
Genomic and Meiotic Changes Accompanying Polyploidization



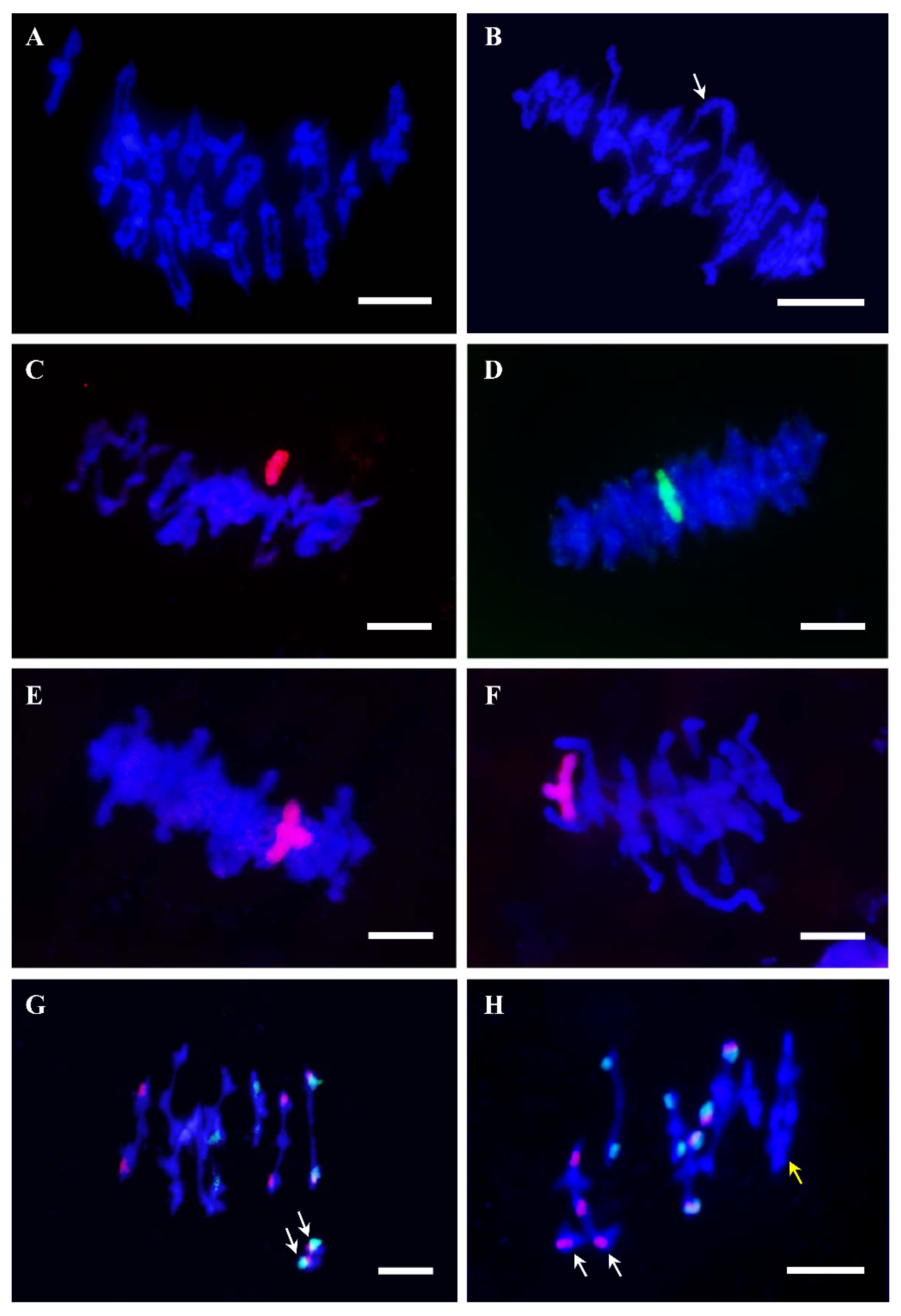{kind=link}
Abstract
:1. Introduction
2. Interspecific Hybridization
3. Mechanisms and Frequency of WGD in Plants
3.1. Pathways for WGD in Plants
3.2. Cellular Mechanisms, and Their Genetic Control, Causing Plants Meiotic Restitution
3.3. Meiotic Restitution in Interspecific Hybrids
3.4. Stress-Induced Meiotic Restitution
3.5. Premeiotic and Postmeiotic WGD in Plants
3.6. Frequency of Unreduced Gametes Formation in Natural Plant Populations
4. Cytological Diploidization of Allopolyploids
4.1. Monocots
4.1.1. Wheat
4.1.2. Wheat-Related Polyploid Species
4.1.3. Other Polyploid Species Included in the Poaceae Family
4.2. Dicots
4.2.1. Brassica napus
4.2.2. Arabidopsis
4.2.3. Solanum tuberosum
5. Readjustments of the Merged Genomes
5.1. Genomic Changes
5.2. Changes in the Activity of Transposable Elements (TEs)
5.3. Changes at the Gene Expression and Regulatory Level
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, W.H. Polyploidy: Biological Relevance; Plenum Press: New York, NY, USA, 1980; pp. 1–583. [Google Scholar] [CrossRef]
- Soltis, D.E.; Visger, C.J.; Soltis, P.S. The Polyploidy Revolution Then...and Now: Stebbins Revisited. Am. J. Bot. 2014, 101, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, J.; Schemske, D.W. Pathways, Mechanisms, and Rates of Polyploid Formation in Flowering Plants. Annu. Rev. Ecol. Syst. 1998, 29, 467–501. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.C. Polyploidy and Diploidy: New Perspectives on Chromosome Pairing and Its Evolutionary Implications. Am. J. Bot. 1982, 69, 1512–1523. [Google Scholar] [CrossRef]
- Ramsey, J.; Schemske, D.W. Neopolyploidy in Flowering Plants. Annu. Rev. Ecol. Syst. 2002, 33, 589–639. [Google Scholar] [CrossRef] [Green Version]
- Soltis, P.S.; Soltis, D.E. The Role of Genetic and Genomic Attributes in the Success of Polyploids. Proc. Natl. Acad. Sci. USA 2000, 97, 7051–7057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltis, D.E.; Soltis, P.S.; Schemske, D.W.; Hancock, J.F.; Thompson, J.N.; Husband, B.C.; Judd, W.S. Autopolyploidy in Angiosperms: Have We Grossly Underestimated the Number of Species? Taxon 2007, 56, 13–30. [Google Scholar] [CrossRef]
- Qiu, T.; Liu, Z.; Liu, B. The Effects of Hybridization and Genome Doubling in Plant Evolution via Allopolyploidy. Mol. Biol. Rep. 2020, 47, 5549–5558. [Google Scholar] [CrossRef]
- Schranz, M.E.; Osborn, T.C. Novel Flowering Time Variation in the Resynthesised Polyploid Brassica Napus. J. Hered. 2000, 91, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Cohen, H.; Fait, A.; Tel-Zur, N. Morphological, Cytological and Metabolic Consequences of Autopolyploidization in Hylocereus (Cactaceae) Species. BMC Plant Biol. 2013, 13, 173. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.; Levy, A.A. Allopolyploidy—A Shaping Force in the Evolution of Wheat Genomes. Cytogenet. Genome Res. 2005, 109, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.L.; Wendel, J.F. Polyploidy and Genome Evolution in Plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef]
- Chen, Z.J.; Ni, Z. Mechanisms of Genomic Rearrangements and Gene Expression Changes in Plant Polyploids. BioEssays 2006, 28, 240–252. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Lu, P.; Tang, K.; Osborn, T.C. Rapid Genome Change in Synthetic Polyploids of Brassica and Its Implications for Polyploid Evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 7719–7723. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, S.D.; Le Mignon, G.; Eber, F.; Coriton, O.; Monod, H.; Clouet, V.; Huteau, V.; Lostanlen, A.; Delourme, R.; Chalhoub, B.; et al. Homeologous Recombination Plays a Major Role in Chromosome Rearrangements That Occur During Meiosis of Brassica Napus Haploids. Genetics 2007, 175, 487–503. [Google Scholar] [CrossRef] [Green Version]
- Gaeta, R.T.; Chris Pires, J. Homoeologous Recombination in Allopolyploids: The Polyploid Ratchet. New Phytol. 2010, 186, 18–28. [Google Scholar] [CrossRef]
- Grandont, L.; Jenczewski, E.; Lloyd, A. Meiosis and Its Deviations in Polyploid Plants. Cytogenet. Genome Res. 2013, 140, 171–184. [Google Scholar] [CrossRef]
- Lipman, M.J.; Chester, M.; Soltis, P.S.; Soltis, D.E. Natural Hybrids between Tragopogon mirus and T. miscellus (Asteraceae): A New Perspective on Karyotypic Changes Following Hybridization at the Polyploid Level. Am. J. Bot. 2013, 100, 2016–2022. [Google Scholar] [CrossRef]
- Shaked, H.; Kashkush, K.; Ozkan, H.; Feldman, M.; Levy, A.A. Sequence Elimination and Cytosine Methylation Are Rapid and Reproducible Responses of the Genome to Wide Hybridization and Allopolyploidy in Wheat. Plant Cell 2001, 13, 1749–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashkush, K.; Feldman, M.; Levy, A.A. Gene Loss, Silencing and Activation in a Newly Synthesized Wheat Allotetraploid. Genetics 2002, 160, 1651–1659. [Google Scholar] [CrossRef] [PubMed]
