
Deciphering the Genetic Inheritance of Tocopherols in Indian Mustard (Brassica juncea L. Czern and Coss)

 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Inheritance Pattern of Tocopherol Contents
2.2. Estimation of Genetic Parameters
2.3. Estimation of Scaling Test and Genes Effects
3. Materials and Methods
3.1. Selection of Genotypes
3.2. Development of Different Breeding Populations
3.3. Experimental Design and Crop Maintenance
3.4. Estimation of Tocopherol Contents
3.5. Statistical Analysis
3.5.1. Estimation of Scaling Test
3.5.2. Components of Generation Means
3.5.3. Estimation of Various Genetic Parameters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goffman, F.D.; Becker, H.C. Genetic variation of tocopherol content in a germplasm collection of Brassica napus L. Euphytica 2002, 125, 189–196. [Google Scholar] [CrossRef]
- Endrigkeit, J.; Wang, X.; Cai, D.; Zhang, C.; Long, Y.; Meng, J.; Jung, C. Genetic mapping, cloning, and functional characterization of the BnaX.VTE4 gene encoding a γ-tocopherol methyltransferase from oilseed rape. Theor. Appl. Genet. 2009, 119, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Marwede, V.; Schierholt, A.; Möllers, C.; Becker, H.C. Genotype × Environment interactions and heritability of tocopherol contents in Canola. Crop Sci. 2004, 44, 728–731. [Google Scholar] [CrossRef]
- Kamal-Eldin, A.; Andersson, R. A multivariate study of the correlation between tocopherol content and fatty acid composition in vegetable oils. J. Am. Oil Chem. Soc. 1997, 74, 375–380. [Google Scholar] [CrossRef]
- Goffman, F.D.; Thies, W.; Velasco, L. Chemotaxonomic value of tocopherols in Brassicaceae. Phytochemistry 1999, 50, 793–798. [Google Scholar] [CrossRef]
- DellaPenna, D. A decade of progress in understanding vitamin E synthesis in plants. J. Plant Physiol. 2005, 162, 729–737. [Google Scholar] [CrossRef]
- Valentin, H.E.; Lincoln, K.; Moshiri, F.; Jensen, P.K.; Qi, Q.; Venkatesh, T.V.; Karunanandaa, B.; Baszis, S.R.; Norris, S.R.; Savidge, B.; et al. The Arabidopsis vitamin E pathway gene5-1 mutant reveals a critical role for phytol kinase in seed tocopherol biosynthesis. Plant Cell 2006, 18, 212–224. [Google Scholar] [CrossRef] [Green Version]
