
The Effect of Thymus vulgaris L. Hydrolate Solutions on the Seed Germination, Seedling Length, and Oxidative Stress of Some Cultivated and Weed Species

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Seed Germination
2.1.1. Germination Percentage (GP)
2.1.2. Coefficient of Velocity of Germination (CVG)
2.1.3. Germination Index (GI) and Germination Rate Index (GRI)
2.1.4. Median Germination Time (t50)
2.1.5. Shoot and Root Length
2.2. Biochemical Parameters
2.2.1. MDA Content
2.2.2. SOD Activity
2.2.3. O2.-/Radicals
3. Discussion
4. Materials and Methods
4.1. Tested Plants
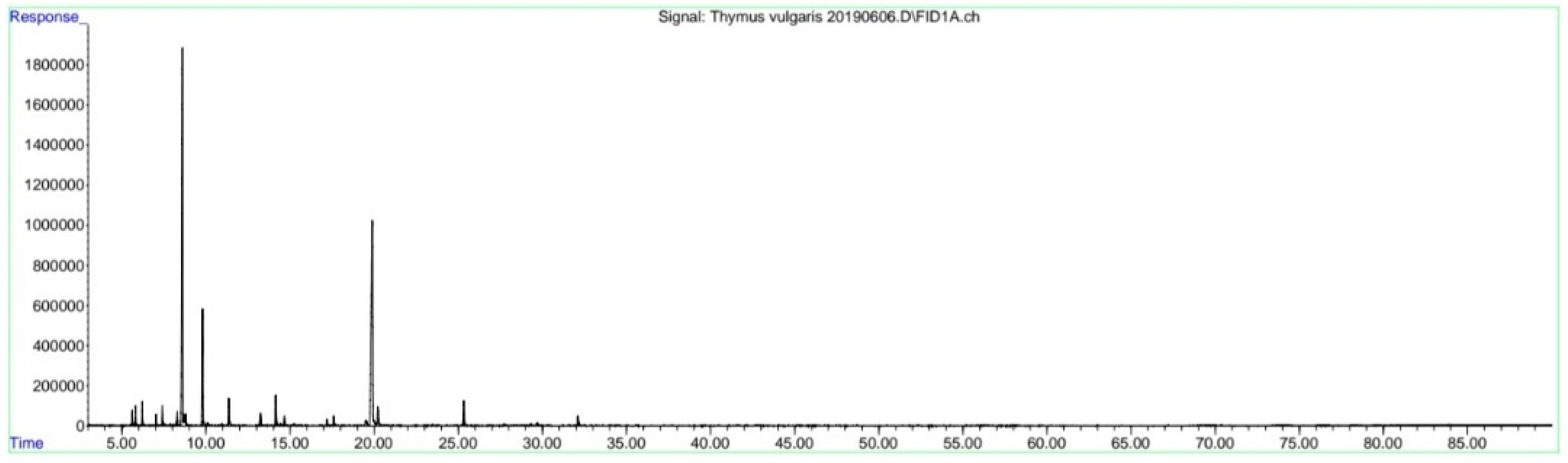
4.2. Hydrolate
4.3. Seed Germination and Seedlings Length
- Germination percentage (GP) [%] [53] represents the final germination percentage of the seed population and is calculated according to Equation (1):where Ng is the number of the germinated seeds and Nt is the total number of the seeds.
- Coefficient of velocity of germination (CVG) [% day−1] [54] represents the time required in order to reach the final germination percentage, and is calculated by Equation (2):where Ti is the time from the start of the experiment to the ith interval; Ni is the number of the seeds germinated in the ith interval (the number corresponding to the ith interval, not the accumulated number); and k is the total number of the intervals.
- Germination index (GI) [%] [55] reflects the germination speed; thus, a higher GI value indicates a faster germination rate. It is calculated according to Equation (3):where n1, n2…n10 represent the number of germinated seeds on the 1st, 2nd, and subsequent days until the 10th day; 10, 9…1 are the weights that are given to the number of germinated seeds on the 1st, 2nd, and subsequent days until the 10th day.
- Median germination time (t50) [time] [56] represents the time required in order to reach the 50% of the final germination and is calculated by Equation (4):where N is the final number of germinated seeds; Ni and Nj are the total number of seeds germinated in adjacent counts at time Ti and Tj, when Ni < < Nj.
- Germination rate index (GRI) (% day−1) [57] (after a modification) reflects the germination speed without distinguishing between the days with higher or lower germination since the percentage is evenly spread across the time frame. It is calculated according to Equation (5):where G1 is the germination percentage × 100 on the 1st day after sowing; G2 is the germination percentage × 100 on the 2nd day after sowing, etc., and Gx is the germination percentage × 100 on the xth day after sowing.
