
BnKAT2 Positively Regulates the Main Inflorescence Length and Silique Number in Brassica napus by Regulating the Auxin and Cytokinin Signaling Pathways


 , ,
, ,
Abstract
:1. Introduction
2. Results
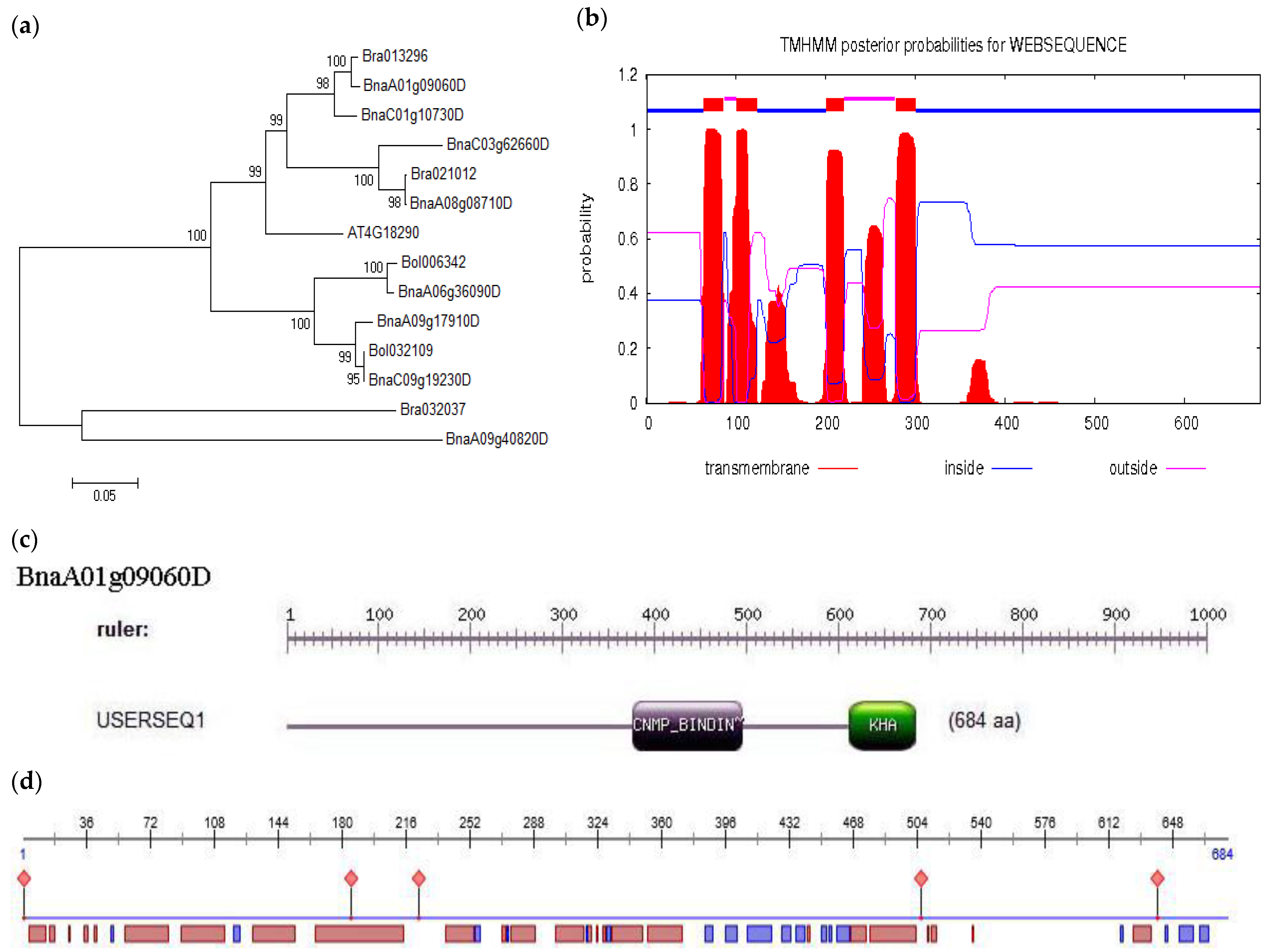
2.1. Cloning and Bioinformatics Analysis of BnKAT2 in B. napus
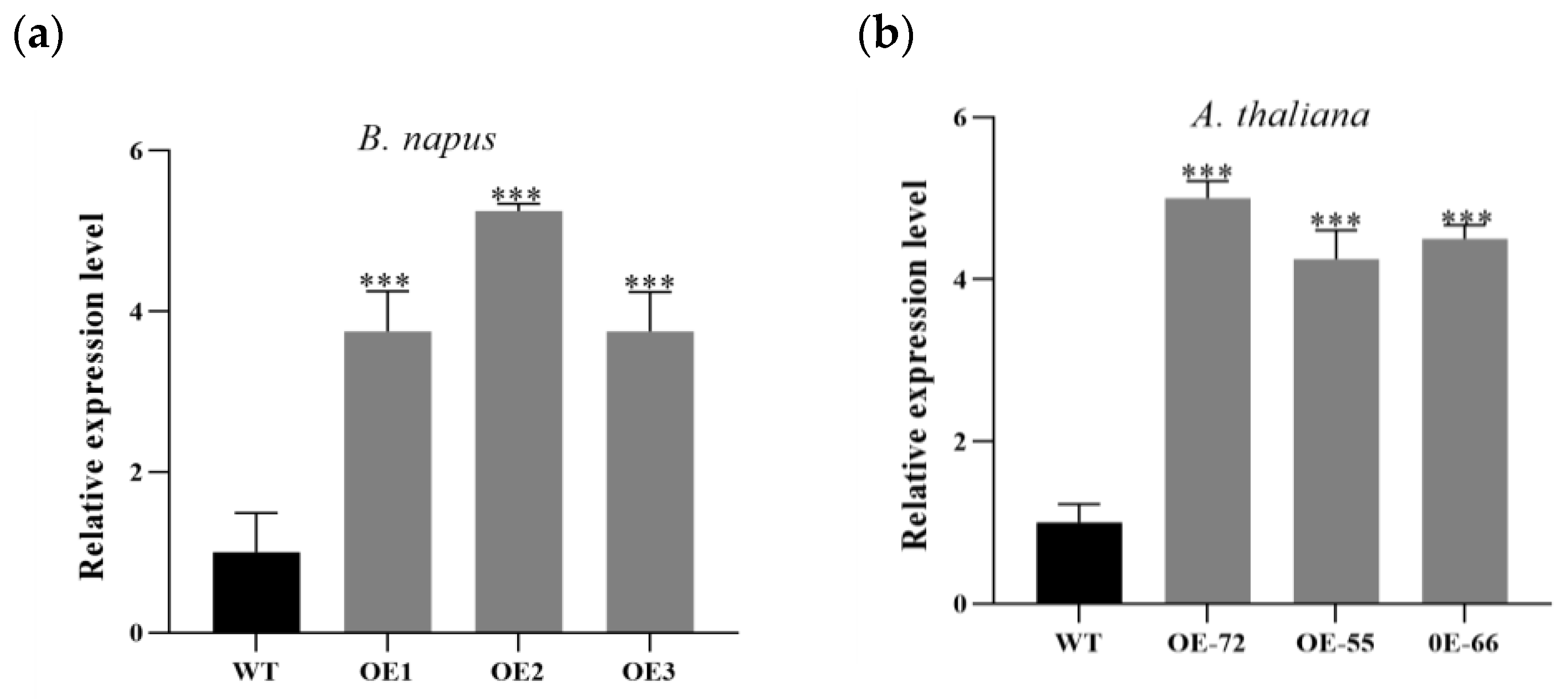
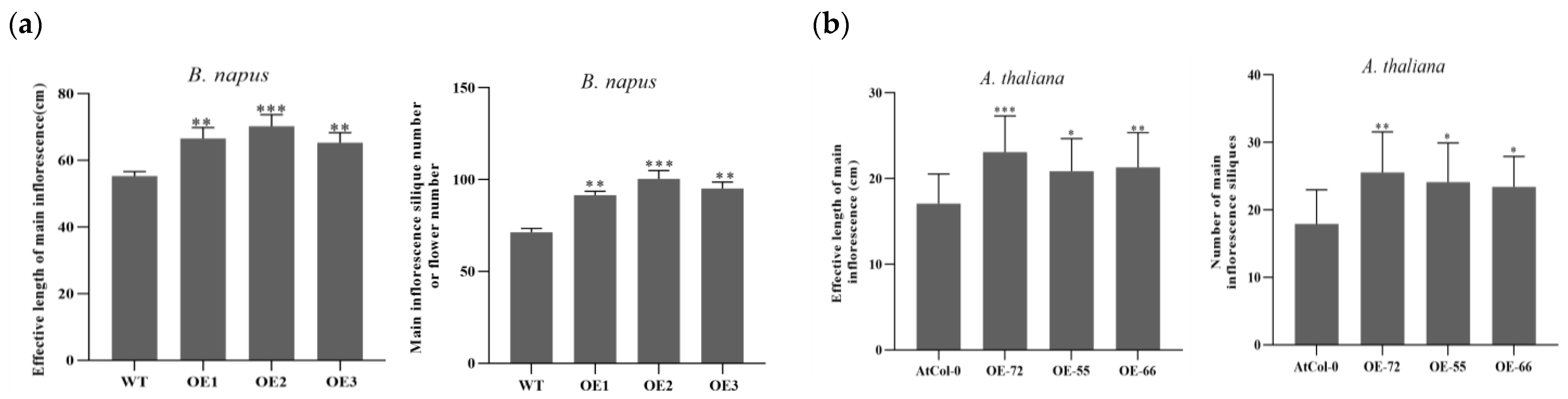
2.2. Improvement Effects of BnKAT2 on the Effective Inflorescence Length and Silique Number
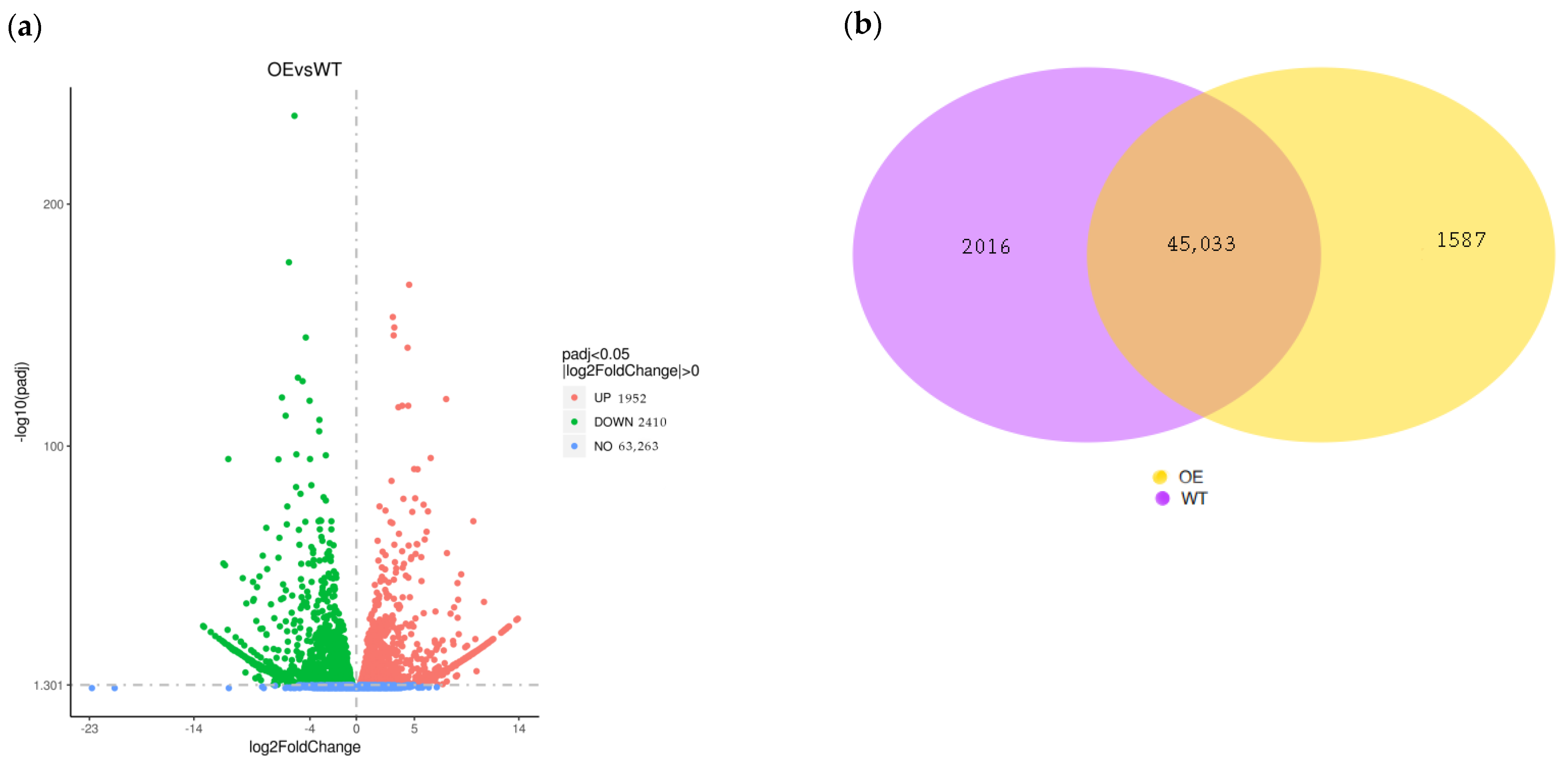
2.3. Analysis of Differentially Expressed Genes
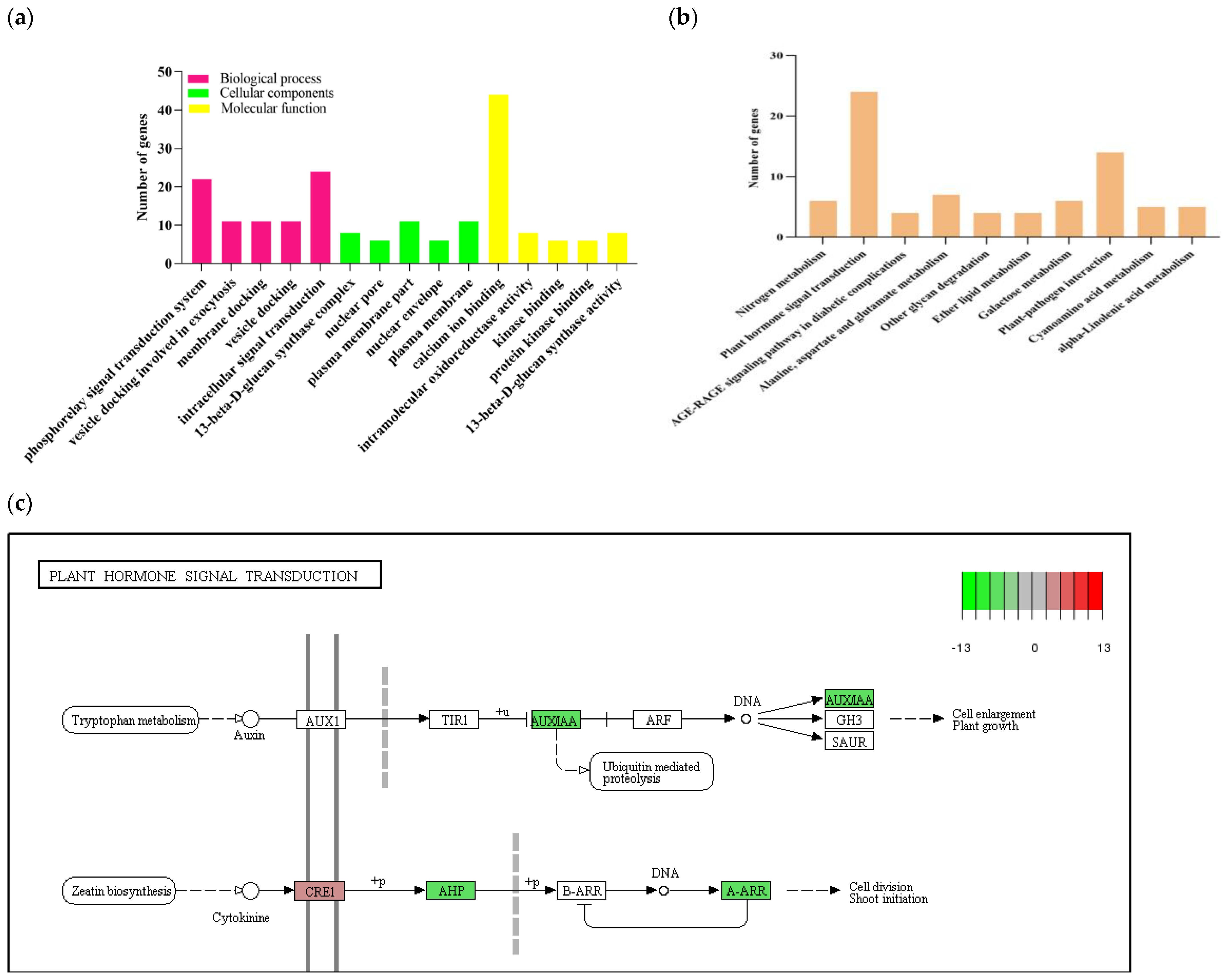
2.4. GO and KEGG Enrichment Analysis on the DEGs
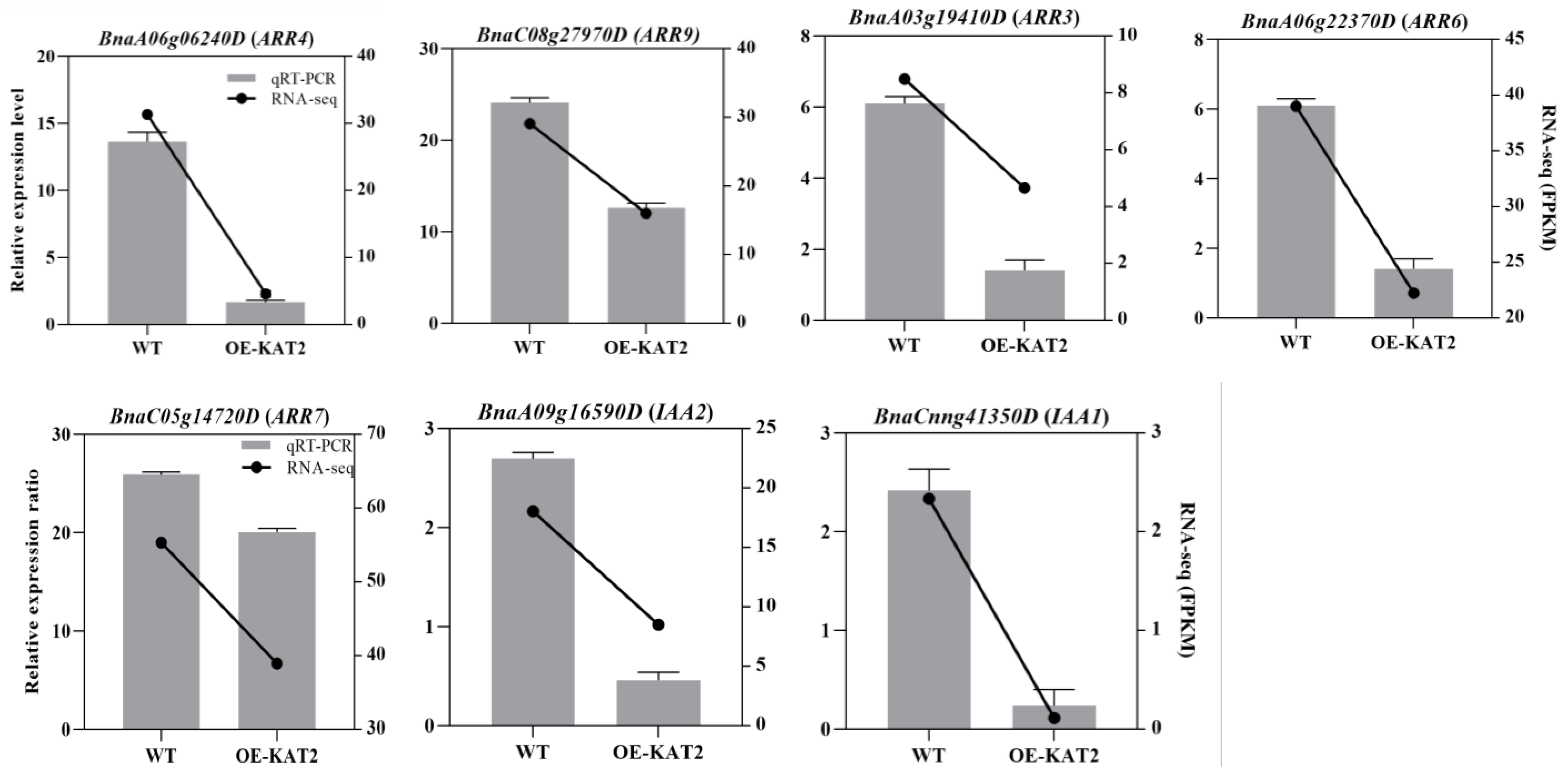
2.5. DEGs Verification Using qRT-PCR
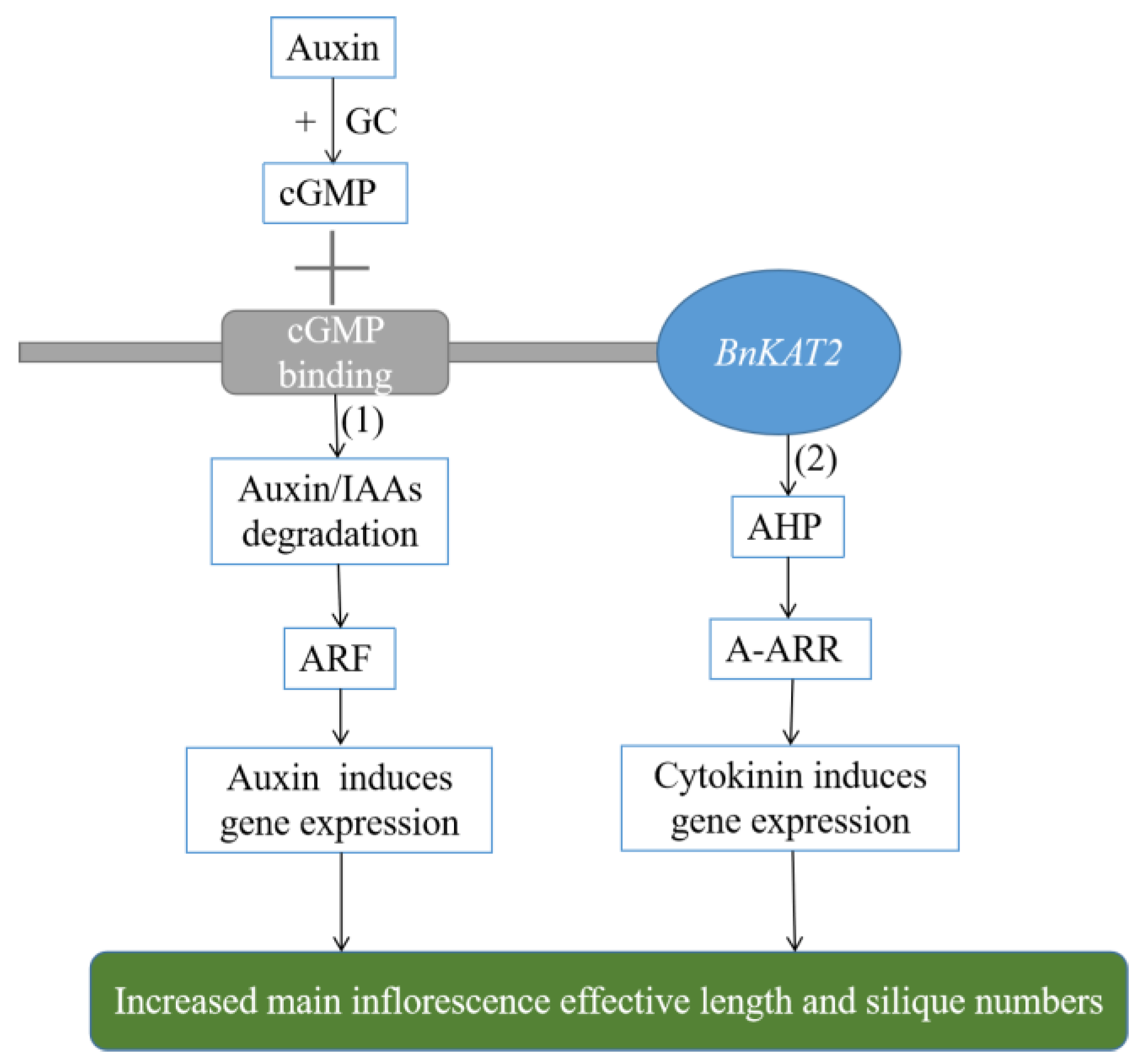
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growing Conditions
4.2. Cloning of BnKAT2 in B. napus
4.3. Bioinformatics Analysis
4.4. Plasmid Construction and Plant Transformation
4.5. Phenotypic Examination of the Transgenic Plants
4.6. RNA-Seq
4.7. QRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prakash, S.; Wu, X.; Bhat, S.R. History, evolution and domestication of Brassica crops. Plant Breed. Rev. 2012, 35, 19–84. [Google Scholar] [CrossRef]
- Li, H.; Feng, H.; Guo, C.; Yang, S.; Huang, W.; Xiong, X.; Liu, J.; Chen, G.; Liu, Q.; Xiong, L.; et al. High-throughput phenotyping accelerates the dissection of the dynamic genetic architecture of plant growth and yield improvement in rapeseed. Plant Biotechnol. J. 2020, 18, 2345–2353. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Shu, C.; Lin, C.; Xu, J.; Liu, K. Identification of a major QTL for silique length and seed weight in oilseed rape (Brassica napus L.). Theor. Appl. Genet. 2012, 125, 285–296. [Google Scholar] [CrossRef]
