
Response Mechanisms of “Hass” Avocado to Sequential 1–methylcyclopropene Applications at Different Maturity Stages during Cold Storage

 , ,
, ,
Abstract
:1. Introduction
2. Results
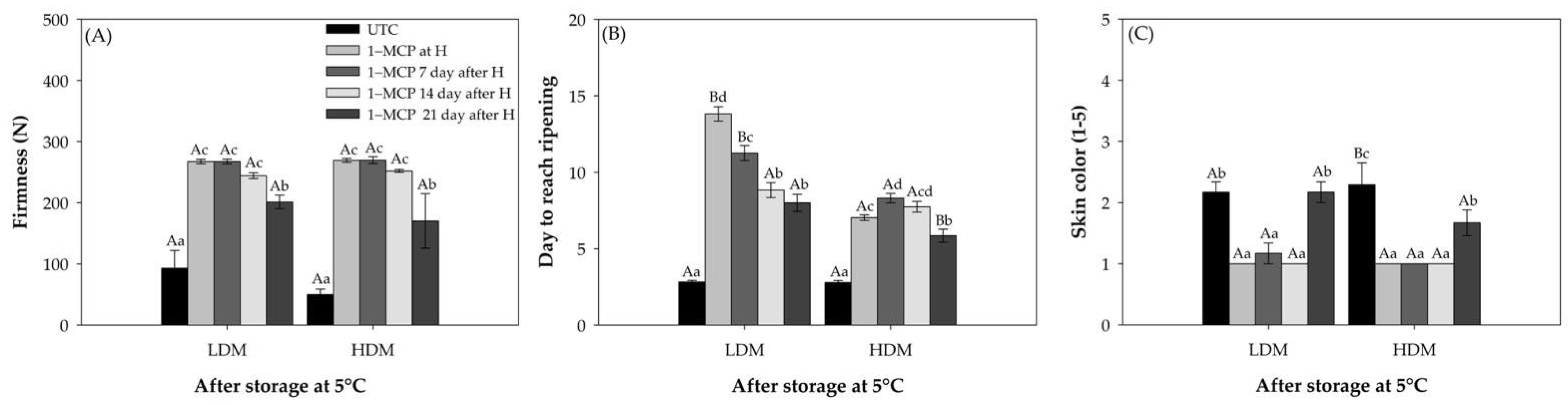
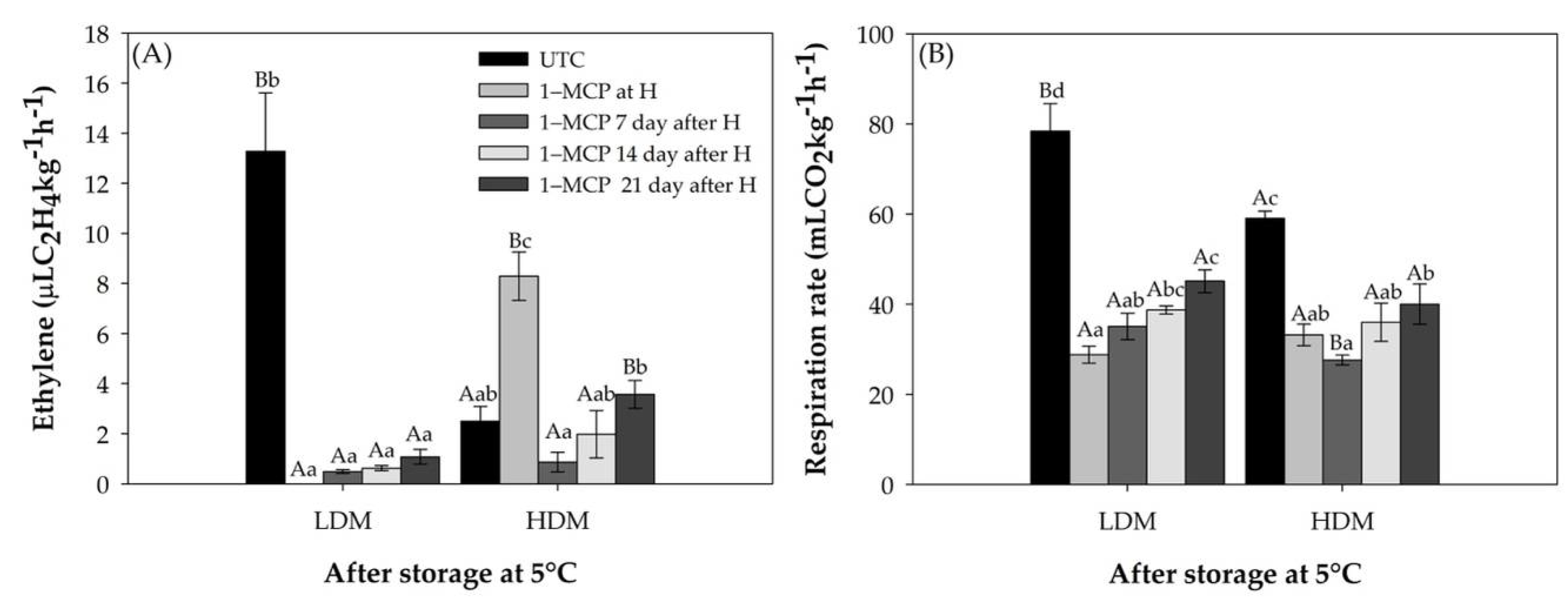
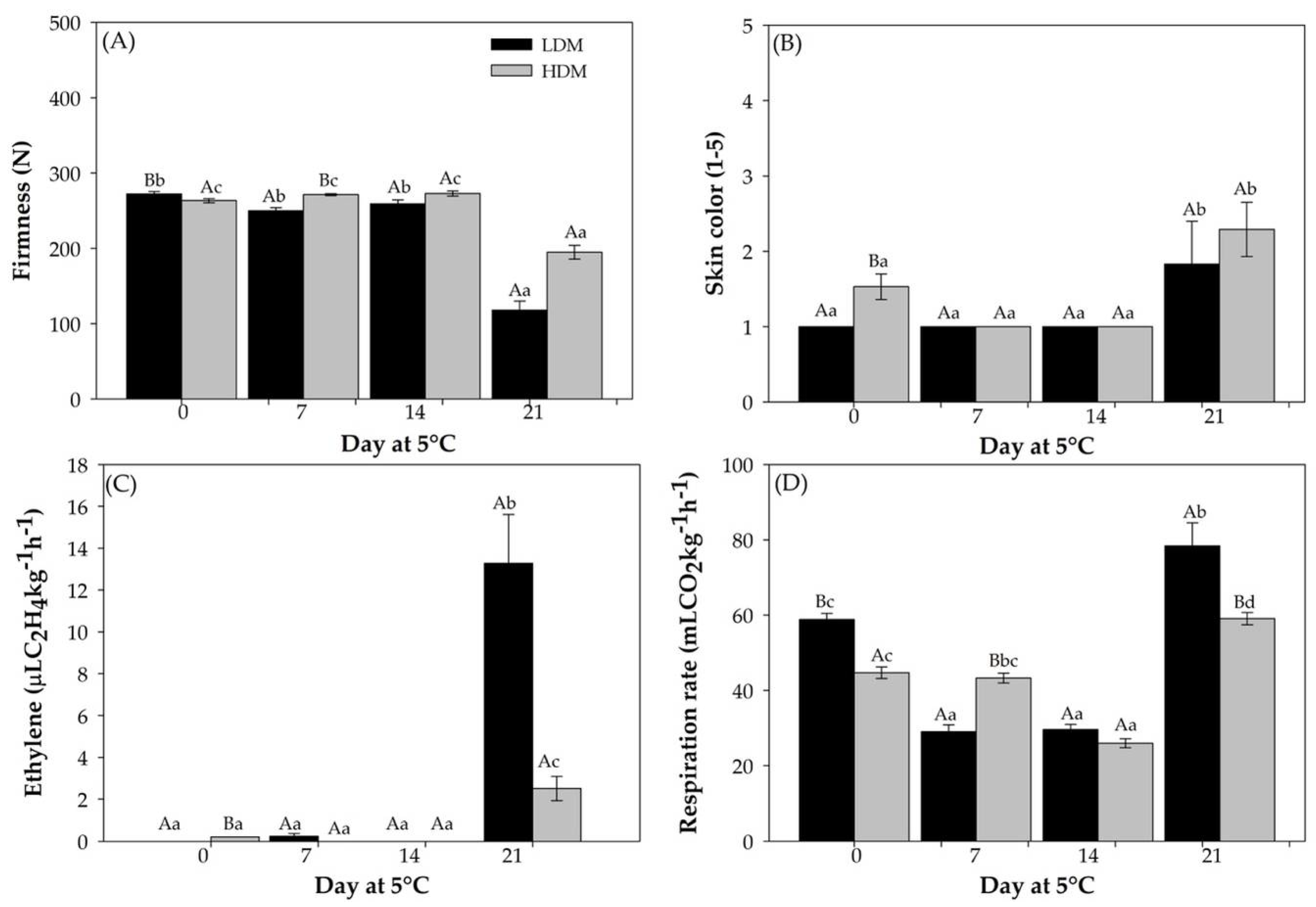
2.1. Ripening Related Physiological Parameters
2.1.1. Ripening Parameters
2.1.2. Physiological Parameters
2.2. Fruit Characteristics at the 1–MCP Application Time
2.2.1. Ripening Parameters
2.2.2. Physiological Parameters
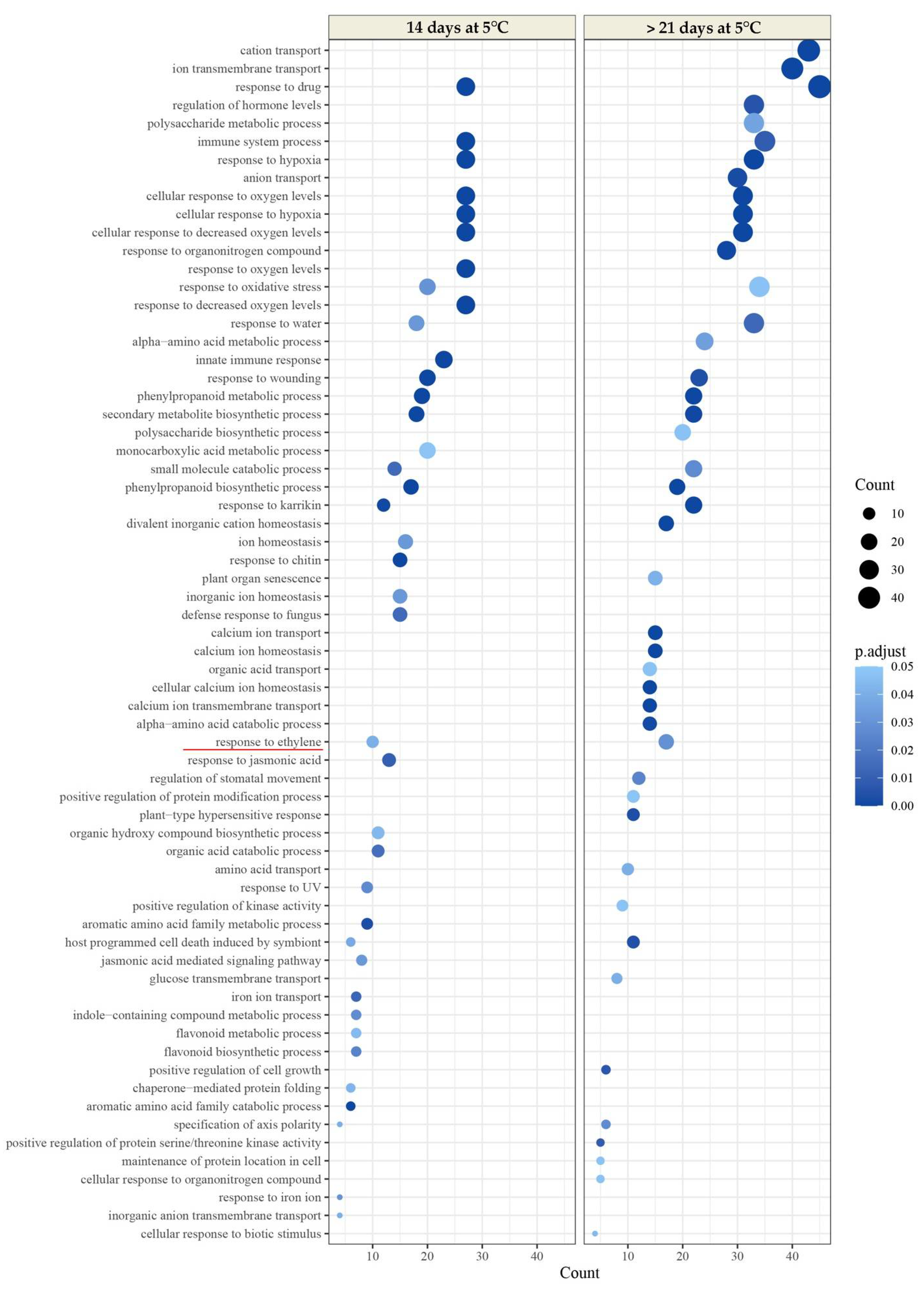
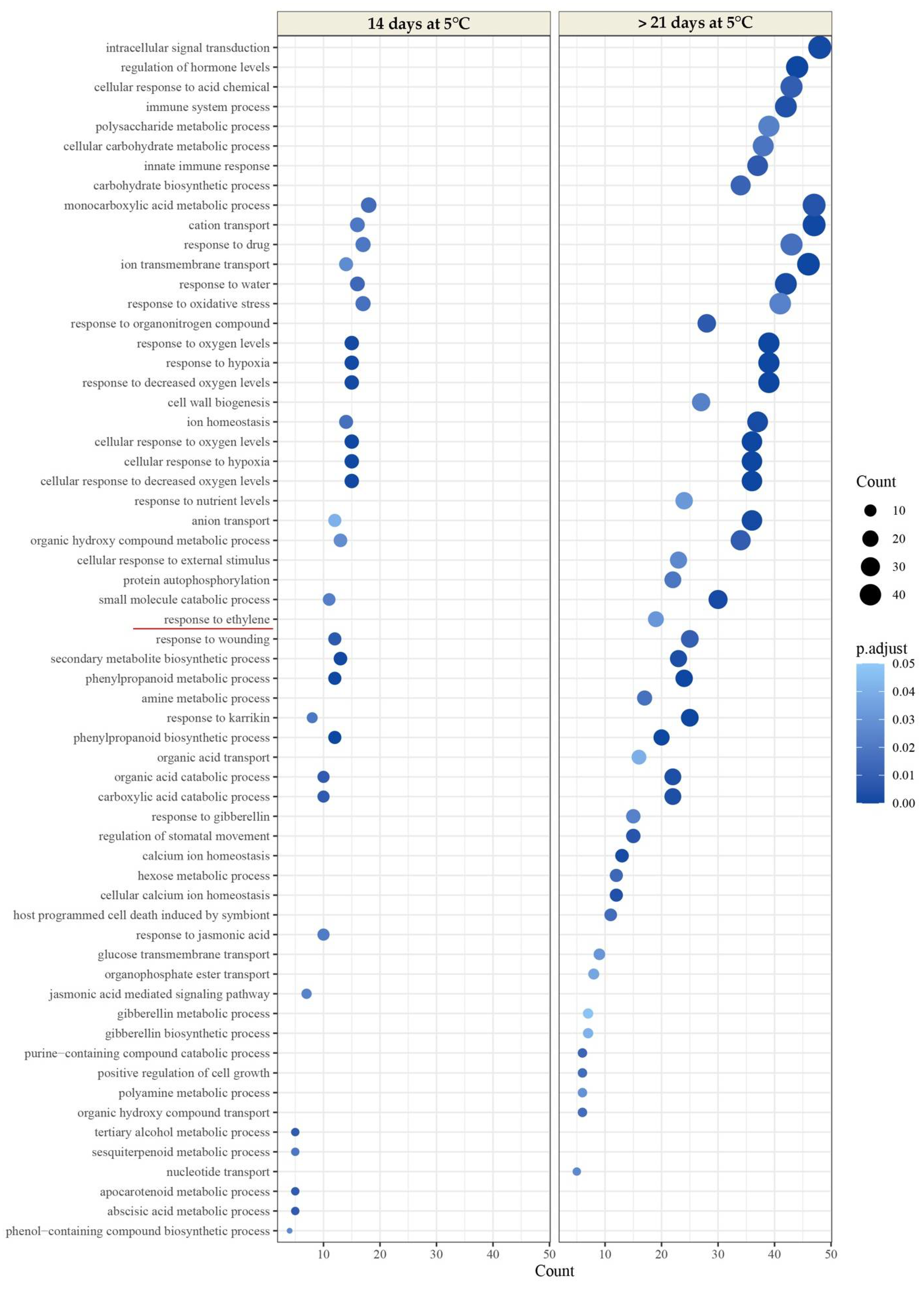
2.2.3. Transcriptomic Analysis
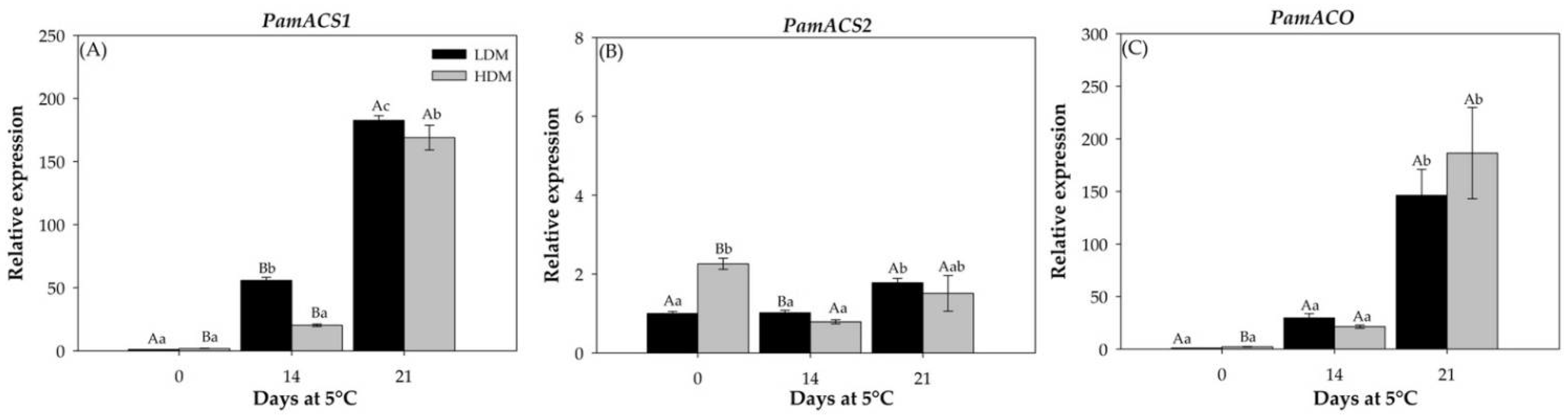
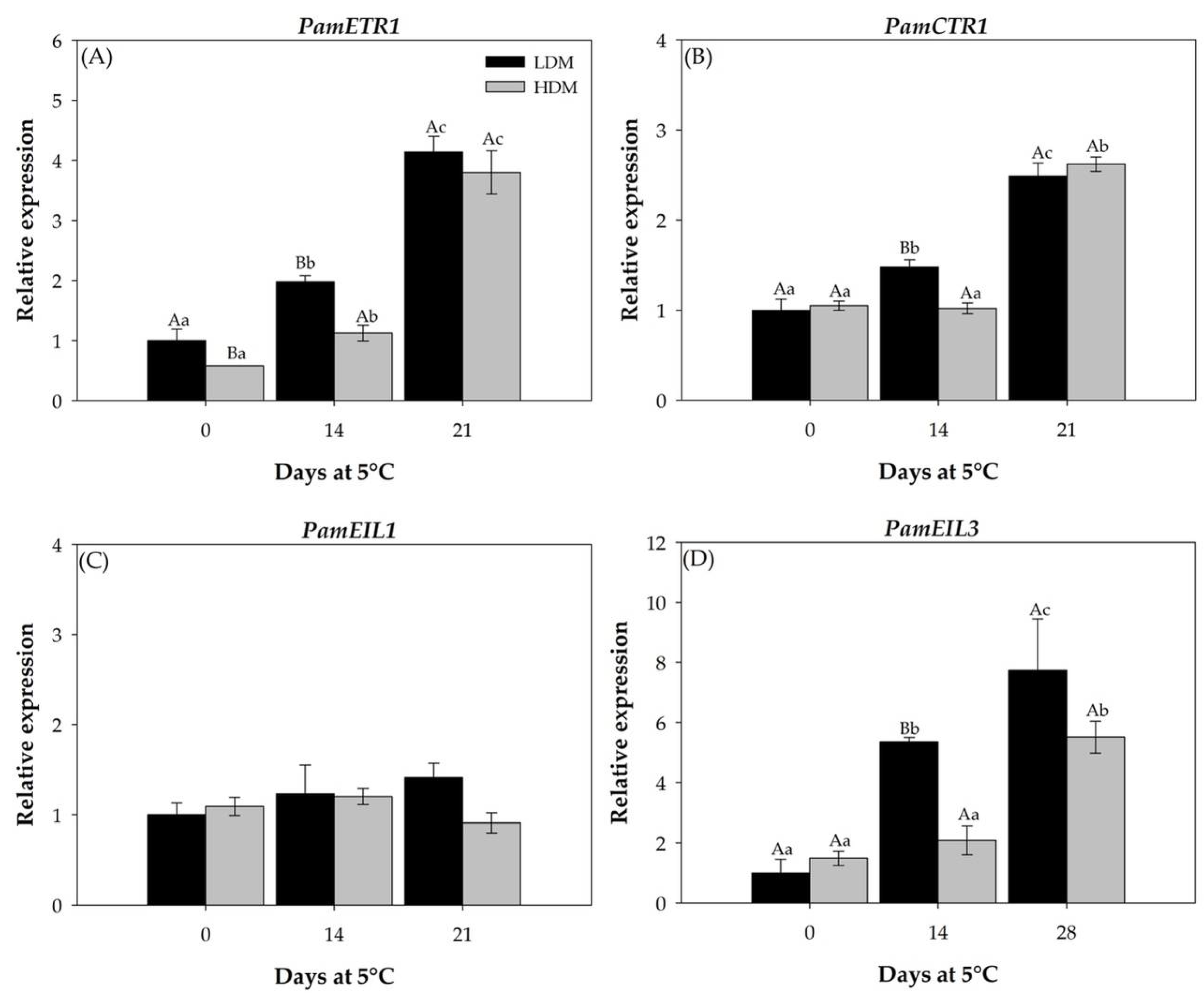
2.2.4. Expression Patterns of Genes Involved in Ethylene Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Dry Matter (DM)
4.3. 1–MCP Applications
4.4. Mesocarp Firmness
4.5. Skin Color
4.6. Respiration and Ethylene Production Rates
4.7. Bioinformatic Analysis
4.7.1. RNA-Seq Experiment
4.7.2. Sequencing Data Analysis
4.7.3. Differential Gene Expression Analysis
4.8. cDNA Synthesis and Real-Time Quantitative PCR Assays