- Hunter, S.C.; Cahoon, E.B. Enhancing vitamin E in oilseeds: Unraveling tocopherol and tocotrienol biosynthesis. Lipids 2007, 42, 97–108. [Google Scholar] [CrossRef]
- Fritsche, S.; Wang, X.; Li, J.; Stich, B.; Kopisch-Obuch, F.J.; Endrigkeit, J.; Leckband, G.; Dreyer, F.; Friedt, W.; Meng, J.; et al. A candidate gene-based association study of tocopherol content and composition in rapeseed (Brassica napus). Front. Plant Sci. 2012, 3, 129. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, C.; Li, L.; Fritsche, S.; Endrigkeit, J.; Zhang, W.; Long, Y.; Jung, C.; Meng, J. Unraveling the genetic basis of seed tocopherol content and composition in rapeseed (Brassica napus L.). PLoS ONE 2012, 7, e50038. [Google Scholar] [CrossRef] [Green Version]
- Goffman, F.D.; Becker, H.C. Diallel analysis for tocopherol contents in seeds of rapeseed. Crop Sci. 2001, 41, 1072–1079. [Google Scholar] [CrossRef]
- Soll, J.; Douce, R.; Schultz, G. Site of biosynthesis of a-tocopherol in spinach chloroplasts. FEBS Lett. 1980, 112, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Fritsche, S.; Wang, X.; Nichelmann, L.; Suppanz, I.; Hadenfeldt, S.; Endrigkeit, J.; Meng, J.; Jung, C. Genetic and functional analysis of tocopherol biosynthesis pathway genes from rapeseed (Brassica napus L.). Plant Breed. 2014, 133, 470–479. [Google Scholar] [CrossRef]
- Van Eenennaam, A.; Lincoln, K.; Durrett, T.P.; Valentin, H.E.; Shewmaker, C.K.; Thorne, G.M.; Jiang, J.; Baszis, S.R.; Levering, C.K.; Aasen, E.D.; et al. Engineering vitamin e content: From arabidopsis mutant to soy oil. Plant Cell 2003, 15, 3007–3019. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, Z.; Sun, X.; Tang, K. Current opinions on the functions of tocopherol based on the genetic manipulation of tocopherol biosynthesis in plants. J. Integr. Plant Biol. 2008, 50, 1057–1069. [Google Scholar] [CrossRef]
- Marwede, V.; Gul, M.K.; Becker, H.C.; Ecke, W. Mapping of QTL controlling tocopherol content in winter oilseed rape. Plant Breed. 2005, 124, 20–26. [Google Scholar] [CrossRef]