- Madlung, A.; Masuelli, R.W.; Watson, B.; Reynolds, S.H.; Davison, J.; Comai, L. Remodeling of DNA Methylation and Phenotypic and Transcriptional Changes in Synthetic Arabidopsis Allotetraploids. Plant Physiol. 2002, 129, 733–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madlung, A.; Tyagi, A.P.; Watson, B.; Jiang, H.; Kagochi, T.; Doerge, R.W.; Martienssen, R.; Comai, L. Genomic Changes in Synthetic Arabidopsis Polyploids. Plant J. 2005, 41, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J. Genetic and Epigenetic Mechanisms for Gene Expression and Phenotypic Variation in Plant Polyploids. Annu. Rev. Plant Biol. 2007, 58, 377–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaeta, R.T.; Pires, J.C.; Iniguez-Luy, F.; Leon, E.; Osborn, T.C. Genomic Changes in Resynthesized Brassica Napus and Their Effect on Gene Expression and Phenotype. Plant Cell 2007, 19, 3403–3417. [Google Scholar] [CrossRef] [Green Version]
- Adams, K.L.; Wendel, J.F. Novel Patterns of Gene Expression in Polyploid Plants. Trends Genet. 2005, 21, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Comai, L.; Tyagi, A.P.; Winter, K.; Holmes-Davis, R.; Reynolds, S.H.; Stevens, Y.; Byers, B. Phenotypic Instability and Rapid Gene Silencing in Newly Formed Arabidopsis Allotetraploids. Plant Cell 2000, 12, 1551–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tian, L.; Madlung, A.; Lee, H.S.; Chen, M.; Lee, J.J.; Watson, B.; Kagochi, T.; Comai, L.; Chen, Z.J. Stochastic and Epigenetic Changes of Gene Expression in Arabidopsis Polyploids. Genetics 2004, 167, 1961–1973. [Google Scholar] [CrossRef] [Green Version]
- Stupar, R.M.; Bhaskar, P.B.; Yandell, B.S.; Rensink, W.A.; Hart, A.L.; Ouyang, S.; Veilleux, R.E.; Busse, J.S.; Erhardt, R.J.; Buell, C.R.; et al. Phenotypic and Transcriptomic Changes Associated With Potato Autopolyploidization. Genetics 2007, 176, 2055–2067. [Google Scholar] [CrossRef] [Green Version]
- Mayrose, I.; Zhan, S.H.; Rothfels, C.J.; Magnusn-Ford, K.; Barker, M.S.; Rieseberg, L.H.; Otto, S.P. Recently Formed Polyploid Plants Diversify at Lower Rates. Science 2011, 333, 1257. [Google Scholar] [CrossRef] [Green Version]
- Sémon, M.; Wolfe, K.H. Consequences of Genome Duplication. Curr. Opin. Genet. Dev. 2007, 17, 505–512. [Google Scholar] [CrossRef]
- Tate, J.A.; Joshi, P.; Soltis, K.A.; Soltis, P.S.; Soltis, D.E. On the Road to Diploidization? Homoeolog Loss in Independently Formed Populations of the Allopolyploid Tragopogon miscellus (Asteraceae). BMC Plant Biol. 2009, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.G.; Huang, S.Z.; Pin, A.L.; Adams, K.L. Extensive Divergence in Alternative Splicing Patterns after Gene and Genome Duplication during the Evolutionary History of Arabidopsis. Mol. Biol. Evol. 2010, 27, 1686–1697. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Lin, S.Y.; Chi, W.T.; Charng, Y.Y. Recent Gene Duplication and Subfunctionalization Produced a Mitochondrial GrpE, the Nucleotide Exchange Factor of the Hsp70 Complex, Specialized in Thermotolerance to Chronic Heat Stress in Arabidopsis. Plant Physiol. 2012, 158, 747–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of Duplicate Genes in Exon-Intron Structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Smet, R.; Adams, K.L.; Vandepoele, K.; Van Montagu, M.C.E.; Maere, S.; Van De Peer, Y. Convergent Gene Loss Following Gene and Genome Duplications Creates Single-Copy Families in Flowering Plants. Proc. Natl. Acad. Sci. USA 2013, 110, 2898–2903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roulin, A.; Auer, P.L.; Libault, M.; Schlueter, J.; Farmer, A.; May, G.; Stacey, G.; Doerge, R.W.; Jackson, S.A. The Fate of Duplicated Genes in a Polyploid Plant Genome. Plant J. 2013, 73, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Koh, J.; Soltis, P.S.; Soltis, D.E. Homeolog Loss and Expression Changes in Natural Populations of the Recently and Repeatedly Formed Allotetraploid Tragopogon mirus (Asteraceae). BMC Genome 2010, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, T.; Benavente, E. The Mode and Regulation of Chromosome Pairing in Wheat-Alien Hybrids (Ph Genes, an Updated View). In Alien Introgression in Wheat: Cytogenetics, Molecular Biology, and Genomics, 1st ed.; Molnár-Láng, M., Ceoloni, C., Doležel, J., Eds.; Springer: Cham, Switzerland, 2015; pp. 133–162. [Google Scholar] [CrossRef]
- Svačina, R.; Sourdille, P.; Kopecký, D.; Bartoš, J. Chromosome Pairing in Polyploid Grasses. Front. Plant Sci. 2020, 11, 1056. [Google Scholar] [CrossRef]
- Jiao, Y.; Wickett, N.J.; Ayyampalayam, S.; Chanderbali, A.S.; Landherr, L.; Ralph, P.E.; Tomsho, L.P.; Hu, Y.; Liang, H.; Soltis, P.S.; et al. Ancestral Polyploidy in Seed Plants and Angiosperms. Nature 2011, 473, 97–100. [Google Scholar] [CrossRef]
- Paterson, A.H.; Wang, X.; Li, J.; Tang, H. Ancient and Recent Polyploidy in Monocots. In Polyploidy Genome Evolution; Soltis, P.S., Soltis, D.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 93–108. [Google Scholar] [CrossRef]
- Wendel, J.F. The Wondrous Cycles of Polyploidy in Plants. Am. J. Bot. 2015, 102, 1753–1756. [Google Scholar] [CrossRef] [Green Version]