- Cai, G.; Yang, Q.; Chen, H.; Yang, Q.; Zhang, C.; Fan, C.; Zhou, Y. Genetic dissection of plant architecture and yield-related traits in Brassica napus. Sci. Rep. 2016, 16, 21625. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.L.; Zhao, J.X.; Xiang, Y.; Ren, T.B.; Cheng, G.P. Hybrid Brassica napus: Correlation analysis of main inflorescence length and plant traits. Chin. Agric. Sci. Bull. 2018, 34, 42–48. [Google Scholar]
- Tuncturk, M.; Ciftci, V. Relationship between yield and some yield components in rapeseed (Brassica napus ssp. oleifera L.) cultivars by using correlation and path analysis. Pak. J. Bot. 2007, 39, 81–84. [Google Scholar] [CrossRef]
- Ma, N.; Yuan, J.; Li, M.; Li, J.; Zhang, L.; Liu, L.; Naeem, M.S.; Zhang, C. Ideotype population exploration: Growth, photosynthesis, and yield components at different planting densities in winter oilseed rape (Brassica napus L.). PLoS ONE 2014, 9, e114232. [Google Scholar] [CrossRef]
- Khan, S.; Anwar, S.; Kuai, J.; Ullah, S.; Fahad, S.; Zhou, G. Optimization of nitrogen rate and planting density for improving uield, nitrogen use efficiency, and lodging resistance in oilseed rape. Front. Plant Sci. 2017, 8, 532. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.Y.; Cui, C.; Wang, Q.; Tang, Z.L.; Xu, X.F.; Ling, N.; Ying, J.M.; Li, J.N.; Zhou, Q.Y. Genome-Wide Association Analysis of silique density on racemes and its component traits in Brassica napus. Sci. Agric. Sin. 2018, 51, 1014–1033. [Google Scholar]
- Phillips, A.L.; Huttly, A.K. Cloning of two gibberellin-regulated cDNAs from A. thaliana by subtractive hybridization: Expression of the tonoplast water channel, gamma-TIP, is increased by GA3. Plant Mol. Biol. 1994, 24, 603–615. [Google Scholar] [CrossRef]
- Shi, J.X.; Malitsky, S.; De Oliveira, S.; Branigan, C.; Franke, R.B.; Schreiber, L.; Aharoni, A. SHINE transcription factors act redundantly to pattern the archetypal surface of Arabidopsis thaliana Flower Organs. PLoS Genet. 