4.9. Experimental Design and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hernández, I.; Uarrota, V.; Paredes, D.; Fuentealba, C.; Defilippi, B.G.; Campos-Vargas, R.; Meneses, C.; Hertog, M.; Pedreschi, R. Can Metabolites at Harvest Be Used as Physiological Markers for Modelling the Softening Behaviour of Chilean “Hass” Avocados Destined to Local and Distant Markets? Postharvest Biol. Technol. 2021, 174, 111457. [Google Scholar] [CrossRef]
- Uarrota, V.G.; Hernandez, I.; Ponce Guequen, E.; Vidal Cruz, J.; Fuentealba, C.; Defilippi, B.G.; Lindh, V.; Zulueta, C.; Chirinos, R.; Campos, D.; et al. Unravelling Factors Associated with ‘Blackspot’ Disorder in Stored Hass Avocado (Persea Americana Mill.) Fruit. J. Hortic. Sci. Biotechnol. 2020, 95, 804–815. [Google Scholar] [CrossRef]
- Defilippi, B.G.; Robledo, P.; Ferreyra, R.; Soto, S.; Saavedra, J. Preharvest Factors Influencing “Hass” Avocado (Persea Americana Mill.) Quality during Long Term Storage. Acta Hortic. 2015, 1071, 137–142. [Google Scholar] [CrossRef]
