
Foliar Application of Salicylic Acid to Mitigate Water Stress in Tomato

 , , , , and
, , , , and 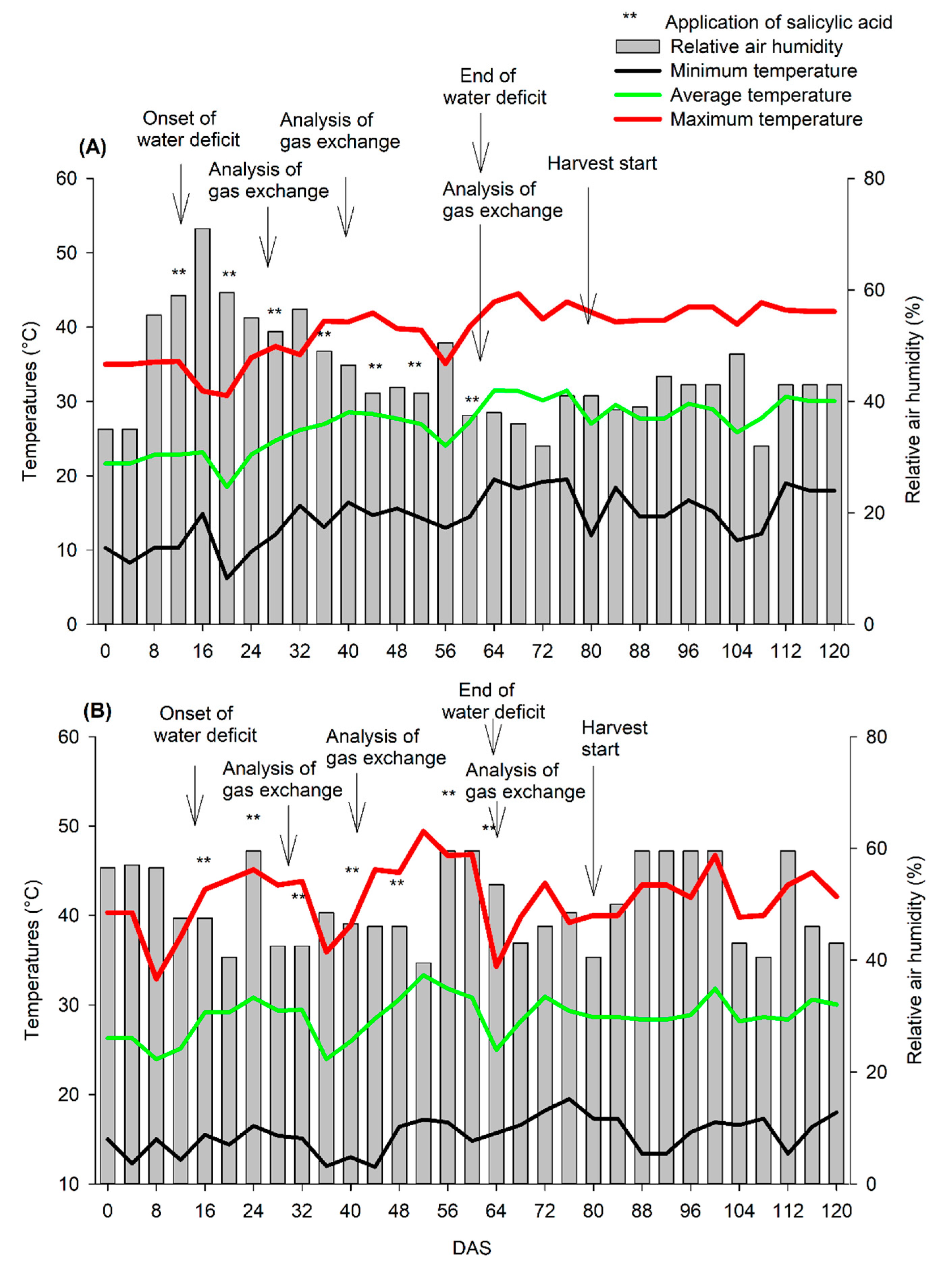{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation Conditions
2.2. Treatments
2.3. Foliar Application
2.4. Gas Exchange
2.5. Productive Characteristics
2.6. Statistical Analysis
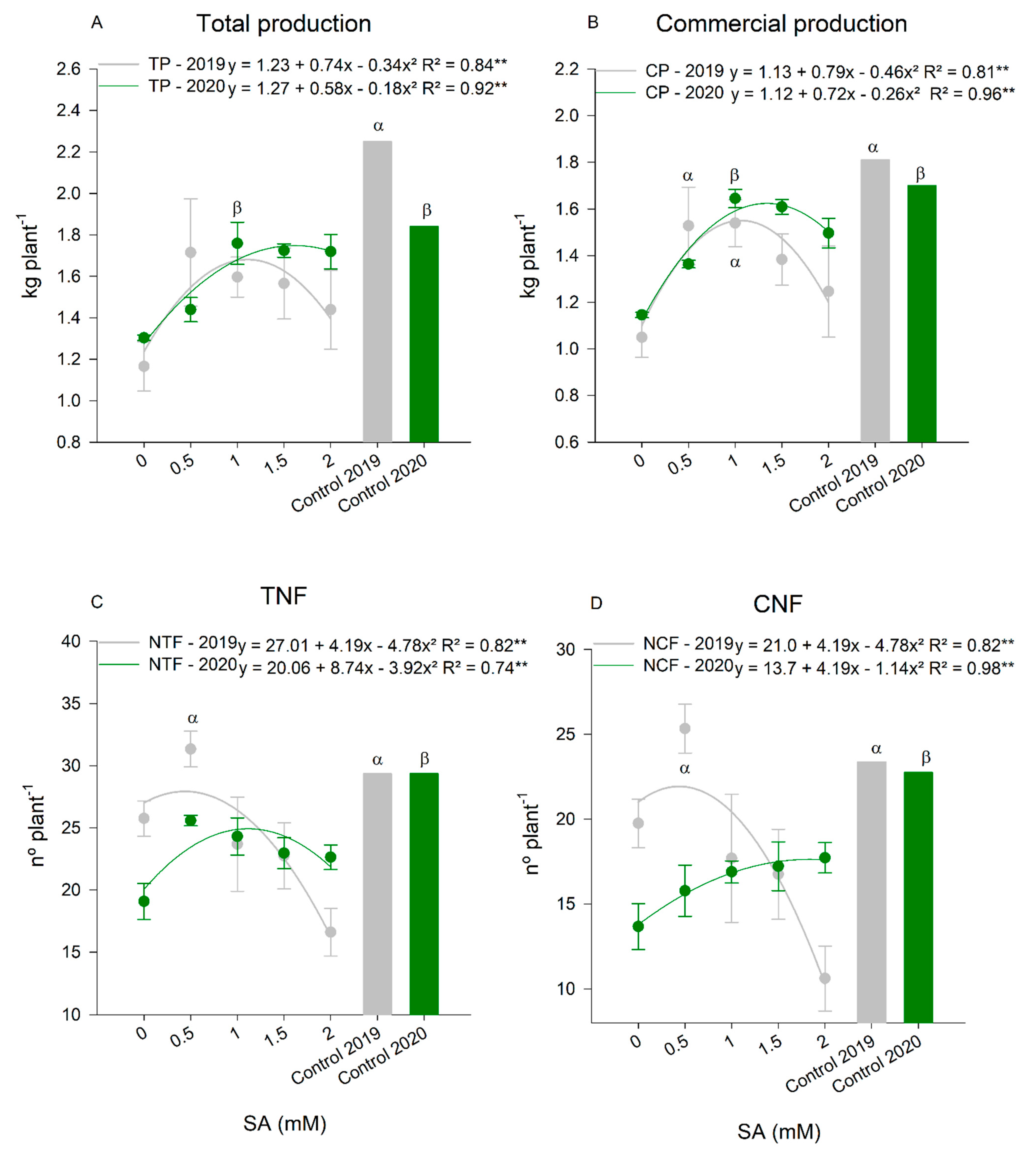
3. Results
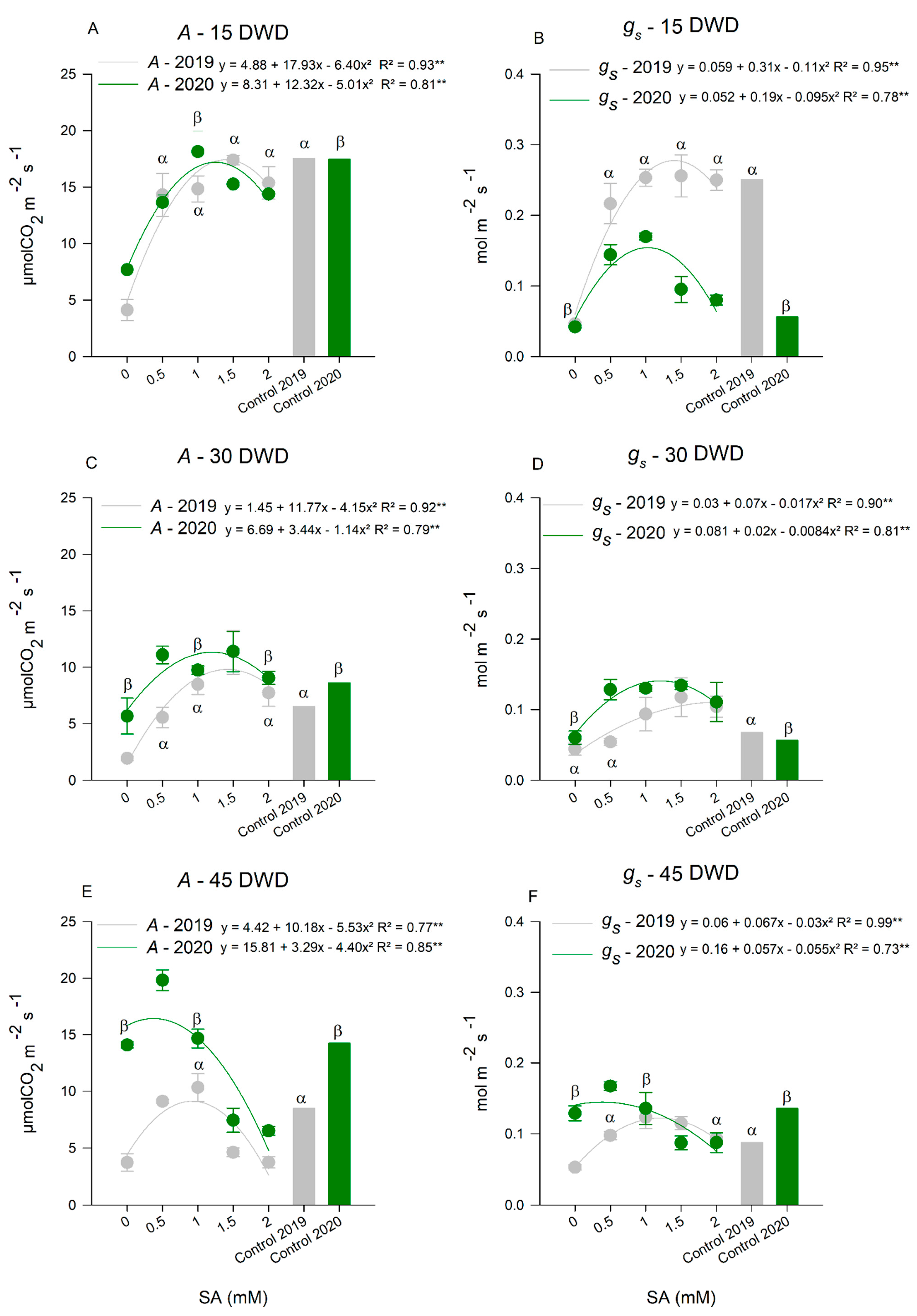