2011, 7, e1001388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; He, Y.; Wang, Y.; Zhao, S.; Chen, X.; Ye, T.; Wu, Y.; Wu, Y.A. thaliana PLC2 is involved in auxin-modulated reproductive development. Plant J. 2015, 84, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Di Fino, L.M.; D’Ambrosio, J.M.; Tejos, R.; van Wijk, R.; Lamattina, L.; Munnik, T.; Pagnussat, G.C.; Laxalt, A.M. Arabidopsis thaliana phosphatidylinositol-phospholipase C2 (PLC2) is required for female gametogenesis and embryo development. Planta 2016, 245, 717–728. [Google Scholar] [CrossRef] [Green Version]
- Han, D.P.; Zhou, C.; Zheng, Y.Z.; Fu, D.J.; Zhou, Q.H. Genome-wide association analysis of main inflorescence traits in Brassica napus. J. Nucl. Agric. Sci. 2018, 32, 463–476. [Google Scholar]
- Zhu, J.; Zhang, G.Q.; Chang, Y.H.; Chang, Y.H.; Li, X.C.; Yang, J.; Huang, X.Y.; Yu, Q.B.; Chen, H.; Wu, T.L.; et al. AtMYB103 is a crucial regulator of several pathways affecting Arabidopsis thaliana anther development. Sci. China Life Sci. 2010, 53, 1112–1122. [Google Scholar] [CrossRef]
- Guo, X.; Lu, W.; Ma, Y.; Qin, Q.; Hou, S. The BIG gene is required for auxin-mediated organ growth in Arabidopsis thaliana. Planta 2013, 237, 1135–1147. [Google Scholar] [CrossRef]
- Yang, P.; Smalle, J.; Lee, S.; Yan, N.; Emborg, T.J.; Vierstra, R.D. Ubiquitin C-terminal hydrolases 1 and 2 affect shoot architecture in Arabidopsis thaliana. Plant J. 2007, 51, 441–457. [Google Scholar] [CrossRef]
- Lebaudy, A.; Vavasseur, A.; Hosy, E.; Dreyer, I.; Leonhardt, N.; Thibaud, J.B.; Véry, A.A.; Simonneau, T.; Sentenac, H. Plant adaptation to fluctuating environment and biomass production are strongly dependent on guard cell potassium channels. Proc. Natl. Acad. Sci. USA 2008, 105, 5271–5276. [Google Scholar] [CrossRef] [Green Version]
- Lebaudy, A.; Pascaud, F.; Véry, A.A.; Alcon, C.; Dreyer, I.; Thibaud, J.B.; Lacombe, B. Preferential KAT1-KAT2 heteromerization determines inward K+ current properties in Arabidopsis thaliana guard cells. J. Biol. Chem. 2010, 285, 6265–6274. [Google Scholar] [CrossRef] [Green Version]