- Rivera, S.A.; Ferreyra, R.; Robledo, P.; Selles, G.; Lu, M.; Saavedra, J.; Defilippi, B.G. Identification of Preharvest Factors Determining Postharvest Ripening Behaviors in ‘ Hass ’ Avocado under Long Term Storage. Sci. Hortic. 2017, 216, 29–37. [Google Scholar] [CrossRef]
- Blakey, R.J.; Bower, J.P.; Bertling, I. Influence of Water and ABA Supply on the Ripening Pattern of Avocado (Persea Americana Mill.) Fruit and the Prediction of Water Content Using Near Infrared Spectroscopy. Postharvest Biol. Technol. 2009, 53, 72–76. [Google Scholar] [CrossRef]
- Shezi, S.; Magwaza, L.S.; Tesfay, S.Z.; Mditshwa, A. Biochemical Changes in Response to Canopy Position of Avocado Fruit (Cv. ‘Carmen’ and ‘Hass’) during Growth and Development and Relationship with Maturity. Sci. Hortic. 2020, 265, 109227. [Google Scholar] [CrossRef]
- Hershkovitz, V.; Friedman, H.; Goldschmidt, E.E.; Pesis, E. The Role of the Embryo and Ethylene in Avocado Fruit Mesocarp Discoloration. J. Exp. Bot. 2009, 60, 791–799. [Google Scholar] [CrossRef] [Green Version]
- Gwanpua, S.G.; Qian, Z.; East, A.R. Modelling Ethylene Regulated Changes in ‘Hass’ Avocado Quality. Postharvest Biol. Technol. 2018, 136, 12–22. [Google Scholar] [CrossRef]