- De Storme, N.; Mason, A. Plant Speciation through Chromosome Instability and Ploidy Change: Cellular Mechanisms, Molecular Factors and Evolutionary Relevance. Curr. Plant Biol. 2014, 1, 10–33. [Google Scholar] [CrossRef] [Green Version]
- Robert, H. Plant Hybridization before Mendel; Princeton University Press: Princeton, NJ, USA; Elsevier: Princeton, NJ, USA, 1929. [Google Scholar]
- Zirkle, C. The Beginnings of Plant Hybridization. Morris Arboretum Monographs. I; University of Pennsylvania Press, Philadelphia (Humphrey Milford, Oxford University Press): London, UK, 1935; pp. 1–248. [Google Scholar]
- López-Caamal, A.; Tovar-Sánchez, E. Genetic, Morphologica, and Chemical Patterns of Plant Hybridization. Rev. Chil. Hist. Nat. 2014, 87, 16. [Google Scholar] [CrossRef] [Green Version]
- Mendel, G. Versuche Über Pflanzen-Hybriden. In Verhandlungen des Naturforschenden Vereines in Brünn; Verlage des Vereines: Brünn, Czechoslovakia, 1866; pp. 3–47. [Google Scholar]
- De Vries, H. Das Spaltungsgesetz Der Bastarde. Ber. Der Dtsch. Bot. Ges. 1900, 18, 83–90. [Google Scholar] [CrossRef]
- Correns, C. Mendel’s Regel Über Das Verhalten Der Nachkommenschaft Der Rassenbastarde. Ber. Der Dtsch. Bot. Ges. 1900, 18, 158–168. [Google Scholar]
- Tschermak, E. Ueber Künstliche Kreuzung Bei Pisum Sativum. Ber. Der Dtsch. Bot. Ges. 1900, 18, 232–239. [Google Scholar] [CrossRef]
- Lotsy, J.P. Evolution by Means of Hybridization; Martinus Nijhoff: The Hague, The Netherlands, 1916; p. 166. [Google Scholar]
- Winge, Ø. The Chromosomes, Their Numbers and General Importance. C.R. Trav. Lab. Carlsb. 1917, 13, 131–275. [Google Scholar]
- Stebbins, G.L. Variation and Evolution in Plants; Columbia University Press: New York, NY, USA, 1950; pp. 1–643. [Google Scholar] [CrossRef]
- Soltis, D.E.; Soltis, P.S. Molecular Data and the Dynamic Nature of Polyploidy. CRC Crit. Rev. Plant Sci. 1993, 12, 243–273. [Google Scholar] [CrossRef]
- Müntzing, A. Outlines to a Genetic Monograph of the Genus Galeopsis. Hereditas 1930, 13, 185–341. [Google Scholar] [CrossRef]
- Rieseberg, L.H. Hybrid Origins of Plant Species. Annu. Rev. Ecol. Syst. 2003, 28, 359–389. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D. Evolution of Plant Breeding Systems. Curr. Biol. 2006, 16, R726–R735. [Google Scholar] [CrossRef] [Green Version]
- Ollerton, J.; Winfree, R.; Tarrant, S. How Many Flowering Plants Are Pollinated by Animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Aide, T.M. The Influence of Wind and Animal Pollination on Variation in Outcrossing Rates. Evolution 1986, 40, 434–435. [Google Scholar] [CrossRef] [Green Version]
- Tiffin, P.; Olson, M.S.; Moyle, L.C. Asymmetrical Crossing Barriers in Angiosperms. Proc. Biol. Sci. 2001, 268, 861–867. [Google Scholar] [CrossRef] [Green Version]
- Moreira-Hernández, J.I.; Muchhala, N. Importance of Pollinator-Mediated Interspecific Pollen Transfer for Angiosperm Evolution. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 191–217. [Google Scholar] [CrossRef]
- Yakimowski, S.B.; Rieseberg, L.H. The Role of Homoploid Hybridization in Evolution: A Century of Studies Synthesizing Genetics and Ecology. Am. J. Bot. 2014, 101, 1247–1258. [Google Scholar] [CrossRef]
- Bedinger, P.A.; Broz, A.K.; Tovar-Mendez, A.; McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiol. 2017, 173, 79–90. [Google Scholar] [CrossRef]
- Whitney, K.D.; Ahern, J.R.; Campbell, L.G.; Albert, L.P.; Kin, M.S. Patterns of Hybridization in Plants. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 175–182. [Google Scholar] [CrossRef]
- Ellstrand, N. Dangerous Liaisons? When Cultivated Plants Mate with Their Wild Relatives; Johns Hopkins University Press: Baltimore, MD, USA, 2003; p. 244. [Google Scholar]
- Darlington, C.D. Recent Advances in Cytology, 2nd ed.; P. Blakiston’s Son & Co.: Philadelphia, PA, USA, 1937; pp. 1–671. [Google Scholar]
- Buggs, R.J.A.; Soltis, P.S.; Soltis, D.E. Does Hybridization between Divergent Progenitors Drive Whole-Genome Duplication? Mol. Ecol. 2009, 18, 3334–3339. [Google Scholar] [CrossRef] [Green Version]
- Soltis, D.E.; Buggs, R.J.A.; Doyle, J.J.; Soltis, P.S. What We Still Don’t Know about Polyploidy. Taxon 2010, 59, 1387–1403. [Google Scholar] [CrossRef]
- de Wet, J.M. Origins of Polyploids. Polyploidy Biol. Revel. 1979, 13, 3–15. [Google Scholar] [CrossRef]
- Harlan, J.R.; de Wet, J.M.J. On Ö. Winge and a Prayer: The Origins of Polyploidy. Bot. Rev. 1975, 41, 361–390. [Google Scholar] [CrossRef]
- Bretagnolle, F.; Thompson, J.D. Gametes with the Somatic Chromosome Number: Mechanisms of Their Formation and Role in the Evolution of Autopolyploid Plants. New Phytol. 1995, 129, 1–22. [Google Scholar] [CrossRef]
- Brownfield, L.; Köhler, C. Unreduced Gamete Formation in Plants: Mechanisms and Prospects. J. Exp. Bot. 2011, 62, 1659–1668. [Google Scholar] [CrossRef] [Green Version]