- Özer, H.; Oral, E.; Dogru, U. Relationships between yield and yield components on currently improved spring rapeseed cultivars. Turk. J. Agric. For. 1999, 23, 603–607. [Google Scholar]
- Chen, W.; Zhang, Y.; Liu, X.; Chen, B.; Tu, J.; Tingdong, F. Detection of QTL for six yield-related traits in oilseed rape (Brassica napus) using DH and immortalized F(2) populations. Theor. Appl. Genet. 2007, 115, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCF (TIRI)-dependent degradation of AUX, IAA proteins. Nature 2001, 414, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Abel, S.; Nguyen, M.D.; Theologis, A. The PS-IAA4/5-like family of early auxin-inducible mRNAs in Arabidopsis thaliana. J. Mol. Biol. 1995, 251, 533–549. [Google Scholar] [CrossRef]
- Reed, J.W. Roles and activities of Aux/IAA proteins in Arabidopsis thaliana. Trends Plant Sci. 2001, 6, 420–425. [Google Scholar] [CrossRef]
- Ross, J.J.; O’Neill, D.P.; Wolbang, C.M.; Symons, G.M.; Reid, J.B. Auxin- gibberell in interactions and their role in plant growth. J. Plant Growth Regul. 2001, 20, 336–353. [Google Scholar] [CrossRef]
- Yin, C.; Gan, L.; Ng, D.; Zhou, X.; Xia, K. Decreased panicle derived indole- 3- acetic acid reduces gibberellin A1 level in the uppermost internode, causing panicle enclosure in male sterile rice Zhenshan 97A. J. Exp. Bot. 2007, 58, 2441–2449. [Google Scholar] [CrossRef] [Green Version]
- Chandler, J.; Werr, W. DORNRÖSCHEN, DORNRÖSCHEN-LIKE, and PUCHI redundantly control floral meristem identity and organ initiation in Arabidopsis. J. Exp. Bot. 2017, 68, 3457–3472. [Google Scholar] [CrossRef]
- Su, Z.; Zhao, L.; Zhao, Y.; Li, S.; Won, S.; Cai, H.; Wang, L.; Li, Z.; Chen, P.; Qin, Y. The THO complex non-cell-autonomously represses female germline specification through the TAS3-ARF3 module. Curr. Biol. 2017, 27, 1597–1609. [Google Scholar] [CrossRef] [Green Version]
