
Genetic Analysis in Grain Legumes [Vigna radiata (L.) Wilczek] for Yield Improvement and Identifying Heterotic Hybrids

 ,
,  , ,
, ,  , ,
, ,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussions
2.1. Analysis of Variance (ANOVA) for Combining Ability
2.2. Mean Evaluation of Parents and Their Crosses
2.3. General Combining Ability Effects
2.4. Specific Combining Ability Effect
2.5. Heterosis
3. Materials and Methods
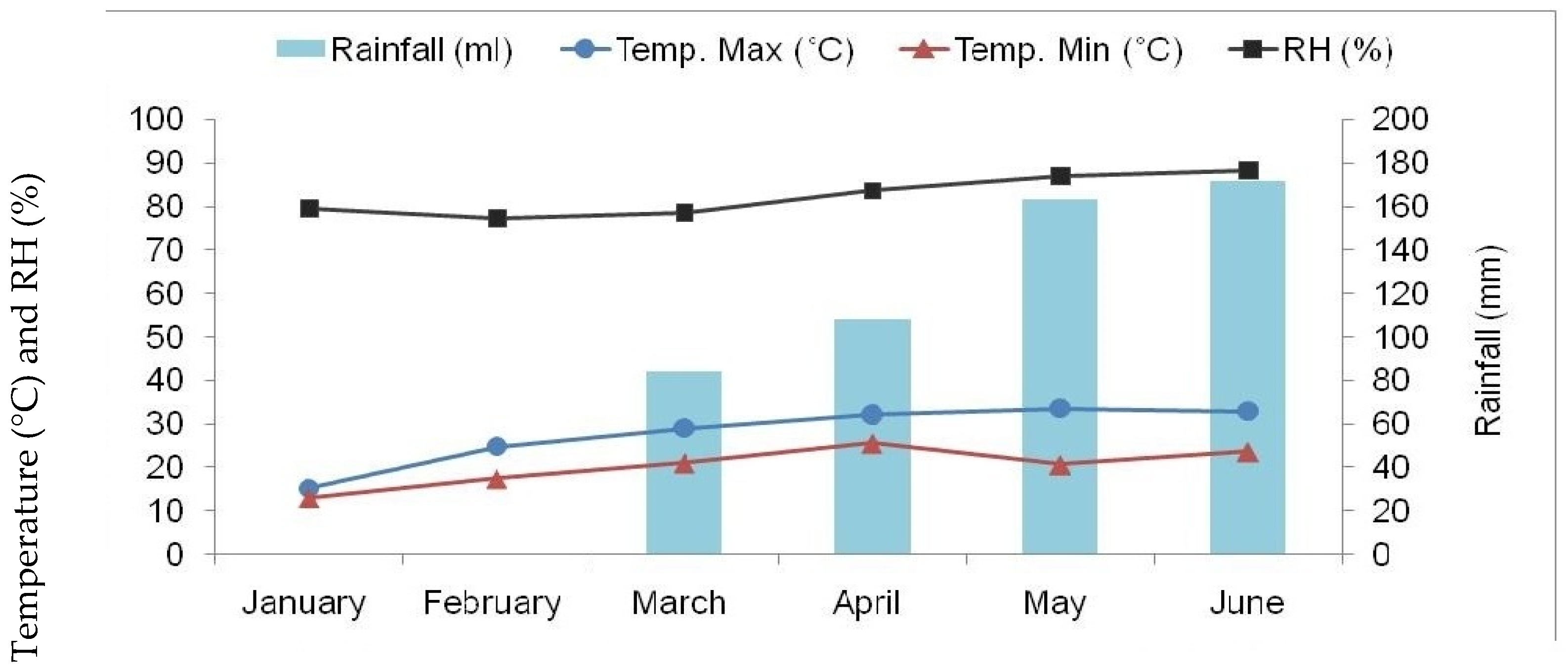
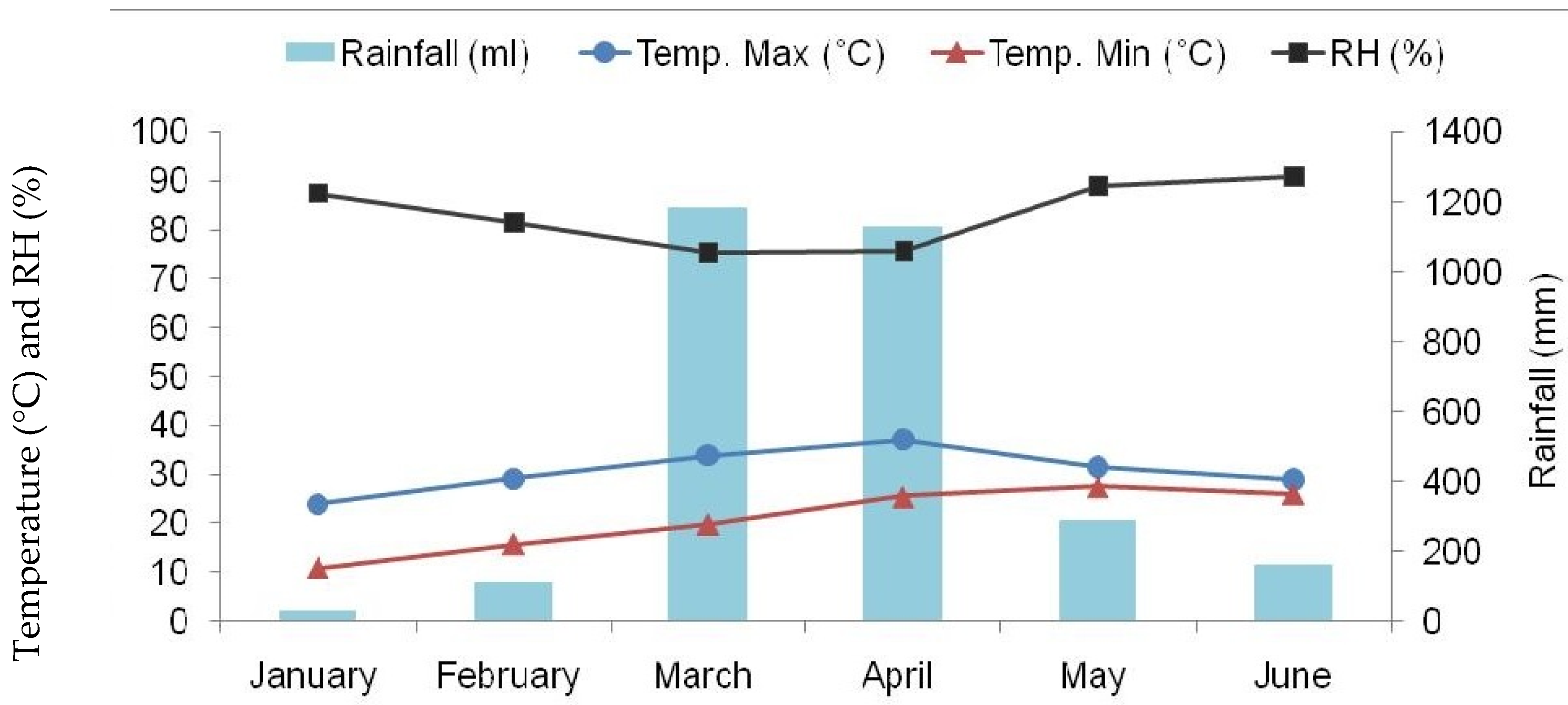
3.1. Experimental Site and Climate
3.2. Experimental Materials and Crossing Technique
3.3. Crop Management
3.4. Data Collection
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References and Notes
- Kumar, K.; Prasad, Y.; Mishra, S.B.; Pandey, S.S.; Kumar, R. Study on Genetic Variability, Correlation and Path Analysis with Grain Yield and Yield Attributing Traits in Green Gram [Vigna radiata (L.) Wilczek]. Bioscan 2013, 8, 1551–1555. [Google Scholar]
- Mubarak, A.E. Nutritional Composition and Antinutritional Factors of Mung Bean Seeds (Phaseolus aureus) as Affected by Some Home Traditional Processes. Food Chem. 2005, 89, 489–495. [Google Scholar] [CrossRef]