- de Storme, N.; Geelen, D. Sexual Polyploidization in Plants—Cytological Mechanisms and Molecular Regulation. New Phytol. 2013, 198, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Shamina, N.; Dorogova, N.; Goncharov, N.; Orlova, A.; Trunova, S. Abnormalities of Spindle and Cytokine Behavior Leading to the Formation of Meiotic Restitution Nuclei in Intergeneric Cereal Hybrids. Cell Biol. Int. 1999, 23, 863–870. [Google Scholar] [CrossRef]
- Shamina, N.V.; Silkova, O.G.; Seriukova, E.G. Monopolar Spindles in Meiosis of Intergeneric Cereal Hybrids. Cell Biol. Int. 2003, 27, 657–664. [Google Scholar] [CrossRef]
- Shamina, N.; Dorogova, N.; Trunova, S. Radial Spindle and the Phenotype of the Maize Meiotic Mutant, Dv. Cell Biol. Int. 2000, 24, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Conicella, C.; Genualdo, G.; Errico, A.; Frusciante, L.; Monti, L.M. Meiotic Restitution Mechanisms and 2n Pollen Formation in a Solanum tuberosum Dihaploid and in Dihaploid x Wild Species Hybrids. Plant Breed. 1996, 115, 157–161. [Google Scholar] [CrossRef]
- Conicella, C.; Capo, A.; Cammareri, M.; Errico, A.; Shamina, N.; Monti, L.M. Elucidation of Meiotic Nuclear Restitution Mechanisms in Potato through Analysis of Microtubular Cytoskeleton. Euphytica 2003, 133, 107–115. [Google Scholar] [CrossRef]
- D’Erfurth, I.; Jolivet, S.; Froger, N.; Catrice, O.; Novatchkova, M.; Simon, M.; Jenczewski, E.; Mercier, R. Mutations in AtPS1 (Arabidopsis thaliana Parallel Spindle 1) Lead to the Production of Diploid Pollen Grains. PLoS Genet. 2008, 4, e1000274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Storme, N.; Geelen, D. The Arabidopsis Mutant jason Produces Unreduced First Division Restitution Male Gametes through a Parallel/Fused Spindle Mechanism in Meiosis II. Plant Physiol. 2011, 155, 1403–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra Lopez-Lavalle, L.A.; Orjeda, G. Occurrence and Cytological Mechanism of 2n Pollen Formation in a Tetraploid Accession of Ipomoea batatas (Sweet Potato). J. Hered. 2002, 93, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.F.; Wei, Z.Z.; Li, D.; Li, B. Using SSR Markers to Study the Mechanism of 2n Pollen Formation in Populus × Euramericana (Dode) Guinier and P. popularis. Ann. For. Sci. 2009, 66, 506. [Google Scholar] [CrossRef]
- Liu, B.; Jin, C.; De Storme, N.; Schotte, S.; Schindfessel, C.; De Meyer, T.; Geelen, D. A Hypomorphic Mutant of PHD Domain Protein Male Meiocytes Death 1. Genes 2021, 12, 516. [Google Scholar] [CrossRef]
- Li, Y.; Shen, Y.; Cai, C.; Zhong, C.; Zhu, L.; Yuan, M.; Ren, H. The Type II Arabidopsis Formin14 Interacts with Microtubules and Microfilaments to Regulate Cell Division. Plant Cell 2010, 22, 2710–2726. [Google Scholar] [CrossRef] [Green Version]
- Iwanaga, M.; Peloquin, S.J. Origin and Evolution of Cultivated Tetraploid Potatoes via 2n Gametes. Theor. Appl. Genet. 1982, 61, 161–169. [Google Scholar] [CrossRef]
- Watanabe, K.; Peloquin, S.J. Occurrence of 2n Pollen and Ps Gene Frequencies in Cultivated Groups and Their Related Wild Species in Tuber-Bearing Solanums. Theor. Appl. Genet. 1989, 78, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Mo, X.; Gui, M.; Wu, X.; Jiang, Y.; Ma, L.; Shi, Z.; Luo, Y.; Tang, W. Cytological, Molecular Mechanisms and Temperature Stress Regulating Production of Diploid Male Gametes in Dianthus caryophyllus L. Plant Physiol. Biochem. 2015, 97, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, T.W.; Bingham, E.T. Abnormal Meiosis in Alfalfa, Medicago sativa: Cytology of 2N Egg and 4N Pollen Formation. Can. J. Genet. Cytol. 1983, 25, 107–112. [Google Scholar] [CrossRef]
- Pagliarini, M.S.; Takayama, S.Y.; De Freitas, P.M.; Carraro, L.R.; Adamowski, E.V.; Silva, N.; Batista, L.A.R. Failure of Cytokinesis and 2n Gamete Formation in Brazilian Accessions of Paspalum. Euphytica 1999, 108, 129–135. [Google Scholar] [CrossRef]
- Boldrini, K.R.; Pagliarini, M.S.; Do Valle, C.B. Abnormal Timing of Cytokinesis in Microsporogenesis in Brachiaria humidicola (Poaceae: Paniceae). J. Genet. 2006, 85, 225–228. [Google Scholar] [CrossRef] [PubMed]
- De Storme, N.; Copenhaver, G.P.; Geelen, D. Production of Diploid Male Gametes in Arabidopsis by Cold-Induced Destabilization of Postmeiotic Radial Microtubule Arrays. Plant Physiol. 2012, 160, 1808–1826. [Google Scholar] [CrossRef] [Green Version]
- Premachandran, M.N.; Lalitha, R. Microsporogenesis in a Fertile Saccharum Officinarum × Erianthus Arundinaceus Hybrid with Floral Abnormalities. J. Sugarcane Res. 2012, 2, 11–17. [Google Scholar]
- Gómez-Rodríguez, V.M.; Rodríguez-Garay, B.; Barba-Gonzalez, R. Meiotic Restitution Mechanisms Involved in the Formation of 2n Pollen in Agave tequilana Weber and Agave angustifolia Haw. Springerplus 2012, 1, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.Y.; Spielman, M.; Coles, J.P.; Li, Y.; Ghelani, S.; Bourdon, V.; Brown, R.C.; Lemmon, B.E.; Scott, R.J.; Dickinson, H.G. TETRASPORE Encodes a Kinesin Required for Male Meiotic Cytokinesis in Arabidopsis. Plant J. 2003, 34, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Chen, J.G.; Ellis, B.E. AtMPK4 Is Required for Male-Specific Meiotic Cytokinesis in Arabidopsis. Plant J. 2011, 67, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Krupnova, T.; Stierhof, Y.D.; Hiller, U.; Strompen, G.; Müller, S. The Microtubule-Associated Kinase-like Protein RUNKEL Functions in Somatic and Syncytial Cytokinesis. Plant J. 2013, 74, 781–791. [Google Scholar] [CrossRef]
