
Optimization of Biomass Accumulation and Production of Phenolic Compounds in Callus Cultures of Rhodiola rosea L. Using Design of Experiments

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
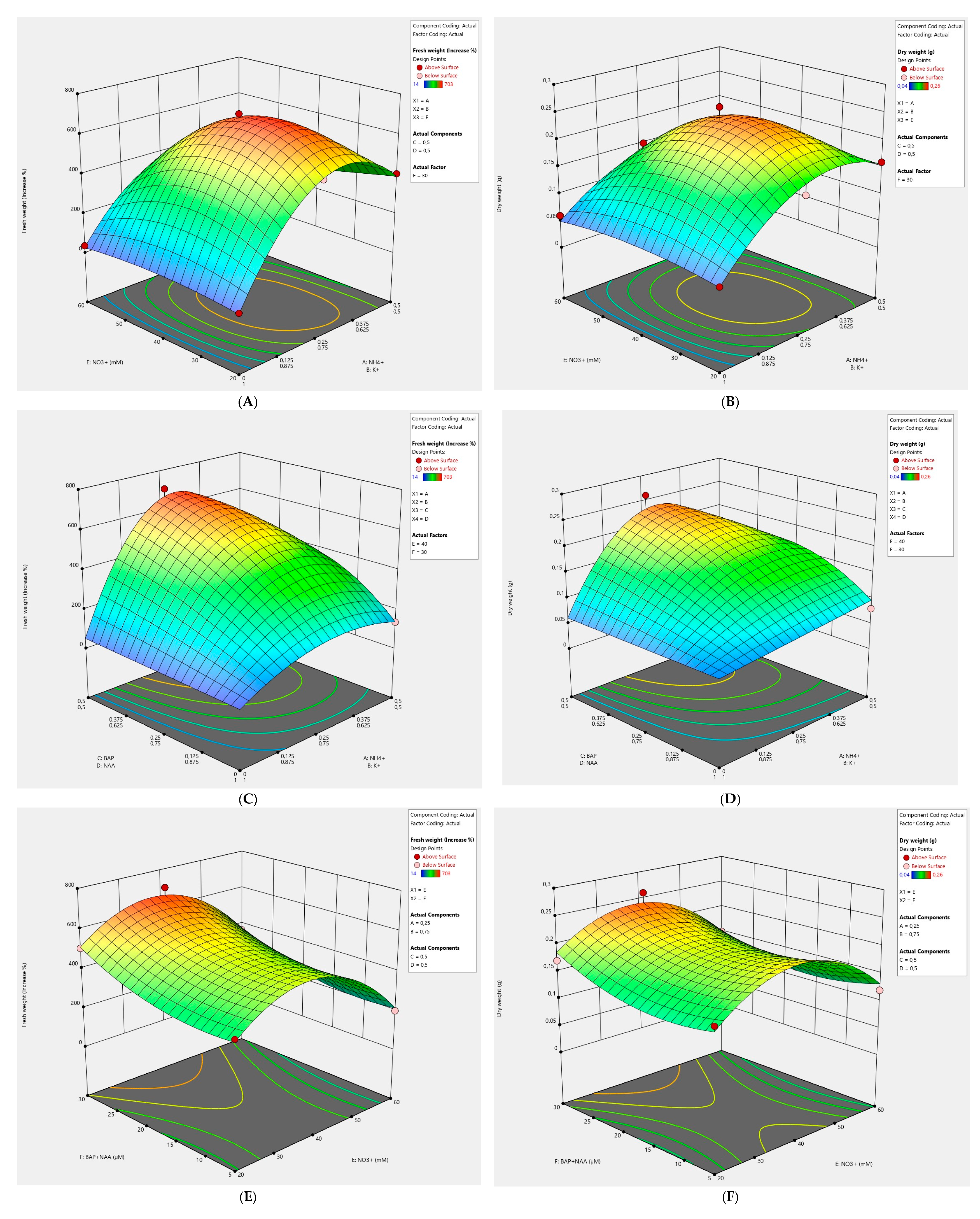
2.1. Effects of the NH4+/K+ Ratio and NO3− Concentration on Callus Culture
2.2. Effects of Ratios BAP/NAA and NH4+/K+ on Callus Culture
2.3. Effects of BAP + NAA and NO3− Concentrations on Callus Culture
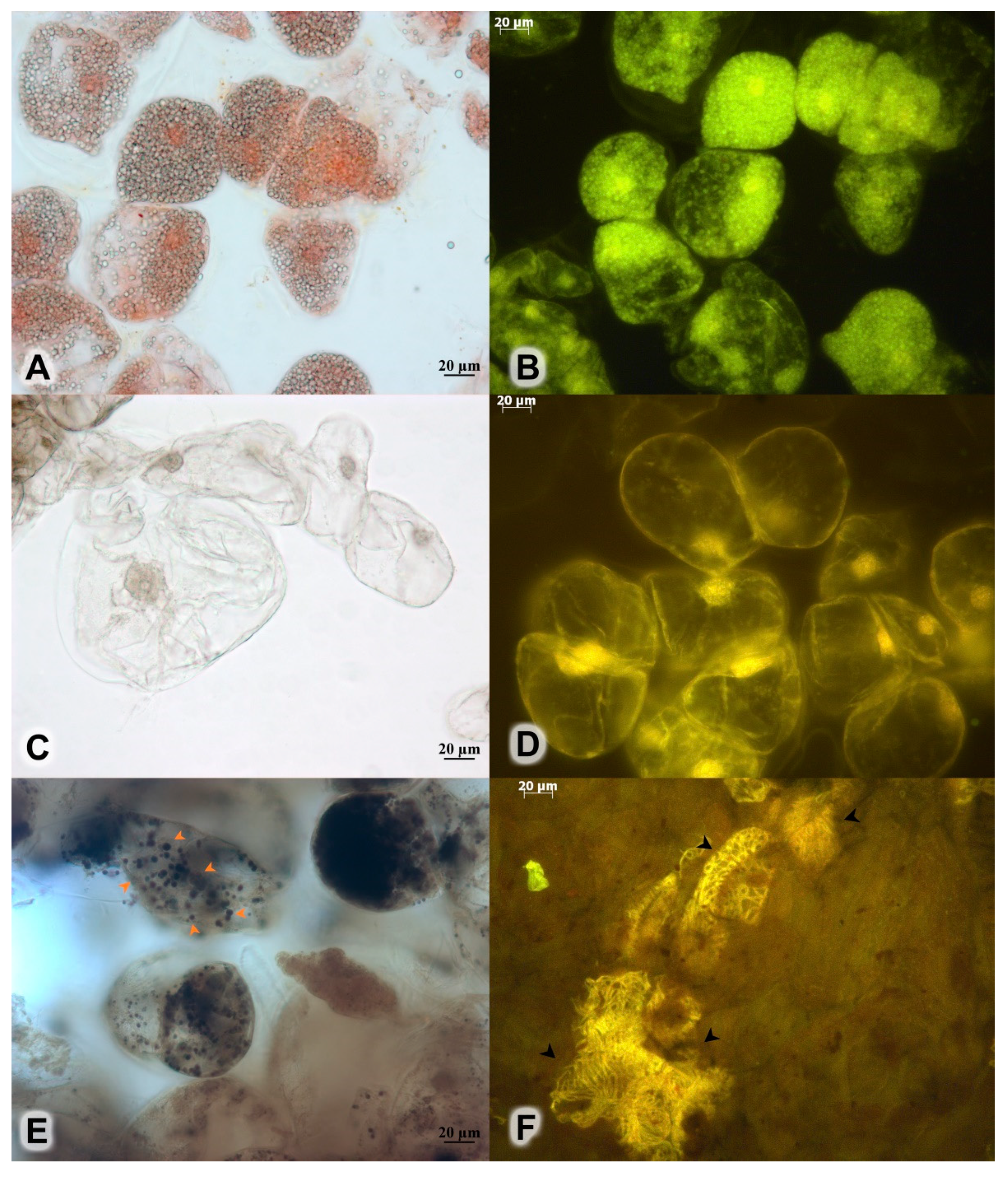
2.4. Histochemical Analysis
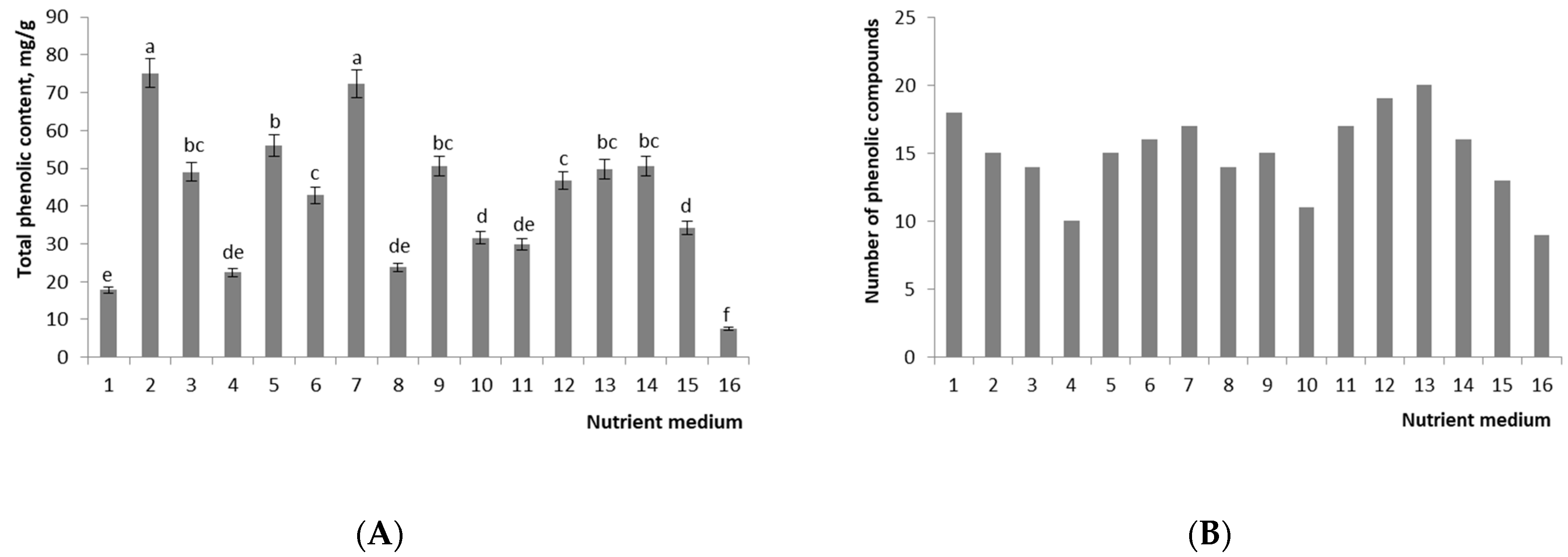
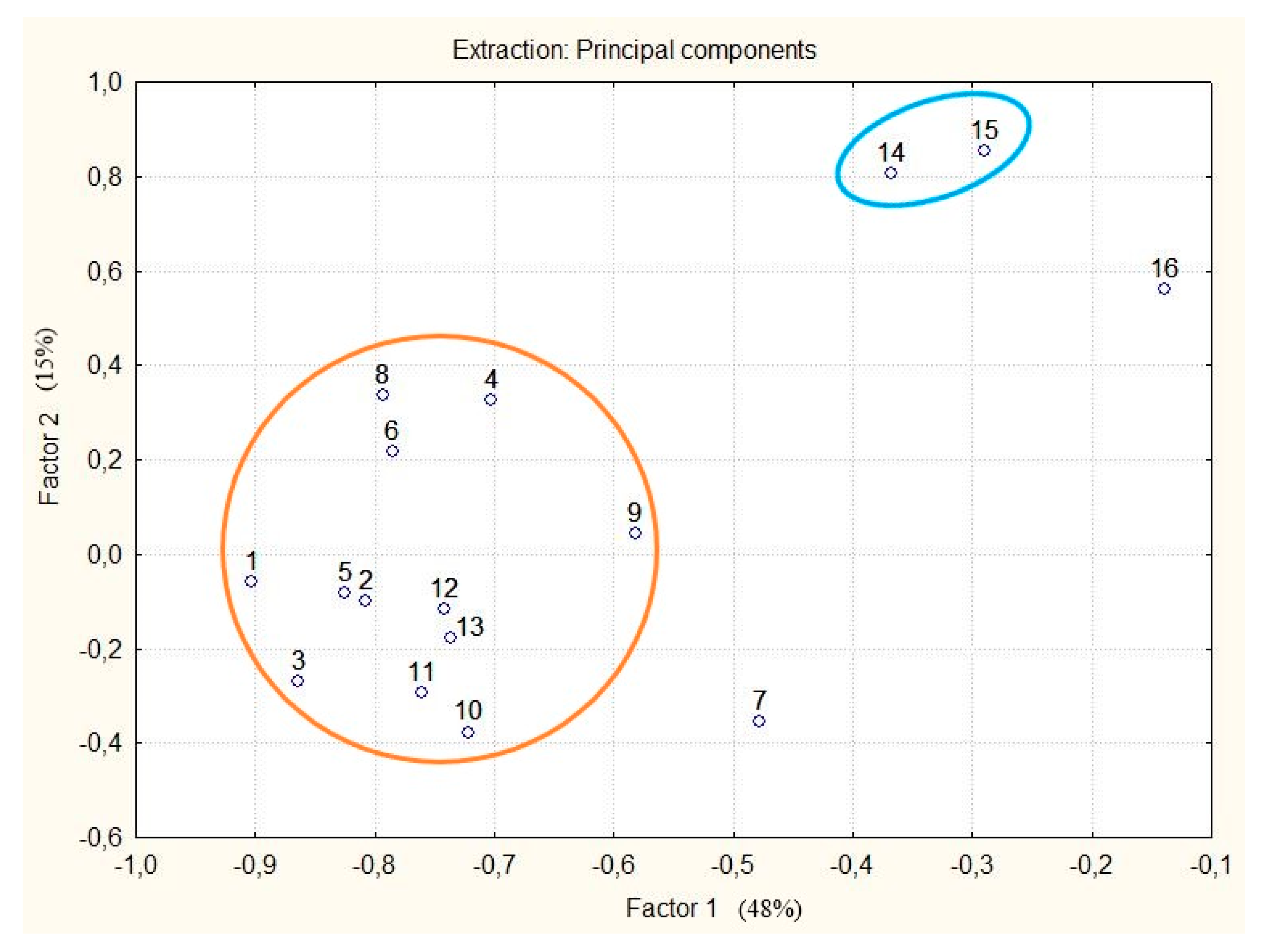
2.5. Biochemical Analysis
2.5.1. The Profile and Levels of Phenolic Compounds in Callus Culture and Seedlings (Control 1)
2.5.2. The Profile and Levels of Phenolic Compounds in Immature Plants (Control 2)
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Establishment of Callus Cultures
4.3. Optimal (Combined) Design
4.4. Effect of MJ Treatment
4.5. Biochemical Analysis
4.5.1. Extraction
4.5.2. HPLC Analysis
4.6. Histochemical Analysis
4.7. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marchev, A.; Haas, C.; Schulz, S. Sage in vitro cultures: A promising tool for the production of bioactive terpenes and phenolic substances. Biotechnol. Lett. 2014, 36, 211–221. [Google Scholar] [CrossRef]
- Grech-Baran, M.; Sykłowska-Baranek, K.; Pietrosiuk, A. Biotechnological approaches to enhance salidroside, rosin and its derivatives production in selected Rhodiola spp. in vitro cultures. Phytochem. Rev. 2015, 14, 657–674. [Google Scholar] [CrossRef] [Green Version]
- Zheleznichenko, T.; Banaev, E.; Asbaganov, S.; Voronkova, M.; Kukushkina, T.; Filippova, E.; Mazurkova, N.; Shishkina, L.; Novikova, T. Nitraria schoberi L. hairy root culture as a source of compounds with antiviral activity against influenza virus subtypes A(H5N1) and A(H3N2). 3 Biotech 2018, 8, 260. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.M.; Chen, H.C.; Wu, C.S.; Wu, P.Y.; Wen, K.C. Rhodiola plants: Chemistry and biological activity. J. Food Drug Anal. 2015, 23, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Marchev, A.S.; Dinkova-Kostova, A.T.; György, Z. Rhodiola rosea L.: From golden root to green cell factories. Phytochem. Rev. 2016, 15, 515–536. [Google Scholar] [CrossRef] [Green Version]
