
Seed Germination Ecology of Semiparasitic Weed Pedicularis kansuensis in Alpine Grasslands


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Seed Morphology, Mass, and Imbibition
2.2. Effect of Temperature and Light on Seed Germination
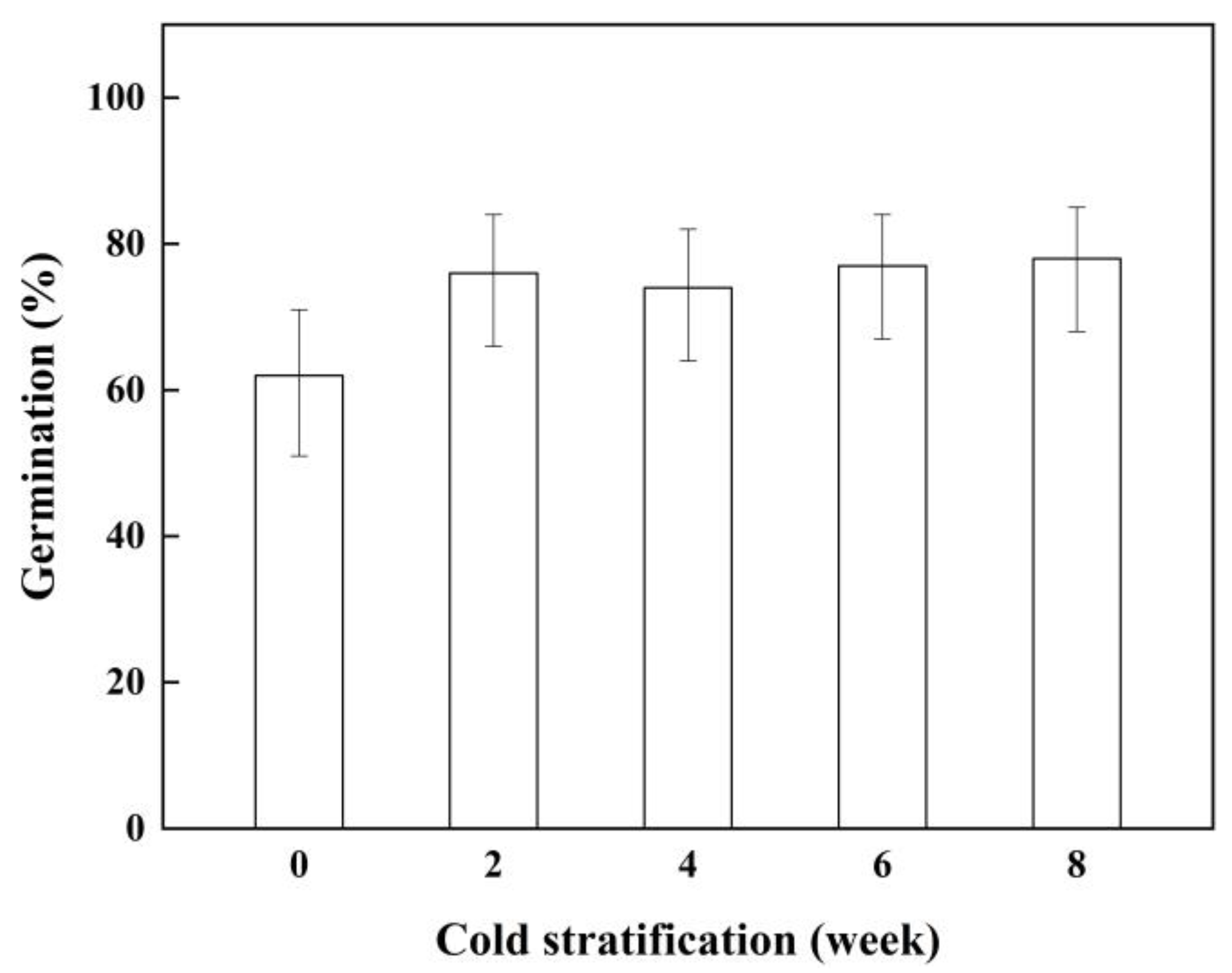
2.3. Effect of Cold Stratification on Seed Germination
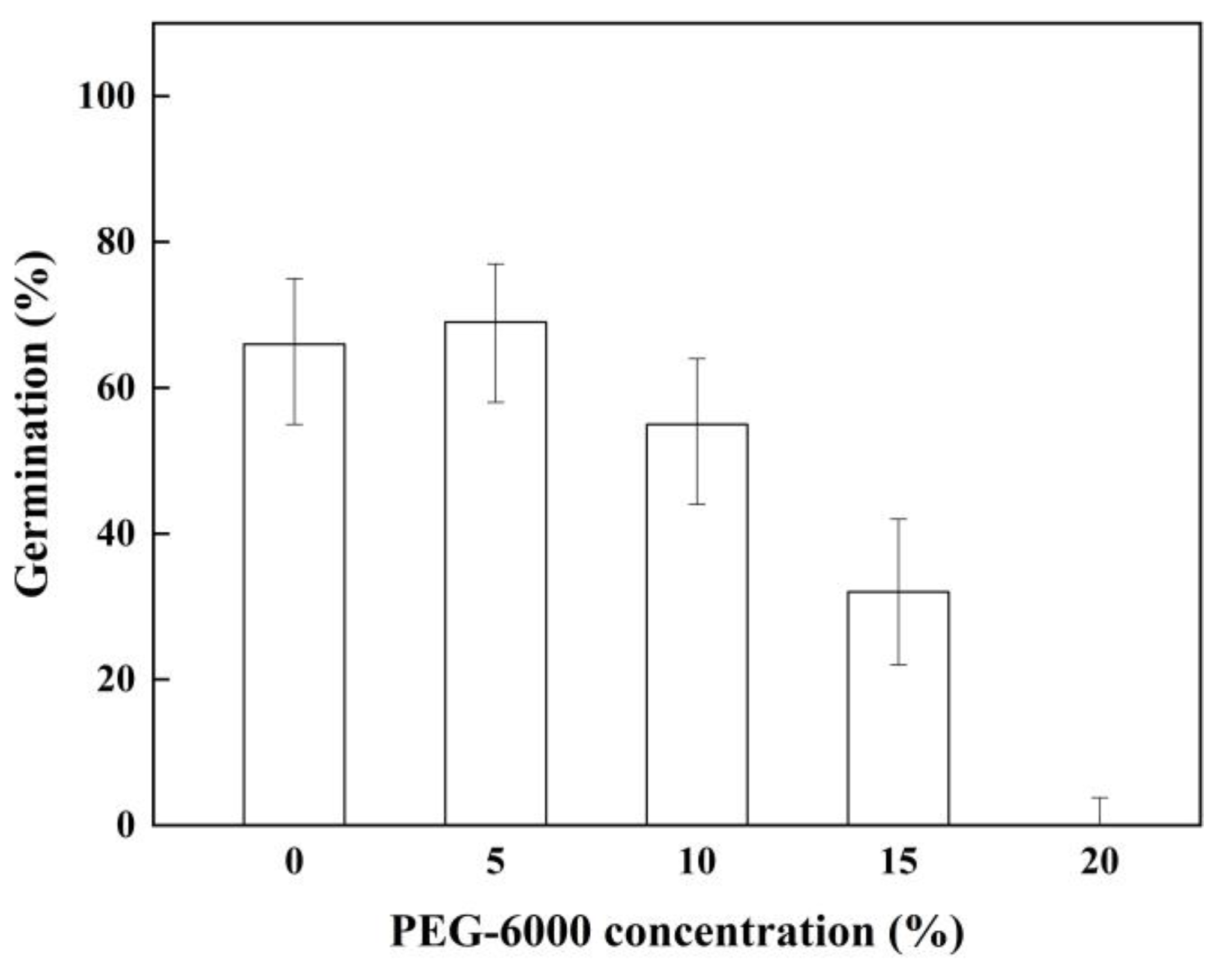
2.4. Effect of Drought Stress on Seed Germination
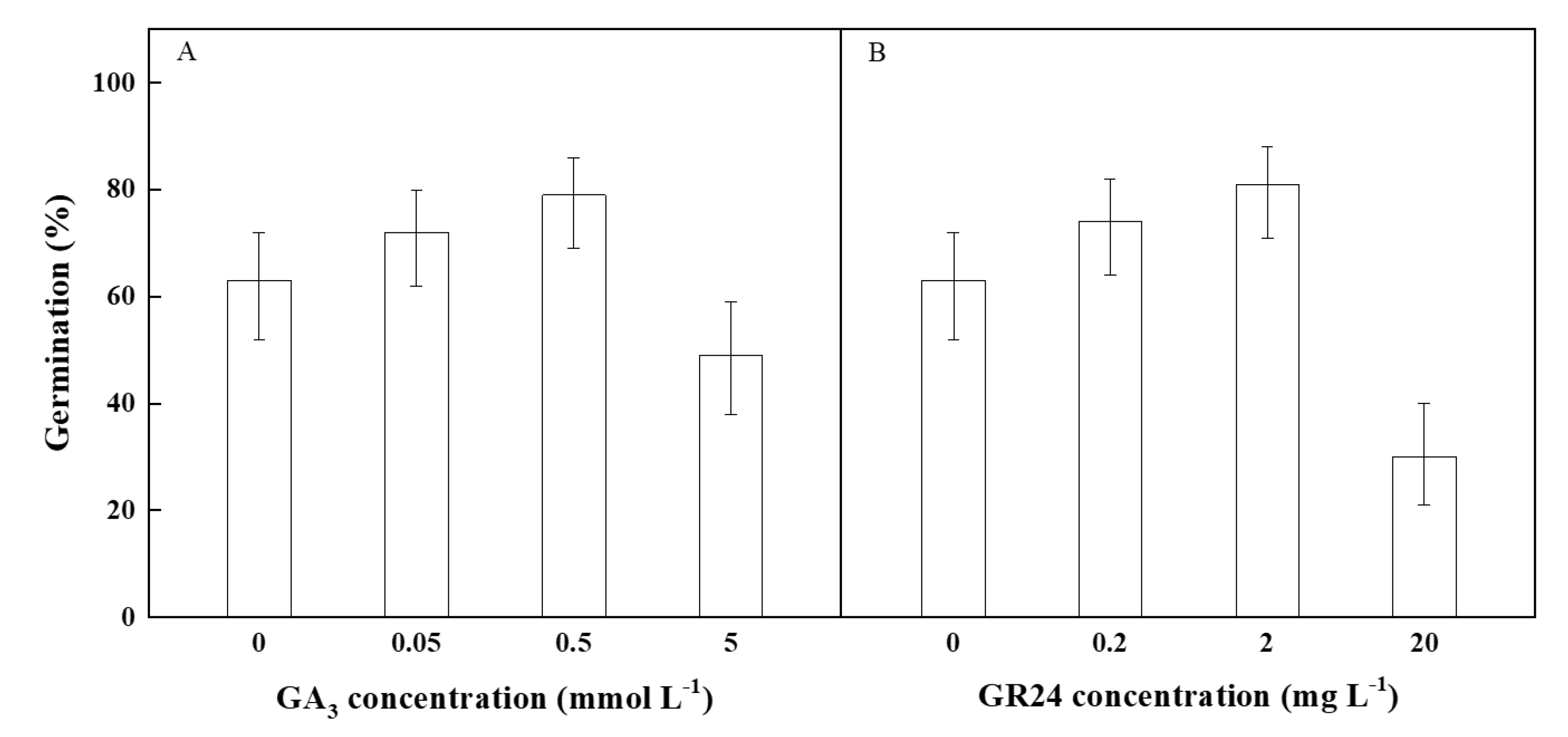
2.5. Effect of Plant Hormones on Seed Germination
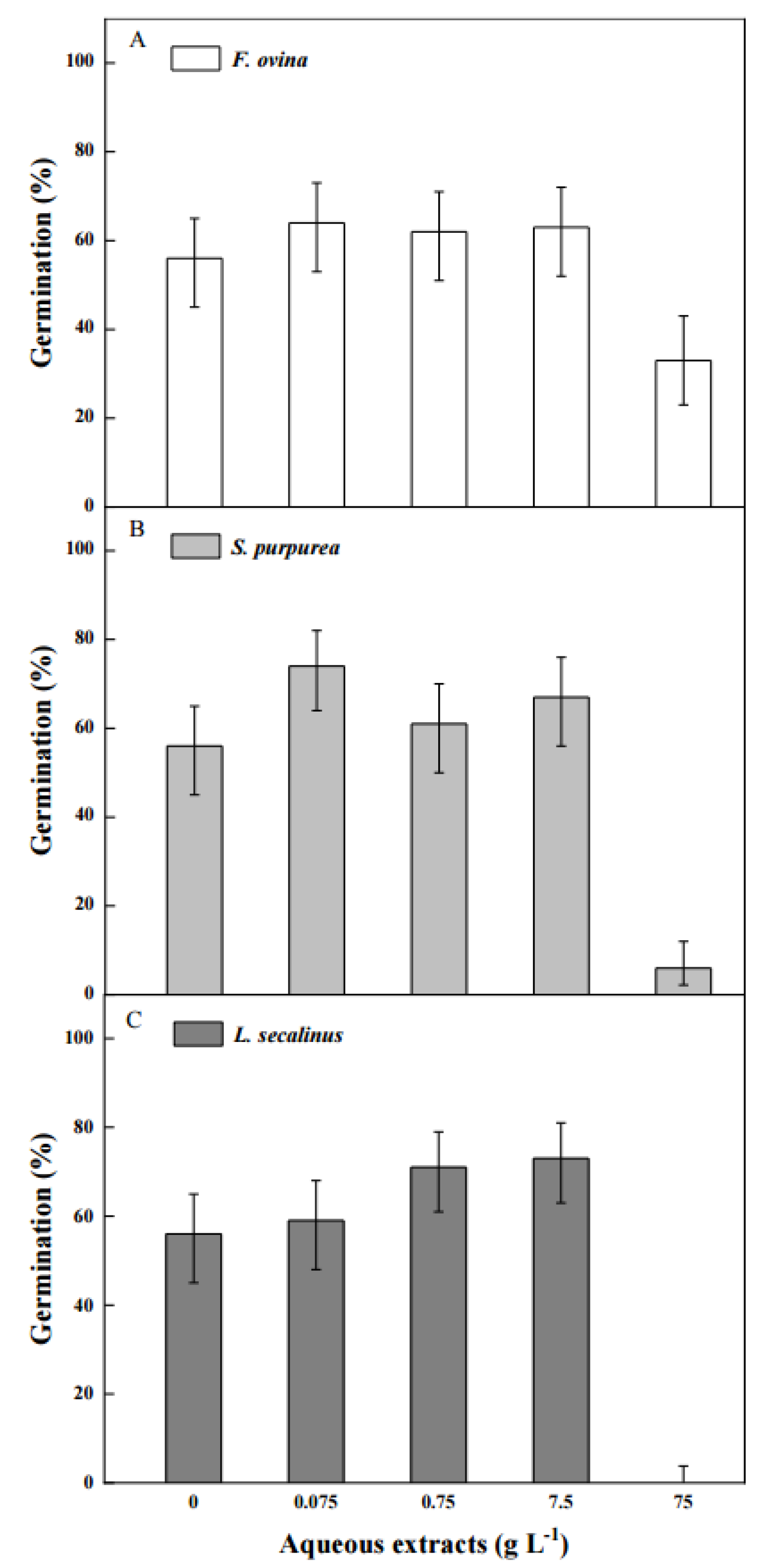
2.6. Effect of Aqueous Extracts of Native Dominant Plants on Seed Germination
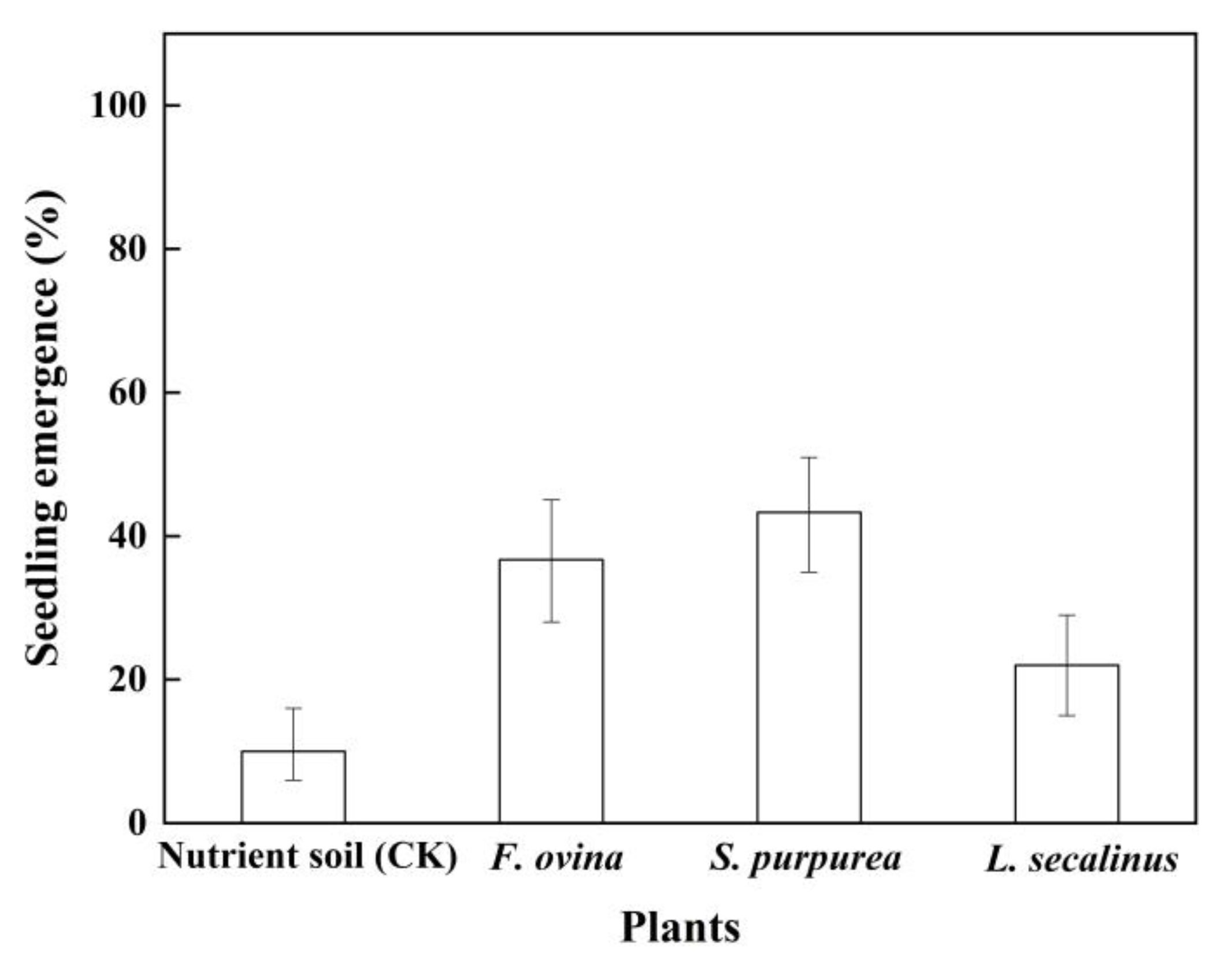
2.7. Effect of Transplantation of Native Dominant Plants on Seedling Emergence
3. Discussion
4. Materials and Methods
4.1. Seed Collection
4.2. Seed Morphology, Mass, and Imbibition
4.3. General Seed-Germination Test
4.4. Effect of Temperature and Light on Seed Germination
4.5. Effect of Cold Stratification on Seed Germination
4.6. Effect of Drought Stress on Seed Germination
4.7. Effect of Plant Hormones on Seed Germination
4.8. Effect of Aqueous Extracts of Native Dominant Plants on Seed Germination
4.9. Transplantation of Native Dominant Plants on Seedling Emergence
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hua, L.M.; Squires, V.R. Managing China’s pastoral lands, Current problems and future prospects. Land Use Policy 2015, 43, 129–137. [Google Scholar] [CrossRef]