- Mazhar, M.; Joyce, D.; Hofman, P.; Vu, N. Factors Contributing to Increased Bruise Expression in Avocado (Persea Americana M.) Cv. ‘Hass’ Fruit. Postharvest Biol. Technol. 2018, 143, 58–67. [Google Scholar] [CrossRef]
- Hershkovitz, V.; Saguy, S.I.; Pesis, E. Postharvest Application of 1–MCP to Improve the Quality of Various Avocado Cultivars. Postharvest Biol. Technol. 2005, 37, 252–264. [Google Scholar] [CrossRef]
- Hernández, I.; Fuentealba, C.; Olaeta, J.A.; Poblete-Echeverría, C.; Defilippi, B.G.; González-Agüero, M.; Campos-Vargas, R.; Lurie, S.; Pedreschi, R. Effects of Heat Shock and Nitrogen Shock Pre-Treatments on Ripening Heterogeneity of Hass Avocados Stored in Controlled Atmosphere. Sci. Hortic. 2017, 225, 408–415. [Google Scholar] [CrossRef]
- Burdon, J.; Lallu, N.; Haynes, G.; McDermott, K.; Billing, D. The Effect of Delays in Establishment of a Static or Dynamic Controlled Atmosphere on the Quality of “Hass” Avocado Fruit. Postharvest Biol. Technol. 2008, 49, 61–68. [Google Scholar] [CrossRef]
- Arpaia, M.L. Preharvest Factors Influencing Postharvest Quality of Tropical and Subtropical Fruit. HortScience 1994, 29, 982–985. [Google Scholar] [CrossRef] [Green Version]
