosynthesis and Mineral Nutrient Absorption. Biol. Trace Element Res. 2010, 142, 67–76. [Google Scholar] [CrossRef]
- Hattori, T.; Inanaga, S.; Araki, H.; An, P.; Morita, S.; Luxova, M.; Lux, A. Application of silicon enhanced drought tolerance in Sorghum bicolor. Physiol. Plant. 2005, 123, 459–466. [Google Scholar] [CrossRef]
- Mukhtar, A.; Ummara, Q. Silicon application and drought tolerance mechanism of sorghum. Afr. J. Agric. Res. 2011, 6, 594–607. [Google Scholar] [CrossRef]
- Saud, S.; Li, X.; Chen, Y.; Zhang, L.; Fahad, S.; Hussain, S.; Sadiq, A.; Chen, Y. Silicon Application Increases Drought Tolerance of Kentucky Bluegrass by Improving Plant Water Relations and Morphophysiological Functions. Sci. World J. 2014, 2014, 368694. [Google Scholar] [CrossRef]
- Shen, X.; Zhou, Y.; Duan, L.; Li, Z.; Eneji, A.E.; Li, J. Silicon effects on photosynthesis and antioxidant parameters of soybean seedlings under drought and ultraviolet-B radiation. J. Plant Physiol. 2010, 167, 1248–1252. [Google Scholar] [CrossRef]
- Rodríguez, D.; Goudriaan, J.; Oyarzábal, M.; Pomar, M.C. Phosphorus nutrition and water stress tolerance in wheat plants. J. Plant Nutr. 1996, 19, 29–39. [Google Scholar] [CrossRef]
- Ma, J.F.; Goto, S.; Tamai, K.; Ichii, M. Role of Root Hairs and Lateral Roots in Silicon Uptake by Rice. Plant Physiol. 2001, 127, 1773–1780. [Google Scholar] [CrossRef]
- Zhan, A.; Schneider, H.; Lynch, J.P. Reduced Lateral Root Branching Density Improves Drought Tolerance in Maize. Plant Physiol. 2015, 168, 1603–1615. [Google Scholar] [CrossRef] [Green Version]
- Miao, B.-H.; Han, X.-G.; Zhang, W.-H. The ameliorative effect of silicon on soybean seedlings grown in potassium-deficient medium. Ann. Bot. 2010, 105, 967–973. [Google Scholar] [CrossRef] [Green Version]
- Luyckx, M.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Silicon and Plants: Current Knowledge and Technological Perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Munné-Bosch, S.; Llusià, J.; Filella, I. Leaf reflectance and photo- and antioxidant protection in field-grown summer-stressed Phillyrea angustifolia. Optical signals of oxidative stress? New Phytol. 2004, 162, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Song, A.; Li, P.; Fan, F.; Li, Z.; Liang, Y. The Effect of Silicon on Photosynthesis and Expression of Its Relevant Genes in Rice (Oryza sativa L.) under High-Zinc Stress. PLoS ONE 2014, 9, e113782. [Google Scholar] [CrossRef]
- Basso, A.S.; Miguez, F.E.; Laird, D.A.; Horton, R.; Westgate, M. Assessing potential of biochar for increasing water-holding capacity of sandy soils. GCB Bioenergy 2012, 5, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, H.M.; Al-Wabel, M.I.; Usman, A.R.A.; Al-Omran, A. Effect of Conocarpus Biochar Application on the Hydraulic Properties of a Sandy Loam Soil. Soil Sci. 2013, 178, 165–173. [Google Scholar] [CrossRef]
- Kammann, C.I.; Linsel, S.; Gößling, J.W.; Koyro, H.W. Influence of biochar on drought tolerance of Chenopodium quinoa Willd and on soil–plant relations. Plant Soil 2011, 345, 195–210. [Google Scholar] [CrossRef]
- Jeffery, S.; Verheijen, F.G.A.; van der Velde, M.; Bastos, A.C. A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Delta Agricultural Weather Center. Delta Research and Extension Center, Mississippi State University. Available online: http://ext.msstate.edu/anr/drec/weather.cgi. (accessed on 13 May 2022).
- Hewitt, E.J.; Bureaux, C.A.; Royal, F. Sand and Water Culture Methods Used in the Study of Plant Nutrition. In Commonwealth Bureau of Horticulture and Plantation Crops, East Malling. Technical Communication 22, 2nd ed.; 1966; Available online: https://agris.fao.org/agris-search/search.do?recordID=US201300126318 (accessed on 13 May 2022).