- Zhao, W.H.; Li, J.L.; Qi, J.G.; Liu, X. Analysis on the problems and actuality of the steppe ecological security in Xinjiang and the solving measures. Arid Zone Res. 2005, 22, 45–50, (In Chinese with English abstract). [Google Scholar]
- Liu, Y.Y. Ecological Factors of P. kansuensis Maxim. Expansion in Bayanbulak Grassland. Ph.D. Dissertation, Xinjiang University, Urumqi, China, 2018. (In Chinese with English abstract). [Google Scholar]
- Sui, X.L.; Patrick, K.; Li, W.J.; Yang, M.Q.; Guan, K.Y.; Li, A.R. Identity and distribution of weedy Pedicularis kansuensis Maxim. (Orobanchaceae) in Tianshan Mountains of Xinjiang, morphological; anatomical and molecular evidence. J. Arid Land 2016, 8, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Majewska, M.L.; Rola, K.; Stefanowicz, A.M.; Nobis, M.; Baszkowski, J.; Zubek, S. Do the impacts of alien invasive plants differ from expansive native ones? An experimental study on arbuscular mycorrhizal fungi communities. Biol. Fertil. Soils 2018, 54, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Li, W.J.; Sui, X.L.; Kuss, P.; Liu, Y.Y.; Li, A.R.; Guan, K.Y. Long-distance dispersal after the last glacial maximum (LGM) led to the disjunctive distribution of Pedicularis kansuensis (Orobanchaceae) in the Qinghai-Tibetan Plateau and Tianshan Altay region. PLoS ONE 2016, 11, e0165700. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.S. It is an urgent task to recover and comprehensively manage the grassland ecology of Bayanbulak. Chin. J. Agric. Resour. Reg. Plan. 2006, 27, 21–25, (In Chinese with English abstract). [Google Scholar]