- Pesis, E.; Ackerman, M.; Ben-Arie, R.; Feygenberg, O.; Feng, X.; Apelbaum, A.; Goren, R.; Prusky, D. Ethylene Involvement in Chilling Injury Symptoms of Avocado during Cold Storage. Postharvest Biol. Technol. 2002, 24, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Serek, M.; Woltering, E.J.; Sisler, E.C.; Frello, S.; Sriskandarajah, S. Controlling Ethylene Responses in Flowers at the Receptor Level. Biotechnol. Adv. 2006, 24, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Watkins, C.B. The Use of 1-Methylcyclopropene (1–MCP) on Fruits and Vegetables. Biotechnol. Adv. 2006, 24, 389–409. [Google Scholar] [CrossRef]
- Jeong, J.; Huber, D.J.; Sargent, S.A. Influence of 1-Methylcyclopropene (1–MCP) on Ripening and Cell-Wall Matrix Polysaccharides of Avocado (Persea Americana) Fruit. Postharvest Biol. Technol. 2002, 25, 241–256. [Google Scholar] [CrossRef]
- Jeong, J.; Huber, D.J. Suppression of Avocado (Persea Americana Mill.) Fruit Softening and Changes in Cell Wall Matrix Polysaccharides and Enzyme Activities: Differential Responses to 1–MCP and Delayed Ethylene Application. J. Am. Soc. Hort. Sci. 2004, 129, 752–759. [Google Scholar] [CrossRef] [Green Version]