- Khanna, K.; Mishra, K.P.; Ganju, L.; Singh, S.B. Golden root: A wholesome treat of immunity. Biomed. Pharmacother 2017, 87, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Brinckmann, J.A.; Cunningham, A.B.; Harter, D.E.V. Running out of time to smell the roseroots: Reviewing threats and trade in wild Rhodiola rosea L. J. Ethnopharmacol. 2021, 269, 113710. [Google Scholar] [CrossRef] [PubMed]
- György, Z.; Tolonen, A.; Pakonen, M.; Neubauer, P.; Hohtola, A. Enhancement of the production of cinnamyl glycosides in CCA cultures of Rhodiola rosea through biotransformation of cinnamyl alcohol. Plant Sci. 2004, 166, 229–236. [Google Scholar] [CrossRef]
- Ma, L.Q.; Gao, D.Y.; Wang, Y.N.; Wang, H.H.; Zhang, J.X.; Pang, X.B.; Hu, T.S.; Lu, S.Y.; Li, G.F.; Ye, H.C.; et al. Effects of overexpression of endogenous phenylalanine ammonia-lyase (PALrs1) on accumulation of salidroside in Rhodiola sachalinensis. Plant Biol. 2008, 10, 323–333. [Google Scholar] [CrossRef]
- György, Z.; Hohtola, A. Production of cinnamyl glycosides in compact callus aggregate cultures of Rhodiola rosea through biotransformation of cinnamyl alcohol. In Protocols for In Vitro Cultures and Secondary Metabolite Analysis of Aromatic and Medicinal Plants. Methods in Molecular Biology; Jain, S.M., Saxena, P., Eds.; Humana Press: New York, NY, USA, 2009; Volume 547, pp. 305–312. [Google Scholar]
- Martin, J.; Pomahacova, B.; Dusek, J.; Duskova, J. In vitro culture establishment of Schizandra chinensis (Turz.) Baill. and Rhodiola rosea L., two adaptogenic compounds producing plants. J. Phytol. 2010, 2, 80–87. [Google Scholar]
- Furmanowa, M.; Hartwich, M.; Alfermann, A.W. Rosavin as a product of glycosylation by Rhodiola rosea (roseroot) cell cultures. Plant Cell Tissue Organ Cult. 1999, 56, 105–110. [Google Scholar] [CrossRef]
- Grech-Baran, M.; Sykłowska-Baranek, K.; Krajewska-Patan, A. Biotransformation of cinnamyl alcohol to rosavins by non-transformed wild type and hairy root cultures of Rhodiola kirilowii. Biotechnol. Lett. 2014, 36, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, S.; Raghuvanshi, R.; Bhardwaj, P.; Sood, H.; Saxena, S.; Chaurasia, O.P. Influence of light quality on growth, secondary metabolites production and antioxidant activity in callus culture of Rhodiola imbricata Edgew. J. Photochem. Photobiol. B 2018, 183, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Condra, L.W. Reliability Improvement with Design of Experiments; Marcel Dekker AG: New York, NY, USA, 2001. [Google Scholar]