- Yan, D.J.; Zhou, Q.W.; Lu, H.; Wu, C.C.; Zhao, B.Y.; Cao, D.D.; Ma, F.; Liu, X.X. The disaster.; ecological distribution and control of poisonous weeds in natural grasslands of Xinjiang Uygur Autonomous Region. Sci. Agric. Sin. 2015, 48, 565–582, (In Chinese with English abstract). [Google Scholar]
- Xu, H.G.; Ding, H.; Li, M.Y.; Sheng, Q.; Guo, J.Y.; Han, Z.M.; Huang, Z.G.; Sun, H.Y.; He, S.P.; Wu, H.R.; et al. The distribution and economic losses of alien species invasion in China. Biol. Invasions 2006, 8, 1495–1500. [Google Scholar] [CrossRef]
- Osipitan, O.A.; Mesgaran, M.B.; Hanson, B.D.; Fernandez-Bayo, J.D. Leveraging tomato crop residues and pomace for biosolarization to deplete weed seedbank in tomato fields. Agric. Sci. Technol. 2022, 2, 22–31. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Gill, G.; Preston, C. Influence of environmental factors on seed germination and seedling emergence of rigid ryegrass (Lolium rigidum). Weed Sci. 2006, 54, 1004–1012. [Google Scholar] [CrossRef]
- Cohen, O.; Riov, J.; Katan, J.; Gamliel, A.; Bar, P. Reducing persistent seed banks of invasive plants by soil solarization—The case of Acacia saligna. Weed Sci. 2008, 56, 860–865. [Google Scholar] [CrossRef]