- Oh, S.A.; Bourdon, V.; Dickinson, H.G.; Twell, D.; Park, S.K. Arabidopsis Fused Kinase TWO-IN-ONE Dominantly Inhibits Male Meiotic Cytokinesis. Plant Reprod. 2014, 27, 7–17. [Google Scholar] [CrossRef]
- Shamina, N.V.; Shatskaya, O.A. Two Novel Meiotic Restitution Mechanisms in Haploid Maize (Zea Mays L.). Russ. J. Genet. 2011, 47, 438–445. [Google Scholar] [CrossRef]
- Enns, L.C.; Kanaoka, M.M.; Torii, K.U.; Comai, L.; Okada, K.; Cleland, R.E. Two Callose Synthases, GSL1 and GSL5, Play an Essential and Redundant Role in Plant and Pollen Development and in Fertility. Plant Mol. Biol. 2005, 58, 333–349. [Google Scholar] [CrossRef]
- Zhang, Z.; Kang, X. Cytological Characteristics of Numerically Unreduced Pollen Production in Populus tomentosa Carr. Euphytica 2010, 173, 151–159. [Google Scholar] [CrossRef]
- Tavoletti, S. Cytological Mechanisms of 2n Egg Formation in a Diploid Genotype of Medicago sativa Subsp. Falcata. Euphytica 1994, 75, 1–8. [Google Scholar] [CrossRef]
- Ravi, M.; Marimuthu, M.P.A.; Siddiqi, I. Gamete Formation without Meiosis in Arabidopsis. Nature 2008, 451, 1121–1124. [Google Scholar] [CrossRef]
- D’Erfurth, I.; Jolivet, S.; Froger, N.; Catrice, O.; Novatchkova, M.; Mercier, R. Turning Meiosis into Mitosis. PLoS Biol. 2009, 7, e1000124. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Goel, S.; Meeley, R.B.; Dantec, C.; Parrinello, H.; Michaud, C.; Leblanc, O.; Grimanelli, D. Production of Viable Gametes without Meiosis in Maize Deficient for an ARGONAUTE Protein. Plant Cell 2011, 23, 443–458. [Google Scholar] [CrossRef] [Green Version]
- Grimanelli, D.; Leblanc, O.; Espinosa, E.; Perotti, E.; González De León, D.; Savidan, Y. Mapping Diplosporous Apomixis in Tetraploid Tripsacum: One Gene or Several Genes? Heredity 1998, 80, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, P.J.; Bakx-Schotman, J.M.T. Formation of Unreduced Megaspores (Diplospory) in Apomictic Dandelions (Taraxacum officinale, s.l.) is Controlled by a Sex-Specific Dominant Locus. Genetics 2004, 166, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Noyes, R.D. Inheritance of Apomeiosis (Diplospory) in Fleabanes (Erigeron, Asteraceae). Heredity 2004, 94, 193–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozias-Akins, P. Apomixis: Developmental Characteristics and Genetics. CRC Crit. Rev. Plant Sci. 2007, 25, 199–214. [Google Scholar] [CrossRef]
- Kaushal, P.; Dwivedi, K.K.; Radhakrishna, A.; Srivastava, M.K.; Kumar, V.; Roy, A.K.; Malaviya, D.R. Partitioning Apomixis Components to Understand and Utilize Gametophytic Apomixis. Front. Plant Sci. 2019, 10, 256. [Google Scholar] [CrossRef] [PubMed]
- Agashe, B.; Prasad, C.K.; Siddiqi, I. Identification and Analysis of DYAD: A Gene Required for Meiotic Chromosome Organisation and Female Meiotic Progression in Arabidopsis. Development 2002, 129, 3935–3943. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Aguilar, M.; Michaud, C.; Leblanc, O.; Grimanelli, D. Inactivation of a DNA Methylation Pathway in Maize Reproductive Organs Results in Apomixis-like Phenotypes. Plant Cell 2010, 22, 3249–3267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corral, J.M.; Vogel, H.; Aliyu, O.M.; Hensel, G.; Thiel, T.; Kumlehn, J.; Sharbel, T.F. A Conserved Apomixis-Specific Polymorphism Is Correlated with Exclusive Exonuclease Expression in Premeiotic Ovules of Apomictic Boechera Species. Plant Physiol. 2013, 163, 1660–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- d’Erfurth, I.; Cromer, L.; Jolivet, S.; Girard, C.; Horlow, C.; Sun, Y.; To, J.P.C.; Berchowitz, L.E.; Copenhaver, G.P.; Mercier, R. The CYCLIN-A CYCA1;2/TAM Is Required for the Meiosis I to Meiosis II Transition and Cooperates with OSD1 for the Prophase to First Meiotic Division Transition. PLoS Genet. 2010, 6, e1000989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cromer, L.; Heyman, J.; Touati, S.; Harashima, H.; Araou, E.; Girard, C.; Horlow, C.; Wassmann, K.; Schnittger, A.; de Veylder, L.; et al. OSD1 Promotes Meiotic Progression via APC/C Inhibition and Forms a Regulatory Network with TDM and CYCA1;2/TAM. PLoS Genet. 2012, 8, e1002865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulankova, P.; Riehs-Kearnan, N.; Nowack, M.K.; Schnittger, A.; Riha, K. Meiotic Progression in Arabidopsis Is Governed by Complex Regulatory Interactions between SMG7, TDM1, and the Meiosis I–Specific Cyclin TAM. Plant Cell 2010, 22, 3791–3803. [Google Scholar] [CrossRef] [Green Version]
- Kihara, H.; Lilienfeld, F. A New Synthesized 6x-wheat. Hereditas 1949, 35, 307–319. [Google Scholar] [CrossRef]