- Niedz, R.P.; Evens, T.J. Design of experiments (DoE)—History, concepts, and relevance to in vitro culture. In Vitro Cell. Dev. Biol. Plant 2016, 52, 547–562. [Google Scholar] [CrossRef]
- Niedz, R.P.; Evens, T.J. The effects of nitrogen and potassium nutrition on the growth of nonembryogenic and embryogenic tissue of sweet orange (Citrus sinensis (L.) Osbeck). BMC Plant Biol. 2008, 8, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butenko, R.G. The Culture of Isolated Tissues and the Physiology of Plant Morphogenesis; Nauka Publications: Moscow, Russia, 1964; 350p. [Google Scholar]
- Ramage, C.M.; Williams, R.R. Inorganic nitrogen requirements during shoot organogenesis in tobacco leaf discs. J. Exp. Bot. 2002, 53, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Cousson, A.; Van Tran Thanh, K. Influence of ionic composition of the culture medium on de novo flower formation in tobacco thin cell layers. Can. J. Bot. 1993, 71, 506–511. [Google Scholar] [CrossRef]
- Poothong, S.; Reed, B.M. Optimizing shoot culture media for Rubus germplasm: The effects of NH4+, NO3−, and total nitrogen. In Vitro Cell. Dev. Biol. Plant 2016, 52, 265–275. [Google Scholar] [CrossRef]
- Niedz, R.P.; Evens, T.J. Regulating plant tissue growth by mineral nutrition. In Vitro Cell. Dev. Biol. Plant 2007, 43, 370–381. [Google Scholar] [CrossRef]
- Sheng, C.Z.; Hu, T.Q.; Bi, H.; Yuan, Y.J.; Jiang, Y. Effects of plant growth substances on induction and culture of callus from Rhodiola quadrifida. Zhongguo Zhong Yao Za Zhi 2005, 30, 1237–1240. [Google Scholar]
- Tasheva, K.; Kosturkova, G. Bulgarian golden root in vitro cultures for micropropagation and reintroduction. Cent. Eur. J. Biol. 2010, 5, 853–863. [Google Scholar] [CrossRef]
- Ghiorghită, G.; Hârtan, M.; Maftei, D. Some considerations regarding the in vitro culture of Rhodiola rosea L. Rom. Biotechnol. Lett. 2011, 16, 5902–5908. [Google Scholar]
- Shi, L.; Wang, C.; Zhou, X.; Zhang, Y.; Liu, Y.; Ma, C. Production of salidroside and tyrosol in cell suspension cultures of Rhodiola crenulata. Plant Cell Tiss. Organ. Cult. 2013, 114, 295–303. [Google Scholar] [CrossRef]