4.4. Biochemical Analysis of the Tested Plants
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aćimović, M.; Tešević, V.; Smiljanić, K.; Cvetković, M.; Stanković, J.; Kiprovski, B.; Sikora, V. Hydrolates–by-products of essential oil distillation: Chemical composition, biological activity and potential uses. Adv. Technol. 2020, 9, 54–70. [Google Scholar] [CrossRef]
- D’Amato, S.; Serio, A.; Chavez López, C.; Paparella, A. Hydrosols: Biological activity and potential as antimicrobials for food applications. Food Control 2018, 86, 126–137. [Google Scholar] [CrossRef]
- Andola, H.C.; Purohit, V.K.; Chauhan, R.S.; Arunachalam, K. Standardize quality standards for aromatic hydrosols. Med. Plants 2014, 6, 161–162. [Google Scholar] [CrossRef]
- Labadie, C.; Cerutti, C.; Carlin, F. Fate and control of pathogenic and spoilage micro-organisms in orange blossom (Citrus aurantium), and rose flower (Rosa centifolia) hydrosols. J. Appl. Microbiol. 2016, 121, 1568–1579. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V. Thymus vulgaris. In Medicinal Spices and Vegetables from Africa–Therapeutic Potential against Metabolic, Inflammatory, Infectious and Systemic Diseases; Kuete, V., Ed.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2017; p. 599. [Google Scholar] [CrossRef]
- Nabavi, S.M.; Marchese, A.; Izadi, M.; Curti, V.; Daglia, M.; Nabavi, S.F. Plants belonging to the genus Thymus as antibacterial agents: From farm to pharmacy. Food Chem. 2015, 173, 339–347. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Przychodna, M.; Sopata, S.; Bodalska, A.; Fecka, I. Thymol and thyme essential oil-new insights into selected therapeutic applications. Molecules 2020, 9, 4125. [Google Scholar] [CrossRef]
- Moisa, C.; Lupitu, A.; Pop, G.; Chambre, D.; Copolovici, L.; Cioca, G.; Bungau, S.G.; Copolovici, D.M. Variation of the chemical composition of Thymus vulgaris essential oils by phenological stage. Rev. Chim. 2019, 70, 633–637. [Google Scholar] [CrossRef]
- Hay, Y.O.; Abril-Sierra, M.A.; Sequeda-Castañeda, L.G.; Bonnafous, C.; Raynaud, C. Evaluation of combinations of essential oils and essential oils with hydrosols on antimicrobial and antioxidant activities. J. Pharm. Pharmacogn. Res. 2018, 6, 216–230. [Google Scholar]
- Rota, M.; Herrera, A.; Martínez, R.M.; Sotomayor, J.A.; Jordán, M.J. Antimicrobial activity and chemical composition of Thymus vulgaris, Thymus zygis and Thymus hyemalis essential oils. Food Control 2008, 19, 681–687. [Google Scholar] [CrossRef]
- Yildiz, S.; Turan, S.; Kiralan, M.; Ramadan, M.F. Antioxidant properties of thymol, carvacrol, and thymoquinone and its efficiencies on the stabilization of refined and stripped corn oils. J. Food Meas. Charact. 2021, 15, 621–632. [Google Scholar] [CrossRef]
- Beena; Kumar, D.; Rawat, D.S. Synthesis and antioxidant activity of thymol and carvacrol based Schiff bases. Bioorg. Med. Chem. Lett. 2013, 23, 641–645. [Google Scholar] [CrossRef] [PubMed]
- Guarda, A.; Rubilar, J.F.; Miltz, J.; Galotto, M.J. The antimicrobial activity of microencapsulated thymol and carvacrol. Int. J. Food Microbiol. 2011, 146, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Gavaric, N.; Smole Mozina, S.; Kladar, N.; Bozin, B. Chemical Profile, Antioxidant and Antibacterial Activity of Thyme and Oregano Essential Oils, Thymol and Carvacrol and Their Possible Synergism. J. Essent. Oil-Bear. Plants 2015, 18, 1013–1021. [Google Scholar] [CrossRef]