- Wagenaar, E.B. Meiotic Restitution and the Origin of Polyploidy. i. Influence of Genotype on Polyploid Seedset in a Triticum crassum × T. turgidum Hybrid. Can. J. Genet. Cytol. 1968, 10, 836–843. [Google Scholar] [CrossRef]
- Jauhar, P.P.; Dogramaci-Altuntepe, M.; Peterson, T.S.; Almouslem, A.B. Seedset on Synthetic Haploids of Durum Wheat: Cytological and Molecular Investigations. Crop Sci. 2000, 40, 1742–1749. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Nasuda, S. Durum Wheat as a Candidate for the Unknown Female Progenitor of Bread Wheat: An Empirical Study with a Highly Fertile F1 Hybrid with Aegilops tauschii Coss. Theor. Appl. Genet. 2004, 109, 1710–1717. [Google Scholar] [CrossRef]
- Tiwari, V.K.; Rawat, N.; Neelam, K.; Randhawa, G.S.; Singh, K.; Chhuneja, P.; Dhaliwal, H.S. Development of Triticum turgidum Subsp. Durum—Aegilops longissima Amphiploids with High Iron and Zinc Content through Unreduced Gamete Formation in F1 Hybrids. Genome 2008, 51, 757–766. [Google Scholar] [CrossRef]
- You-wei, Y.; Zhang, L.; Yen, Y.; Zheng, Y.; Liu, D. Cytological Evidence on Meiotic Restitution in Pentaploid F1 Hybrids between Synthetic Hexaploid Wheat and Aegilops variabilis. Caryologia 2010, 63, 354–358. [Google Scholar] [CrossRef]
- Matsuoka, Y. Evolution of Polyploid Triticum Wheats under Cultivation: The Role of Domestication, Natural Hybridization and Allopolyploid Speciation in Their Diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, T.; Lacadena, J.R. Wheat Univalent Orientation at Anaphase I in Wheat-Rye Derivatives. Chromosoma 1982, 84, 653–661. [Google Scholar] [CrossRef]
- Cai, X.; Xu, S.S.; Zhu, X. Mechanism of Haploidy-Dependent Unreductional Meiotic Cell Division in Polyploid Wheat. Chromosoma 2010, 119, 275–285. [Google Scholar] [CrossRef]
- Pignone, D. Non-Reductional Meiosis in a Triticum turgidum ×Aegilops longissima Hybrid and in Backcrosses of Its Amphidiploid with T. turgidum (Poaceae). Plant Syst. Evol. 1993, 187, 127–134. [Google Scholar] [CrossRef]
- Silkova, O.G.; Shchapova, A.I.; Shumny, V.K. Meiotic Restitution in Amphihaploids in the Tribe Triticeae. Russ. J. Genet. 2011, 47, 383–393. [Google Scholar] [CrossRef]
- Silkova, O.G.; Loginova, D.B. Sister Chromatid Separation and Monopolar Spindle Organization in the First Meiosis as Two Mechanisms of Unreduced Gametes Formation in Wheat-Rye Hybrids. Plant Reprod. 2016, 29, 199–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, A.V.; Ortiz, A.M.; Silvestri, M.C.; Custodio, A.R.; Moretzsohn, M.C.; Lavia, G.I. Occurrence of 2n Microspore Production in Diploid Interspecific Hybrids between the Wild Parental Species of Peanut (Arachis hypogaea L., Leguminosae) and Its Relevance in the Genetic Origin of the Cultigen. Crop Sci. 2020, 60, 2420–2436. [Google Scholar] [CrossRef]
- Wang, C.J.; Zhang, L.Q.; Dai, S.F.; Zheng, Y.L.; Zhang, H.G.; Liu, D.C. Formation of Unreduced Gametes Is Impeded by Homologous Chromosome Pairing in Tetraploid Triticum turgidum × Aegilops tauschii Hybrids. Euphytica 2010, 175, 323–329. [Google Scholar] [CrossRef]
- Ressurreição, F.; Barão, A.; Viegas, W.; Delgado, M. Haploid Independent Unreductional Meiosis in Hexaploid Wheat. In Meiosis Molecular Mechanism Cytogenetics Diversity; Swan, A., Ed.; InTechOpen: London, UK, 2012; pp. 321–330. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Dong, Y. Fertility and Meiotic Mechanisms of Hybrids between Chromosome Autoduplication Tetraploid Wheats and Aegilops Species. Genome 1992, 35, 379–384. [Google Scholar] [CrossRef]
- Zhang, L.Q.; Yen, Y.; Zheng, Y.L.; Liu, D.C. Meiotic Restriction in Emmer Wheat Is Controlled by One or More Nuclear Genes That Continue to Function in Derived Lines. Sex. Plant Reprod. 2007, 20, 159–166. [Google Scholar] [CrossRef]
- Zhang, L.Q.; Liu, D.C.; Zheng, Y.L.; Yan, Z.H.; Dai, S.F.; Li, Y.F.; Jiang, Q.; Ye, Y.Q.; Yen, Y. Frequent Occurrence of Unreduced Gametes in Triticum turgidum-Aegilops tauschii Hybrids. Euphytica 2010, 172, 285–294. [Google Scholar] [CrossRef]
- Tanaka, M. Newly Synthesized Amphidiploids from the Hybrids Emmer Wheats x Aegilops squarrosa Varieties. Wheat Inf. Serv. 1961, 8, 8. [Google Scholar]
- Xu, S.J.; Joppa, L.R. Mechanisms and Inheritance of First Division Restitution in Hybrids of Wheat, Rye, and Aegilops squarrosa. Genome 1995, 38, 607–615. [Google Scholar] [CrossRef]
- Xu, S.J.; Joppa, L.R. First-Division Restitution in Hybrids of Langdon Durum Disomic Substitution Lines with Rye and Aegilops squarrosa. Plant Breed. 2000, 119, 233–241. [Google Scholar] [CrossRef]
- Hao, M.; Luo, J.; Zeng, D.; Zhang, L.; Ning, S.; Yuan, Z.; Yan, Z.; Zhang, H.; Zheng, Y.; Feuillet, C.; et al. QTug.Sau-3B Is a Major Quantitative Trait Locus for Wheat Hexaploidization. Genes Genome Genet. 2014, 4, 1943–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, Y.; Nasuda, S.; Ashida, Y.; Nitta, M.; Tsujimoto, H.; Takumi, S.; Kawahara, T. Genetic Basis for Spontaneous Hybrid Genome Doubling during Allopolyploid Speciation of Common Wheat Shown by Natural Variation Analyses of the Paternal Species. PLoS ONE 2013, 8, e68310. [Google Scholar] [CrossRef] [Green Version]
