
Isolation, Identification, and Biocontrol Potential of Root Fungal Endophytes Associated with Solanaceous Plants against Potato Late Blight (Phytophthora infestans)



Abstract
:1. Introduction
2. Results
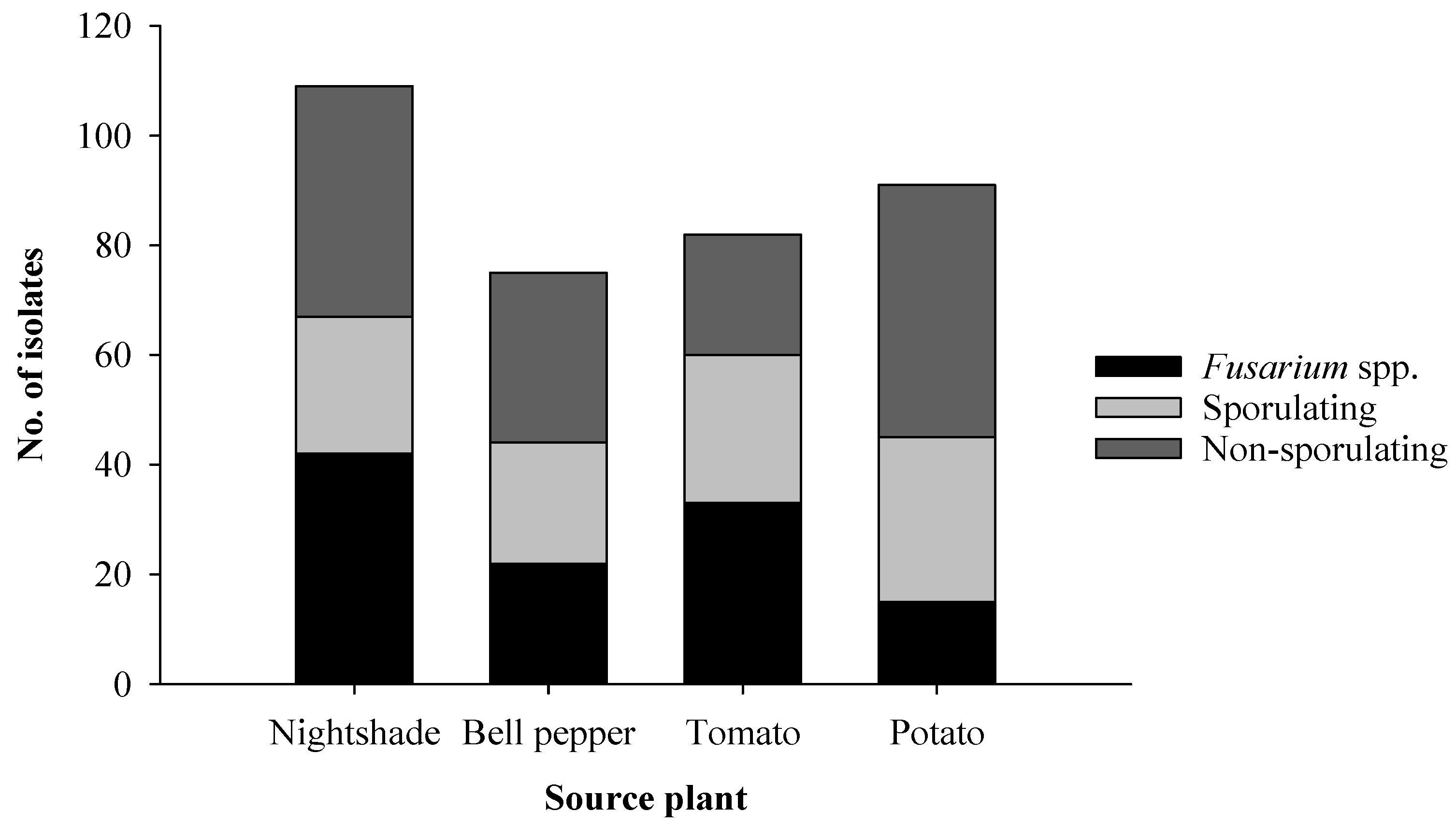
2.1. Isolation of Endophytic Fungi from Roots of Solanaceous Plants
2.2. Screening of Endophytes for Anti-Oomycete Activity
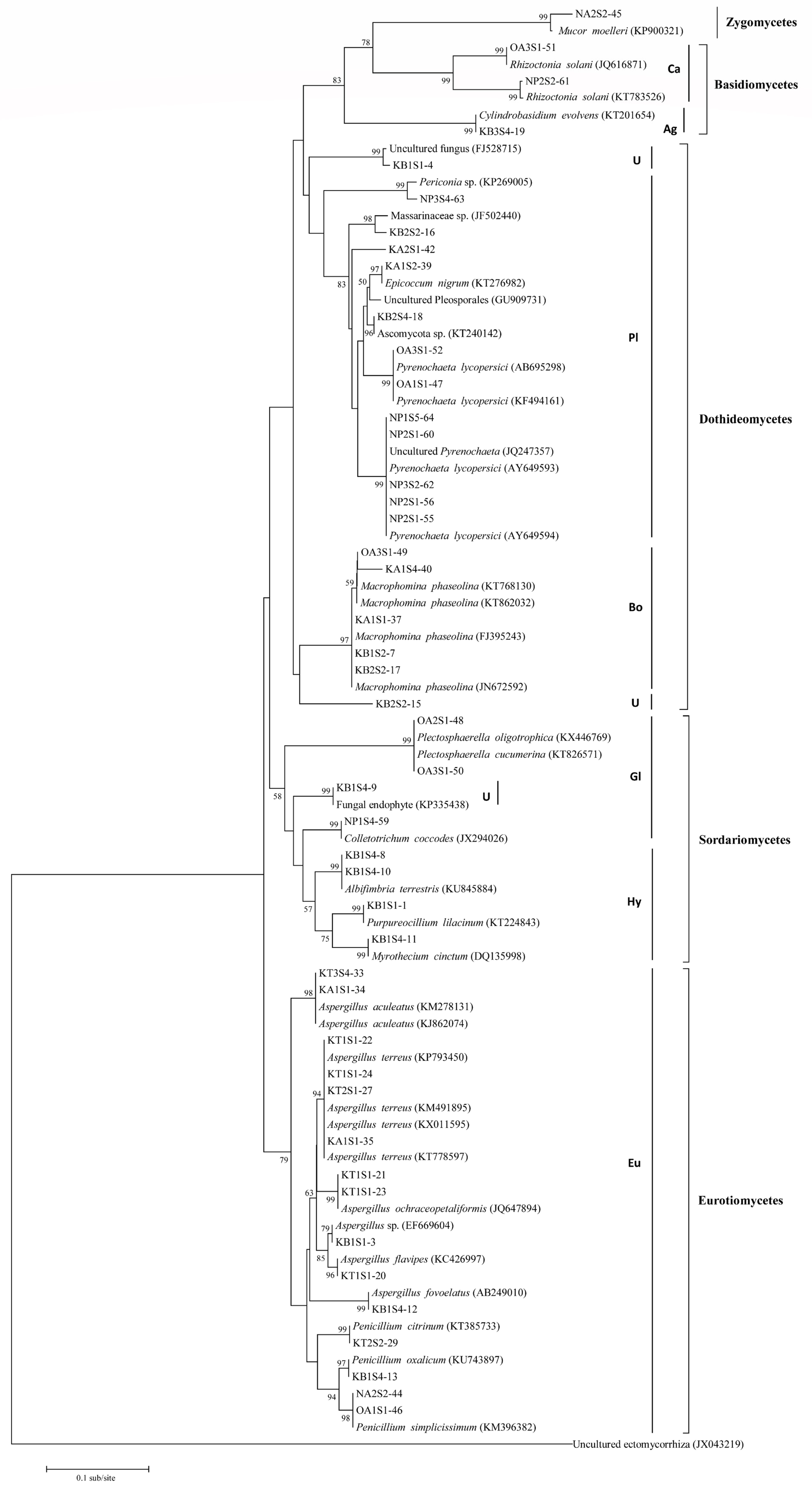
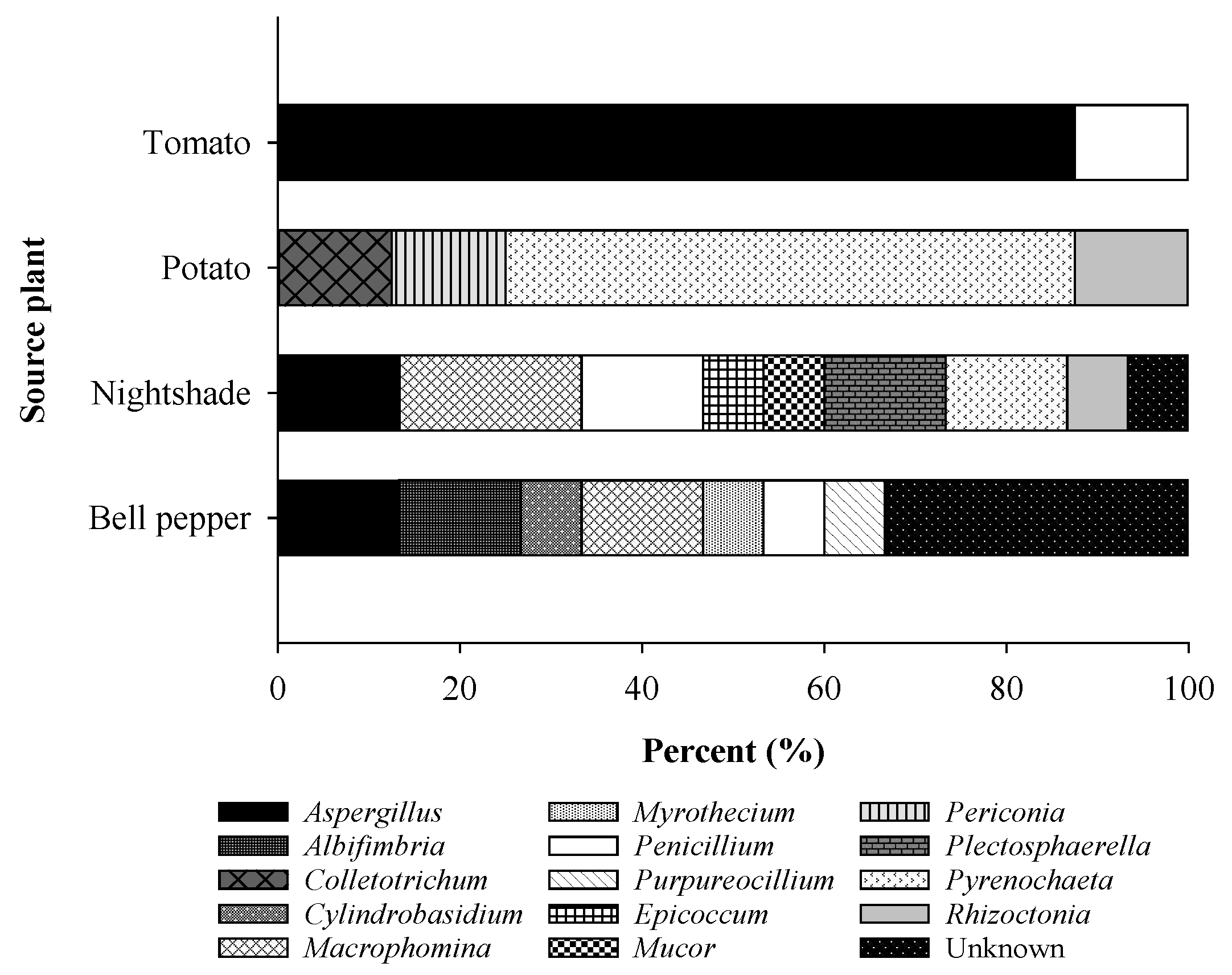
2.3. Characterization of the Fungal Endophytes
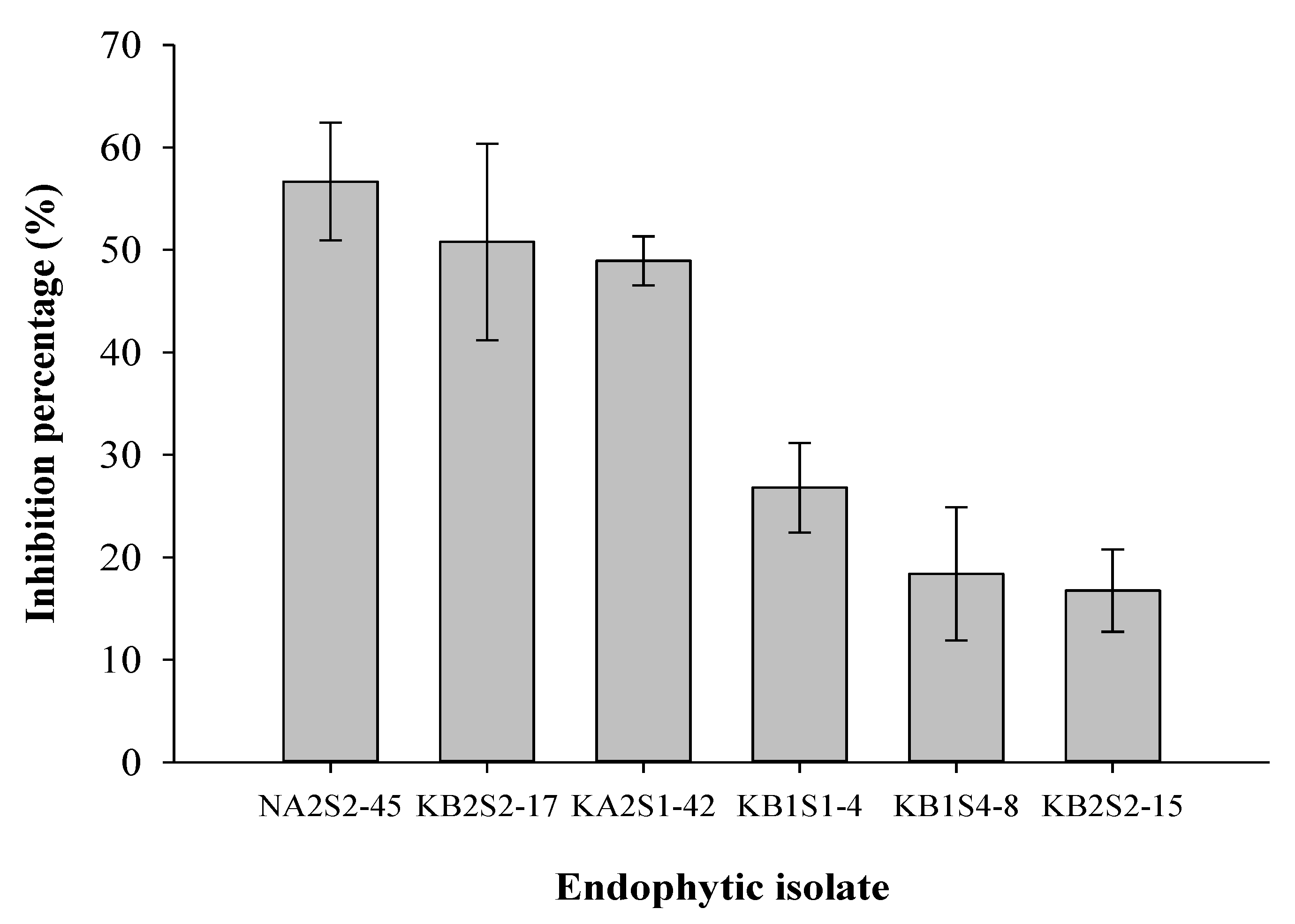
2.4. In Vitro Activity of Endophytic Fungi against Mycelial Growth of P. infestans
2.5. Potential of Endophytes Secreting Volatile Organic Compounds Active against P. infestans
2.6. Effect of Crude Extracts from Selected Endophytes on Sporangial Germination of P. infestans
2.7. In Vivo Activity of the Crude Extracts against P. infestans
3. Discussion
4. Materials and Methods
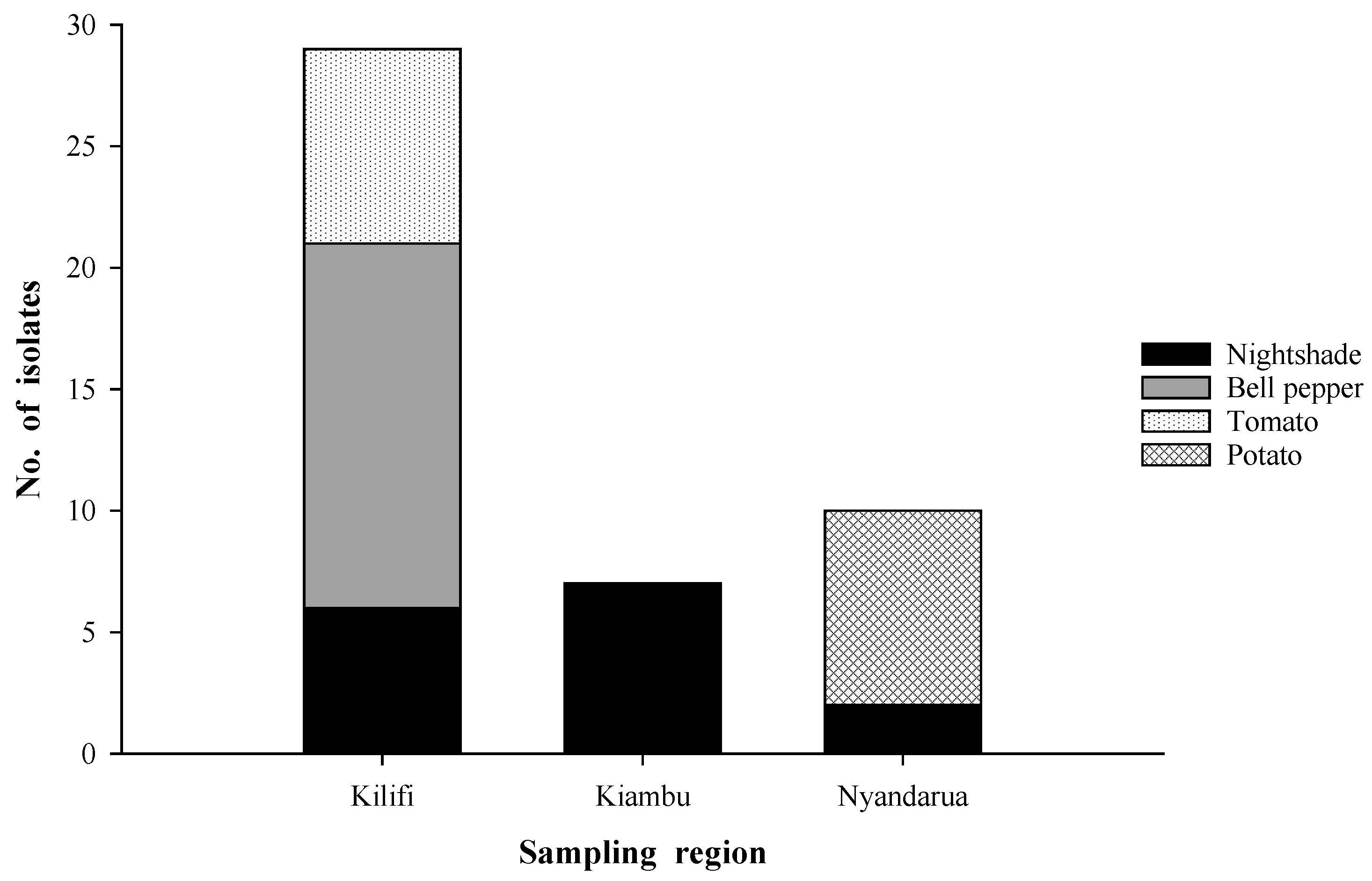
4.1. Sampling Regions and Collection of Plant Materials
4.2. Isolation of Fungal Root Endophytes and P. infestans
4.3. Primary Screening of Endophytes for Anti-Oomycete Activity against P. infestans
4.4. DNA Extraction, PCR Conditions, and Sequencing
4.5. Establishment the Anti-Oomycete Activity of the Endophytes
4.6. Impact of Volatile Organic Compounds Extracted from the Endophytes on Mycelial Growth of P. infestans
4.7. Suppressive Activity of Crude Extracts of Root Endophytic Fungi against Sporangial Germination of P. infestans
4.8. Activity of the Crude Extracts against P. infestans on Detached Potato Leaflets
4.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haverkort, A.J.; Struik, P.C.; Visser, R.G.F.; Jacobsen, E. Applied biotechnology to combat late blight in potato caused by Phytophthora Infestans. Potato Res. 2009, 52, 249–264. [Google Scholar] [CrossRef]
- Kromann, P.; Miethbauer, T.; Ortiz, O.; Forbes, G.A. Review of potato biotic constraints and experiences with integrated pest management interventions. In Integrated Pest Management; Pimental, D., Peshin, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 245–268. [Google Scholar]
- Ivanov, A.A.; Ukladov, E.O.; Golubeva, T.S. Phytophthora infestans: An Overview of Methods and Attempts to Combat Late Blight. J. Fungi 2021, 7, 1071. [Google Scholar] [CrossRef] [PubMed]
- Fry, W.E.; Birch, P.R.J.; Judelson, H.S.; Grünwald, N.J.; Danies, G.; Everts, K.L.; Gevens, A.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; et al. Five reasons to consider Phytophthora infestans a reemerging pathogen. Phytopathology 2015, 105, 966–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olanya, O.M.; Adipala, E.; Hakiza, J.J.; Kedera, J.C.; Ojiambo, P.; Mukalazi, J.M.; Forbes, G.; Nelson, R. Epidemiology and population dynamics of Phytophthora infestans in sub Saharan Africa. Afr. Crop Sci. J. 2001, 9, 185–194. [Google Scholar] [CrossRef]
- Chepsergon, J.; Motaung, T.E.; Bellieny-Rabelo, D.; Moleleki, L.N. Organize, Don’t Agonize: Strategic Success of Phytophthora Species. Microorganisms 2020, 8, 917. [Google Scholar] [CrossRef]