- Brito, C.; Hansen, H.; Espinoza, L.; Faúndez, M.; Olea, A.F.; Pino, S.; Díaz, K. Assessing the control of postharvest gray mold disease on tomato fruit using mixtures of essential oils and their respective hydrolates. Plants 2021, 10, 1719. [Google Scholar] [CrossRef] [PubMed]
- Andrés, M.F.; González-Coloma, A.; Muñoz, R.; De la Peña, F.; Fernando, J.L.; Burillo, J. Nematicidal potential of hydrolates from the semi industrial vapor-pressure extraction of Spanish aromatic plants. Environ. Sci. Pollut. Res. 2018, 25, 29834–29840. [Google Scholar] [CrossRef]
- Sardari, S.; Mobaien, A.; Ghassemifard, L.; Kamali, K.; Khavasi, N. Therapeutic effect of thyme (Thymus vulgaris) essential oil on patients with COVID19: A randomized clinical trial. J. Adv. Med. Biomed. Res. 2021, 29, 83–91. [Google Scholar] [CrossRef]
- Rezatofighi, S.E.; Seydabadi, A.; Seyyed Nejad, S.M. Evaluating the efficacy of Achillea millefolium and Thymus vulgaris extracts against newcastle disease virus in Ovo. Jundishapur J. Microbiol. 2014, 7, e9016. [Google Scholar] [CrossRef] [Green Version]
- Lelešius, R.; Karpovaitė, A.; Mickienė, R.; Drevinskas, T.; Tiso, N.; Ragažinskienė, O.; Kubilienė, L.; Maruška, A.; Šalomskas, A. In vitro antiviral activity of fifteen plant extracts against avian infectious bronchitis virus. BMC Vet. Res. 2019, 29, 178. [Google Scholar] [CrossRef] [Green Version]
- Vimalanathan, S.; Hudson, J. Anti-influenza virus activity of essential oils and vapors. Am. J. Essent. 2014, 2, 47–53. [Google Scholar]
- Feriotto, G.; Marchetti, N.; Costa, V.; Beninati, S.; Tagliati, F.; Mischiati, C. Chemical composition of essential oils from Thymus vulgaris, Cymbopogon citratus, and Rosmarinus officinalis, and their effects on the HIV-1 Tat protein function. Chem. Biodivers. 2018, 15, e1700436. [Google Scholar] [CrossRef]
- Catella, C.; Camero, M.; Lucente, S.; Fracchiolla, G.; Sblano, S.; Tempesta, M.; Martella, V.; Buonavoglia, C.; Lanave, G. Virucidal and antiviral effects of Thymus vulgaris essential oil on feline coronavirus. Res. Vet. Sci. 2021, 137, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Kaewprom, K.; Chen, Y.H.; Lin, C.F.; Chiou, M.T.; Lin, C.N. Antiviral activity of Thymus vulgaris and Nepeta cataria hydrosols against porcine reproductive and respiratory syndrome virus. Thai J. Vet. Med. 2017, 47, 25–33. [Google Scholar]
- Sağdiç, O.; Ozturk, I.; Tornuk, F. Inactivation of non-toxigenic and toxigenic Escherichia coli O157:H7 inoculated on minimally processed tomatoes and cucumbers: Utilization of hydrosols of Lamiaceae spices as natural food sanitizers. Food Control 2013, 30, 7–14. [Google Scholar] [CrossRef]
- Tornuk, F.; Cankurt, H.; Ozturk, I.; Sagdic, O.; Bayram, O.; Yetim, H. Efficacy of various plant hydrosols as natural food sanitizers in reducing Escherichia coli O157:H7 and Salmonella typhimurium on fresh cut carrots and apples. Int. J. Food Microbiol. 2011, 148, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, I.; Tornuk, F.; Caliskan-Aydogan, O.; Durak, M.Z.; Sagdic, O. Decontamination of iceberg lettuce by some plant hydrosols. LWT 2016, 74, 48–54. [Google Scholar] [CrossRef]