- Abbas, T.; Zahir, A.; Naveed, M. Field application of allelopathic bacteria to control invasion of little seed canary grass in wheat. Environ. Sci. Pollut. Res. 2020, 28, 9120–9132. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, A.H.M.; Jhala, A.J.; Singh, M. Factors affecting germination of Citronmelon (Citrullus lanatus var.citroides). Weed Sci. 2014, 62, 45–50. [Google Scholar] [CrossRef]
- Kayanifard, M.; Mohsenzadeh, S. Allelopathic analysis of four ecotypes of Ajowan. Iran. J. Sci. Technol. Trans. A-Sci. 2017, 41, 971–978. [Google Scholar] [CrossRef]
- Yue, Y.H.; Jin, G.L.; Lu, W.H.; Gong, K.; Han, W.Q.; Liu, W.H.; Wu, X.E. Effect of environmental factors on the germination and emergence of drunken horse grass (Achnatherum inebrians). Weed Sci. 2021, 69, 62–68. [Google Scholar] [CrossRef]
- Imad, M.; Idrees, M.; Hadi, F.; Memon, N.H.; Zhang, Z.Y. Allelopathic effect of Parthenium hysterophorus extract on seed germination and seedling growth of selected eight plants. Pak. J. Bot. 2021, 53, 2187–2197. [Google Scholar] [CrossRef]
- Pawłowski, T.A.; Bujarska-Borkowska, B.; Suszka, J.; Tylkowski, T.; Chmielarz, P.; Klupczyńska, E.A.; Staszak, A.M. Temperature regulation of primary and secondary seed dormancy in Rosa canina L., findings from proteomic analysis. Int. J. Mol. Sci. 2020, 21, 7008. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. The control of seed dormancy and germination by temperature; light and nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Pereira, R.G.; Cala, A.; Fernández-Aparicio, M.; Molinillo, J.M.G.; Boaventura, M.A.D.; Macias, F.A. Gibberellic and kaurenoic hybrid strigolactone mimics for seed germination of parasitic weeds. Pest Manag. Sci. 2017, 73, 2529–2537. [Google Scholar] [CrossRef]