- Leach, S.S. Effects of copper and copper fungicide soil residues on Phytophthora infestans. Am. Potato J. 1966, 43, 431–438. [Google Scholar] [CrossRef]
- Bruck, R.I.; Fry, W.E.; Apple, A.E. Effect of metalaxyl, an acylalanine fungicide, on developmental stages of Phytophthora infestans. Phytopathology 1980, 70, 597–601. [Google Scholar] [CrossRef]
- Wale, S.; Platt, H.W.; Cattlin, N.D. Introduction. In Diseases, Pests and Disorders of Potatoes: A Colour Handbook; Wale, S.H., Platt, W., Cattlin, N.D., Eds.; Manson Publishing: London, UK, 2008; pp. 6–8. [Google Scholar]
- Cohen, Y.; Rubin, A.E.; Galperin, M. Effective control of two genotypes of Phytophthora infestans in the field by three oxathiapiprolin fungicidal mixtures. PLoS ONE 2021, 16, e0258280. [Google Scholar] [CrossRef]
- Rani, L.; Thapa, K.; Kanojia, N.; Sharma, N.; Singh, S.; Grewal, A.; Srivastav, A.L.; Kaushal, J. An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Prod. 2021, 283, 124657. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Choi, G.J.; Lee, H.B.; Lee, S.-W.; Lim, H.K.; Jang, K.S.; Son, S.W.; Lee, S.O.; Cho, K.Y.; Sung, N.D.; et al. Some fungal endophytes from vegetable crops and their anti-oomycete activities against tomato late blight. Lett. Appl. Microbiol. 2007, 44, 332–337. [Google Scholar] [CrossRef]
- Portz, D.; Koch, E.; Slusarenko, A.J. Effects of garlic (Allium sativum) juice containing allicin on Phytophthora infestans and downy mildew of cucumber caused by Pseudoperonospora cubensis. Eur. J. Plant Pathol. 2008, 122, 197–206. [Google Scholar] [CrossRef]
- Andrade-Linares, D.R.; Grosch, R.; Restrepo, S.; Krumbein, A.; Franken, P. Effects of dark septate endophytes on tomato plant performance. Mycorrhiza 2011, 21, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Tabet, D.; Sandroni, M.; Benavent-Celma, C.; Seematti, J.; Andersen, C.; Grenville-Briggs, L. The hunt for sustainable biocontrol of oomycete plant pathogens, a case study of Phytophthora infestans. Fungal Biol. Rev. 2022, 40, 53–69. [Google Scholar] [CrossRef]
- Miller, S.L. Functional diversity in fungi. Can. J. Bot. 1995, 73, 50–57. [Google Scholar] [CrossRef]
- Berg, G.; Krechel, A.; Lottmann, J.; Faltin, F.; Ulrich, A.; Hallmann, J.; Grosch, R. Endophytes: A new source for multi-target Biological Control Agents? IOBC/WPRS Bull. 2004, 27, 161–165. [Google Scholar]
- van Loon, L.C.; Bakker, P.A.H.M.; Pieterse, C.M.J. Systemic resistance induced by rhizospere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [Green Version]
- Kloepper, J.W.; Ryu, C.-M. Bacterial endophytes as elicitors of induced systemic resistance. In Microbial Root Endophytes, SOIL Biology; Schulz, B., Boyle, C., Sieber, T.N., Eds.; Springer Science + Business Media Verlag: Berlin, Germany, 2006; Volume 9, pp. 33–52. [Google Scholar]
- Feng, Y.; Shen, D.; Song, W. Rice endophyte Pantoea agglomerans YS19 promotes host plant growth and affects allocations of host photosynthates. J. Appl. Microbiol. 2006, 100, 938–945. [Google Scholar] [CrossRef]
- Mizubuti, E.S.G.; Júnior, L.V.; Gregory, F.A. Management of late blight with alternative products. Pest. Technol. 2007, 1, 106–116. [Google Scholar]