- Varel, V.H. Carvacrol and thymol reduce swine waste odor and pathogens: Stability of oils. Curr. Microbiol. 2002, 44, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Chen, B.; Zhang, J.; Wang, P.; Feng, G. The environmental fate of thymol, a novel botanical pesticide, in tropical agricultural soil and water. Toxicol. Environ. Chem. 2017, 99, 223–232. [Google Scholar] [CrossRef]
- Azirak, S.; Karaman, S. Allelopathic effect of some essential oils and components on germination of weed species. Acta Agric. Scand.-B Soil Plant Sci. 2008, 58, 88–92. [Google Scholar] [CrossRef]
- de Assis Alves, T.; Fontes Pinheiro, P.; Praça-Fontes, M.M.; Andrade-Vieira, L.F.; Barelo Corrêa, K.; de Assis Alves, T.; da Cruz, F.A.; Lacerda Júnior, V.; Ferreira, A.; Bastos Soares, T.C. Toxicity of thymol, carvacrol and their respective phenoxyacetic acids in Lactuca sativa and Sorghum bicolor. Ind. Crops Prod. 2018, 114, 59–67. [Google Scholar] [CrossRef]
- Balboul, T.; Ophir, A.; Dotan, A. Essential oils as natural root-repellent herbicides for drip irrigation systems. Polym. Adv. Technol. 2022, 1–10. [Google Scholar] [CrossRef]
- Pinheiro, P.F.; Costa, A.V.; de Assis Alves, T.; Galter, I.N.; Pinheiro, C.A.; Pereira, A.F.; Oliveira, C.M.; Fontes, M.M. Phytotoxicity and cytotoxicity of essential oil from leaves of Plectranthus amboinicus, carvacrol, and thymol in plant bioassays. J. Agric. Food Chem. 2015, 63, 8981–8990. [Google Scholar] [CrossRef] [PubMed]
- Kordali, S.; Cakir, A.; Ozer, H.; Cakmakci, R.; Kesdek, M.; Mete, E. Antifungal, phytotoxic and insecticidal properties of essential oil isolated from Turkish Origanum acutidens and its three components, carvacrol, thymol and p-cymene. Bioresour. Technol. 2008, 99, 8788–8795. [Google Scholar] [CrossRef] [PubMed]
- Marco, C.A.; Teixeira, E.; Simplício, A.; Oliveira, C.; Costa, J.; Feitosa, J. Chemical composition and allelopathyc activity of essential oil of Lippia sidoides Cham. Chil. J. Agric. Res. 2021, 72, 157–160. [Google Scholar] [CrossRef] [Green Version]
- Hemada, M.; El-Darier, S. Comparative study on composition and biological activity of essential oils of two Thymus species grown in Egypt. Am.-Eurasian J. Agric. Environ. Sci. 2011, 11, 647–654. [Google Scholar]
- Arouiee, H.; Ghasemimohsenabad, S.; Azizi, M.; Neamati, S.H. Allelopathic effects of some medicinal plants extracts on seed germination and growth of common weeds in Mashhad Area. In Proceedings of the 8th International Symposium on Biocontrol and Biotechnology, Pattaya, Thailand, 4–6 October 2010. [Google Scholar]
- Linhart, Y.B.; Gauthier, P.; Keefover-Ring, K.; Thompson, J.D. Variable phytotoxic effects of Thymus vulgaris (Lamiaceae) terpenes on associated species. Int. J. Plant Sci. 2015, 176, 20–30. [Google Scholar] [CrossRef]
- Sadeghi, H.; Khazaei, F.; Sheidaei, S.; Yari, L. Effect of seed size on seed germination behavior of safflower (Carthamus tinctorius L.). J. Agric. Biol. Sci. 2011, 6, 5–8. [Google Scholar]
- Moles, A.T.; Westoby, M. Seed size and plant strategy across the whole life cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Salamci, E.; Kordali, S.; Kotan, R.; Cakir, A.; Kaya, Y. Chemical compositions, antimicrobial and herbicidal effects of essential oils isolated from Turkish Tanacetum aucheranum and Tanacetum chiliophyllum var. chiliophyllum. Biochem. Syst. Ecol. 2007, 35, 569–581. [Google Scholar] [CrossRef]