- Mirzaghaderi, G.; Fathi, N. Unreduced Gamete Formation in Wheat: Aegilops triuncialis Interspecific Hybrids Leads to Spontaneous Complete and Partial Amphiploids. Euphytica 2015, 206, 67–75. [Google Scholar] [CrossRef]
- Silkova, O.G.; Dobrovolskaya, O.B.; Shchapova, A.I.; Shumny, V.K. Features of the Regulation of Meiotic Restitution in Androgenic Haploids of Wheat-Rye Substitution Lines 2R(2D)1, 2R(2D)3, and 6R(6A) (Triticum aestivum L., Cultivar Saratovskaya 29/Secale cereale L., Cultivar Onokhoiskaya). Russ. J. Genet. 2009, 45, 1062–1066. [Google Scholar] [CrossRef]
- Oleszczuk, S.; Lukaszewski, A.J. The Origin of Unusual Chromosome Constitutions among Newly Formed Allopolyploids. Am. J. Bot. 2014, 101, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.B.; Ramanna, M.S.; De Jong, J.H.; Jacobsen, E.; Van Tuyl, J.M. Indeterminate Meiotic Restitution (IMR): A Novel Type of Meiotic Nuclear Restitution Mechanism Detected in Interspecific Lily Hybrids by GISH. Theor. Appl. Genet. 2001, 103, 219–230. [Google Scholar] [CrossRef]
- Stebbins, G.L. Types of Polyploids: Their Classification and Significance. Adv. Genet. 1947, 1, 403–429. [Google Scholar] [CrossRef] [PubMed]
- Gaut, B.S.; Doebley, J.F. DNA Sequence Evidence for the Segmental Allotetraploid Origin of Maize. Proc. Natl. Acad. Sci. USA 1997, 94, 6809–6814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boff, T.; Schifino-Wittmann, M.T. Segmental Allopolyploidy and Paleopolyploidy in Species of Leucaena Benth: Evidence from Meiotic Behaviour Analysis. Hereditas 2003, 138, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, W.; Ushio, Y.; Seo, A.; Nakato, N.; Kono, M.; Kudoh, H.; Tobe, H.; Murakami, N. Evidence for Hybrid Origin and Segmental Allopolyploidy in Eutetraploid and Aneutetraploid Lepisorus thunbergianus (Polypodiaceae). Syst. Bot. 2010, 35, 20–29. [Google Scholar] [CrossRef]
- Xiong, Z.; Gaeta, R.T.; Pires, J.C. Homoeologous Shuffling and Chromosome Compensation Maintain Genome Balance in Resynthesized Allopolyploid Brassica napus. Proc. Natl. Acad. Sci. USA 2011, 108, 7908–7913. [Google Scholar] [CrossRef] [Green Version]
- Chester, M.; Gallagher, J.P.; Symonds, V.V.; Da Silva, A.V.C.; Mavrodiev, E.V.; Leitch, A.R.; Soltis, P.S.; Soltis, D.E. Extensive Chromosomal Variation in a Recently Formed Natural Allopolyploid Species, Tragopogon miscellus (Asteraceae). Proc. Natl. Acad. Sci. USA 2012, 109, 1176–1181. [Google Scholar] [CrossRef] [Green Version]
- De Storme, N.; Geelen, D. The Impact of Environmental Stress on Male Reproductive Development in Plants: Biological Processes and Molecular Mechanisms. Plant. Cell Environ. 2014, 37, 1–18. [Google Scholar] [CrossRef]
- Fuchs, L.K.; Jenkins, G.; Phillips, D.W. Anthropogenic Impacts on Meiosis in Plants. Front. Plant Sci. 2018, 9, 1429. [Google Scholar] [CrossRef]
- Mable, B.K. ‘Why Polyploidy Is Rarer in Animals than in Plants’: Myths and Mechanisms. Biol. J. Linn. Soc. 2004, 82, 453–466. [Google Scholar] [CrossRef] [Green Version]
- Vanneste, K.; Baele, G.; Maere, S.; Van De Peer, Y. Analysis of 41 Plant Genomes Supports a Wave of Successful Genome Duplications in Association with the Cretaceous-Paleogene Boundary. Genome Res. 2014, 24, 1334–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, A.S.; Pires, J.C. Unreduced Gametes: Meiotic Mishap or Evolutionary Mechanism? Trends Genet. 2015, 31, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Loginova, D.B.; Silkova, O.G. Mechanisms of Unreduced Gamete Formation in Flowering Plants. Russ. J. Genet. 2017, 53, 741–756. [Google Scholar] [CrossRef]
- De Storme, N.; Geelen, D. Pre-Meiotic Endomitosis in the Cytokinesis-Defective Tomato Mutant pmcd1 Generates Tetraploid Meiocytes and Diploid Gametes. J. Exp. Bot. 2013, 64, 2345–2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falistocco, E.; Tosti, N.; Falcinelli, M. Cytomixis in Pollen Mother Cells of Diploid Dactylis, One of the Origins of 2n Gametes. J. Hered. 1995, 86, 448–453. [Google Scholar] [CrossRef]
- Spielman, M.; Preuss, D.; Li, F.L.; Browne, W.E.; Scott, R.J.; Dickinson, H.G. Tetraspore is Required for Male Meiotic Cytokinesis in Arabidopsis thaliana. Development 1997, 124, 2645–2657. [Google Scholar] [CrossRef] [PubMed]
- De Haan, A.; Maceira, N.O.; Lumaret, R.; Delay, J. Production of 2n Gametes in Diploid Subspecies of Dactylis glomerata L. 2. Occurrence and Frequency of 2n Eggs. Ann. Bot. 1992, 69, 345–350. [Google Scholar] [CrossRef]
- Maceira, N.O.; De Haan, A.A.; Aret, R.L.; Billon, M.; Delay, J. Production of 2n Gametes in Diploid Subspecies of Dactylis glomerata L. 1. Occurrence and Frequency of 2n Pollen. Ann. Bot. 1992, 69, 335–343. [Google Scholar] [CrossRef]
- Bretagnolle, F. Pollen Production and Spontaneous Polyploidization in Diploid Populations of Anthoxanthum alpinum. Biol. J. Linn. Soc. 2001, 72, 241–247. [Google Scholar] [CrossRef]