- Wijewardana, C.; Hock, M.; Henry, B.; Reddy, K.R. Screening Corn Hybrids for Cold Tolerance using Morphological Traits for Early-Season Seeding. Crop Sci. 2015, 55, 851–867. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | PH (cm) | NN (no.) | LA (cm2) | LDW (g) | SDW (g) | RDW (g) | SHDW (g) | RS | TDW (g) | RL (cm) | RSA (cm2) | RD (mm) | RV (cm3) | RT (no.) | RF (no.) | RC (no.) | CT (°C) | SPAD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | *** | NS | ** | NS | ** | * | * | NS | ** | NS | NS | NS | NS | *** | NS | NS | NS | NS |
| WS Trt | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | ** | ** |
| Si Trt | *** | ** | ** | NS | ** | * | NS | * | ** | * | * | *** | *** | *** | NS | ** | NS | NS |
| Cul X WS Trt | *** | NS | *** | ** | ** | ** | ** | *** | * | * | NS | NS | * | ** | NS | NS | NS | NS |
| Cul X Si Trt | NS | NS | NS | NS | NS | NS | NS | * | NS | NS | NS | * | NS | NS | NS | NS | NS | * |
| WS Trt X Si Trt | *** | NS | ** | NS | * | NS | NS | *** | NS | * | NS | NS | NS | *** | NS | *** | NS | NS |
| Cul X WS Trt X Si Trt | NS | NS | NS | NS | NS | NS | NS | ** | NS | NS | NS | NS | NS | NS | NS | NS | * | NS |
| Growth and Developmental Traits | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | PH (cm) | NN (no.) | LA (cm2) | LDW (g) | SDW (g) | RDW (g) | SHDW (g) | RS | TDW (g) | RL (cm) | RBN (no.) | RV (cm3) |
| Cultivar | *** | NS | * | ** | ** | NS | ** | NS | ** | NS | *** | NS |
| WS Trt | *** | *** | *** | *** | *** | NS | *** | *** | *** | NS | NS | NS |
| Cul X WS Trt | * | NS | NS | NS | NS | * | NS | ** | NS | NS | * | NS |
| Si Trt | ** | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS |
| Cul X Si Trt | NS | NS | NS | NS | NS | NS | NS | ** | NS | NS | NS | NS |
| WS Trt X Si Trt | NS | NS | NS | NS | NS | * | NS | ** | NS | NS | NS | NS |
| Cul X WS Trt X Si Trt | ** | ** | NS | * | NS | NS | * | * | NS | NS | NS | NS |
| Physiological traits | ||||||||||||
| Source of variation | Pnet (µmol m−2s−1) | Cond (mol m−2 s−1) | Ci/Ca | Fv’/Fm’ | ETR (µmol electron m−2s−1) | Trans (mmol m−2 s−1) | WUE (µmol CO2 mol−1 H2O) | |||||
| Cultivar | NS | NS | NS | * | NS | NS | NS | |||||
| WS Trt | NS | *** | *** | ** | * | *** | *** | |||||
| Cul X WS Trt | NS | * | NS | NS | * | NS | NS | |||||
| Si Trt | *** | ** | ** | *** | *** | *** | *** | |||||
| Cul X Si Trt | * | * | NS | NS | NS | NS | NS | |||||
| WS Trt X Si Trt | NS | * | *** | NS | NS | *** | *** | |||||
| Cul X WS Trt X Si Trt | NS | NS | NS | NS | * | NS | NS | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sah, S.K.; Reddy, K.R.; Li, J. Silicon Enhances Plant Vegetative Growth and Soil Water Retention of Soybean (Glycine max) Plants under Water-Limiting Conditions. Plants 2022, 11, 1687. https://doi.org/10.3390/plants11131687
Sah SK, Reddy KR, Li J. Silicon Enhances Plant Vegetative Growth and Soil Water Retention of Soybean (Glycine max) Plants under Water-Limiting Conditions. Plants. 2022; 11(13):1687. https://doi.org/10.3390/plants11131687
Chicago/Turabian StyleSah, Saroj Kumar, Kambham Raja Reddy, and Jiaxu Li. 2022. "Silicon Enhances Plant Vegetative Growth and Soil Water Retention of Soybean (Glycine max) Plants under Water-Limiting Conditions" Plants 11, no. 13: 1687. https://doi.org/10.3390/plants11131687