- De Vrieze, M.; Germanier, F.; Vuille, N.; Weisskopf, L. Combining Different Potato-Associated Pseudomonas Strains for Improved Biocontrol of Phytophthora infestans. Front. Microbiol. 2018, 9, 2573. [Google Scholar] [CrossRef]
- Ng, K.K.; Webster, J.M. Oomycete activity of Xenorhabdus bovienii. Can. J. Plant Pathol. 1997, 2, 125–132. [Google Scholar] [CrossRef]
- Daayf, F.; Adam, L.; Fernando, W.G.D. Comparative screening of bacteria for biological control of potato late blight (strain US-8), using in-vitro, detached-leaves, and whole-plant testing systems. Can. J. Plant Pathol. 2003, 25, 276–284. [Google Scholar] [CrossRef]
- Stephan, D.; Schmitt, A.; Carvalho, S.M.; Seddon, B.; Koch, E. Evaluation of biocontrol preparations and plant extracts for the control of Phytophthora infestans on potato leaves. Eur. J. Plant Pathol. 2005, 112, 235–246. [Google Scholar] [CrossRef]
- Loliam, B.; Morinaga, T.; Chaiyanan, S. Biocontrol of Phytophthora infestans, fungal pathogen of seedling damping off disease in economic plant nursery. Psyche 2012, 2012, 324317. [Google Scholar] [CrossRef]
- Miles, L.A.; Lopera, C.A.; González, S.; Cepero de García, M.C.; Franco, A.E.; Restrepo, S. Exploring the biocontrol potential of fungal endophytes from an Andean Colombian Paramo ecosystem. BioControl 2012, 57, 697–710. [Google Scholar] [CrossRef]
- Gupta, J. Efficacy of biocontrol agents against Phytophthora infestans on potato. Int. J. Eng. Sci. 2016, 6, 2249–2252. [Google Scholar]
- Al-Mughrabi, K.I. Biological control of Phytophthora infestans of potatoes using Trichoderma atroviride. Pest. Technol. 2008, 2, 104–108. [Google Scholar]
- Andrade-Linares, D.R.; Grosch, R.; Franken, P.; Rexer, K.-H.; Kost, G.; Restrepo, S.; de Garcia, M.C.C.; Maximova, E. Colonization of roots of cultivated Solanum lycopersicum by dark septate and other ascomycetous endophytes. Mycologia 2011, 103, 710–721. [Google Scholar] [CrossRef]
- Kannan, K.P.; Muthumary, J. Comparative analysis of endophytic mycobiota in different tissues of medicinal plants. Afr. J. Microbiol. Res. 2012, 6, 4219–4225. [Google Scholar]
- de Vries, S.; von Dahlen, J.K.; Schnake, A.; Ginschel, S.; Schulz, B.; Rose, L.E. Broad-spectrum inhibition of Phytophthora infestans by root endophytes. FEMS Microbiol. Ecol. 2018, 94, fiy037. [Google Scholar] [CrossRef] [Green Version]
- Kusari, P.; Kusari, S.; Spiteller, M.; Kayser, O. Endophytic fungi harbored in Cannabis sativa L.: Diversity and potential as biocontrol agents against host plant-specific phytopathogens. Fungal Divers. 2012, 60, 137–151. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Lv, Y.; Zhang, F.; Chen, J.; Cui, J.; Xing, Y.; Li, X.; Guo, S. Diversity and antimicrobial activity of endophytic fungi associated with the alpine plant Saussurea involucrata. Biol. Pharm. Bull. 2010, 33, 1300–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokati, D.; Herrera, J.; Poudel, R. Soil influences colonization of root-associated fungal endophyte communities of maize, wheat and their progenitors. J. Mycol. 2016, 2016, 8062073. [Google Scholar] [CrossRef]