- Chand, S.; Patidar, O.P.; Chaudhary, R.; Saroj, R.; Chandra, K.; Meena, V.K.; Limbalkar, O.M.; Patel, M.K.; Pardeshi, P.; Vasisth, P. Rapeseed-mustard breeding in India: Scenario, achievements and research needs. In Brassica Breeding and Biotechnology; IntechOpen: London, UK, 2021; p. 174. ISBN 9781839686986. [Google Scholar]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli-Michael, M.; Govindjee; Sarin, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochim. Biophys. Acta-Bioenergy 2010, 1797, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- García-Navarro, E.; Fernández-Martínez, J.M.; Pérez-Vich, B.; Velasco, L. Genetic analysis of reducedγ-tocopherol content in ethiopian mustard seeds. Sci. World J. 2016, 2016, 7392603. [Google Scholar] [CrossRef] [Green Version]
- García-Moreno, M.J.; Fernández-Martínez, J.M.; Velasco, L.; Pérez-Vich, B. Genetic basis of unstable expression of high gamma-tocopherol content in sunflower seeds. BMC Plant Biol. 2012, 12, 71. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, M.A.; Sarin, N.B. Antioxidant value addition in human diets: Genetic transformation of Brassica juncea with γ-TMT gene for increased α-tocopherol content. Transgenic Res. 2007, 16, 109–113. [Google Scholar] [CrossRef]
- Siger, A.; Michalak, M.; Cegielska-Taras, T.; Szała, L.; Lembicz, J.; Nogala-Kałucka, M. Genotype and environment effects on tocopherol and plastochromanol-8 contents of winter oilseed rape doubled haploid lines derived from F1 plants of the cross between yellow and black seeds. Ind. Crop. Prod. 2015, 65, 134–141. [Google Scholar] [CrossRef]
- Gawrysiak-Witulska, M.; Siger, A.; Wawrzyniak, J.; Nogala-Kalucka, M. Changes in tocochromanol content in seeds of Brassica napus L. during adverse conditions of storage. J. Am. Oil Chem. Soc. 2011, 88, 1379–1385. [Google Scholar] [CrossRef]
- Gawrysiak-Witulska, M.; Siger, A.; Rusinek, R. Degradation of tocopherols during rapeseed storage in simulated conditions of industrial silos. Int. Agrophysics 2016, 30, 39–45. [Google Scholar] [CrossRef]
- Capitani, M.; Mateo, C.M.; Nolasco, S.M. Effect of temperature and storage time of wheat germ on the oil tocopherol concentration. Braz. J. Chem. Eng. 2011, 28, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Lavedrine, F.; Ravel, A.; Poupard, A.; Alary, J. Effect of geographic origin, variety and storage on tocopherol concentrations in walnuts by HPLC. Food Chem. 1997, 58, 135–140. [Google Scholar] [CrossRef]
- Johnson, H.W.; Robinson, H.F.; Comstock, R.E. Estimates of genetic and environmental variability in Soybeans 1. Agron. J. 1955, 47, 314–318. [Google Scholar] [CrossRef]