- Adkins, M.F.; Hofman, P.J.; Stubbings, B.A.; Macnish, A.J. Manipulating Avocado Fruit Ripening with 1-Methylcyclopropene. Postharvest Biol. Technol. 2005, 35, 33–42. [Google Scholar] [CrossRef]
- Ochoa-Ascencio, S.; Hertog, M.L.A.T.M.; Nicolaï, B.M. Modelling the Transient Effect of 1–MCP on ‘Hass’ Avocado Softening: A Mexican Comparative Study. Postharvest Biol. Technol. 2009, 51, 62–72. [Google Scholar] [CrossRef]
- Feng, X.; Apelbaum, A.; Sisler, E.; Goren, R. Control of ethylene responses in avocado fruit with 1– MCP. Postharvest Biol. Technol. 2000, 20, 143–150. [Google Scholar] [CrossRef]
- Woolf, A.B.; Requejo-tapia, C.; Cox, K.A.; Jackman, R.C.; Gunson, A.; Lu, M.; White, A. 1–MCP Reduces Physiological Storage Disorders of ‘ Hass ’ Avocados. Postharvest Biol. Technol. 2005, 35, 43–60. [Google Scholar] [CrossRef]
- Zoffoli, J.P.; Callejas, M.J.; Beltran, T. 1–MCP: Una Herramienta Efectiva Para Controlar La Maduracion de Palta y Relacionarla Con La Madurez de Cosecha. Aconex 2003, 24–30. Available online: https://hdl.handle.net/20.500.14001/29104 (accessed on 6 June 2022).