- Ibáñez, M.D.; Blázquez, M.A. Phytotoxicity of essential oils on selected weeds: Potential hazard on food crops. Plants 2018, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Verdeguer, M.; Torres-Pagan, N.; Muñoz, M.; Jouini, A.; García-Plasencia, S.; Chinchilla, P.; Berbegal, M.; Salamone, A.; Agnello, S.; Carrubba, A.; et al. Herbicidal activity of Thymbra capitata (L.) Cav. essential oil. Molecules 2020, 25, 2832. [Google Scholar] [CrossRef]
- Saad, M.; Gouda, N.; Abdelgaleil, S. Bioherbicidal activity of terpenes and phenylpropenes against Echinochloa crus-galli. J. Environ. Sci. Health-B Pestic. Food Contam. Agric. Wastes 2019, 54, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Matković, A.; Marković, T.; Vrbničanin, S.; Sarić-Krsmanović, M.; Božić, D. Chemical composition and in vitro herbicidal activity of five essential oils on seeds of Johnson grass (Sorghum halepense L. Pers.). Lek. Sirovine 2018, 38, 44–50. [Google Scholar] [CrossRef]
- Zunino, M.P.; Zygadlo, J.A. Effect of monoterpenes on lipid oxidation in maize. Planta 2004, 219, 303–309. [Google Scholar] [CrossRef]
- De Martino, L.; Mancini, E.; Rolim de Almeida, L.F.; De Feo, V. The antigerminative activity of twenty-seven monoterpenes. Molecules 2010, 15, 6630–6637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelini, L.G.; Carpanese, G.; Cioni, P.L.; Morelli, I.; Macchia, M.; Flamini, G. Essential oils from Mediterranean Lamiaceae as weed germination inhibitors. J. Agric. Food Chem. 2003, 51, 6158–6164. [Google Scholar] [CrossRef] [PubMed]
- Šućur, J.; Konstantinović, B.; Crnković, M.; Bursić, V.; Samardžić, N.; Malenčić, Đ.; Prvulović, D.; Popov, M.; Vuković, G. Chemical composition of Ambrosia trifida L. and its allelopathic influence on crops. Plants 2021, 10, 2222. [Google Scholar] [CrossRef]
- Aćimović, M.; Cvetković, M.; Stanković Jeremić, J.; Pezo, L.; Varga, A.; Čabarkapa, I.; Kiprovski, B. Biological activity and profiling of Salvia sclarea essential oil obtained by steam and hydrodistillation extraction methods via chemometrics tools. Flavour Fragr. J. 2022, 37, 20–32. [Google Scholar] [CrossRef]
- Aćimović, M.; Lončar, B.; Stanković Jeremić, J.; Cvetković, M.; Pezo, L.; Pezo, M.; Todosijević, M.; Tešević, V. Weather conditions influence on lavandin essential oil and hydrolate quality. Horticulturae 2022, 8, 281. [Google Scholar] [CrossRef]
- Voll, E.; Voll, C.E.; Filho, R.V. Allelopathic effects of aconitic acid on wild poinsettia (Euphorbia heterophy L. La) and morningglory (Ipomoea grandifo Lia). J. Environ. Sci. Health-B 2005, 40, 69–75. [Google Scholar] [CrossRef]
- Marinov-Serafimov, P.; Dimitrova, T.S.; Golubinova, I.; Ilieva, A. Study of suitability of some solutions in allelopathic researches. Herbologia 2007, 8, 1–10. [Google Scholar]
- ISTA-International Seed Testing Association. International rules for seed testing. Seed Sci. Technol. 1993, 21, 142–168. [Google Scholar]
- Jones, K.W.; Sanders, D. The influence of soaking paper seed in water or potassium salt solutions on germination at three temperatures. J. Seed Technol. 1987, 11, 97–102. [Google Scholar] [CrossRef]
- Bench, A.R.; Fenner, M.; Edwards, P. Changes in germinability, ABA content and ABA embryonic sensitivity in developing seeds of Sorghum bicolor (L.) Moench induced by water stress during grain filling. New Phytol. 1991, 118, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Basra, S.M.A.; Ahmad, N.; Hafeez, K. Thermal hardening: A new seed vigor enhancement tool in rice. J. Integr. Plant Biol. 2005, 47, 187–193. [Google Scholar] [CrossRef]