- Singh, P.; Narayanan, S.S. Biometrical Techniques in Plant Breeding, 1st ed.; Kayani Publishers: Ludhiyana, India, 1993; ISBN 741339390. [Google Scholar]
- Mather, K.; Jinks, J.L. Biometrical Genetics: The Study of Continuous Variation, 3rd ed.; Chapman and Hall: London, UK, 1982; ISBN 0412228904. [Google Scholar]
- Alza, J.; Fernandez-Martinez, J. Genetic analysis of yield and related traits in sunflower (Helianthus annuus L.) in dryland and irrigated environments. Euphytica 1997, 95, 243–251. [Google Scholar] [CrossRef]
- Sharma, S.; Sain, R.; Sharma, R. Genetics of spike length in durum wheat. Euphytica 2003, 130, 155–161. [Google Scholar] [CrossRef]
- Singh, R.K.; Chaudhary, B.D. Biometrical Methods in Quantitative Genetic Analysis, 1st ed.; Kalyani Publishers: New Delhi, India, 1999; ISBN 8176633070. [Google Scholar]
- Kearsey, M.J.; Pooni, H.S. The Genetical Analysis of Quantitative Traits, 2nd ed.; Chapman and Hall: London, UK, 2004; ISBN 0748740821. [Google Scholar]
- Said, A. Generation mean analysis in wheat (Triticum aestivum L.) under drought stress conditions. Ann. Agric. Sci. 2014, 59, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Amin, I.A. Genetic behaviour of some agronomic traits in two durum wheat crosses under heat stress. J. Agric. Res. 2013, 58, 53–66. [Google Scholar]
- Mather, K. Biometrical Genetics: The Study of Continuous Variation; Methuen: London, UK, 1949. [Google Scholar]
- Smith, H. Fixing transgressive vigor in Nicotiana rustica. In Heterosis; Gowen, J.W., Ed.; Iowa State College Press: Ames, IA, USA, 1952; pp. 161–174. ISBN 19531603834. [Google Scholar]
- El-Tahawey, M.A.F.A.; Kandeel, A.M.; Youssef, S.M.S.; El-Salam, M.M.M.A. Heterosis, potence ratio, combining ability and correlation of some economic traits in diallel crosses of pumpkins. Egypt. J. Plant Breed. 2015, 19, 419–439. [Google Scholar] [CrossRef]
- Hayman, B.I.; Mather, K. The description of genic interactions in continuous variation. Biometrics 1955, 11, 69. [Google Scholar] [CrossRef]
- Hayman, B.I. The separation of epistatic from additive and dominance variation in generation means. Heredity 1958, 12, 371–390. [Google Scholar] [CrossRef] [Green Version]