- Krizek, B.A. Auxin regulation of A. thaliana flower development involves members of the AINTEGUMENTA-LIKE/PLETHORA (AIL/PLT) family. J. Exp. Bot. 2011, 62, 3311–3319. [Google Scholar] [CrossRef] [Green Version]
- Isner, J.C.; Nühse, T.; Maathuis, F.J. The cyclic nucleotide cGMP is involved in plant hormone signaling and alters phosphorylation of Arabidopsis thaliana root proteins. J. Exp. Bot. 2012, 63, 3199–3205. [Google Scholar] [CrossRef] [Green Version]
- Nan, W.; Wang, X.; Yang, L.; Hu, Y.; Wei, Y.; Liang, X.; Mao, L.; Bi, Y. Cyclic GMP is involved in auxin signaling during Arabidopsis thaliana root growth and development. J. Exp. Bot. 2014, 65, 1571–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehring, C.; Turek, I.S. Cyclic nucleotide monophosphates and their cyclases in plant signaling. Front. Plant Sci. 2017, 8, 1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- To, J.P.; Haberer, G.; Ferreira, F.J.; Deruere, J.; Mason, M.G.; Schaller, G.E.; Alonso, J.M.; Ecker, J.R.; Kieber, J.J. Type-A Arabidopsis thaliana response regulators are partially redundant negative regulators of cytokinin signaling. Plant Cell 2004, 16, 658–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiba, T.; Aoki, K.; Sakakibara, H.; Mizuno, T. Arabidopsis thaliana response regulator, ARR22, ectopic expression of which results in phenotypes similar to the wol cytokinin-receptor mutant. Plant Cell Physiol. 2004, 45, 1063–1077. [Google Scholar] [CrossRef]
- Leibfried, A.; To, J.P.; Busch, W.; Stehling, S.; Kehle, A.; Demar, M.; Kieber, J.J.; Lohmann, J.U. WUSCHEL controls meristem function by direct regulation of cytokinin-inducible response regulators. Nature 2005, 438, 1172–1175. [Google Scholar] [CrossRef]
- Andersen, S.U.; Kieber, J.J.; Lohmann, J.U. Role of A-type ARABIDOPSIS RESPONSE REGULATORS in meristem maintenance and regeneration. Eur. J. Cell Biol. 2010, 89, 279–284. [Google Scholar] [CrossRef]
- Giulini, A.; Wang, J.; Jackson, D. Control of phyllotaxy by the cytokinin-inducible response regulator homologue ABPHYL1. Nature 2004, 430, 1031–1034. [Google Scholar] [CrossRef]