- Tachibana, N.; Wanezaki, S.; Nagata, M.; Motoyama, T.; Kohno, M.; Kitagawa, S. Intake of Mung Bean Protein Isolate Reduces Plasma Triglyceride Level in Rats. Funct. Foods Health Dis. 2013, 3, 365–376. [Google Scholar] [CrossRef]
- Botinestean, C.; Hadaruga, N.G.; Hadaruga, D.I.; Jianu, I. Fatty Acids Composition by Gas Chromatography–Mass Spectrometry (GC-MS) and Most Important Physical-Chemicals Parameters of Tomato Seed Oil. J. Agroaliment. Process Technol. 2012, 18, 89–94. [Google Scholar]
- Nair, R.; Schreinemachers, P. Global Status and Economic Importance of Mungbean. In The Mungbean Genome; Springer: Cham, Switzerland, 2020; pp. 1–8. [Google Scholar] [CrossRef]
- AIS. Agric Inf. Serv. (AIS), Dep. Agric. Extension Minist. Agric. Gov. People’s Repub: Dhaka, Bangladesh, 2020.
- Kumar, A.; Kant, R.; Singh, S.K.; Mishra, S.B.; Kumar, H. Line × Tester Analysis for Yield and Yield Component Traits in Green gram [Vigna radiata (L.) Wilczek]. Int. J. Agric. Stat. Sci. 2017, 13, 259–263. [Google Scholar]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Genotypic Variability for Nutrient, Antioxidant, Yield and Yield Contributing Traits in Vegetable Amaranth. J. Food Agric. Environ. 2014, 12, 168–174. Available online: https://www.wflpublisher.com/Abstract/5378 (accessed on 1 May 2022).