- Jinks, J.L.; Jones, R.M. Estimation of the components of heterosis. Genetics 1958, 43, 223–224. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.C. Epistasis—The essential role of gene interactions in the structure and evolution of genetic systems. Nat. Rev. Genet. 2008, 9, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Khattab, S.A.M.; Esmail, R.M.; Al-Ansary, A.E.M.F. Genetical analysis of some quantitative traits in bread wheat (Triticum aestivum L.). New York Sci. J. 2010, 3, 152–157. [Google Scholar]
- Shahrokhi, M.; Khorasani, S.K.; Ebrahimi, A. Generation mean analysis: A case study of yield and yield components in ksc704 maize (Zea mays L.). Generations 2013, 4, 2686–2693. [Google Scholar]
- Cukadar-Olmedo, B.; Miller, J.F. Inheritance of the Stay Green Trait in Sunflower. Crop Sci. 1997, 37, 150–153. [Google Scholar] [CrossRef]
- Kearsey, M.J. Biometrical genetics in breeding. Plant Breed 1993, 163–183. [Google Scholar] [CrossRef]
- García-Moreno, M.J.; Fernández-Martínez, J.M.; Velasco, L.; Pérez-Vich, B. Molecular tagging and candidate gene analysis of the high gamma-tocopherol trait in safflower (Carthamus tinctorius L.). Mol. Breed. 2010, 28, 367–379. [Google Scholar] [CrossRef]
- Hass, C.G.; Tang, S.; Leonard, S.; Traber, M.G.; Miller, J.F.; Knapp, S.J. Three non-allelic epistatically interacting methyltransferase mutations produce novel tocopherol (vitamin E) profiles in sunflower. Theor. Appl. Genet. 2006, 113, 767–782. [Google Scholar] [CrossRef] [PubMed]
- Dwiyanti, M.S.; Yamada, T.; Sato, M.; Abe, J.; Kitamura, K. Genetic variation of γ-tocopherol methyltransferase gene contributes to elevated α-tocopherol content in soybean seeds. BMC Plant Biol. 2011, 11, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shintani, D.; DellaPenna, D. Elevating the vitamin e content of plants through metabolic engineering. Science 1998, 282, 2098–2100. [Google Scholar] [CrossRef]
- Schledz, M.; Seidler, A.; Beyer, P.; Neuhaus, G. A novel phytyltransferase from Synechocystis sp. PCC 6803 involved in tocopherol biosynthesis. FEBS Lett. 2001, 499, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Dähnhardt, D.; Falk, J.; Appel, J.; van der Kooij, T.A.; Schulz-Friedrich, R.; Krupinska, K. The hydroxyphenylpyruvate dioxygenase