- Bogner, C.W.; Kariuki, G.M.; Elashry, A.; Sichtermann, G.; Buch, A.; Mishra, B.; Thines, M.; Grundler, F.M.W.; Schouten, A. Fungal root endophytes of tomato from Kenya and their nematode biocontrol potential. Mycol. Prog. 2016, 15, 30. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Vitorino, L.C.; Silva, F.G.; Soares, M.A.; Souchie, E.L.; Lima, W.C. The isolation and characterization of endophytic microorganisms from Hyptis marrubioides Epling roots. Afr. J. Biotechnol. 2012, 11, 12766–12772. [Google Scholar]
- Karst, J.; Piculell, B.; Brigham, C.; Booth, M.; Hoeksema, J.D. Fungal communities in soils along a vegetative ecotone. Mycologia 2013, 105, 61–70. [Google Scholar] [CrossRef]
- Lau, M.K.; Arnold, A.E.; Johnson, N.C. Factors influencing communities of foliar fungal endophytes in riparian woody plants. Fungal Ecol. 2013, 6, 365–378. [Google Scholar] [CrossRef]
- Chowdhary, K.; Kaushik, N. Fungal endophyte diversity and bioactivity in the Indian medicinal plant Ocimum sanctum. PLoS ONE 2015, 10, e0141444. [Google Scholar] [CrossRef]
- Nartey, L.; Pu, Q.; Zhu, W.; Zhang, S.; Li, J.; Yao, Y.; Hu, X. Antagonistic and plant growth promotion effects of Mucor moelleri, a potential biocontrol agent. Microbiol. Res. 2022, 255, 126922. [Google Scholar] [CrossRef]
- El-Hasan, A.; Walker, F.; Schöne, J.; Buchenauer, H. Antagonistic effect of 6-pentyl-alpha-pyrone produced by Trichoderma harzianum toward Fusarium moniliforme. J. Plant Dis. Prot. 2007, 114, 62–68. [Google Scholar] [CrossRef]
- El-Hasan, A.; Walker, F.; Schöne, J.; Buchenauer, H. Detection of viridiofungin A and other antifungal metabolites excreted by Trichoderma harzianum active against different plant pathogens. Eur. J. Plant Pathol. 2009, 124, 457–470. [Google Scholar] [CrossRef]
- El-Hasan, A.; Walker, F.; Klaiber, I.; Schöne, J.; Pfannstiel, J.; Voegele, R.T. New Approaches to Manage Asian Soybean Rust (Phakopsora pachyrhizi) Using Trichoderma spp. or Their Antifungal Secondary Metabolites. Metabolites 2022, 12, 507. [Google Scholar] [CrossRef]
- Matroudi, S.; Zamani, M.R.; Motallebi, M. Antagonistic effects of three species of Trichoderma sp. on Sclerotinia sclerotiorum, the causal agent of canola stem rot. Egypt. J. Biol. 2009, 11, 37–44. [Google Scholar]
- Tripathi, A.; Sharma, N.; Tripathi, N. Biological control of plant diseases: An overview and the Trichoderma system as biocontrol agents. In Management of Fungal Plant Pathogens; Arya, A., Perelló, A.E., Eds.; CAB International: Oxfordshire, UK, 2010; pp. 110–121. [Google Scholar]
- Massimo, N.C.; Devan, M.M.N.; Arendt, K.R.; Wilch, M.H.; Riddle, J.M.; Furr, S.H.; Steen, C.; U’Ren, J.M.; Sandberg, D.C.; Arnold, E. Fungal endophytes in above-ground tissues of desert plants: Infrequent in culture, but highly diverse and distinctive symbionts. Microb. Ecol. 2015, 70, 61–76. [Google Scholar] [CrossRef]
- Vila, L.; Lacadena, V.; Fontanet, P.; del Pozo, M.A.; Segundo, B.S. A protein from the mold Aspergillus giganteus is a potent inhibitor of fungal plant pathogens. Mol. Plant-Microbe Interact. 2001, 14, 1327–1331. [Google Scholar] [CrossRef] [Green Version]