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Variability, heritability and genetic association in vegetable amaranth. Spanish J. Agril. Res. 2015, 13, 0702. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Variability in Composition of Vitamins and Mineral Antioxidants in Vegetable Amaranth. Genetika 2015, 47, 85–96. [Google Scholar] [CrossRef]
- Latha, V.S.; Eswari, K.B.; Sudheer, K.S. Combining Ability Analysis for Seed Yield and Its Component Characters in Green Gram (Vigna radiata (L.) Wilczek). Int. J. Chem. Stud. 2018, 6, 237–242. [Google Scholar]
- Patil, A.B.; Desai, N.C.; Mule, P.N.; Khandelwal, V. Combining Ability for Yield and Component Characters in Mungbean. Legume Res. 2011, 34, 190–195. [Google Scholar]
- Sopan, S.Z.; Sao, A.; Nanda, H.C.; Nair, S.K. Combining Ability Analysis for Seed Yield, Its Contributing Traits and Protein Content in Mungbean (Vigna radiata (L.) Wilczek). Int. J. Curr. Microbiol. App. Sci. 2018, 6, 761–764. [Google Scholar]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Genetic Variation and Interrelationships among Antioxidant, Quality, and Agronomic Traits in Vegetable Amaranth. Turk. J. Agric. For. 2016, 40, 526–535. [Google Scholar] [CrossRef]
- Biswas, P.S.; Sarker, U.; Bhuiyan, M.A.R.; Khatun, S. Genetic Divergence in Cold Tolerant Irrigated Rice (Oryza sativa L.). Agriculturists 2006, 4, 15–20. [Google Scholar]
- Hasan, M.J.; Kulsum, M.U.; Majumder, R.R.; Sarker, U. Genotypic Variability for Grain Quality Attributes in Restorer Lines of Hybrid Rice. Genetika 2020, 52, 973–989. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutritional and Bioactive Constituents and Scavenging Capacity of Radicals in Amaranthus hypochondriacus. Sci. Rep. 2020, 10, 19962. [Google Scholar] [CrossRef]
- Hasan-Ud-Daula, M.; Sarker, U. Variability, Heritability, Character Association, and Path Coefficient Analysis in Advanced Breeding Lines of Rice (Oryza sativa L.). Genetika 2020, 52, 711–726. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Color Attributes, Betacyanin, and Carotenoid Profiles, Bioactive Components, and Radical Quenching Capacity in Selected Amaranthus gangeticus Leafy Vegetables. Sci. Rep. 2021, 11, 11559. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutraceuticals, Antioxidant Pigments, and Phytochemicals in the Leaves of Amaranthus spinosus and Amaranthus viridis Weedy Species. Sci. Rep. 2019, 9, 20413. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarty, T.; Sarker, U.; Hasan, M.; Rahman, M.M. Variability in Mineral Compositions, Yield and Yield Contributing Traits of Stem Amaranth (Amaranthus lividus). Genetika 2018, 50, 995–1010. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Nutraceuticals, Phytochemicals, and Radical Quenching Ability of Selected Drought-Tolerant Advance Lines of Vegetable Amaranth. BMC Plant Biol. 2020, 20, 564. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Polyphenol and Flavonoid Profiles and Radical Scavenging Activity in Selected Leafy Vegetable Amaranthus gangeticus. BMC Plant Biol. 2020, 20, 499. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Lin, Y.P.; Oba, S.; Yoshioka, Y.; Ken, H. Prospects and potentials of underutilized leafy Amaranths as vegetable use for health-promotion. Plant Physiol. Biochem. 2022, 182, 104–123. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Iqbal, M.A.; Hossain, M.N.; Oba, S.; Ercisli, S.; Muresan, C.C.; Marc, R.A. Colorant Pigments, Nutrients, Bioactive Components, and Antiradical Potential of Danta Leaves (Amaranthus lividus). Antioxidants 2022, 11, 1206. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S.; Alsanie, W.F.; Gaber, A. Characterization of Phytochemicals, Nutrients, and Antiradical Potential in Slim Amaranth. Antioxidants 2022, 11, 1089. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Azam, M.G.; Talukder, M.Z.A. Genetic Variation in Mineral Profiles, Yield Contributing Agronomic Traits, and Foliage Yield of Stem Amaranth. Genetika 2022, 54, 91–108. [Google Scholar] [CrossRef]
- Sarker, U.; Rabbani, M.G.; Oba, S.; Eldehna, W.M.; Al-Rashood, S.T.; Mostafa, N.M.; Eldahshan, O.A. Phytonutrients, Colorant Pigments, Phytochemicals, and Antioxidant Potential of Orphan Leafy Amaranthus Species. Molecules 2022, 27, 2899. [Google Scholar] [CrossRef]
- Srivastava, R.L.; Singh, G. Heterosis for Yield and its Contributing Characters in Mungbean (Vigna radiata (L.) wilczek). Indian J. Sci. Res. 2013, 4, 131–134. [Google Scholar]
- Alam, A.S.M.S.; Sarker, U.; Mian, M.A.K. Line × Tester Analysis in Hybrid Rice (Oryza sativa L.). Ann. Bangladesh Agric. 2007, 11, 37–44. [Google Scholar]
- Azad, A.K.; Sarker, U.; Ercisli, S.; Assouguem, A.; Ullah, R.; Almeer, R.; Sayed, A.A.; Peluso, I. Evaluation of Combining Ability and Heterosis of Popular Restorer and Male Sterile Lines for the Development of Superior Rice Hybrids. Agronomy 2022, 12, 965. [Google Scholar] [CrossRef]