- Mahonen, A.P.; Bishopp, A.; Higuchi, M.; Nieminen, K.M.; Kinoshita, K.; Tormakangas, K.; Keda, Y.; Oka, A.; Kakimoto, T.; Helariutta, Y. Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 2006, 311, 94–98. [Google Scholar] [CrossRef]
- Schwarz, I.; Scheirlinck, M.T.; Otto, E.; Bartrina, I.; Schmidt, R.C.; Schmülling, T. Cytokinin regulates the activity of the inflorescence meristem and components of seed yield in oilseed rape. J. Exp. Bot. 2020, 71, 7146–7159. [Google Scholar] [CrossRef]
- Werner, S.; Bartrina, I.; Novák, O.; Strnad, M.; Werner, T.; Schmülling, T. The cytokinin status of the epidermis regulates aspects of vegetative and reproductive development in Arabidopsis thaliana. Front. Plant Sci. 2021, 23, 613488. [Google Scholar] [CrossRef]
- Powell, G.K.; Hommes, N.G.; Kuo, J.; Castle, L.A.; Morris, R.O. Inducible expression of cytokinin biosynthesis in Agrobacterium tumefaciens by plant phenolics. Mol. Plant Microbe Interact. 1988, 1, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Besnard, F.; Rozier, F.; Vernoux, T. The AHP6 cytokinin signaling inhibitor mediates an auxin-cytokinin crosstalk that regulates the timing of organ initiation at the shoot apical meristem. Plant Signal. Behav. 2014, 9, 1559–2324. [Google Scholar] [CrossRef] [Green Version]
- McLean, R.M.; Wang, N.X. Potassium. Adv. Food Nutr. Res. 2021, 96, 89–121. [Google Scholar] [CrossRef] [PubMed]
- Thiel, G.; Blatt, M.R.; Fricker, M.D. Modulation of K+ channels in Vicia stomatal guard cells by peptide homologs to the auxin binding protein C-terminus. Proc. Natl. Acad. Sci. USA 1994, 90, 11493–11497. [Google Scholar] [CrossRef] [Green Version]