- Esechie, H. Interaction of salinity and temperature on the germination of sorghum. J. Agron. Crop Sci. 1994, 172, 194–199. [Google Scholar] [CrossRef]
- Mandal, S.; Mitra, A.; Mallick, N. Biochemical characterization of oxidative burst during interaction between Solanum lycopersicum and Fusarium oxysporum f. sp. lycopersici. Physiol. Mol. Plant Pathol. 2008, 72, 56–61. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Tested Plant | Concentration | GP (%) | CVG (% Day−1) | GI (−) | t50 (Time) | GRI (%/Day) |
|---|---|---|---|---|---|---|
| soybean | control | 93 | 23.48 | 627 | 2.78 | 33.94 |
| 10% | 74 | 30.58 | 572 | 2.48 | 31.41 | |
| 20% | 71 | 28.18 | 529 | 2.48 | 27.82 | |
| 50% | 66 | 30.56 | 510 | 2.39 | 27.21 | |
| 100% | 60 | 24.00 | 410 | 2.77 | 19.21 | |
| maize | control | 97 | 22.35 | 633 | 4.15 | 27.45 |
| 10% | 80 | 19.66 | 473 | 4.80 | 19.36 | |
| 20% | 82 | 18.34 | 455 | 4.56 | 18.13 | |
| 50% | 65 | 18.36 | 361 | 4.69 | 15.81 | |
| 100% | 21 | 23.33 | 141 | 3.25 | 8.71 | |
| onion | control | 85 | 23.94 | 580 | 3.69 | 26.29 |
| 10% | 75 | 19.74 | 445 | 4.07 | 16.42 | |
| 20% | 33 | 17.94 | 179 | 4.56 | 6.30 | |
| 50% | 21 | 17.95 | 114 | 4.59 | 4.07 | |
| 100% | 10 | 13.51 | 36 | 7.00 | 1.42 | |
| sunflower | control | 89 | 33.21 | 711 | 1.81 | 44.89 |
| 10% | 96 | 34.78 | 780 | 1.77 | 49.72 | |
| 20% | 88 | 19.43 | 515 | 4.18 | 18.13 | |
| 50% | 83 | 21.01 | 518 | 3.81 | 18.42 | |
| 100% | 73 | 25.70 | 519 | 2.68 | 23.79 | |
| alfalfa | control | 83 | 35.78 | 681 | 2.17 | 36.08 |
| 10% | 85 | 32.69 | 675 | 2.18 | 35.73 | |
| 20% | 76 | 23.03 | 506 | 2.76 | 24.99 | |
| clover | control | 93 | 37.65 | 776 | 2.29 | 41.29 |
| 10% | 95 | 38.00 | 791 | 2.28 | 42.34 | |
| 20% | 90 | 23.87 | 613 | 2.90 | 30.06 | |
| carrot | control | 39 | 16.46 | 192 | 4.85 | 7.09 |
| 10% | 14 | 11.76 | 35 | 8.20 | 1.70 | |
| 20% | 1 | 12.50 | 3 | 7.50 | 0.12 | |
| wheat | control | 100 | 43.10 | 868 | 1.73 | 52.47 |
| 10% | 89 | 18.39 | 495 | 4.36 | 18.22 | |
| 20% | 66 | 22.53 | 433 | 3.65 | 17.26 | |
| 50% | 9 | 20.93 | 56 | 4.17 | 2.14 |
| Tested Plant | Concentration | GP (%) | CVG (% Day−1) | GI (−) | t50 (Time) | GRI (%/Day) |
|---|---|---|---|---|---|---|
| red-root amaranth | control | 83 | 24.78 | 578 | 2.56 | 30.41 |
| 10% | 26 | 14.86 | 111 | 6.29 | 4.03 | |
| cockspur | control | 22 | 19.47 | 129 | 3.92 | 4.60 |
| 10% | 20 | 15.15 | 88 | 6.33 | 3.30 | |
| 20% | 9 | 15.25 | 40 | 5.75 | 1.47 | |
| 50% | 4 | 17.39 | 21 | 5.33 | 0.71 | |
| common purslane | control | 100 | 38.46 | 840 | 1.88 | 51.41 |
| 10% | 46 | 22.77 | 304 | 3.88 | 13.86 | |
| Johnson grass | control | 23 | 13.30 | 80 | 7.50 | 3.35 |
| 10% | 17 | 12.06 | 46 | 7.90 | 2.10 | |
| 20% | 6 | 11.76 | 15 | 8.00 | 0.71 | |
| 50% | 1 | 11.11 | 2 | 8.50 | 0.11 | |
| common lambsquarters | control | 51 | 20.00 | 306 | 4.19 | 12.95 |
| 10% | 47 | 18.58 | 264 | 4.43 | 9.91 | |
| 20% | 1 | 14.29 | 4 | 6.50 | 0.14 | |
| black nightshade | control | 84 | 13.25 | 290 | 7.68 | 12.16 |
| 10% | 100 | 14.14 | 393 | 6.47 | 15.13 | |
| 20% | 95 | 14.80 | 403 | 6.34 | 15.27 | |
| 50% | 14 | 11.57 | 33 | 8.25 | 1.64 |
| Tested Plant | Variable | Shoot Length (mm) | Root Length (mm) |
|---|---|---|---|
| onion | control | 13.70 ± 10.15 a | 7.65 ± 6.40 a |
| 10% | 6.59 ± 5.69 b | 2.86 ± 2.80 b | |