- Vasiliauskas, R.; Stenlid, J. Fungi inhabiting stems of Picea abies in a managed stand in Lithuania. For. Ecol. Manag. 1998, 109, 119–126. [Google Scholar] [CrossRef]
- Vasiliauskas, R.; Stenlid, J. Population structure and genetic variation in Cylindrobasidium evolvens. Mycol. Res. 1998, 102, 1453–1458. [Google Scholar] [CrossRef]
- Warrior, P.; Rehberger, L.A.; Beach, M.; Grau, P.A.; Kirfman, G.W.; Conley, J.M. Commercial development and introduction of DiTeraTM, a new nematicide. Pest. Manag. Sci. 1999, 59, 376–379. [Google Scholar] [CrossRef]
- Hoagland, R.E.; Boyette, D.C.; Abbas, H.K. Myrothecium verrucaria isolates and formulations as bioherbicide agents for kudzu. Biocontrol Sci. Technol. 2007, 17, 721–731. [Google Scholar] [CrossRef]
- Lombard, L.; Houbraken, J.; Decock, C.; Samson, R.A.; Meijer, M.; Réblová, M.; Groenewald, J.Z.; Crous, P.W. Generic hyper-diversity in Stachybotriaceae. Persoonia 2016, 36, 156–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurakov, A.V.; Lavrent, R.B.; Nechitailo, T.Y.; Golyshin, P.N.; Zvyagintsev, D.G. Diversity of facultatively anaerobic microscopic mycelial fungi in soils. Microbiology 2008, 77, 90–98. [Google Scholar] [CrossRef]
- Zou, X.; Niu, S.; Ren, J.; Li, E.; Liu, X.; Che, Y. Verrucamides A–D, antibacterial cyclopeptides from Myrothecium verrucaria. J. Nat. Prod. 2011, 74, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Walker, H.L.; Tilley, A.M. Evaluation of an isolate of Myrothecium verrucaria from sicklepod (Senna obtusifolia) as a potential mycoherbicide agent. Biol. Control 1997, 10, 104–112. [Google Scholar] [CrossRef]
- Li, Z.; Chang, P.; Gao, L.; Wang, X. The endophytic fungus Albifimbria verrucaria from wild grape as an antagonist of Botrytis cinerea and other grape pathogens. Phytopathology 2019, 110, 843–850. [Google Scholar] [CrossRef]
- Banerjee, D.; Strobel, G.A.; Booth, E.; Geary, B.; Sears, J.; Spakowicz, D.; Busse, S. An endophytic Myrothecium inundatum producing volatile organic compounds. Mycosphere 2010, 1, 229–240. [Google Scholar]
- Boddy, L. Interactions between fungi and other microbes. In The Fungi, 3rd ed.; Watkinson, S.C., Boddy, L., Money, N.P., Eds.; Academic Press: London, UK, 2016; pp. 337–360. [Google Scholar]
- Linkies, A.; Jacob, S.; Zink, P.; Maschemer, M.; Maier, W.; Koch, E. Characterization of cultural traits and fungicidal activity of strains belonging to the fungal genus Chaetomium. J. Appl. Microbiol. 2021, 131, 375–391. [Google Scholar] [CrossRef]
- Bae, S.J.; Mohanta, T.K.; Chung, J.Y.; Ryu, M.; Park, G.; Shim, S.; Hong, S.-B.; Seo, H.; Bae, D.-W.; Bae, I.; et al. Trichoderma metabolites as biological control agents against Phytophthora pathogens. Biol. Control 2016, 92, 128–138. [Google Scholar] [CrossRef]
- Chandrakala, A.; Chadrashekar, S.C.; Jyothi, G.; Ravikumar, B.M. Effect of cell-free culture filtrates of bio-control agents on the spore germination and infection by Phytophthora infestans causing late blight of potato. Glob. J. Biol. Agric. Health Sci. 2012, 1, 40–45. [Google Scholar]