- Omoarelojie, L.O.; Kulkarni, M.G.; Finnie, J.F.; Pospisil, T.; Strnad, M.; Van Staden, J. Synthetic strigolactone (rac-GR24) alleviates the adverse effects of heat stress on seed germination and photosystem II function in lupine seedlings. Plant Physiol. Biochem. 2020, 155, 965–979. [Google Scholar] [CrossRef]
- Wu, B.C.; Sun, M.; Zhang, H.; Yang, D.; Lin, C.; Khan, I.; Wang, X.S.; Zhang, X.Q.; Nie, G.; Feng, G.Y.; et al. Transcriptome analysis revealed the regulation of gibberellin and the establishment of photosynthetic system promote rapid seed germination and early growth of seedling in pearl millet. Biotechnol. Biofuels 2021, 14, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Cui, B.C.; Duan, S.S.; Chen, J.J.; Fan, H.; Lu, B.B.; Zheng, J.H. Moving north in China, the habitat of Pedicularis kansuensis in the context of climate change. Sci. Total Environ. 2019, 697, 133979. [Google Scholar] [CrossRef] [PubMed]
- Bao, G.S.; Song, M.L.; Wang, Y.Q.; Saikkonen, K.; Li, C.J. Does Epichloë endophyte enhance host tolerance to root hemiparasite? Microb. Ecol. 2020, 82, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.H.; Hou, Y.; Li, F.; Guo, C.C.; Jia, T.H.; Degen, A.A.; White, A.; Ding, L.M.; Long, R.J. Inhibitory action of allelochemicals from Artemisia Nanschanica to control Pedicularis kansuensis, an annual weed of alpine grasslands. Aust. J. Bot. 2017, 65, 305–314. [Google Scholar] [CrossRef]