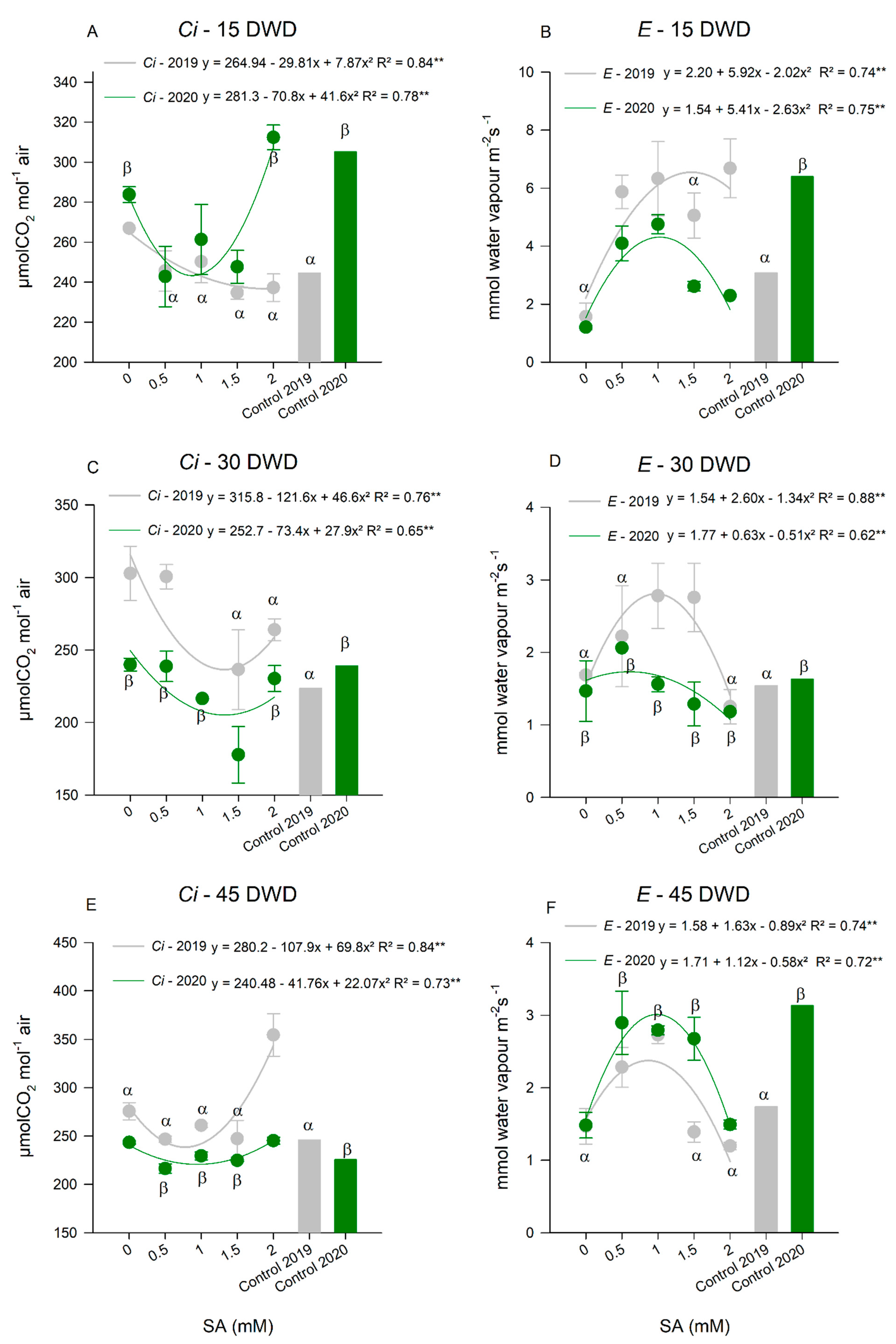
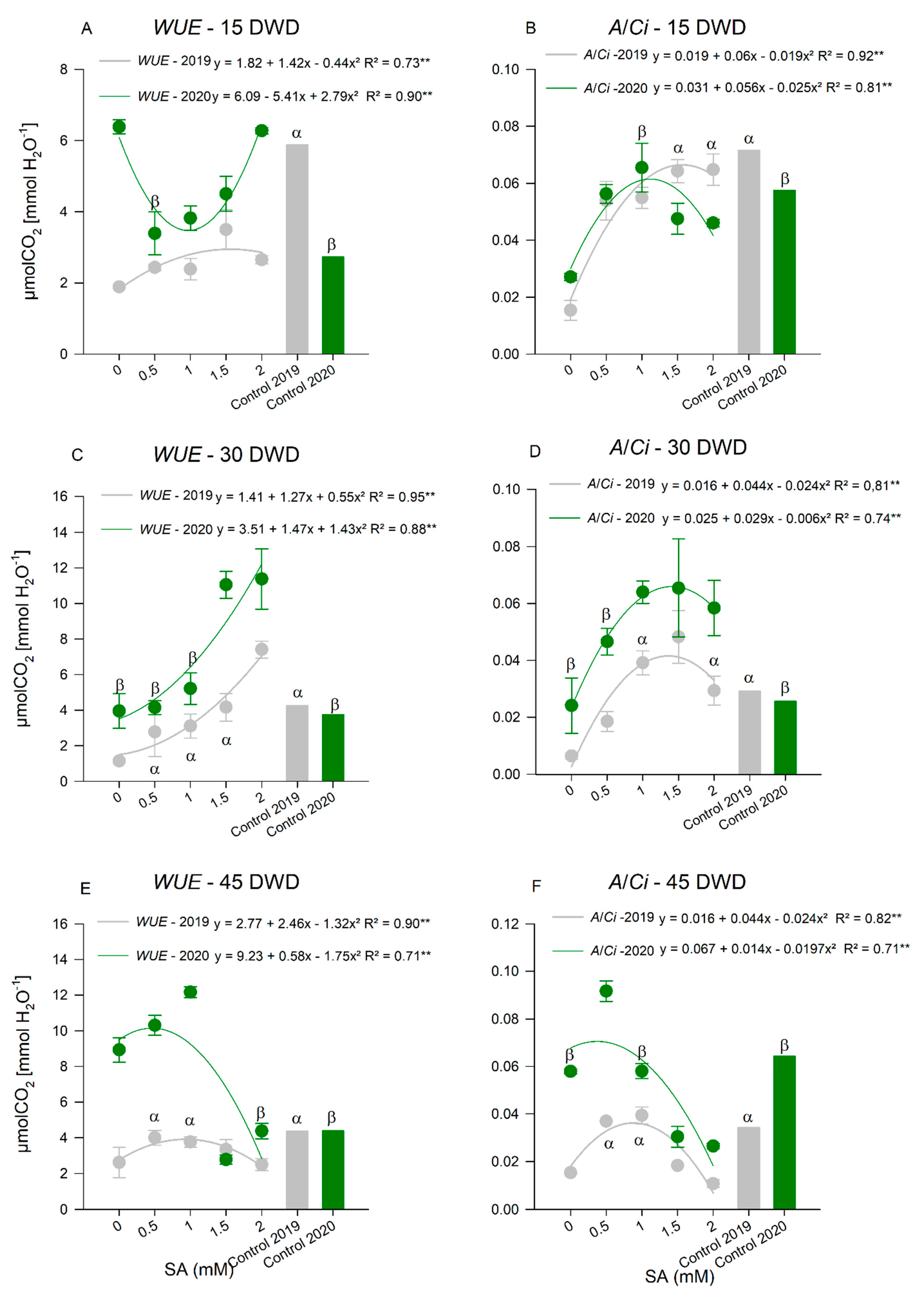
Gas Exchange
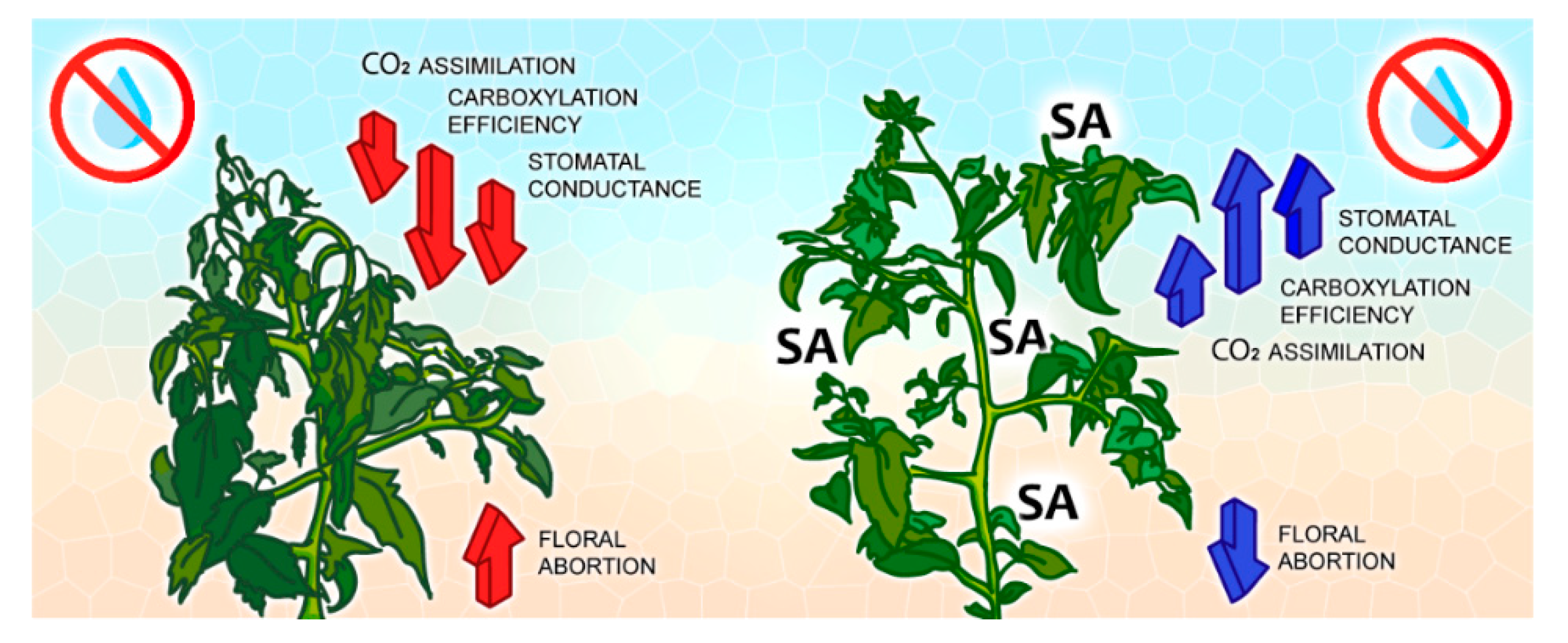
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gomes, A.M.F.; Nhantumbo, N.; Ferreira-Pinto, M.; Massinga, R.; Ramalho, J.C.; Ribeiro-Barros, A. Breeding Elite Cowpea [Vigna unguiculata (L.) Walp] Varieties for Improved Food Security and Income in Africa: Opportunities