- Blatt, M.R.; Grabov, A. Signal redundancy, gates and integration in the control of ion channels for stomatal movement. J. Exp. Bot. 1997, 48, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Senn, A.P.; Goldsmith, M.H. Regulation of electro-genic proton pumping by auxin and fusicoccin as related to the growth of Avena coleoptiles. Plant Physiol. 2019, 88, 131–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frias, I.; Caldeira, M.T.; Perez-Castineira, J.R.; Navarro-Avino, J.P.; Culianez-Macia, F.A.; Kuppinger, O.; Stransky, H.; Pagès, M.; Hager, A.; Serrano, R. A major isoform of the maize plasma membrane H+-ATPase: Characterization and induction by auxin in coleoptiles. Plant Cell 1996, 8, 1533–1544. [Google Scholar] [CrossRef] [Green Version]
- Christian, M.; Steffens, B.D. How does auxin enhance cell elongation? roles of auxin-binding proteins and potassium channels in growth control. Plant Biol. 2010, 8, 346–352. [Google Scholar] [CrossRef]
- Becker, D.; Hedrich, R. Channelling auxin action: Modulation of ion transport by indole-3-acetic acid. Plant Mol. Biol. 2002, 49, 349–356. [Google Scholar] [CrossRef]
- Hager, A. Role of the plasma membrane H+-ATPase in auxin-induced elongation growth: Historical and new aspects. J. Plant Res. 2003, 116, 483–505. [Google Scholar] [CrossRef]
- Chen, L.; Chen, L.; Zhang, X.; Liu, T.; Niu, S.; Wen, J.; Yi, B.; Ma, C.; Tu, J.; Fu, T.; et al. Identification of miRNAs that regulate silique development in Brassica napus. Plant Sci. 2018, 269, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.H.; Cheng, Y.; Ma, S.J.; He, C.; Zhen, W.Y.; Zhou, K.J.; Ma, Y.S. Genetics of effective silique number on main inflorescence of new germplasm 12R1402(Brassica napus L.). Chin. J. Oil Crop Sci. 2016, 38, 287–291. [Google Scholar]
- Wilmowicz, E.; Ku’ko, A.; Burchardt, S.; Przywieczerski, T. Molecular and hormonal aspects of drought-triggered flower shedding in yellow lupine. Int. J. Mol. Sci. 2019, 20, 3731. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wan, Y.; Meng, X.; Zhang, X.; Yao, M.; Miu, W.; Zhu, D.; Yuan, D.; Lu, K.; Li, J.; et al. Genome-wide identification and analysis of MKK and MAPK gene families in Brassica species and response to stress in Brassica napus. Int. J. Mol. Sci. 2021, 22, 544. [Google Scholar] [CrossRef] [PubMed]