- Wang, H.C.; Zhao, J.Z.; Zhou, H.K. Effects of temperature and moisture on seed germination of Pedicularis kansuensis Maxim. J. Anhu Agric. Sci. 2008, 36, 14873–14875, (In Chinese with English abstract). [Google Scholar]
- Sui, X.L.; Li, A.R.; Guan, K.Y. Impacts of climatic changes as well as seed germination characteristics on the population expansion of Pedicularis verticillata. Ecol. Environ. Sci. 2013, 22, 1099–1104, (In Chinese with English abstract). [Google Scholar]
- Baskin, J.M.; Davis, B.H.; Baskin, C.C.; Gleason, S.M.; Cordell, S. Physical dormancy in seeds of Dodonaea viscosa (Sapindales; Sapindaceae) from Hawaii. Seed Sci. Res. 2004, 14, 81–90. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds, Ecology.; Biogeography.; and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014; pp. 1–1586. [Google Scholar]
- Wang, G.Y.; Baskin, C.C.; Baskin, J.M.; Yang, X.J.; Liu, G.F.; Zhang, X.S.; Ye, X.H.; Huang, Z.Y. Timing of seed germination in two alpine herbs on the southeastern Tibetan plateau, the role of seed dormancy and annual dormancy cycling in soil. Plant Soil. 2017, 421, 465–476. [Google Scholar] [CrossRef]
- Nishimoto, R.K.; Mc Carty, L.B. Fluctuating temperature and light influence seed germination of goosegrass (Eleusine indica). Weed Sci. 1997, 45, 426–429. [Google Scholar] [CrossRef]