and Challenges. In Legume Crops—Characterization and Breeding for Improved Food Security; IntechOpen: London, UK, 2019; pp. 626–640. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Fugita, M.; Oku, H.; Islam, M.T. Plant Tolerance to Environmental Stress: Role of Phytoprotectants, 1st ed.; CRC Press: Boca Raton, FL, USA, 2019; p. 488. [Google Scholar]
- Ullah, H.; Luc, P.D.; Gautam, A.; Datta, A. Growth, Yield and Silicon Uptake of Rice (Oryza sativa) As Influenced by Dose and Timing of Silicon Application under Water-Deficit Stress. Arch. Agron. Soil Sci. 2018, 64, 318–330. [Google Scholar] [CrossRef]
- Sirisuntornlak, N.; Ghafoori, S.; Datta, A.; Arirob, W. Seed Priming and Soil Incorporation with Silicon Influence Growth and Yield of Maize under Water-Deficit Stress. Arch. Agron. Soil Sci. 2019, 65, 197–207. [Google Scholar] [CrossRef]
- Chakma, R.; Biswas, A.; Saekong, P.; Ullah, H.; Datta, A. Foliar Application and Seed Priming of Salicylic Acid Affect Growth, Fruit Yield, and Quality of Grape Tomato under Drought Stress. Sci. Hortic. 2021, 280, 109904. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Tomatoes Production—Crops. 2019. Available online: http://faostat3.fao.org/search/tomatoes (accessed on 3 January 2022).