- Islam, M.M.; Sarker, U.; Rasul, M.G.; Rahman, M.M. Heterosis in local boro rice (Oryza sativa L.). Bangladesh J. Pl. Breed. Genet. 2010, 23, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Reddy, B.R.K.; Reddy, K.H.P.; Reddy, D.M.; Sudhakar, P.; Reddy, B.R. Heterosis for Yield, Yield Attributes and Water use Efficiency related Traits in Mungbean. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 634–643. [Google Scholar] [CrossRef]
- Viraj, R.; Pithia, M.S.; Solanki, H.V. Combining Ability Analysis for Yield and Yield Components in Mungbean [Vigna radiata (L.) Wilczek]. Legum. Res. Int. J. 2020, 43, 151–154. [Google Scholar] [CrossRef]
- Griffing, B. A Generalised Treatment of the Use of Diallel Crosses in Quantitative Inheritance. Heredity 1956, 10, 31–50. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Biswas, P.S.; Prasad, B.; Mian, M.A.K. Correlated Response, Relative Selection Efficiency and Path Analysis in Cold Tolerant Rice. Bangladesh J. Pl. Breed. Genet. 2001, 14, 33–36. [Google Scholar]
- Sarker, U.; Mian, M.A.K. Genetic Variability, Character Association and Path Analysis for Yield and Its Components in Rice. J. Asiat. Soc. Bangladesh Sci. 2003, 29, 47–54. [Google Scholar]
- Sarker, U.; Mian, M.A.K. Genetic Variations and Correlations between Floral Traits in Rice. Bangladesh J. Agril. Res. 2004, 29, 553–558. [Google Scholar]
- Azam, M.G.; Sarker, U.; Banik, B.R. Genetic Variability of Yield and Its Contributing Characters on CIMMYT Maize Inbreds under Drought Stress. Bangladesh J. Agric. Res. 2014, 39, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.N.; Sarker, U.; Raihan, M.S.; Al-Huqail, A.A.; Siddiqui, M.H.; Oba, S. Influence of Salinity Stress on Color Parameters, Leaf Pigmentation, Polyphenol and Flavonoid Contents, and Antioxidant Activity of Amaranthus lividus Leafy Vegetables. Molecules 2022, 27, 1821. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Ercisli, S.; Assouguem, A.; Alotaibi, A.; Ullah, R. Bioactive Phytochemicals and Quenching Activity of Radicals in Selected Drought-Resistant Amaranthus tricolor Vegetable Amaranth. Antioxidants 2022, 11, 578. [Google Scholar] [CrossRef]
- Kumar, B.S.; Prakash, M.; Gokulakrishnan, J. Combining Ability Analysis in Mungbean (Vigna radiata (L.) Wilczek). Crop Improv. 2010, 37, 165–167. [Google Scholar]
- Griffing, B. Concept of General and Specific Combining Ability in Relation to Diallel Crossing Systems. Aust. J. Biol. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]
- Nath, A.; Maloo, S.R.; Nath, S.; Yadav, G.S. Combining Ability Analysis for Seed Protein and Methionine Content in Green Gram [Vigna radiata L. wilczek]. Int. J. Com. Sci. 2018, 52, 34–39. [Google Scholar] [CrossRef]
- Sarker, U.; Mian, M.A.K. Line × Tester Analysis for Yield and Its Components in Rice (Oryza sativa L.). J. Asiatic Soc. Bangladesh Sci. 2002, 28, 71–81. [Google Scholar]
- Sarker, U.; Rasul, M.G.; Mian, M.A.K. Combining Ability of CMS and Restorer Lines in Rice. Bangladesh J. Pl. Breed. Genet. 2003, 16, 1769. [Google Scholar] [CrossRef]
- Sarker, U.; Biswas, P.S.; Prasad, B.; Mian, M.A.K. Heterosis and Genetic Analysis in Rice Hybrids. Pakistan J. Biol. Sci. 2002, 5, 1–5. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, N.K.; Kumar, R.; Chandel, D.; Yadav, M.K. Genetic Divergence Studies in Mungbean Germplasm under Arid Environment. Int. J. Com. Sci. 2022, 11, 1617–1619. [Google Scholar]
- Mondal, M.M.A.; Puteh, A.B.; Malek, M.A.; Hasan, M.F.; Rahman, M.H. Pod Maturity Synchrony in Relation to Canopy Structure in Mungbean (Vigna radiata). Int. J. Agric. Biol. 2013, 15, 963–967. [Google Scholar]
- Gupta, S.K.; Singh, S.; Kaur, A. Heterosis for Seed Yield and its Component Traits in Desi X Desi and Desi X Kabuli Crosses of Chickpea (Cicer arietinum L.). Crop Improv. 2003, 30, 203–207. [Google Scholar]
- Kumar, B.S.; Prakash, M. Heterosis for Biometric and Biochemical Components in Mungbean (Vigna radiata (L.) Wilczek). Karnataka J. Agric. Sci. 2011, 24, 523–524. [Google Scholar]
- Sujatha, K.; Kajjidoni, S.T. Genetic Analysis Involving Selected Powdery Mildew Resistant Lines in Mungbean (Vigna radiata (L.) Wilckzek). Mol. Plant Breed. 2013, 4, 38–43. [Google Scholar] [CrossRef]
- Dhuppe, M.V.; Wadikar, P.B.; Pole, S.P. Heterosis in mung Bean. Res. J. Agric. Sci. 2010, 1, 438–440. [Google Scholar]
- Zubair, M.; Ajmal, S.U.; Ali, S. Heterosis for Yield-Related Attributes in Mungbean (Vigna radiata (L.) Wilczek). Pak. J. Bot. 2010, 42, 3209–3214. [Google Scholar]
- Yadav, P.S.; Lavanya, G.R.; Vishwakarma, M.K.; Saxesena, R.R.; Baranwal, D.K.; Singh, S. Heterosis Studies Using Diallel Analysis for Yield and Component Characters in Mungbean (Vigna radiata L. Wilczek). Bioscan 2015, 10, 711–713. [Google Scholar]
- BARI (Bangladesh Agricultural Research Institute). Krishi Projukyi Hatboi (Handbook on Agro-Technology), 5th ed.; Farm Technology Group: Gazipur, Bangladesh, 2014. [Google Scholar]
- PBTools version1.2. Biometrics and Breeding Informatics, PBGB Division, International Rice Research Institute. 2014.