from Synechocystis sp. PCC 6803 is not required for plastoquinone biosynthesis. FEBS Lett. 2002, 523, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Falk, J.; Andersen, G.; Kernebeck, B.; Krupinska, K. Constitutive overexpression of barley 4-hydroxyphenylpyruvate dioxygenase in tobacco results in elevation of the vitamin E content in seeds but not in leaves. FEBS Lett. 2003, 540, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Kearsey, M.J.; Pooni, H.S. The Genetical Analysis of Quantitative Traits; Chapman and Hall, Stanley Thornes: London, UK, 1996; ISBN 0412609800. [Google Scholar]
- Singh, R.K.; Chaudhary, B.D. Biometrical Methods in Quantitative Genetic Analysis; Kalyani Publishers: Ludhiyana, India, 1977; ISBN 19801689021. [Google Scholar]
- Ramli, A.B.; Rafii, M.Y.; Latif, M.A.; Saleh, G.B.; Omar, O.B.; Puteh, A.B. Generation mean analysis of grain quality traits in selected rice populations derived from different amylose characteristics. J. Sci. Food Agric. 2016, 96, 1593–1600. [Google Scholar] [CrossRef]
- Warner, J.N. A method for estimating heritability. Agron. J. 1952, 44, 427–430. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
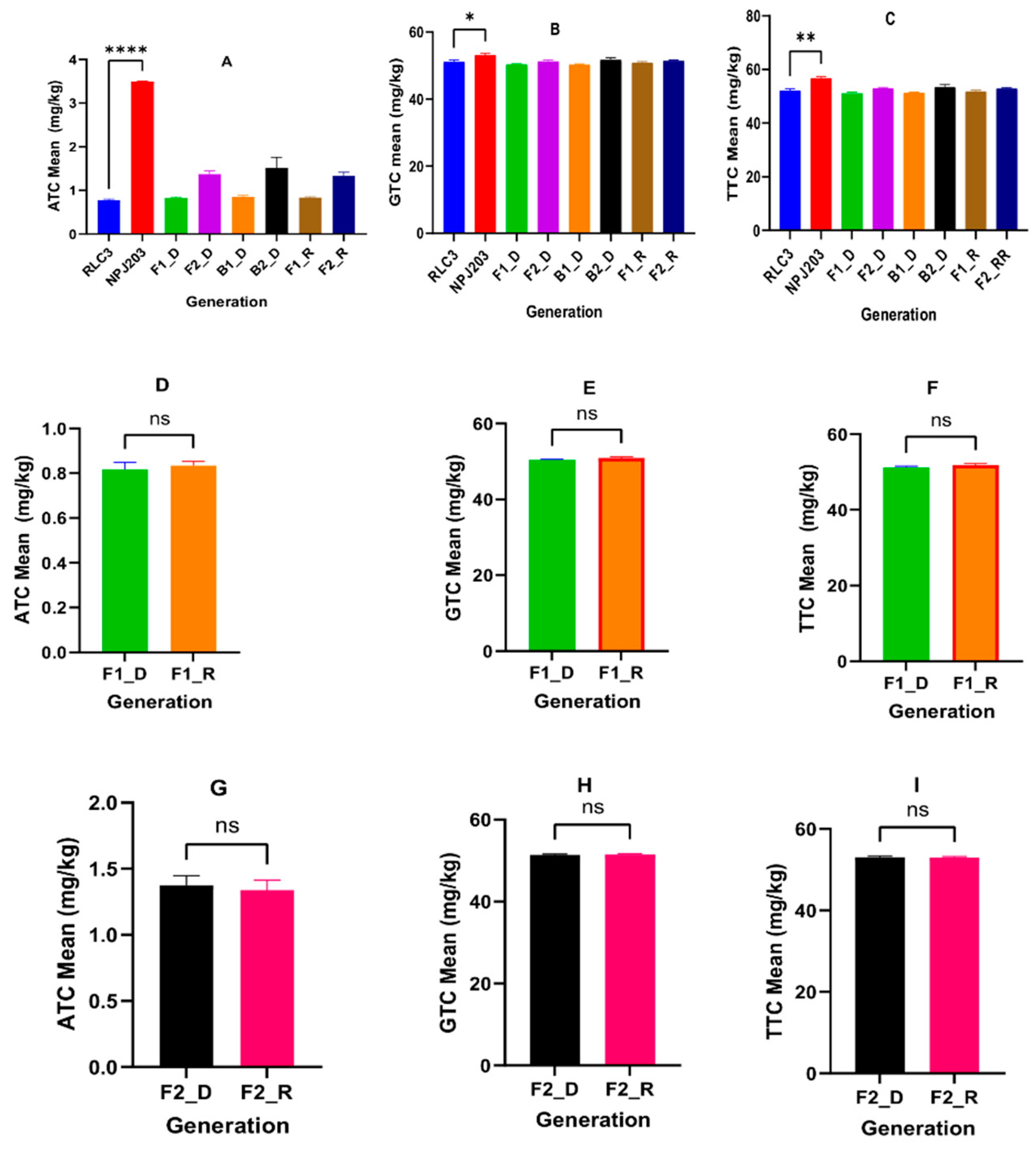
| Generations | RLC3 × NPJ203 (Direct Cross) | NPJ203 × RLC3 (Reciprocal Cross) | ||||||
|---|---|---|---|---|---|---|---|---|
| ATC | GTC | TTC | AGR% | ATC | GTC | TTC | AGR% | |
| RLC3 (P1) | 0.768 ± 0.035 | 51.182 ± 0.515 | 52.138 ± 0.539 | 1.50 | 0.788 ± 0.039 | 51.097 ± 0.346 | 52.037 ± 0.363 | 1.54 |
| NPJ203 (P2) | 3.492 ± 0.008 | 53.223 ± 0.425 | 56.848 ± 0.432 | 6.56 | 3.327 ± 0.094 | 54.47 ± 0.372 | 57.947 ± 0.414 | 6.11 |
| F1 | 0.815 ± 0.031 | 50.418 ± 0.174 | 51.188 ± 0.293 | 1.62 | 0.833 ± 0.035 | 50.903 ± 0.358 | 51.883 ± 0.363 | 1.64 |
| F2 | 1.371 ± 0.077 | 51.396 ± 0.241 | 53.03 ± 0.304 | 2.67 | 1.338 ± 0.077 | 51.48 ± 0.205 | 52.979 ± 0.272 | 2.60 |
| B1 | 0.853 ± 0.045 | 50.366 ± 0.302 | 51.373 ± 0.328 | 1.69 | 1.626 ± 0.285 | 52.527 ± 0.655 | 54.287 ± 0.911 | 1.72 |
| B2 | 1.511 ± 0.245 | 51.766 ± 0.636 | 53.486 ± 0.882 | 2.92 | 0.863 ± 0.047 | 50.278 ± 0.243 | 51.282 ± 0.266 | 3.10 |
| Character $ | Genetic Parameters and Variation Components # | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| H2b | H2n | GA | [h/d] | [E] | [D] | [H] | [F] | √H/D | F/√H×D | [P] | |