- Farooq, M.; Wahid, A.; Kobayashi, N.S.M.A.; Fujita, D.B.S.M.A.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Lu, C.; Zhang, J. Effects of Water Stress on Photosynthesis, Chlorophyll Fluorescence and Photoinhibition in Wheat Plants. Aust. J. Plant Physiol. 1998, 25, 883–892. [Google Scholar] [CrossRef]
- Hura, T.; Hura, K.; Grzesiak, M.; Rzepka, A. Effect of Long-Term Drought Stress on Leaf Gas Exchange and Fluorescence Parameters in C3 and C4 Plants. Acta Physiol. Plant. 2007, 29, 103–113. [Google Scholar] [CrossRef]
- Jones, H.G.; Sutherlan, R.A. Stomatal Control of Xylem Embolism. Plant Cell Environ. 1991, 11, 11–121. [Google Scholar] [CrossRef]
- Franks, P.J. Stomatal Control and Hydraulic Conductance, with Special Reference to Tall Trees. Tree Physiol. 2004, 24, 865–878. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How Plants Cope with Water Stress in the Field? Photosynthesis and Growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Chen, C.; Khatri, K.; Rathore, M.S.; Pandey, S.P. Gracilaria Dura Extract Confers Drought Tolerance in Wheat by Modulating Abscisic Acid Homeostasis. Plant Physiol. Biochem. 2019, 136, 143–154. [Google Scholar] [CrossRef]
- Hussein, Y.; Amin, G.; Azab, A.; Gahin, H. Induction of Drought Stress Resistance in Sesame (Sesamum indicum L.) Plant by Salicylic Acid and Kinetin. J. Plant Sci. 2015, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Norrie, J.; Ali, H.M.; Salem, M.Z.M.; Mahmoud, E.A.; Yessoufou, K. Enhancement of Calibrachoa Growth, Secondary Metabolites and Bioactivity Using Seaweed Extracts. BMC Complement. Altern. Med. 2016, 16, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Mageed, T.A.; Semida, W.M.; Rady, M.M. Moringa Leaf Extract as Biostimulant Improves Water Use Efficiency, Physio-Biochemical Attributes of Squash Plants under Deficit Irrigation. Agric. Water Manag. 2017, 193, 46–54. [Google Scholar] [CrossRef]
- Kazemi, M. Effect of Foliar Application with Salicylic Acid and Methyl Jasmonate on Growth, Flowering, Yield and Fruit Quality of Tomato. Bull. Environ. Pharmacol. Life Sci. 2014, 3, 154–158. [Google Scholar]
- Gorni, P.H.; Brozulato, M.O.; Lourenção, R.S.; Konrad, E.C.G. Increased Biomass and Salicylic Acid Elicitor Activity in Fennel (Foeniculum vulgare Miller). Braz. J. Food Technol. 2017, 20, e2016172. [Google Scholar] [CrossRef]
- Shemi, R.; Wang, R.; Gheith, E.S.M.; Hussain, H.A.; Hussain, S.; Irfan, M.; Wang, L. Effects of Salicylic Acid, Zinc and Glycine Betaine on Morpho-Physiological Growth and Yield of Maize under Drought Stress. Sci. Rep. 2021, 11, 3195. [Google Scholar] [CrossRef] [PubMed]
- Habibi, G. Exogenous Salicylic Acid Alleviates Oxidative Damage of Barley Plants under Drought Stress. Acta Biol. Szeged. 2012, 56, 57–63. [Google Scholar]
- Saheri, F.; Barzin, G.; Pishkar, L.; Boojar, M.M.A.; Babaeekhou, L. Foliar Spray of Salicylic Acid Induces Physiological and Biochemical Changes in Purslane (Portulaca oleracea L.) under Drought Stress. Biologia 2020, 75, 2189–2200. [Google Scholar] [CrossRef]
- Chavoushi, M.; Najafi, F.; Salimi, A.; Angaji, S.A. Effect of Salicylic Acid and Sodium Nitroprusside on Growth Parameters, Photosynthetic Pigments and Secondary Metabolites of Safflower under Drought Stress. Sci. Hortic. 2020, 259, 108823. [Google Scholar] [CrossRef]
- Trivedi, K.; Vijay Anand, K.G.; Kubava, D.; Patidar, R.; Ghosh, A. Drought Alleviatory Potential of Kappaphycus Seaweed Extract and the Role of the Quaternary Ammonium Compounds as Its Constituents Towards Imparting Drought Tolerance in Zea mays L. J. Appl. Phycol. 2018, 30, 2001–2015. [Google Scholar] [CrossRef]
- Trani, P.E.; Kariya, E.A.; Hanai, S.M.; Anbo, R.H.; Bassetto Júnior, O.B.; Purquerio, L.F.V.; Trani, A.L. Calagem e Adubação do Tomate de Mesa; Boletim Técnico IAC, 215; Instituto Agronômico: Campinas, Brazil, 2015. [Google Scholar]
- Companhia de Entrepostos e Armazéns Gerais de São Paulo (CEAGESP). Programa Brasileiro para Modernização da Horticultura. In Normas Técnicas de Classificação do Tomate; Centro de Qualidade em Horticultura: Sao Paulo, Brazil, 2003. [Google Scholar]
- SIGMAPLOT, SYSTAT. 12.5; SYSTAT: San Jose, CA, USA, 2013.