- Li, Y.; Shao, C.H.; Park, S.Y. Production of salidroside and polysaccharides in Rhodiola sachalinensis using airlift bioreactor systems. Acta Physiol. Plant 2014, 36, 2975–2983. [Google Scholar] [CrossRef]
- Kapoor, S.; Sharma, A.; Bhardwaj, P. Enhanced production of phenolic compounds in compact allus Aggregate suspension cultures of Rhodiola imbricata Edgew. Appl. Biochem. Biotechnol. 2019, 187, 817–837. [Google Scholar] [CrossRef]
- Mirmazloum, I.; Kiss, A.; Ladányi, M. Production of cinnamyl alcohol glycosides by biotransformation in roseroot callus cells. Plant Cell Tiss. Organ Cult. 2019, 139, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Rattan, S.; Rattan, S.; Sood, A.; Kumar, P.; Kumar, A.; Kumar, D.; Kumar, D.; Warghat, A.R.; Warghat, A.R. Phenylethanoids, phenylpropanoids, and phenolic acids quantification vis-à-vis gene expression profiling in leaf and root derived callus lines of Rhodiola imbricata (Edgew.). Ind. Crops Prod. 2020, 154, 112708. [Google Scholar] [CrossRef]
- Stepanova, A.; Malunova, M.; Salamaikina, S.; Selimov, R.; Solov’eva, A. Establishment of Rhodiola quadrifida hairy roots and callus culture to produce bioactive compounds. Phyton-Int. J. Exp. Bot. 2021, 90, 543–552. [Google Scholar] [CrossRef]
- Kolewe, M.E.; Gaurav, V.; Roberts, S.C. Pharmaceutically active natural product synthesis and supply via plant cell culture technology. Mol. Pharm. 2008, 5, 243–256. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Gundlach, H.; Müller, M.J.; Kutchan, T.M.; Zenk, M.H. Jasmonic acid is a signal transducer in elicitor-induced plant cell cultures. Proc. Natl. Acad. Sci. USA 1992, 89, 2389–2393. [Google Scholar] [CrossRef] [Green Version]
- Chu, U.C.; Adelberg, J.; Lowe, K. Use of DoE methodology to optimize the regeneration of high-quality, single-copy transgenic Zea mays L. (maize) plants. In Vitro Cell. Dev. Biol. Plant 2019, 55, 678–694. [Google Scholar] [CrossRef] [Green Version]
- Erst, A.A.; Petruk, A.A.; Zibareva, L.N.; Erst, A.S. Morphological, histochemical and biochemical features of cultivated Rhodiola rosea (Altai Mountains ecotype). Contemp. Probl. Ecol. 2021, 14, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Montes, E.O.; Oliver-Salvador, C.; Durán-Figueroa, N. Optimization of direct shoot regeneration using cotyledonary explants and true leaves from lettuce cv. Romaine (Lactuca sativa L.) by surface response methodology. Plant Growth Regul. 2015, 77, 327–334. [Google Scholar] [CrossRef]