- Owino, W.O.; Nakano, R.; Kubo, Y.; Inaba, A. Differential Regulation of Genes Encoding Ethylene Biosynthesis Enzymes and Ethylene Response Sensor Ortholog during Ripening and in Response to Wounding in Avocados. J. Am. Soc. Hort. Sci. 2002, 127, 520–527. [Google Scholar] [CrossRef] [Green Version]
- McGarvey, D.J.; Sirevåg, R.; Christoffersen, R.E. Ripening-Related Gene from Avocado Fruit: Ethylene-Inducible Expression of the MRNA and Polypeptide. Plant Physiol. 1992, 98, 554–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- In, B.C.; Strable, J.; Binder, B.M.; Falbel, T.G.; Patterson, S.E. Morphological and Molecular Characterization of Ethylene Binding Inhibition in Carnations. Postharvest Biol. Technol. 2013, 86, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Tian, M.S.; Prakash, S.; Zhang, N.; Ross, G.S. Chilling-Induced Ethylene Biosynthesis in Braeburn Apples. Plant Growth Regul. 2002, 38, 249–257. [Google Scholar] [CrossRef]
- Fonseca, S.; Monteiro, L.; Barreiro, M.G.; Pais, M.S. Expression of Genes Encoding Cell Wall Modifying Enzymes Is Induced by Cold Storage and Reflects Changes in Pear Fruit Texture. J Exp. Bot. 2005, 56, 2029–2036. [Google Scholar] [CrossRef] [Green Version]
- El-Sharkawy, I.; Jones, B.; Gentzbittel, L.; Lelièvre, J.; Pech, J.C.; Latché, A. Differential Regulation of ACC Synthase Genes in Cold-Dependent and -Independent Ripening in Pear Fruit. Plant Cell Environ. 2004, 27, 1197–1210. [Google Scholar] [CrossRef]
- Hershkovitz, V.; Friedman, H.; Goldschmidt, E.E.; Feygenberg, O.; Pesis, E. Induction of Ethylene in Avocado Fruit in Response to Chilling Stress on Tree. J. Plant Physiol. 2009, 166, 1855–1862. [Google Scholar] [CrossRef]
- Rendón-Anaya, M.; Ibarra-Laclette, E.; Méndez-Bravo, A.; Lan, T.; Zheng, C.; Carretero-Paulet, L.; Perez-Torres, C.A.; Chacón-López, A.; Hernandez-Guzmán, G.; Chang, T.H.; et al. The Avocado Genome Informs Deep Angiosperm Phylogeny, Highlights Introgressive Hybridization, and Reveals Pathogen-Influenced Gene Space Adaptation. Proc. Natl. Acad. Sci. USA 2019, 116, 17081–17089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisler, E.C.; Serek, M. Inhihitors of Ethylene Responses in Plants at the Receptor Level: Recent Developments. Physiol. Plant. 1997, 100, 577–582. [Google Scholar] [CrossRef]
- Manriquez, D.; Quintana, J.; Retamales, J. Inhibition of Ripen-Ing by 1–MCP in Hass Avocado Fruit. In Biology and Biotechnology of the Plant Hormone Ethylene III; Vendrell, M., Klee, H., Pech, J.C., Romojaro, F., Eds.; Springer: Amsterdam, The Netherlands, 2003; pp. 461–462. [Google Scholar]
- Feng, X.; Apelbaum, A.; Sisler, E.C.; Goren, R. Control of Ethylene Activity in Various Plant Systems by Structural Analogues of 1-Methylcyclopropene. Plant Growth Regul. 2004, 42, 29–38. [Google Scholar] [CrossRef]
- Lemmer, D.; Bezuidenhout, J.; Sekhune, S.; Ramokone, P.; Letsoalo, L.; Malumane, T.R.; Chibi, P.; Nxudu, Y.; Kruger, F.J. Semi-Commercial Evaluation of SmartFreshTM with South African Export Avocados in Static Containers at the Westfalia Packhouse during 2002. S. Afr. Avocados Grow. Assoc. Yearb. 2003, 26, 55–64. [Google Scholar]
- Lemmer, D.; Kruger, F.J. Laboratory Based Evaluation of 1-Methyl Cyclopropene (1–MCP): With Five South African Commercial Export Avocado Cultivars. In Proceedings of the V World Avocado Congress, Granada–Málaga, Spain, 19–24 October 2003; pp. 617–622. [Google Scholar]
- Magwaza, L.S.; Tesfay, S.Z. A Review of Destructive and Non-Destructive Methods for Determining Avocado Fruit Maturity. Food Bioprocess Technol. 2015, 8, 1995–2011. [Google Scholar] [CrossRef]
- Jeong, J.; Huber, D.J.; Sargent, S.A. Delay of Avocado (Persea Americana) Fruit Ripening by 1-Methylcyclopropene and Wax Treatments. Postharvest Biol. Technol. 2003, 28, 247–257. [Google Scholar] [CrossRef]
- Pesis, E.; Krilo, V.; Feygenberg, O.; Ackerman, M.; Ben-Arie, R.; Prusky, D. Postharvest Effect of 1–MCP on Ripening of Avocado Cv. Ettinger. In Biology and Biotechnology of the Plant Hormone Ethylene III; Springer: Amsterdam, The Netherlands, 2003; pp. 397–401. [Google Scholar]
- Golding, J.B.; Shearer, D.; Mcglasson, W.B.; Wyllie, S.G. Relationships between Respiration, Ethylene, and Aroma Production in Ripening Banana. J. Agric. Food Chem. 1999, 47, 1646–1651. [Google Scholar] [CrossRef]
- Golding, J.B.; Shearer, D.; Wyllie, S.G.; Mcglasson, W.B. Application of 1–MCP and Propylene to Identify Ethylene-Dependent Ripening Processes in Mature Banana Fruit. Postharvest Biol. Technol. 1998, 14, 87–98. [Google Scholar] [CrossRef]
- Fan, X.; Mattheis, J.P. Impact of 1-Methylcyclopropene and Methyl Jasmonate on Apple Volatile Production. J. Agric. Food Chem. 1999, 47, 2847–2853. [Google Scholar] [CrossRef]
- Fan, X.; Argenta, L.; Mattheis, J.P. Inhibition of Ethylene Action by 1-Methylcyclopropene Prolongs Storage Life of Apricots. Postharvest Biol. Technol. 2000, 20, 135–142. [Google Scholar] [CrossRef]