- Karimi, H.R. Evaluation the Effect of diurnal temperature fluctuations on seed germination of Pistacia vera L. J. Agric. Sci. Technol. B 2011, 1, 616–620. [Google Scholar]
- Camacho, M.E.; Heitman, J.L.; Gannon, T.W.; Amoozegar, A.; Leon, R.G. Seed germination responses to soil hydraulic conductivity and polyethylene glycol (PEG) osmotic solutions. Plant Soil. 2021, 462, 175–188. [Google Scholar] [CrossRef]
- López, A.S.; López, D.R.; Arana, M.V.; Batlla, D.; Marchelli, P. Germination response to water availability in populations of Festuca pallescens along a Patagonian rainfall gradient based on hydrotime model parameters. Sci Rep. 2021, 11, 10653. [Google Scholar] [CrossRef] [PubMed]
- Almodares, A.; Hadi, M.R.; Dosti, B. Effects of salt stress on germination percentage and seedling growth in sweet sorghum cultivars. J. Biol. Sci. 2007, 7, 1492–1495. [Google Scholar] [CrossRef] [Green Version]
- Reza, Y.A.; Rashidi, S.; Moradi, P.; Mastinu, A. Germination and seedling growth responses of Zygophyllum fabago; Salsola kali L. and Atriplex canescens to PEG-induced drought stress. Environments 2020, 7, 10653. [Google Scholar]
- Tafvizi, F.; Tafreshi, S.A.H.; Toluei, Z.; Toghyani, M. Stress responses of the green microalga; Dunaliella salina to PEG-induced drought. J. Mar. Biol. Assoc. UK 2020, 100, 1043–1052. [Google Scholar] [CrossRef]
- Liu, B.H.; Liu, X.H.; Liu, F.C.; Ma, H.L.; Ma, B.Y.; Peng, L. Stress tolerance of Xerocomus badius and its promotion effect on seed germination and seedling growth of annual ryegrass under salt and drought stresses. AMB Express 2021, 11, 15–23. [Google Scholar] [CrossRef]