- Bagherieh-Najjar, M.B.; Nezamdoost, T. Optimization of shikonin production in Onosma dichroantha callus using response surface methodology. Plant Cell Tiss. Organ Cult. 2016, 126, 399–409. [Google Scholar] [CrossRef]
- Yu, K.W.; Hahn, E.J.; Paek, K.Y. Effects of NH4+: NO3− ratio and ionic strength on adventitious root growth and ginsenoside production in bioreactor culture of Panax ginseng C.A. Meyer. Acta Hortic. 2001, 560, 259–262. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Yi, F.; Cai, M. Effects of amino acids, nitrate, and ammonium on the growth and taxol production in cell cultures of Taxus yunnanensis. Plant Growth Regul. 2003, 41, 265–268. [Google Scholar] [CrossRef]
- Prakash, G.; Srivastava, A.K. Statistical media optimization for cell growth and azadirachtin production in Azadirachta indica (A. Juss) suspension cultures. Process Biochem. 2005, 40, 3795–3800. [Google Scholar] [CrossRef]
- Irshad, M.; Debnath, B.; Mitra, S. Accumulation of anthocyanin in callus cultures of red-pod okra (Abelmoschus esculentus (L.) Hongjiao) in response to light and nitrogen levels. Plant Cell Tiss. Organ Cult. 2018, 134, 29–39. [Google Scholar] [CrossRef]
- Oleszkiewicz, T.; Kruczek, M.; Baranski, R. Repression of carotenoid accumulation by nitrogen and NH4+ supply in carrot callus cells in vitro. Plants 2021, 10, 1813. [Google Scholar] [CrossRef]
- Martinez, M.E.; Jorquera, L.; Poirrier, P.; Díaz, K.; Chamy, R. Effect of the carbon source and plant growth regulators (PGRs) in the induction and maintenance of an in vitro callus culture of Taraxacum officinale (L) weber Ex F.H. Wigg. Agronomy 2021, 11, 1181. [Google Scholar] [CrossRef]
- Small, C.C.; Degenhardt, D. Plant growth regulators for enhancing revegetation success in reclamation: A review. Ecol. Eng. 2018, 118, 43–51. [Google Scholar] [CrossRef]
- Goyal, S.; Ramawat, K.G. Synergistic effect of morphactin on cytokinin-induced production of isoflavonoids in cell cultures of Pueraria tuberosa (Roxb. ex. Willd.) DC. Plant Growth Regul. 2008, 55, 175–181. [Google Scholar] [CrossRef]
- Muraseva, D.S.; Kostikova, V.A. In vitro propagation of Spiraea betulifolia subsp. aemiliana (Rosaceae) and comparative analysis of phenolic compounds of microclones and intact plants. Plant Cell Tiss. Organ Cult. 2021, 144, 493–504. [Google Scholar] [CrossRef]