| 20% | 0.85 ± 1.98 c | 0.59 ± 1.44 c | |
| 50% | 1.08 ± 1.64 c | 0.60 ± 0.98 c | |
| 100% | 0.17 ± 0.53 c | 0.17 ± 0.53 c | |
| maize | control | 25.26 ± 12.87 a | 69.29 ± 25.76 a |
| 10% | 19.49 ± 17.00 b | 54.31 ± 49.06 b | |
| 20% | 13.80 ± 14.75 c | 29.66 ± 28.76 c | |
| 50% | 10.78 ± 14.81 c | 20.09 ± 27.81 d | |
| 100% | 4.87 ± 10.05 d | 5.53 ± 13.48 e | |
| sunflower | control | 34.41 ± 21.64 a | 50.01 ± 38.16 b |
| 10% | 38.00 ± 19.68 a | 64.51 ± 46.27 a | |
| 20% | 29.31 ± 23.66 b | 40.90 ± 41.06 b | |
| 50% | 9.47 ± 7.91 c | 13.18 ± 10.70 c | |
| 100% | 7.32 ± 6.54 c | 12.26 ± 11.13 c | |
| soybean | control | 27.87 ± 13.03 a | 13.92 ± 8.65 b |
| 10% | 14.76 ± 11.18 b | 19.91 ± 19.83 a | |
| 20% | 12.13 ± 9.80 b | 18.56 ± 18.65 a | |
| 50% | 8.94 ± 7.37 c | 11.70 ± 12.40 b | |
| 100% | 7.70 ± 7.41 c | 6.28 ± 6.61 c | |
| wheat | control | 114.12 ± 19.17 a | 114.11 ± 19.14 a |
| 10% | 30.28 ± 26.61 b | 40.36 ± 34.36 b | |
| 20% | 22.47 ± 26.56 c | 22.92 ± 26.09 c | |
| 50% | 3.72 ± 13.82 d | 2.63 ± 9.69 d | |
| carrot | control | 3.65 ± 7.38 a | 4.32 ± 6.95 a |
| 10% | 0.37 ± 1.07 b | 1.65 ± 4.54 b | |
| 20% | 0.04 ± 0.40 b | 0.04 ± 0.40 c | |
| white clover | control | 12.74 ± 4.99 a | 31.85 ± 16.72 a |
| 10% | 9.66 ± 3.47 b | 25.64 ± 10.86 b | |
| 20% | 6.99 ± 3.26 c | 17.17 ± 10.18 c | |
| alfalfa | control | 12.08 ± 7.71 a | 28.51 ± 19.45 a |
| 10% | 10.35 ± 5.35 b | 27.29 ± 17.64 a | |
| 20% | 7.50 ± 5.44 c | 12.46 ± 11.92 b |
| Tested Plant | Variable | Shoot Length (mm) | Root Length (mm) |
|---|---|---|---|
| common purslane | control | 10.49 ± 2.55 a | 19.76 ± 4.53 a |
| 10% | 1.54 ± 1.78 b | 1.71 ± 2.01 b | |
| red-root amaranth | control | 15.61 ± 8.32 a | 14.43 ± 7.72 a |
| 10% | 1.81 ± 3.29 b | 1.96 ± 3.41 b | |
| common lambsquarters | control | 4.12 ± 5.63 a | 7.65 ± 12.77 a |
| 10% | 2.07 ± 3.21 b | 1.89 ± 2.46 b | |
| 20% | - | 0.05 ± 0.50 c | |
| cockspur grass | control | 8.90 ± 17.75 a | 10.70 ± 23.20 a |
| 10% | 6.13 ± 13.74 ab | 4.70 ± 11.07 bc | |
| 20% | 3.08 ± 10.33 bc | 1.47 ± 5.88 b | |
| Johnson grass | control | 10.48 ± 21.54 a | 10.38 ± 22.58 a |
| 10% | 2.38 ± 9.94 bc | 1.78 ± 7.33 bc | |
| 20% | 5.52 ± 15.09 b | 4.81 ± 13.25 b | |
| 50% | 0.42 ± 2.96 c | - | |
| black nightshade | control | 2.57 ± 5.48 c | 9.69 ± 19.86 c |
| 10% | 10.96 ± 3.80 a | 43.88 ± 13.68 a | |
| 20% | 8.77 ± 4.60 b | 19.64 ± 15.25 b | |
| 50% | 1.22 ± 4.17 d | 2.97 ± 9.20 d |
| Tested Plant | Variable | LP nmol/g FW | SOD U/g FW | nmol O2.-//g FW |
|---|---|---|---|---|
| wheat | control | 69.40 ± 16.60 a | 119.09 ± 0.81 a | 58.01 ± 4.06 a |
| 10% | 67.91 ± 8.23 a | 124.46 ± 0.38 b | 89.84 ± 25.86 a | |
| 20% | 48.09 ± 4.86 a | 122.96 ± 0.50 b | 252.08 ± 63.13 ab | |
| 50% | 55.27 ± 8.72 a | 125.39 ± 1.31 b | 319.63 ± 103.44 b | |
| maize | control | 62.00 ± 1.90 a | 120.44 ± 0.14 d | 15.83 ± 3.88 a |
| 10% | 38.66 ± 4.75 a | 118.91 ± 0.21 bc | 34.25 ± 9.08 bc | |
| 20% | 69.25 ± 27.45 a | 119.53 ± 0.42 c | 19.66 ± 3.71 ab | |
| 50% | 63.19 ± 23.16 a | 118.35 ± 0.07 b | 46.38 ± 1.21 c | |
| 100% | 49.51 ± 8.59 a | 116.40 ± 0.00 a | 5.11 ± 0.63 a | |
| sunflower | control | 52.20 ± 5.65 a | 82.66 ± 0.57 c | 57.63 ± 6.15 a |
| 10% | 42.03 ± 10.17 a | 50.60 ± 16.59 ab | 115.60 ± 7.12 b | |
| 20% | 43.00 ± 6.69 a | 33.85 ± 11.59 a | 56.08 ± 16.80 a | |
| 50% | 41.13 ± 4.83 a | 67.20 ± 5.48 bc | 76.26 ± 18.75 ab | |