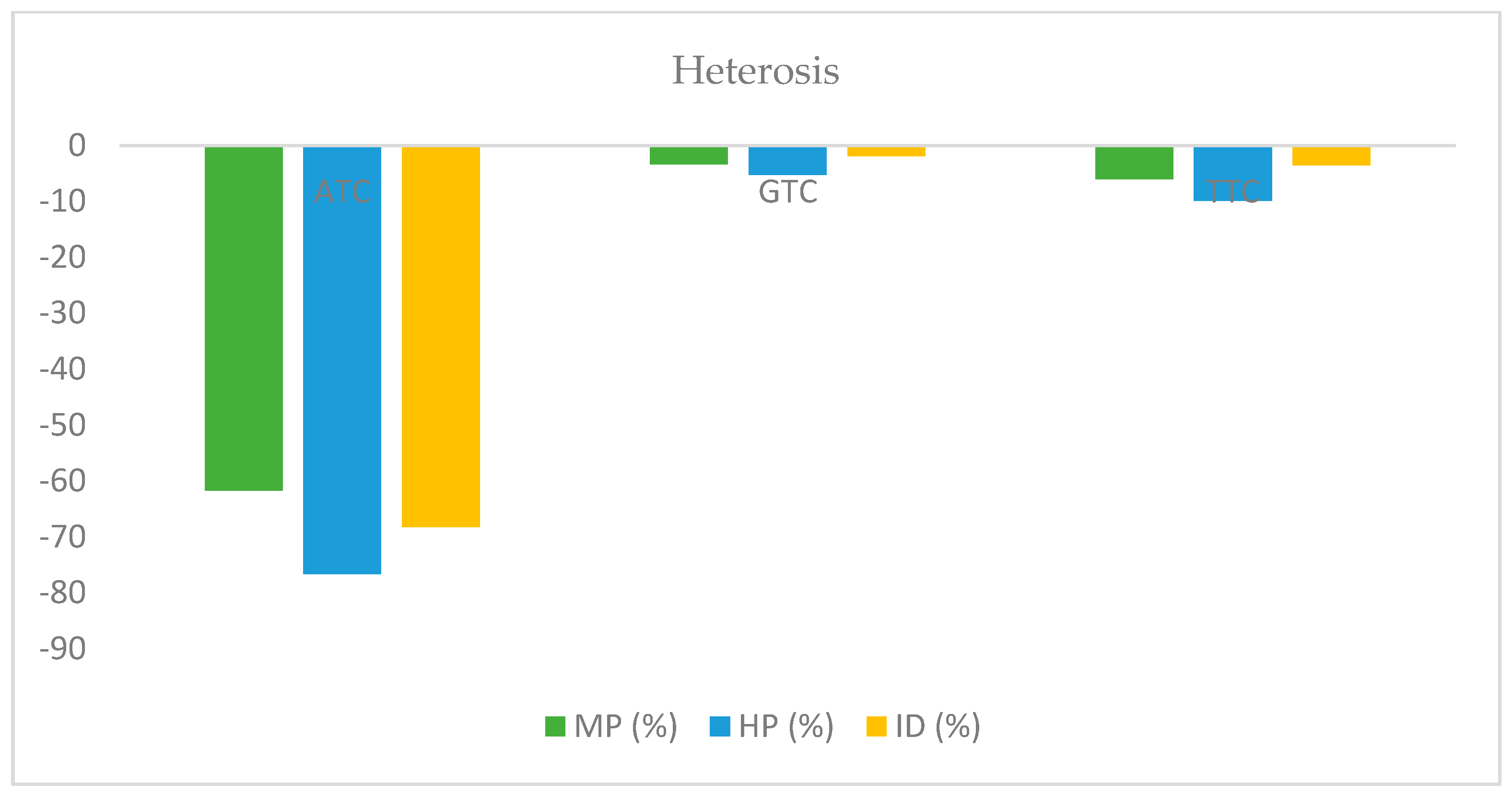
| ATC | 0.997 | 0.677 | 1.724 | 3.140 | 0.002 | 0.952 | 0.899 | −0.87 | 0.972 | −0.941 | −0.965 |
| GTC | 0.932 | 0.938 | 4.764 | 2.219 | 0.476 | 9.714 | 3.227 | −4.69 | 0.576 | −0.838 | −1.749 |
| TTC | 0.949 | 0.806 | 6.533 | 2.702 | 0.563 | 17.946 | 6.360 | −10.06 | 0.595 | −0.942 | −1.403 |
| Character | Scaling Test | Gene Effects by Six Parameter Model | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | [m] | [d] | [h] | [i] | [j] | [l] | Epistasis | |
| ATC | −0.122 | 1.284 ** | 0.408 | 0.377 | 1.371 ** | −0.659 ** | −2.069 ** | −0.754 | 1.406 ** | 1.916 ** | Duplicate |
| GTC | 0.869 | 0.11 | −0.343 | 0.661 | 51.396 ** | −1.4 ** | −3.106 | −1.321 | −0.759 | 2.3 | Absent |
| TTC | 0.581 | 1.065 | −0.758 | 1.202 | 53.03 ** | −2.113 ** | −5.709 | −2.404 | 0.485 | 4.05 | Absent |
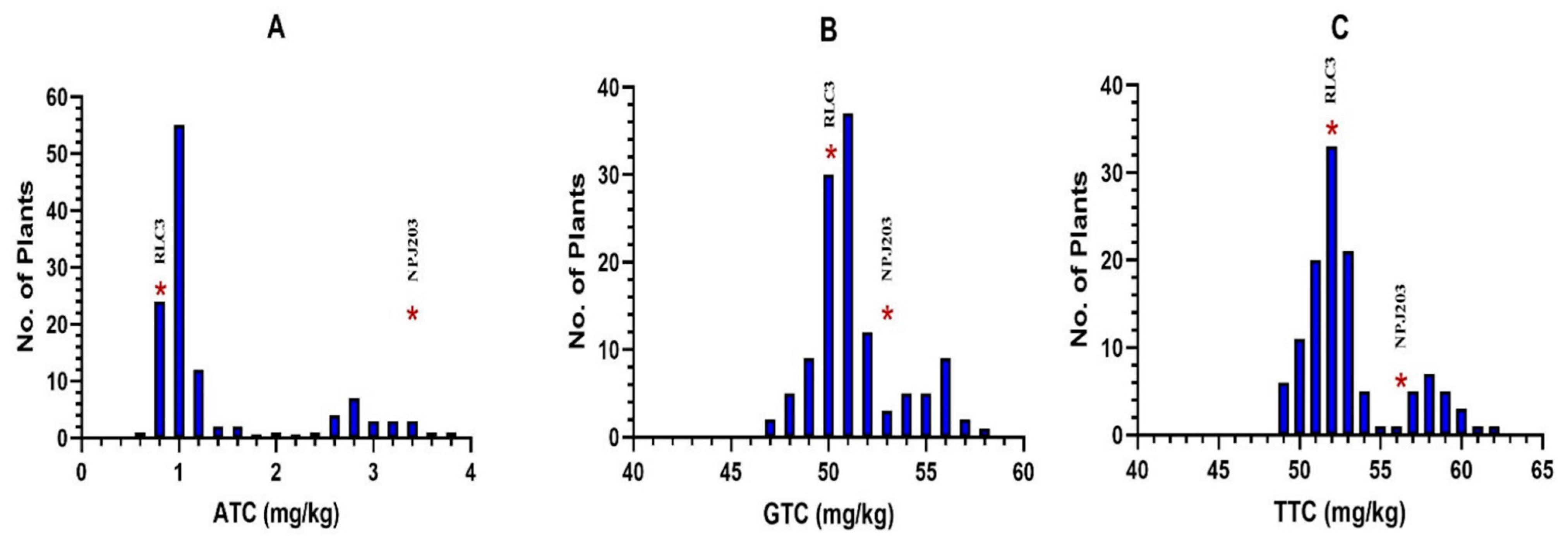
| S.N. | Genotypes | Pedigree/Source | Origin | Tocopherol Content Traits (mg/kg) | ||||
|---|---|---|---|---|---|---|---|---|
| ATC | GTC | DTC | AGR | TTC | ||||
| 1 | RLC3 | JM06003 × JM06020 | PAU, Ludhiana, Indian | 0.74 | 51.17 | 0.19 | 0.01 | 52.10 |
| 2 | NPJ203 | (EJ9913 × SEJ8) × Laxmi | IARI, New Delhi, Indian | 3.49 | 53.14 | 0.15 | 0.07 | 56.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meena, V.K.; Taak, Y.; Chaudhary, R.; Chand, S.; Patel, M.K.; Muthusamy, V.; Yadav, S.; Saini, N.; Vasudev, S.; Yadava, D.K. Deciphering the Genetic Inheritance of Tocopherols in Indian Mustard (Brassica juncea L. Czern and Coss). Plants 2022, 11, 1779. https://doi.org/10.3390/plants11131779
Meena VK, Taak Y, Chaudhary R, Chand S, Patel MK, Muthusamy V, Yadav S, Saini N, Vasudev S, Yadava DK. Deciphering the Genetic Inheritance of Tocopherols in Indian Mustard (Brassica juncea L. Czern and Coss). Plants. 2022; 11(13):1779. https://doi.org/10.3390/plants11131779
Chicago/Turabian StyleMeena, Vijay Kamal, Yashpal Taak, Rajat Chaudhary, Subhash Chand, Manoj Kumar Patel, Vignesh Muthusamy, Sangita Yadav, Navinder Saini, Sujata Vasudev, and Devendra Kumar Yadava. 2022. "Deciphering the Genetic Inheritance of Tocopherols in Indian Mustard (Brassica juncea L. Czern and Coss)" Plants 11, no. 13: 1779. https://doi.org/10.3390/plants11131779