- Karaaslan, M.; Ozden, M.; Vardin, H.; Yilmaz, F.M. Optimisation of phenolic compound biosynthesis in grape (Bogazkere Cv.) callus culture. Afr. J. Biotechnol. 2013, 12, 3922–3933. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, X.; Mohsin, A.; Yin, Z.; Zhuang, Y.; Zhou, B.; Du, L.; Yin, X.; Liu, N.; Wang, Z.; et al. Embryogenic callus induction, cell suspension culture, and spectrum-effect relationship between antioxidant activity and polyphenols composition of Siraitia grosvenorii cultured cells. Ind. Crops Prod. 2022, 176, 114380. [Google Scholar] [CrossRef]
- Mizukami, H.; Tabira, Y.; Ellis, B.E. Methyl jasmonate-induced rosmarinic acid biosynthesis in Lithospermum erythrorhizon cell suspension cultures. Plant Cell Rep. 1993, 12, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Shabania, L.; Ehsanpoura, A.A.; Asgharib, G.; Emamib, J. Glycyrrhizin production by in vitro cultured Glycyrrhiza glabra elicited by methyl jasmonate and salicylic acid. Russ. J. Plant Physiol. 2009, 56, 621–626. [Google Scholar] [CrossRef]
- Martin, K.P.; Sabovljevic, A.; Madassery, J. High-frequency transgenic plant regeneration and plumbagin production through methyl jasmonate elicitation from hairy roots of Plumbago indica L. J. Crop. Sci. Biotech. 2011, 14, 205–212. [Google Scholar] [CrossRef]
- Ram, M.; Prasad, K.V.; Singh, S.K.; Hada, B.S.; Kumar, S. Influence of salicylic acid and methyl jasmonate elicitation on anthocyanin production in callus cultures of Rosa hybrida L. Plant Cell Tiss. Organ Cult. 2013, 113, 459–467. [Google Scholar] [CrossRef]
- Chodisetti, B.; Rao, K.; Gandi, S.; Giri, A. Gymnemic acid enhancement in the suspension cultures of Gymnema sylvestre by using the signaling molecules–methyl jasmonate and salicylic acid. In Vitro Cell. Dev. Biol. Plant 2015, 51, 88–92. [Google Scholar] [CrossRef]
- Erst, A.A.; Zibareva, L.N.; Filonenko, E.S.; Zheleznichenko, T.V. Influence methyl jasmonate on production of ecdysteroids from hairy roots of Silene linicola C.C.Gmelin. Russ. J. Bioorg. Chem. 2019, 7, 76–82. [Google Scholar] [CrossRef]