| 100% | 37.24 ± 6.66 a | 71.64 ± 0.26 bc | 99.39 ± 5.96 b | |
| soybean | control | 95.20 ± 4.59 a | 107.96 ± 0.86 c | 22.38 ± 1.15 ab |
| 10% | 96.92 ± 8.93 a | 24.91 ± 0.99 b | 30.09 ± 1.71 bc | |
| 20% | 97.15 ± 16.08 a | 23.01 ± 4.43 b | 18.21 ± 2.40 a | |
| 50% | 86.53 ± 8.75 a | 10.68 ± 1.05 a | 33.75 ± 5.81 c | |
| 100% | 89.89 ± 24.37 a | 5.69 ± 2.94 a | 19.71 ± 2.77 ab | |
| onion | control | 36.87 ± 5.63 a | 111.58 ± 2.46 a | 90.44 ± 42.56 a |
| 10% | 37.92 ± 8.46 a | 97.41 ± 2.74 a | 290.53 ± 25.37 a | |
| 20% | 35.17 ± 5.62 a | 116.34 ± 14.87 a | 817.26 ± 117.71 b | |
| carrot | control | 18.03 ± 2.20 a | 99.19 ± 6.91 a | 60.83 ± 14.08 a |
| 10% | 18.54 ± 0.81 a | 150.95 ± 14.96 b | 165.92 ± 33.20 b | |
| white clover | control | 43.73 ± 8.47 a | 23.46 ± 2.68 c | 46.48 ± 6.47 b |
| 10% | 38.65 ± 3.78 a | 8.72 ± 0.95 b | 25.45 ± 2.64 a | |
| 20% | 60.90 ± 10.18 a | 2.47 ± 0.63 a | 30.07 ± 4.56 ab | |
| alfalfa | control | 97.48 ± 20.25 ab | 188.94 ± 1.74 b | 17.77 ± 7.08 a |
| 10% | 51.35 ± 11.12 a | 175.61 ± 0.78 a | 24.64 ± 7.01 a | |
| 20% | 115.74 ± 19.05 b | 282.83 ± 1.20 c | 49.91 ± 6.47 b |
| Tested Plant | Variable | LP nmol/g FW | SOD U/g FW | nmol O2.-//g FW |
|---|---|---|---|---|
| common lambsquarters | control | 18.48 ± 2.38 a | 79.52 ± 2.91 a | 154.49 ± 8.15 a |
| 10% | 165.73 ± 0.67 b | 334.59 ± 95.64 a | 3366.46 ± 412.48 b | |
| red-root amaranth | control | 43.88 ± 2.81 b | 80.56 ± 3.06 a | 77.48 ± 1.87 a |
| 10% | 31.41 ± 2.42 a | 101.83 ± 10.51 a | 82.48 ± 19.03 a | |
| cockspur grass | control | 37.81 ± 1.41 a | 142.56 ± 7.65 a | 151.52 ± 45.71 a |
| 10% | 32.90 ± 0.20 a | 126.47 ± 8.79 a | 256.70 ± 89.31 a | |
| 20% | 83.96 ± 3.75 b | 313.76 ± 9.55 b | 536.88 ± 20.80 b | |
| black nightshade | control | 48.92 ± 4.32 a | 84.67 ± 2.40 a | 305.66 ± 126.34 a |
| 10% | 48.46 ± 9.95 a | 236.92 ± 58.78 b | 1105.43 ± 9.45 a | |
| 20% | 30.48 ± 2.91 a | 179.82 ± 3.76 ab | 644.68 ± 460.14 a | |
| 50% | 35.23 ± 6.61 a | 100.55 ± 2.79 a | 420.61 ± 85.40 a | |
| Johnson grass | control | 37.40 ± 1.77 ab | 77.59 ± 2.43 a | 224.38 ± 14.53 a |
| 10% | 33.36 ± 0.12 a | 166.76 ± 57.79 ab | 241.64 ± 46.46 a | |
| 20% | 74.51 ± 2.82 c | 337.81 ± 31.68 c | 731.04 ± 290.72 a | |
| 50% | 43.63 ± 2.98 b | 228.08 ± 3.93 b | 338.85 ± 8.69 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konstantinović, B.; Popov, M.; Samardžić, N.; Aćimović, M.; Šućur Elez, J.; Stojanović, T.; Crnković, M.; Rajković, M. The Effect of Thymus vulgaris L. Hydrolate Solutions on the Seed Germination, Seedling Length, and Oxidative Stress of Some Cultivated and Weed Species. Plants 2022, 11, 1782. https://doi.org/10.3390/plants11131782
Konstantinović B, Popov M, Samardžić N, Aćimović M, Šućur Elez J, Stojanović T, Crnković M, Rajković M. The Effect of Thymus vulgaris L. Hydrolate Solutions on the Seed Germination, Seedling Length, and Oxidative Stress of Some Cultivated and Weed Species. Plants. 2022; 11(13):1782. https://doi.org/10.3390/plants11131782
Chicago/Turabian StyleKonstantinović, Bojan, Milena Popov, Nataša Samardžić, Milica Aćimović, Jovana Šućur Elez, Tijana Stojanović, Marina Crnković, and Miloš Rajković. 2022. "The Effect of Thymus vulgaris L. Hydrolate Solutions on the Seed Germination, Seedling Length, and Oxidative Stress of Some Cultivated and Weed Species" Plants 11, no. 13: 1782. https://doi.org/10.3390/plants11131782