- Szymańska, R.; Ślesak, I.; Orzechowska, A.; Kruk, J. Physiological and Biochemical Responses to High Light and Temperature Stress in Plants. Environ. Exp. Bot. 2017, 139, 165–177. [Google Scholar] [CrossRef]
- Heerden, P.D.R.; Swanepoel, J.W.; Kruger, G.H.J. Modulation of Photosynthesis by Drought in Two Desert Scrub Species Exhibiting C3-mode CO2 Assimilation. Environ. Exp. Bot. 2009, 61, 124–136. [Google Scholar] [CrossRef]
- Murtaza, G.; Rasool, F.; Habib, R.; Javed, T.; Sardar, K.; Ayub, M.M.; Rasool, A. A Review of Morphological, Physiological and Biochemical Responses of Plants under Drought Stress Conditions. Imp. J. Interdiscip. Res. 2016, 2, 1600–1606. [Google Scholar]
- Li, D.; Wang, M.; Zhang, T.; Chen, X.; Li, C.; Liu, Y.; Brestic, M.; Chen, T.H.H.; Yang, X. Glycinebetaine Mitigated the Photoinhibition of Photosystem II at High Temperature in Transgenic Tomato Plants. Photosynth. Res. 2021, 147, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, T.; Wang, M.; Liu, Y.; Brestic, M.; Chen, T.H.H.; Yang, X. Genetic Engineering of the Biosynthesis of Glycine Betaine Modulates Phosphate Homeostasis by Regulating Phosphate Acquisition in Tomato. Front. Plant Sci. 2019, 9, 1995. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Brestic, M. Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef] [Green Version]
- Sohag, A.A.M.; Tahjib-Ul-Arif, M.; Brestic, M.; Afrin, S.; Sakil, M.A.; Hossain, T.; Hossain, M.A.; Hossain, M.A. Exogenous Salicylic Acid and Hydrogen Peroxide Attenuate Drought Stress in Rice. Plant Soil Environ. 2020, 66, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Aires, E.S.; Ferraz, A.K.L.; Carvalho, B.L.; Teixeira, F.P.; Rodrigues, J.D.; Ono, E.O. Foliar Application of Salicylic Acid Intensifies Antioxidant System and Photosynthetic Efficiency in Tomato Plants. Bragantia 2022, 81, e1522. [Google Scholar] [CrossRef]
- Sehar, Z.; Jahan, B.; Masood, A.; Anjum, N.A.; Khan, N.A. Hydrogen Peroxide Potentiates Defense System in Presence of Sulfur to Protect Chloroplast Damage and Photosynthesis of Wheat under Drought Stress. Physiol. Plant. 2020, 2, 172. [Google Scholar] [CrossRef]
- Negi, J.; Matsuda, O.; Nagasawa, T.; Oba, Y.; Takahashi, H.; Kawai-Yamada, M.; Uchimiya, H.; Hashimoto, M.; Iba, K. CO2 Regulator SLAC1 and its Homologues Are Essential for Anion Homeostasis in Plant Cells. Nature 2008, 452, 483–486. [Google Scholar] [CrossRef]
- Iqbal, N.; Fatma, M.; Gautam, H.; Sehar, Z.; Rasheed, F.; Khan, M.I.R.; Khan, N.A. Salicylic Acid Increases Photosynthesis of Drought Grown Mustard Plants Effectively with Sufficient-N via Regulation of Ethylene, Abscisic Acid, and Nitrogen-Use Efficiency. J. Plant Growth Regul. 2022, 43, 1–12. [Google Scholar] [CrossRef]
- Samui, I.; Skalicky, M.; Sarkar, S.; Brahmachari, K.; Sau, S.; Ray, K.; Hossain, A.; Ghosh, A.; Nanda, M.K.; Bell, R.W.; et al. Productivity, Nutritional Quality and Water Use Efficiency of Tomato (Solanum lycopersicum L.) are Influenced by Drip Irrigation and Straw Mulch in the Coastal Saline Ecosystem of Ganges Delta, India. Sustainability 2020, 12, 6779. [Google Scholar] [CrossRef]
- Pakmehr, A.; Rastgo, M.; Shekari, F.; Saba, J.; Zangani, E. Effects of Seed Priming with Salicylic Acid on Some Morphophysiological Traits and Yield of Cowpea under Drought Stress Condition. Iran. J. Field Crops Res. 2012, 9, 606–614. [Google Scholar] [CrossRef]
- Richards, R.A.; Condon, A.G.; Rebetzke, G.J. Traits to Improve Yield in Dry Environment. In Application of Physiology in Wheat Breeding; Reynolds, M.P., Ortiz-Monasterit, J.I., McNab, A., Eds.; CIMMYT: Mexico City, Mexico, 2001; pp. 88–100. [Google Scholar]