- Tellenbach, C.; Sumarah, M.W.; Grünig, C.R.; Miller, J.D. Inhibition of Phytophthora species by secondary metabolites produced by the dark septate endophyte Phialocephala europaea. Fungal Ecol. 2013, 6, 12–18. [Google Scholar] [CrossRef]
- Bae, H.; Roberts, D.P.; Lim, H.-S.; Strem, M.D.; Park, S.-C.; Ryu, C.-M.; Melnick, R.L.; Bailey, B.A. Endophytic Trichoderma isolates from tropical environments delay disease onset and induce resistance against Phytophthora capsici in hot pepper using multiple mechanisms. Mol. Plant-Microbe Interact. 2011, 24, 336–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte, E.E.; Hoskins, B. Recommended soil organic matter tests. In Recommended Soil Testing Procedures for the Northeastern United States, The Northeast Coordinating Committee for Soil Testing (NEC 1812), Northeastern Regional Publication Cooperative Bulletin No. 493, 3rd ed.; Agricultural Experiment Station, Univ. of Delaware: Newark, NY, USA, 2011; pp. 63–74. [Google Scholar]
- Liu, D.; Coloe, S.; Baird, R.; Pedersen, J. Rapid mini-preparation of fungal DNA for PCR. J. Clin. Microbiol. 2000, 38, 471. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. PCR protocols: A guide to methods and applications. In Amplification and Direct Sequencing of Fungal Ribosomal DNA Genes for Phylogenetics; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Edgington, L.V.; Khew, K.L.; Barron, G.L. Fungitoxic spectrum of benzimidazole compounds. Phytopathology 1971, 61, 42–44. [Google Scholar] [CrossRef]
- El-Hasan, A.; Schöne, J.; Höglinger, B.; Walker, F.; Voegele, R.T. Assessment of the antifungal activity of selected biocontrol agents and their secondary metabolites against Fusarium graminearum. Eur. J. Plant Pathol. 2018, 150, 91–103. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Accession Numbers | Host Plant | Best BLAST Match (Accession Number) | Identity (%) |
|---|---|---|---|---|
| KB1S2-7 | MG214581 | Capsicum annum | Macrophomina phaseolina (KT862032) | 99 |
| NA2S2-45 | MG214587 | Solanum nigrum | Mucor moelleri (KP900321) | 99 |
| KA1S1-37 | MG214583 | S. nigrum | Macrophomina phaseolina (FJ395243) | 100 |
| OA3S1-49 | MG214585 | S. nigrum | Macrophomina phaseolina (KT768130) | 100 |
| KA1S4-40 | MG214584 | S. nigrum | Macrophomina phaseolina (KT768130) | 100 |
| OA3S1-51 | MG214604 | S. nigrum | Rhizoctonia solani (JQ616871) | 100 |
| NP2S2-61 | MG214605 | S. tuberosum | Rhizoctonia solani (KT783526) | 99 |
| KA1S1-34 | MG214568 | S. nigrum | Aspergillus aculeatus (KJ862074) | 100 |
| KB1S1-3 | MG214573 | C. annum | Aspergillus sp. (EF669604) | 99 |
| KT1S1-20 | MG214569 | Lycopersicon esculentum | Aspergillus flavipes (KC426997) | 100 |
| OA1S1-46 | MG214592 | S. nigrum | Penicillium simplicissimum (KM396382) | 99 |
| KB1S4-8 | MG214560 | C. annum | Albifimbria terrestris (KU845884) | 99 |
| KB2S2-17 | MG214582 | C. annum | Macrophomina phaseolina (JN672592) | 100 |
| KB2S4-18 | MG214565 | C. annum | Ascomycota sp. (KT240142) | 100 |
| KB1S4-10 | MG214561 | C. annum | Albifimbria terrestris (KU845884) | 99 |
| KT1S1-21 | MG214571 | L. esculentum | Aspergillus ochraceopetaliformis (JQ647894) | 100 |