- Cox, D.J.; Frey, K.J. Combining Ability and the Selection of Parents for Interspecific Oat Matings. Crop Sci. 1984, 24, 963–967. [Google Scholar] [CrossRef]
- Sarker, U.; Rasul, M.G.; Mian, M.A.K. Heterosis and Combining Ability in Rice. Bangladesh J. Pl. Breed. Genet. 2002, 15, 17–26. [Google Scholar]
- Abrham, S.; Mandefro, N.; Sentayehu, A. Heterosis and Study of Hot Pepper (Capsicum annuum L.) Genotypes in Southern Ethiopia. Int. J. Plant Breed. Genet. 2017, 11, 63–70. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Source of Variance | d.f. | Mean Square | ||||||
|---|---|---|---|---|---|---|---|---|
| DF | DM | Plant Height | Pods per Plant | Pod Length | Seeds per Pod | Yield per Plant | ||
| Replications | 2 | 3.45 | 5.54 | 0.25 | 2.49 | 0.03 | 2.78 | 0.03 |
| Genotypes | 20 | 51.78 ** | 44.23 ** | 248.19 ** | 265.22 ** | 2.74 ** | 256.73 ** | 2.66 ** |
| Parents | 5 | 29.12 ** | 66.45 ** | 301.60 ** | 73.81 ** | 5.65 ** | 80.89 ** | 5.29 ** |
| Crosses | 14 | 65.67 ** | 38.43 ** | 102.88 ** | 346.93 ** | 1.89 ** | 248.05 ** | 1.82 ** |
| Error | 40 | 24.54 ** | 24.23 ** | 1.58 | 3.39 | 0.51 | 5.58 | 0.22 |
| GCA | 5 | 14.00 ** | 14.43 ** | 72.28 ** | 109.02 ** | 1.41 * | 40.06 ** | 1.45 * |
| SCA | 15 | 5.67 | 10.22 | 85.22 ** | 81.26 ** | 0.57 | 100.42 ** | 0.52 ** |
| Error | 40 | 0.87 | 0.76 | 0.50 | 1.11 | 0.15 | 1.65 | 0.07 |
| GCA/SCA | 2.47 | 1.41 | 0.85 | 1.34 | 2.47 | 0.40 | 2.79 | |
| Parents/Crosses | Days to Flowering | Days to Maturity | Plant Height | Pods per Plant | Pod Length | Seeds per Pod | Yield per Plant |
|---|---|---|---|---|---|---|---|
| Parents | |||||||
| P1 | 43.5 ± 1.20 | 70.5 ± 1.17 | 40.5 ± 2.20 | 34.2 ± 2.17 | 8.1 ± 0.56 | 11 ± 1.16 | 7.8 ± 0.46 |
| P2 | 43.0 ± 1.72 | 71.5 ± 1.46 | 46.4 ± 0.72 | 21.4 ± 0.46 | 8.5 ± 0.51 | 10 ± 1.53 | 8.9 ± 0.36 |
| P3 | 42.5 ± 2.19 | 69.5 ± 2.08 | 50.6 ± 0.19 | 29.0 ± 1.08 | 8.1 ± 0.85 | 12 ± 2.00 | 7.8 ± 0.48 |
| P4 | 43.5 ± 1.71 | 68.5 ± 1.88 | 61.2 ± 0.71 | 33.4 ± 0.88 | 6.2 ± 0.24 | 12 ± 3.00 | 7.1 ± 0.83 |
| P5 | 42.0 ± 2.51 | 71.5 ± 1.53 | 38.8 ± 1.51 | 15.0 ± 0.53 | 9.9 ± 0.70 | 11 ± 2.00 | 9.9 ± 0.55 |
| P6 | 48.5 ± 2.00 | 78.5 ± 1.70 | 35.2 ± 1.00 | 24.2 ± 0.70 | 6.1 ± 0.85 | 11 ± 1.53 | 6.0 ± 0.44 |
| Crosses | |||||||
| P1 ×P2 | 44.5 ± 2.40 | 79.5 ± 2.10 | 41.66 ± 1.40 | 34.20 ± 1.10 | 8.00 ± 1.12 | 10 ± 2.08 | 8.60 ± 0.25 |
| P1 ×P3 | 44.0 ± 2.38 | 79.0 ± 1.45 | 37.00 ± 1.38 | 26.75 ± 0.45 | 8.06 ± 0.45 | 9 ± 2.00 | 8.25 ± 0.40 |
| P1 × P4 | 44.5 ± 2.92 | 76.5 ± 1.85 | 46.75 ± 0.92 | 42.75 ± 0.85 | 7.38 ± 0.54 | 10 ± 2.03 | 7.80 ± 0.30 |
| P1 × P5 | 48.5 ± 2.47 | 68.5 ± 1.78 | 43.25 ± 1.47 | 36.25 ± 0.78 | 8.88 ± 0.72 | 12 ± 2.00 | 9.90 ± 0.16 |
| P1 × P6 | 49.0 ± 2.31 | 70.5 ± 2.00 | 39.50 ± 0.31 | 36.25 ± 1.00 | 8.60 ± 0.49 | 12 ± 2.52 | 8.70 ± 0.55 |
| P2 × P3 | 44.5 ± 3.05 | 67.5 ± 3.08 | 37.00 ± 2.05 | 39.00 ± 2.08 | 9.30 ± 0.71 | 12 ± 3.00 | 8.90 ± 0.43 |
| P2 × P4 | 43.5 ± 1.76 | 76.5 ± 2.55 | 46.75 ± 0.76 | 31.60 ± 1.55 | 8.30 ± 0.68 | 12 ± 1.53 | 6.90 ± 0.10 |