- Grigorova, B.; Vassileva, V.; Klimchuk, D.; Vaseva, I.; Demirevska, K.; Feller, U. Drought, High Temperature, and Their Combination Affect Ultrastructure of Chloroplasts and Mitochondria in Wheat (Triticum aestivum L.) Leaves. J. Plant Interact. 2012, 7, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Schneider, E.A.; Whitman, F. Metabolism of Auxin in Higher Plants. Annu. Rev. Plant Physiol. 1974, 25, 487–513. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Hayat, S.; Ahmad, A. Salicylic Acid Influences Net Photosynthetic Rate, Carboxylation Efficiency, Nitrate Reductase Activity, and Seed Yield in Brassica juncea. Photosynthetica 2003, 41, 281–284. [Google Scholar] [CrossRef]
- Majeau, N.; Coleman, J.R. Correlation of Carbonic Anhydrase and Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase Ex-pression in Pea. Plant Physiol. 1994, 104, 1393–1399. [Google Scholar] [CrossRef] [Green Version]
- Tahjib-Ul-Arif, M.; Siddiqui, M.N.; Sohag, A.A.M.; Sakil, M.A.; Rahman, M.M.; Polash, M.A.S.; Tran, L.S.P. Salicylic Acid-Mediated Enhancement of Photosynthesis Attributes and Antioxidant Capacity Contributes to Yield Improvement of Maize Plants under Salt Stress. J. Plant Growth Regul. 2018, 37, 1318–1330. [Google Scholar] [CrossRef]
- Martínez, C.; Pons, E.; Prats, G.; Leon, J. Salicylic Acid Regulates Flowering Time and Links Defence Responses and Reproductive Development. Plant J. 2003, 37, 209–217. [Google Scholar] [CrossRef]
- Pacheco, A.C.; Cabral, C.S.; Fermino, E.S.S.; Aleman, C.C. Salicylic Acid-Induced Changes to Growth, Flowering and Flavonoids Production in Marigold Plants. J. Med. Plant Res. 2013, 7, 3158–3163. [Google Scholar] [CrossRef]
- Fahad, S.; Nie, L.; Chen, Y.; Wu, C.; Xiong, D.; Saud, S.; Hongyan, L.; Cui, K.; Huang, J. Crop Plant Hormones and Environmental Stress. Sustain. Agric. Rev. 2015, 15, 371–400. [Google Scholar] [CrossRef]
- Sariñana-Aldaco, O.; Sánchez-Chávez, E.; Troyo-Diéguez, E.; Tapia-Vargas, L.M.; Díaz-Pérez, J.C.; Preciado-Rangel, P. Foliar Aspersion of Salicylic Acid Improves Nutraceutical Quality and Fruit Yield in Tomato. Agriculture 2020, 10, 482. [Google Scholar] [CrossRef]
- Preciado-Rangel, P.; Reyes-Pérez, J.J.; Ramírez-Rodríguez, S.C.; Salas-Pérez, L.; Fortis-Hernández, M.; Murillo-Amador, B.; Troyo-Diéguez, E. Foliar Aspersion of Salicylic Acid Improves Phenolic and Flavonoid Compounds, and Also the Fruit Yield in Cucumber (Cucumis sativus L.). Plants 2019, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Horváth, E.; Brunner, S.; Bela, K.; Papdi, C.; Szabados, L.; Tari, I.; Csisz´ar, J. Exogenous Salicylic Acid-Triggered Changes in the Glutathione Transferases and Peroxidases Are Key Factors in the Successful Salt Stress Acclimation of Arabidopsis thaliana. Funct. Plant Biol. 2015, 42, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Poór, P.; Borbély, P.G.; Bódi, N.; Bagyánszki, M.; Tari, I. Effects of Salicylic Acid on Photosynthetic Activity and Chloroplast Morphology under Light and Prolonged Darkness. Photosynthetica 2019, 57, 367–376. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aires, E.S.; Ferraz, A.K.L.; Carvalho, B.L.; Teixeira, F.P.; Putti, F.F.; de Souza, E.P.; Rodrigues, J.D.; Ono, E.O. Foliar Application of Salicylic Acid to Mitigate Water Stress in Tomato. Plants 2022, 11, 1775. https://doi.org/10.3390/plants11131775
Aires ES, Ferraz AKL, Carvalho BL, Teixeira FP, Putti FF, de Souza EP, Rodrigues JD, Ono EO. Foliar Application of Salicylic Acid to Mitigate Water Stress in Tomato. Plants. 2022; 11(13):1775. https://doi.org/10.3390/plants11131775
Chicago/Turabian StyleAires, Eduardo Santana, Andrew Kim Lopes Ferraz, Beatriz Lívero Carvalho, Fabricio Palla Teixeira, Fernando Ferrari Putti, Emanuele Possas de Souza, João Domingos Rodrigues, and Elizabeth Orika Ono. 2022. "Foliar Application of Salicylic Acid to Mitigate Water Stress in Tomato" Plants 11, no. 13: 1775. https://doi.org/10.3390/plants11131775