- Ho, T.T.; Murthy, H.N.; Park, S.Y. Methyl jasmonate induced oxidative stress and accumulation of secondary metabolites in plant cell and organ cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erst, A.A.; Zibareva, L.N.; Filonenko, E.S. Variation in phytoecdysteroid accumulation in hairy roots of Silene linicola over extended time periods. J. Plant Biochem. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Peschel, W.; Kump, A.; Horváth, A.; Csupor, D. Age and harvest season affect the phenylpropenoid content in cultivated European Rhodiola rosea L. Ind. Crops Prod. 2016, 83, 787–802. [Google Scholar] [CrossRef]
- Kołodziej, B.; Sugier, D. Influence of plant age on the chemical composition of roseroot (Rhodiola rosea L.). Acta Sci. Pol. Hortorum. Cultus 2013, 12, 147–160. [Google Scholar]
- Kurkin, V.A.; Zapesochnaya, G.G.; Dubichev, A.G. Phenylpropanoids of a callus culture of Rhodiola rosea. Chem. Nat. Compd. 1991, 27, 419–425. [Google Scholar] [CrossRef]
- Erst, A.A.; Yakubov, V.V. Regenerative in vitro capacity of rare species Rhodiola rosea L. from various habitats. Contemp. Probl. Ecol. 2019, 12, 368–376. [Google Scholar] [CrossRef]
- Erst, A.; Erst, A.; Shmakov, A. In vitro propagation of rare species Rhodiola rosea from Altai Mountains. Turczaninowia 2018, 21, 78–86. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Niedz, R.P.; Evens, T.J. A solution to the problem of ion confounding in experimental biology. Nat. Methods 2006, 3, 417. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fresh Weight | Dry Weight | |||
|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | |
| Model | 107.14 | <0.0001 | 35.83 | <0.0001 |
| Linear × Linear Mixture | 274.28 | <0.0001 | 80.74 | <0.0001 |
| NH4+ * K+ *BAP | 140.46 | <0.0001 | 31.82 | <0.0001 |
| NH4+ * K+ *NAA | 22.00 | <0.0001 | 13.40 | 0.0008 |
| NH4+ *BAP*NAA | 38.42 | <0.0001 | 6.61 | 0.0142 |
| NH4+ *BAP* NO3− | 5.78 | 0.0218 | 1.01 | 0.3208 |
| NH4+ *BAP*BAP + NAA | 21.64 | <0.0001 | 6.87 | 0.0126 |
| NH4+ * K+ *BAP* NO3− | 19.65 | <0.0001 | - | - |
| NH4+ * K+ *BAP* BAP + NAA | 19.84 | <0.0001 | 2.74 | 0.1058 |
| NH4+ * K+ *NAA* NO3− | 7.92 | 0.0081 | 1.70 | 0.2002 |
| NH4+ * K+ *NAA* BAP + NAA | 15.35 | 0.0004 | 6.63 | 0.0141 |
| NH4+ *BAP*NAA* NO3− | 76.36 | <0.0001 | 17.88 | 0.0001 |
| NH4+ *BAP*NAA* BAP + NAA | 68.45 | <0.0001 | - | - |
| NH4+ *BAP* NO3−* BAP + NAA | 21.51 | <0.0001 | 25.17 | <0.0001 |
| NH4+ * NAA* NO3−* BAP + NAA | 12.34 | 0.0013 | - | - |
| K+ *BAP* [NO3−]2 | 1.84 | 0.1842 | - | - |