| KT1S1-24 | MG214575 | L. esculentum | Aspergillus terreus (KP793450) | 99 |
| KT2S1-27 | MG214576 | L. esculentum | Aspergillus terreus (KX011595) | 100 |
| KB1S4-13 | MG214590 | C. annum | Penicillium oxalicum (KU743897) | 100 |
| KB3S4-19 | MG214579 | C. annum | Cylindrobasidium evolvens (KT201654) | 100 |
| KT3S4-33 | MG214567 | L. esculentum | Aspergillus aculeatus (KM278131) | 100 |
| KT1S1-22 | MG214574 | L. esculentum | Aspergillus terreus (KT778597) | 100 |
| OA3S1-52 | MG214598 | S. nigrum | Pyrenochaeta lycopersici (AB695298) | 99 |
| KB2S2-16 | MG214586 | C. annum | Massarinaceae sp. (JF502440) | 97 |
| KB1S4-9 | MG214563 | C. annum | Fungal endophyte isolate (KP335438) | 99 |
| NP1S4-59 | MG214578 | S. tuberosum | Colletotrichum coccodes (JX294026) | 99 |
| NA2S2-44 | MG214591 | S. nigrum | Penicillium simplicissimum (KM396382) | 99 |
| KA1S2-39 | MG214580 | S. nigrum | Epicoccum nigrum (KT276982) | 100 |
| OA2S1-48 | MG214595 | S. nigrum | Plectosphaerella oligotrophica (KX446769) | 99 |
| KB1S4-11 | MG214588 | C. annum | Myrothecium cinctum (DQ135998) | 100 |
| OA3S1-50 | MG214594 | S. nigrum | Plectosphaerella cucumerina (KT826571) | 100 |
| KB1S4-12 | MG214570 | C. annum | Aspergillus foveolatus (AB249010) | 100 |
| KT1S1-23 | MG214572 | L. esculentum | Aspergillus ochraceopetaliformis (JQ647894) | 100 |
| KA1S1-35 | MG214577 | S. nigrum | Aspergillus terreus (KM491895) | 99 |
| NP1S5-64 | MG214602 | S. tuberosum | Pyrenochaeta lycopersici (AY649593) | 99 |
| NP2S1-55 | MG214599 | S. tuberosum | Pyrenochaeta lycopersici (AY649594) | 100 |
| NP2S1-60 | MG214603 | S. tuberosum | Uncultured Pyrenochaeta (JQ247357) | 100 |
| KB1S1-1 | MG214596 | C. annum | Purpureocillium lilacinum (KT224843) | 100 |
| NP3S4-63 | MG214593 | S. tuberosum | Periconia sp. (KP269005) | 97 |
| KA2S1-42 | MG214566 | S. nigrum | Uncultured Pleosporales (GU909731) | 94 |
| NP2S1-56 | MG214600 | S. tuberosum | Pyrenochaeta lycopersici (AY649594) | 100 |
| NP3S2-62 | MG214601 | S. tuberosum | Pyrenochaeta lycopersici (AY649594) | 100 |
| OA1S1-47 | MG214597 | S. nigrum | Pyrenochaeta lycopersici (KF494161) | 100 |
| KB1S1-4 | MG214562 | C. annum | Uncultured fungus (FJ528715) | 99 |
| KT2S2-29 | MG214589 | L. esculentum | Penicillium citrinum (KT385733) | 100 |
| KB2S2-15 | MG214564 | C. annum | Uncultured ectomycorrhiza (JX043219) | 91 |
| Isolate | Highest BLAST Affinities | Mycelial Growth Inhibition (%) * | Inhibition Zone (mm) * |
|---|---|---|---|
| T16 | Trichoderma harzianum | 84.5 a | - |
| KB1S2-7 | Macrophomina phaseolina | 78.8 ab | - |
| KA1S1-34 | Aspergillus aculeatus | 76.7 ab | - |
| NA2S2-45 | Mucor moelleri | 73.8 ab | - |
| KB2S2-17 | Macrophomina phaseolina | 72.0 ab | - |
| KA1S1-37 | Macrophomina phaseolina | 70.4 ab | - |
| OA3S1-49 | Macrophomina phaseolina | 70.2 ab | - |
| KA1S4-40 | Macrophomina phaseolina | 69.2 ab | - |
| OA3S1-51 | Rhizoctonia solani | 69.2 ab | - |
| NP2S2-61 | Rhizoctonia solani | 68.3 bc | - |
| KB1S1-3 | Aspergillus sp. | 68.2 bc | - |
| OA1S1-46 | Penicillium simplicissimum | 56.3 dc | - |
| KB2S4-18 | Ascomycota sp. | 54.1 de | - |