| P2 × P5 | 43.5 ± 1.62 | 76.5 ± 2.20 | 43.25 ± 0.62 | 18.75 ± 1.20 | 9.25 ± 0.78 | 10 ± 2.08 | 8.60 ± 0.47 |
| P2 × P6 | 41.5 ± 1.79 | 76.0 ± 1.80 | 39.50 ± 0.79 | 26.25 ± 0.80 | 8.60 ± 0.65 | 10 ± 1.00 | 6.75 ± 0.23 |
| P3 × P4 | 43.5 ± 2.27 | 76.5 ± 3.67 | 49.75 ± 2.27 | 31.75 ± 2.67 | 8.30 ± 1.01 | 13 ± 2.52 | 6.75 ± 0.45 |
| P3 × P5 | 42.5 ± 2.11 | 76.5 ± 2.34 | 34.50 ± 1.11 | 25.00 ± 1.34 | 8.30 ± 0.72 | 11 ± 2.52 | 7.25 ± 0.81 |
| P3 × P6 | 49.0 ± 2.22 | 78.5 ± 1.62 | 33.60 ± 1.22 | 35.20 ± 0.62 | 6.80 ± 0.56 | 11 ± 1.00 | 6.30 ± 0.46 |
| P4 × P5 | 47.5 ± 2.15 | 66.5 ± 2.51 | 57.50 ± 1.15 | 25.50 ± 1.51 | 10.40 ± 0.81 | 12 ± 1.53 | 9.20 ± 0.47 |
| P4 × P6 | 43.0 ± 1.77 | 74.5 ± 1.79 | 44.50 ± 0.77 | 25.75 ± 0.79 | 6.70 ± 0.42 | 11 ± 2.08 | 6.50 ± 0.34 |
| P5 × P6 | 44.0 ± 1.27 | 79.5 ± 1.75 | 43.33 ± 0.27 | 12.00 ± 0.75 | 8.30 ± 0.51 | 12 ± 3.21 | 6.83 ± 0.27 |
| Mean | 44.57 | 73.93 | 43.36 | 28.77 | 8.19 | 12.90 | 7.84 |
| LSD at 5% | 3.05 | 2.67 | 2.07 | 3.04 | 1.16 | 1.90 | 0.77 |
| Parents | Characters | ||||||
|---|---|---|---|---|---|---|---|
| DF | DM | Plant Height | Pods per Plant | Pod Length | Seeds per Pod | Yield per Plant | |
| P1 | −0.37 | −0.03 | 2.86 ** | 6.11 ** | −0.23 | −0.24 | 0.34 * |
| P2 | −1.28 ** | 1.54 ** | −2.92 ** | −1.49 ** | 0.35 * | 0.51 | 0.36 * |
| P3 | −0.55 | −0.86 | −2.25 ** | 1.68 ** | 0.09 | 0.67 | −0.05 |
| P4 | 2.54 ** | 2.24 ** | 3.97 ** | 0.75 | −0.50 * | 3.68 ** | −0.50 ** |
| P5 | −0.27 | 1.22 * | −2.61 ** | −3.21 ** | 0.59 ** | −2.21 ** | 0.46 ** |
| P6 | −0.26 | −0.81 | 0.93 ** | −3.86 ** | −0.29 | −2.43 ** | −0.51 ** |
| SE Gi | 0.27 | 0.51 | 0.23 | 0.34 | 0.13 | 0.41 | 0.08 |
| SE Gi-Gj | 0.42 | 0.69 | 0.35 | 0.53 | 0.20 | 0.64 | 0.13 |
| Crosses | Characters | ||||||
|---|---|---|---|---|---|---|---|
| DF | DM | Plant Height | Pods per Plant | Pod Length | Seeds Per Pod | Yield per Plant | |
| P1 × P2 | −0.75 | 0.85 | 2.74 ** | 0.45 | 0.04 | −16.45 ** | −0.01 |
| P1 × P3 | 2.83 ** | 2.01 | −2.57 ** | −10.64 ** | 0.54 | −2.95 ** | 0.77 * |
| P1 × P4 | −0.09 | −0.83 | 0.95 | 6.37 ** | 0.72 | 0.05 | 0.42 |
| P1 × P5 | 0.26 | −0.39 | 4.03 ** | 24.31 ** | −1.42 | 5.93 ** | 0.82 * |
| P1 × P6 | 2.22 ** | −3.08 * | −3.27 ** | −6.10 ** | −1.11 ** | 2.83 | 0.84 * |
| P2 × P3 | 1.38 | 3.35 * | 7.58 ** | 9.35 ** | −1.14 * | 6.30 ** | 0.68 * |
| P2 × P4 | 2.25 ** | 6.68 *** | 0.38 | 2.65 * | 0.53 * | 4.63 ** | −0.93 ** |
| P2 × P5 | 0.08 | 5.23 ** | 9.74 * | −6.53 ** | 0.28 | 3.18 * | −0.05 |
| P2 × P6 | 2.68 ** | 2.04 | 1.59 | 2.28 | 0.41 | 2.74 | −0.74 * |
| P3 × P4 | 3.02 ** | −0.08 | 8.08 ** | −0.20 | 0.83 | −11.20 ** | −0.37 |
| P3 × P5 | −0.64 | −0.56 | 0.23 | −5.64 ** | −0.26 | −20.66 ** | −0.85 * |
| P3 × P6 | 0.44 | −0.86 | −3.23 ** | 8.02 ** | 0.01 | 6.24 ** | −0.71 * |
| P4 × P5 | 4.36 *** | 2.48 | 17.16 ** | −3.24 * | −0.26 | −2.33 | 0.78 ** |
| P4 × P6 | 4.36 *** | 5.29 *** | 0.66 | −0.76 | −0.37 | −4.76 ** | 0.19 |
| P5 × P6 | −0.34 | −0.71 | 5.85 ** | −10.06 ** | 0.56 | −11.22 ** | −0.45 |