- Lough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, W.H. Potassium transport and signaling in higher plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, 480–484. [Google Scholar] [CrossRef]
- Arocho, A.; Chen, B.; Ladanyi, M.; Pan, Q. Validation of the 2-DeltaDeltaCt calculation as an alternate method of data analysis for quantitative PCR of BCR-ABL P210 transcripts. Diagn. Mol. Pathol. 2006, 15, 56–61. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| BnKAT2 | ATGTCAATCTCTTGCACTAGAAACTTCTTT | TCAAGAGTCTATGCTTTCAAGCTCAC |
| BnKAT2q | CAACATTGTCAATGGCTTCTTTGC | TGGAATGGAGCTGTGGAGCAGA |
| ACT7 | TGGGTTTGCTGGTGACGAT | ACGGATCCTAGCAGCTCAGATGTTGA |
| OE-BnKAT2 | CACCATGTCAATCTCTTGCACTAGAAACTTCTTTG | AGAGTCTATGCTTTCAAGCTCACTG |
| 35S3NDF-RPA3ND | GGAAGTTCATTTCATTTGGAGAG | AGAGTCTATGCTTTCAAGCTCACTG |
| Bar | CGACATCCGCCGTGCCACCGA | CAAATCTCGGTGACGGGCAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, D.; Zhang, Y.; Wang, Z.; Qu, C.; Zhu, D.; Wan, H.; Liang, Y. BnKAT2 Positively Regulates the Main Inflorescence Length and Silique Number in Brassica napus by Regulating the Auxin and Cytokinin Signaling Pathways. Plants 2022, 11, 1679. https://doi.org/10.3390/plants11131679
Yuan D, Zhang Y, Wang Z, Qu C, Zhu D, Wan H, Liang Y. BnKAT2 Positively Regulates the Main Inflorescence Length and Silique Number in Brassica napus by Regulating the Auxin and Cytokinin Signaling Pathways. Plants. 2022; 11(13):1679. https://doi.org/10.3390/plants11131679
Chicago/Turabian StyleYuan, Dashuang, Yin Zhang, Zhen Wang, Cunmin Qu, Dongming Zhu, Huafang Wan, and Ying Liang. 2022. "BnKAT2 Positively Regulates the Main Inflorescence Length and Silique Number in Brassica napus by Regulating the Auxin and Cytokinin Signaling Pathways" Plants 11, no. 13: 1679. https://doi.org/10.3390/plants11131679