| KT1S1-23 | Aspergillus ochraceopetaliformis | 53.5 de | 9.3 cd |
| KA1S1-35 | Aspergillus terreus | 52.0 d–f | 7.0 c–e |
| KB1S4-10 | Albifimbria terrestris | 51.8 d–f | - |
| KT1S1-21 | Aspergillus ochraceopetaliformis | 49.5 d–g | - |
| KT1S1-24 | Aspergillus terreus | 47.0 d–h | - |
| NP1S4-59 | Colletotrichum coccodes | 46.9 d–h | - |
| KT2S1-27 | Aspergillus terreus | 46.5 d–h | - |
| NA2S2-44 | Penicillium simplicissimum | 45.0 d–i | - |
| NP1S5-64 | Pyrenochaeta lycopersici | 43.9 d–j | 10.0 cd |
| KB1S4-13 | Penicillium oxalicum | 43.5 d–j | - |
| NP2S1-55 | Pyrenochaeta lycopersici | 42.9 e–k | 7.6 c–e |
| KB1S4-8 | Albifimbria terrestris | 41.5 e–k | - |
| NP2S1-60 | Uncultured Pyrenochaeta | 39.4 f–l | 17.0 a |
| KB1S1-1 | Purpureocillium lilacinum | 39.0 f–l | 6.5 de |
| KA1S2-39 | Epicoccum nigrum | 39.0 f–l | - |
| NP3S4-63 | Periconia sp. | 38.4 g–m | 11.0 bc |
| KB3S4-19 | Cylindrobasidium evolvens | 37.3 g–m | - |
| KA2S1-42 | Uncultured Pleosporales | 37.0 g–n | 18.3 a |
| NP2S1-56 | Pyrenochaeta lycopersici | 36.5 g–n | 11.3 bc |
| KT3S4-33 | Aspergillus aculeatus | 36.4 g–o | - |
| NP3S2-62 | Pyrenochaeta lycopersici | 34.5 h–o | 7.3 c–e |
| KT1S1-20 | Aspergillus flavipes | 32.2 i–p | - |
| OA1S1-47 | Pyrenochaeta lycopersici | 31.4 j–q | 8.0 c–e |
| KT1S1-22 | Aspergillus terreus | 31.1 j–q | - |
| OA2S1-48 | Plectosphaerella oligotrophica | 30.5 k–q | - |
| KB1S4-11 | Myrothecium cinctum | 28.1 l–r | - |
| OA3S1-50 | Plectosphaerella cucumerina | 26.1 m–r | - |
| OA3S1-52 | Pyrenochaeta lycopersici | 24.7 n–r | - |
| KB1S1-4 | Uncultured fungus | 24.2 o–r | 18.8 a |
| KB1S4-12 | Aspergillus foveolatus | 22.3 p–s | - |
| KB2S2-16 | Massarinaceae sp. | 20.1 q–s | - |
| KB1S4-9 | Fungal endophyte isolate | 20.0 q–s | - |
| KT2S2-29 | Penicillium citrinum | 18.9 rs | 4.8 e |
| KB2S2-15 | Uncultured ectomycorrhiza | 13.3 s | 14.8 ab |
| CEs Source | Inhibition (%) 1 | |
|---|---|---|
| Sporangial Germination 2 | Germ tube Growth 3 | |
| KB2S2-15 | 100.00 a | 100.00 a |
| KA2S1-42 | 100.00 a | 100.00 a |
| KB1S1-4 | 69.38 b | 82.73 b |
| NA2S2-45 | 30.04 c | −1.92 e |
| Acetone (solvent) control | 0.00 d | 0.00 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Hasan, A.; Ngatia, G.; Link, T.I.; Voegele, R.T. Isolation, Identification, and Biocontrol Potential of Root Fungal Endophytes Associated with Solanaceous Plants against Potato Late Blight (Phytophthora infestans). Plants 2022, 11, 1605. https://doi.org/10.3390/plants11121605
El-Hasan A, Ngatia G, Link TI, Voegele RT. Isolation, Identification, and Biocontrol Potential of Root Fungal Endophytes Associated with Solanaceous Plants against Potato Late Blight (Phytophthora infestans). Plants. 2022; 11(12):1605. https://doi.org/10.3390/plants11121605
Chicago/Turabian StyleEl-Hasan, Abbas, Grace Ngatia, Tobias I. Link, and Ralf T. Voegele. 2022. "Isolation, Identification, and Biocontrol Potential of Root Fungal Endophytes Associated with Solanaceous Plants against Potato Late Blight (Phytophthora infestans)" Plants 11, no. 12: 1605. https://doi.org/10.3390/plants11121605