| NH4+ * K+ *BAP* NO3−* BAP + NAA | 4.74 | 0.0364 | 7.54 | 0.0092 |
| NH4+ *BAP*NAA* NO3−* BAP + NAA | 23.44 | <0.0001 | 1.83 | 0.1840 |
| NH4+ * K+ *BAP*[ NO3−]2 | 61.34 | <0.0001 | 16.24 | 0.0003 |
| NH4+ * K+ *BAP*[ BAP + NAA]2 | 7.57 | 0.0095 | 3.79 | 0.0590 |
| Lack of Fit | p = 0.8180 | p = 0.6741 | ||
| R2 | 0.9851 | 0.9413 | ||
| Adjusted R2 | 0.9759 | 0.9150 | ||
| Predicted R2 | 0.9235 | 0.8162 | ||
| Adeq Precision | 40.4608 | 23.0849 | ||
| Std. Dev. | 25.21 | 0.0142 | ||
| Mean | 178.00 | 0.1071 | ||
| C.V.% | 14.16 | 13.26 | ||
| Model type | Reduced Quadratic × Quadratic × Quadratic model | Reduced Quadratic × Quadratic × Quadratic model | ||
| Compound | Spectral Characteristics: λmax, nm | Retention Time (tR), min | Content, mg/g of Air-Dried Material | |
|---|---|---|---|---|
| Rhizomes | Roots | |||
| Gallic acid | 272 | 1.8 | 26.15 | 21.20 |
| Compound 2 | 216,280 | 2.9 | 3.20 | 2.30 |
| Compound 3 | 228,296 | 6.5 | 0.51 | 0.23 |
| Compound 4 | 218,274 | 8.5 | 0.41 | - |
| Rosarin | 253 | 10.7 | 16.33 | 7.41 |
| Rosavin | 253 | 12.5 | 41.73 | 11.73 |
| Rosin | 253 | 13.5 | 25.10 | 11.25 |
| Cinnamyl alcohol | 205,253 | 24.3 | 42.74 | 32.10 |
| Rhodiosin | 277,333,385 | 39.9 | 0.81 | 0.72 |
| Rhodionin | 277,333,385 | 40.5 | 2.11 | 1.74 |
| TPC | 159.09 | 88.68 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erst, A.A.; Petruk, A.A.; Erst, A.S.; Krivenko, D.A.; Filinova, N.V.; Maltseva, S.Y.; Kulikovskiy, M.S.; Banaev, E.V. Optimization of Biomass Accumulation and Production of Phenolic Compounds in Callus Cultures of Rhodiola rosea L. Using Design of Experiments. Plants 2022, 11, 124. https://doi.org/10.3390/plants11010124
Erst AA, Petruk AA, Erst AS, Krivenko DA, Filinova NV, Maltseva SY, Kulikovskiy MS, Banaev EV. Optimization of Biomass Accumulation and Production of Phenolic Compounds in Callus Cultures of Rhodiola rosea L. Using Design of Experiments. Plants. 2022; 11(1):124. https://doi.org/10.3390/plants11010124
Chicago/Turabian StyleErst, Anna A., Anastasia A. Petruk, Andrey S. Erst, Denis A. Krivenko, Nadezhda V. Filinova, Svetlana Y. Maltseva, Maxim S. Kulikovskiy, and Evgeny V. Banaev. 2022. "Optimization of Biomass Accumulation and Production of Phenolic Compounds in Callus Cultures of Rhodiola rosea L. Using Design of Experiments" Plants 11, no. 1: 124. https://doi.org/10.3390/plants11010124