- Giovannoni, J. Molecular Regulation of Fruit Ripening. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 725–749. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Chen, K.S.; Xu, W.P.; Zhang, W.S.; Li, X.; Ferguson, I. Effect of 1–MCP on Postharvest Quality of Loquat Fruit. Postharvest Biol. Technol. 2006, 40, 155–162. [Google Scholar] [CrossRef]
- Yang, S.F.; Hoffman, N.E. Ethylene Biosynthesis and Its Regulation in Higher Plants. Annu. Rev. Plant Physiol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Kende, H. Ethylene Biosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 283–307. [Google Scholar] [CrossRef]
- Chang, C.; Kwok, S.F.; Bleecker, A.B.; Meyerowitz, E.M. Arabidopsis Ethylene-Response Gene ETR1: Similarity of Product to Two-Component Regulators. Science 1993, 262, 539–544. [Google Scholar] [CrossRef]
- Hua, J.; Chang, C.; Sun, Q.; Meyerowitz, E.M. Ethylene Insensitivity Conferred by Arabidopsis EPS Gene. Science 1995, 269, 1712–1714. [Google Scholar] [CrossRef]
- Sakai, H.; Hua, J.; Chen, Q.G.; Chang, C.; Medrano, L.J.; Bleecker, A.B.; Meyerowitz, E.M. ETR2 Is an ETR1-like Gene Involved in Ethylene Signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 5812–5817. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Cureatz, V.; Toivonen, P.M.A. Improved Quality Retention of Packaged “Anjou” Pear Slices Using a 1-Methylcyclopropene (1–MCP) Co-Release Technology. Postharvest Biol. Technol. 2009, 51, 378–383. [Google Scholar] [CrossRef]
- Selvarajah, S.; Bauchot, A.D.; John, P. Internal Browning in Cold-Stored Pineapples Is Suppressed by a Postharvest Application of 1-Methylcyclopropene. Postharvest Biol. Technol. 2001, 23, 167–170. [Google Scholar] [CrossRef]
- Gudenschwager, O.; García-Rojas, M.; Defilippi, B.G.; González-Agüero, M. Identification and Characterization of Two Putative Genes Encoding Acetyl-Coenzyme A Carboxylase Subunits That Are Possibly Associated with Internal Browning during Cold Storage of “Hass” Avocados (Persea Americana Mill.). Postharvest Biol. Technol. 2013, 84, 74–80. [Google Scholar] [CrossRef]
- Pedreschi, R.; Muñoz, P.; Robledo, P.; Becerra, C.; Defilippi, B.G.; Van Eekelen, H.; Mumm, R.; Westra, E.; de Vos, R.C.H. Metabolomics Analysis of Postharvest Ripening Heterogeneity of ‘Hass’ Avocadoes. Postharvest Biol. Technol. J. 2014, 92, 172–179. [Google Scholar] [CrossRef]
- García-rojas, M.; Morgan, A.; Gudenschwager, O.; Zamudio, S.; Campos-vargas, R.; González-agüero, M.; Defilippi, B.G. Scientia Horticulturae Biosynthesis of Fatty Acids-Derived Volatiles in ‘Hass’ Avocado Is Modulated by Ethylene and Storage Conditions during Ripening. Sci. Hortic. 2016, 202, 91–98. [Google Scholar] [CrossRef]
- Defilippi, B.G.; Ejsmentewicz, T.; Covarrubias, M.P.; Gudenschwager, O.; Campos-Vargas, R. Changes in Cell Wall Pectins and Their Relation to Postharvest Mesocarp Softening of “Hass” Avocados (Persea Americana Mill.). Plant Physiol. Biochem. 2018, 128, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Olivares, D.; Alvarez, E.; Veliz, D.; García-Rojas, M.; Díza, C.; Defilippi, B.G. Effects of 1-Methylcyclopropene and Controlled Atmosphere on Ethylene Synthesis and Quality Attributes of Avocado Cvs. Edranol and Fuerte. J. Food Qual. 2020, 20, 14. [Google Scholar] [CrossRef]
- Gudenschwager, O.; González-Agüero, M.; Defilippi, B.G. A General Method for High-Quality RNA Isolation from Metabolite-Rich Fruits. S. Afr. J. Bot. 2012, 83, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Hershkovitz, V.; Friedman, H.; Goldschmidt, E.E.; Pesis, E. Ethylene Regulation of Avocado Ripening Differs between Seeded and Seedless Fruit. Postharvest Biol. Technol. 2010, 56, 138–146. [Google Scholar] [CrossRef]
- Pfaffl, M. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, 2003–2007. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Harvest | Trial | Up | Down |
|---|---|---|---|
| LDM | 14 d at 5 °C vs. Harvest | 701 | 657 |
| >21 d at 5 °C vs. Harvest | 1829 | 2651 | |
| HDM | 14 d at 5 °C vs. Harvest | 477 | 644 |
| >21 d at 5 °C vs. Harvest | 2214 | 3065 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olivares, D.; García-Rojas, M.; Ulloa, P.A.; Riveros, A.; Pedreschi, R.; Campos-Vargas, R.; Meneses, C.; Defilippi, B.G. Response Mechanisms of “Hass” Avocado to Sequential 1–methylcyclopropene Applications at Different Maturity Stages during Cold Storage. Plants 2022, 11, 1781. https://doi.org/10.3390/plants11131781
Olivares D, García-Rojas M, Ulloa PA, Riveros A, Pedreschi R, Campos-Vargas R, Meneses C, Defilippi BG. Response Mechanisms of “Hass” Avocado to Sequential 1–methylcyclopropene Applications at Different Maturity Stages during Cold Storage. Plants. 2022; 11(13):1781. https://doi.org/10.3390/plants11131781
Chicago/Turabian StyleOlivares, Daniela, Miguel García-Rojas, Pablo A. Ulloa, Aníbal Riveros, Romina Pedreschi, Reinaldo Campos-Vargas, Claudio Meneses, and Bruno G. Defilippi. 2022. "Response Mechanisms of “Hass” Avocado to Sequential 1–methylcyclopropene Applications at Different Maturity Stages during Cold Storage" Plants 11, no. 13: 1781. https://doi.org/10.3390/plants11131781