| SE Sij | 0.74 | 1.29 | 0.63 | 0.93 | 0.35 | 1.14 | 0.23 |
| SE Sij-Sik | 1.11 | 1.93 | 0.94 | 1.39 | 0.52 | 1.70 | 0.35 |
| SE Sij-Skl | 1.03 | 1.79 | 0.87 | 1.29 | 0.48 | 1.57 | 0.32 |
| Crosses | DF | DM | Plant Height | Pods per Plant | ||||
| MP (%) | BP (%) | MP (%) | BP (%) | MP (%) | BP (%) | MP (%) | BP (%) | |
| P1 × P2 | 2.89 ** | 2.30 * | 11.97 ** | 11.19 ** | −4.12 ** | −10.91 ** | 23.02 ** | −2.92 ** |
| P1 × P3 | 2.33 * | 1.15 NS | 12.86 ** | 12.06 ** | −18.77 ** | −29.86 ** | −15.35 ** | −21.78 ** |
| P1 × P4 | 2.30 * | 2.30 * | 10.07 ** | 8.51 ** | −8.06 ** | −28.42 ** | 26.48 ** | 25.00 ** |
| P1 × P5 | 13.45 ** | 11.49 ** | −3.52 ** | −4.20 ** | 9.08 ** | 6.94 ** | 47.36 ** | 5.99 ** |
| P1 × P6 | 6.52 ** | 1.03 NS | −5.37 ** | −10.19 ** | 4.36 ** | −2.64 * | 24.14 ** | 5.99 ** |
| P2 × P3 | 4.09 ** | 3.49 ** | −4.26 ** | −5.59 ** | −23.71 ** | −28.04 ** | 54.76 ** | 34.48 ** |
| P2 × P4 | 0.58NS | 2.30 * | 9.29 ** | 6.99 ** | −16.37 ** | −25.85 ** | 15.33 ** | −5.39 ** |
| P2 × P5 | 2.35 * | 1.16 NS | 6.99 ** | 6.99 ** | −13.50 ** | −35.90 ** | 3.02 ** | −12.38 ** |
| P2 × P6 | −9.29 ** | −14.43 ** | 1.33 NS | −3.18 ** | 6.76 ** | 1.89 NS | 15.13 ** | 8.47 ** |
| P3 × P4 | 1.16 NS | −2.30 * | 10.87 ** | 10.07 ** | −11.00 ** | −20.48 ** | 1.76 NS | −4.94 ** |
| P3 × P5 | 0.59 NS | −2.35 * | 8.51 ** | 6.99 ** | −31.00 ** | −8.60 ** | 13.64 ** | −13.79 ** |
| P3 × P6 | 7.69 ** | 1.03 NS | 6.08 ** | −1.27 NS | −9.19 ** | −74.59 ** | 32.33 ** | 21.38 ** |
| P4 × P5 | 11.11 ** | 9.20 ** | −5.00 ** | −6.99 ** | 15.00 ** | 37.40 ** | 5.37 ** | −23.65 ** |
| P4 × P6 | −6.52 ** | −11.34 ** | 1.36 NS | −5.10 ** | 20.27 ** | 15.41 ** | −10.59 ** | −22.90 ** |
| P5 × P6 | −2.76 ** | −9.28 ** | 6.00 ** | 1.27 NS | 17.11 ** | 117.11 ** | −38.78 ** | −50.41 ** |
| Crosses | Pod Length | Seeds per Pod | Yield per Plant | |||||
| MP (%) | BP (%) | MP (%) | BP (%) | MP (%) | BP (%) | |||
| P1 × P2 | 2.56 * | −5.88 * | 2.00 NS | −1.92 NS | 2.99 ** | −3.37 ** | ||
| P1 × P3 | 6.05 ** | −0.49 NS | −19.64 ** | −25.00 ** | 5.77 ** | 5.77 ** | ||
| P1 × P4 | 10.98 ** | 3.94 ** | −14.04 ** | −20.97 ** | 4.70 ** | 12.82 ** | ||
| P1 × P5 | 11.00 ** | −0.22 NS | 14.81 ** | 10.71 ** | 11.86 ** | 20.20 ** | ||
| P1 × P6 | 13.16 ** | 6.17 ** | 11.54 ** | 11.54 ** | 26.09 ** | 11.54 ** | ||
| P2 × P3 | 12.05 ** | 9.41 ** | 9.26 ** | −1.67 NS | 6.59 ** | −3.37 ** | ||
| P2 × P4 | 12.93 ** | −2.35 * | 7.27 ** | −4.84 ** | −13.75 ** | −22.47 ** | ||
| P2 × P5 | 6.32 ** | 3.93 ** | −7.69 ** | −14.29 ** | −8.51 ** | −13.13 ** | ||
| P2 × P6 | 17.81 ** | 1.18 NS | −4.00 ** | −7.69 ** | −9.40 ** | −24.16 ** | ||
| P3 × P4 | 16.08 ** | 2.47 * | 4.92 ** | 3.23 ** | −9.40 ** | −13.46 ** | ||
| P3 × P5 | −2.35 * | −6.74 ** | −6.90 ** | −10.00 ** | −18.08 ** | −26.77 ** | ||
| P3 × P6 | −4.23 ** | −16.05 ** | −7.14 ** | −13.33 ** | −8.70 ** | −19.23 ** | ||
| P4 × P5 | 16.85 ** | 16.85 ** | 3.39 ** | 4.84 ** | 8.24 ** | −7.07 ** | ||
| P4 × P6 | 8.94 ** | 8.06 ** | −8.77 ** | −16.13 ** | −0.76 NS | −8.45 ** | ||