- Kgosi, R.L.; Zwanenburg, B.; Mwakabok, A.S.; Murdoch, A.J. Strigolactone analogues induce suicidal seed germination of Striga spp. in soil. Weed Res. 2012, 52, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.X.; Zhang, M.; Dong, S.Q.; Ma, Y.Q. Conditioning duration and agents involved in broomrape seeds responding to germination stimulants. Plant Growth Regul. 2017, 81, 221–230. [Google Scholar] [CrossRef]
- Uraguchi, D.; Kuwata, K.; Hijikata, Y.; Yamaguchi, R.; Imaizumi, H.; Sathiyanarayanan, A.M.; Rakers, C.; Mori, N.; Akiyama, K.; Irle, S.; et al. A femtomolar-range suicide germination stimulant for the parasitic plant Striga hermonthica. Science 2019, 362, 1301–1305. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Li, K.; Deng, C.; Gong, Y.; Liu, Y.; Wang, L. Seed Germination Ecology of Semiparasitic Weed Pedicularis kansuensis in Alpine Grasslands. Plants 2022, 11, 1777. https://doi.org/10.3390/plants11131777
Hu J, Li K, Deng C, Gong Y, Liu Y, Wang L. Seed Germination Ecology of Semiparasitic Weed Pedicularis kansuensis in Alpine Grasslands. Plants. 2022; 11(13):1777. https://doi.org/10.3390/plants11131777
Chicago/Turabian StyleHu, Jiedong, Kaihui Li, Chengjun Deng, Yanming Gong, Yanyan Liu, and Lei Wang. 2022. "Seed Germination Ecology of Semiparasitic Weed Pedicularis kansuensis in Alpine Grasslands" Plants 11, no. 13: 1777. https://doi.org/10.3390/plants11131777