| P5 × P6 | 36.07 ** | 38.07 ** | 12.96 ** | 8.93 ** | −14.09 ** | −31.01 ** | ||
| Particle Size Distribution | Textural Class | Bulk Density (g/cm3) | pH | SOM (g/kg) | Total N (g/kg) | Exchangeable (Meq 100 g/soil) | Other Nutrients (mg/kg) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sand (%) | Silt (%) | Clay (%) | P | S | Zn | B | ||||||
| 26 | 18 | 56 | Clay loam | 1.42 | 7.3 | 1.25 | 0.065 | 0.17 | 12 | 14 | 0.57 | 0.17 |
| Sl. No. | Symbol | Parents/Cultivars | Pedigree | Sources | Special Features |
|---|---|---|---|---|---|
| 1 | P1 | BMXK1-14004 | Local cross | BARI, BD | High-yielding, medium seed, drought, and mungbean yellow mosaic virus (MYMV)-tolerant |
| 2 | P2 | BARI Mung-1 | Selection from NM92 | BARI, BD | High-yielding, bold seed and MYMV-tolerant |
| 3 | P3 | BINA Mung-8 | MB149 with 400 Gy dose | BINA, BD | High-yielding, small seed, and MYMV-tolerant |
| 4 | P4 | Sukumar | T-1 × K-441-11 | IIPR, India | High-yielding, bold seed and MYMV-tolerant |
| 5 | P5 | Pusa-7 (PS-7) | Selection from P-4092 | IIPR, India | High-yielding, small seed, MYMV susceptible |
| 6 | P6 | Sonali mung | Local | Local | Low-yielding, small seed and golden-colored, MYMV-tolerant |
| Sl. No. | Crosses | Cross Combinations |
|---|---|---|
| 1 | P1 × P2 | BMXK1-14004 × BARI Mung-1 |
| 2 | P1 × P3 | BMXK1-14004 × BINA Mung-8 |
| 3 | P1 × P4 | BMXK1-14004 × Sukumar |
| 4 | P1 × P5 | BMXK1-14004 × Pusa-7 (PS-7) |
| 5 | P1 × P6 | BMXK1-14004 × Sonali mung |
| 6 | P2 × P3 | BARI Mung-1 × BINA Mung-8 |
| 7 | P2 × P4 | BARI Mung-1 × Sukumar |
| 8 | P2 × P5 | BARI Mung-1 × Pusa-7 (PS-7) |
| 9 | P2 × P6 | BARI Mung-1 × Sonali mung |
| 10 | P3 × P4 | BINA Mung-8 × Sukumar |
| 11 | P3 × P5 | BINA Mung-8 × Pusa-7 (PS-7) |
| 12 | P3 × P6 | BINA Mung-8 × Sonali mung |
| 13 | P4 × P5 | Sukumar × Pusa-7 (PS-7) |
| 14 | P4 × P6 | Sukumar × Sonali mung |
| 15 | P5 × P6 | Pusa-7 (PS-7) × Sonali mung |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azam, M.G.; Sarker, U.; Hossain, M.A.; Iqbal, M.S.; Islam, M.R.; Hossain, M.F.; Ercisli, S.; Kul, R.; Assouguem, A.; AL-Huqail, A.A.; et al. Genetic Analysis in Grain Legumes [Vigna radiata (L.) Wilczek] for Yield Improvement and Identifying Heterotic Hybrids. Plants 2022, 11, 1774. https://doi.org/10.3390/plants11131774
Azam MG, Sarker U, Hossain MA, Iqbal MS, Islam MR, Hossain MF, Ercisli S, Kul R, Assouguem A, AL-Huqail AA, et al. Genetic Analysis in Grain Legumes [Vigna radiata (L.) Wilczek] for Yield Improvement and Identifying Heterotic Hybrids. Plants. 2022; 11(13):1774. https://doi.org/10.3390/plants11131774
Chicago/Turabian StyleAzam, Md. Golam, Umakanta Sarker, Md. Amir Hossain, Md. Shahin Iqbal, Md. Rafiqul Islam, Md. Faruk Hossain, Sezai Ercisli, Raziye Kul, Amine Assouguem, Arwa Abdulkreem AL-Huqail, and et al. 2022. "Genetic Analysis in Grain Legumes [Vigna radiata (L.) Wilczek] for Yield Improvement and Identifying Heterotic Hybrids" Plants 11, no. 13: 1774. https://doi.org/10.3390/plants11131774