- Ramsey, J. Unreduced Gametes and Neopolyploids in Natural Populations of Achillea borealis (Asteraceae). Heredity 2006, 98, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Sheidai, M.; Jafari, S.; Taleban, P.; Keshavarzi, M. Cytomixis and Unreduced Pollen Grain Formation in Alopecurus L. and Catbrosa Beauv. (Poaceae). Cytologia 2009, 74, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Kovalsky, I.E.; Solís Neffa, V.G. Evidence of 2n Microspore Production in a Natural Diploid Population of Turnera sidoides Subsp. Carnea and Its Relevance in the Evolution of the T. sidoides (Turneraceae) Autopolyploid Complex. J. Plant Res. 2012, 125, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Kreiner, J.M.; Kron, P.; Husband, B.C. Frequency and Maintenance of Unreduced Gametes in Natural Plant Populations: Associations with Reproductive Mode, Life History and Genome Size. New Phytol. 2017, 214, 879–889. [Google Scholar] [CrossRef] [Green Version]
- Comai, L. The Advantages and Disadvantages of Being Polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, M.L.; Mittelsten Scheid, O. Meiosis in Polyploid Plants. Polyploidy Genome Evol. 2012, 33–55. [Google Scholar] [CrossRef]
- Doyle, J.J.; Egan, A.N. Dating the Origins of Polyploidy Events. New Phytol. 2010, 186, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Boden, S.A.; Shadiac, N.; Tucker, E.J.; Langridge, P.; Able, J.A. Expression and Functional Analysis of TaASY1 during Meiosis of Bread Wheat (Triticum aestivum). BMC Mol. Biol. 2007, 8, 65. [Google Scholar] [CrossRef] [Green Version]
- Boden, S.A.; Langridge, P.; Spangenberg, G.; Able, J.A. TaASY1 Promotes Homologous Chromosome Interactions and Is Affected by Deletion of Ph1. Plant J. 2009, 57, 487–497. [Google Scholar] [CrossRef] [PubMed]
- De Bustos, A.; Pérez, R.; Jouve, N. Characterization of the Gene Mre11 and Evidence of Silencing after Polyploidization in Triticum. Theor. Appl. Genet. 2007, 114, 985–999. [Google Scholar] [CrossRef]
- Sutton, T.; Whitford, R.; Baumann, U.; Dong, C.; Able, J.A.; Langridge, P. The Ph2 Pairing Homoeologous Locus of Wheat (Triticum aestivum): Identification of Candidate Meiotic Genes Using a Comparative Genetics Approach. Plant J. 2003, 36, 443–456. [Google Scholar] [CrossRef]
- Griffiths, S.; Sharp, R.; Foote, T.N.; Bertin, I.; Wanous, M.; Reader, S.; Colas, I.; Moore, G. Molecular Characterization of Ph1 as a Major Chromosome Pairing Locus in Polyploid Wheat. Nature 2006, 439, 749–752. [Google Scholar] [CrossRef] [PubMed]
- Sears, E.R. Nullisomic-Tetrasomic Combinations in Hexaploid Wheat. Chromosom. Manip. Plant Genet. 1966, 29–45. [Google Scholar] [CrossRef]
- Feldman, M.; Levy, A.A. Genome Evolution Due to Allopolyploidization in Wheat. Genetics 2012, 192, 763–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, R.; Chapman, V. Genetic Control of the Cytologically Diploid Behaviour of Hexaploid Wheat. Nature 1958, 182, 713–715. [Google Scholar] [CrossRef]
- Driscoll, C.J. Genetic suppression of homoeologous chromosome pairing in hexaploid wheat. Can. J. Genet. Cytol. 1972, 14, 39–42. [Google Scholar] [CrossRef]
- Martinez-Perez, E.; Shaw, P.; Moore, G. The Ph1 Locus Is Needed to Ensure Specific Somatic and Meiotic Centromere Association. Nature 2001, 411, 204–207. [Google Scholar] [CrossRef]
- Sears, E.R. Genetic Control of Chromosome Pairing in Wheat. Annu. Rev. Genet. 1976, 10, 31–51. [Google Scholar] [CrossRef]
- Mello-Sampayo, T. Genetic Regulation of Meiotic Chromosome Pairing by Chromosome 3D of Triticum aestivum. Nat. New Biol. 1971, 230, 22–23. [Google Scholar] [CrossRef]
- Prieto, P.; Moore, G.; Reader, S. Control of Conformation Changes Associated with Homologue Recognition during Meiosis. Theor. Appl. Genet. 2005, 111, 505–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sears, E.R. Induced Mutant with Homoeologous Pairing in Common Wheat. Can. J. Genet. Cytol. 1977, 19, 585–593. [Google Scholar] [CrossRef]
- Okamoto, M. Asynaptic Effect of Chromosome V. Wheat Inf. Serv. 1957, 5, 6. [Google Scholar]
- Sears, R.E. Intergenomic Chromosome Relationships in Hexaploid Wheat. Proc. Int. Congr. Genet. 1958, 2, 258–259. [Google Scholar]
- Richards, D.M.; Greer, E.; Martin, A.C.; Moore, G.; Shaw, P.J.; Howard, M. Quantitative Dynamics of Telomere Bouquet Formation. PLoS Comput. Biol. 2012, 8, e1002812. [Google Scholar] [CrossRef] [Green Version]
- Moore, G. The Control of Recombination in Wheat by Ph1 and Its Use in Breeding. Methods Mol. Biol. 2014, 1145, 143–153. [Google Scholar] [CrossRef]
- Rey, M.D.; Martín, A.C.; Higgins, J.; Swarbreck, D.; Uauy, C.; Shaw, P.; Moore, G. Exploiting the ZIP4 Homologue within the Wheat Ph1 Locus Has Identified Two Lines Exhibiting Homoeologous Crossover in Wheat-Wild Relative Hybrids. Mol. Breed. 2017, 37, 1–11. [Google Scholar] [CrossRef]
- Calderón, M.C.; Rey, M.D.; Martín, A.; Prieto, P. Homoeologous Chromosomes From Two Hordeum Species Can Recognize and Associate During Meiosis in Wheat in the Presence of the Ph1 Locus. Front. Plant Sci. 2018, 9, 585. [Google Scholar] [CrossRef] [Green Version]
- Calderón, M.C.; Prieto, P. Wild and Cultivated Homoeologous Barley Chromosomes Can Associate and Recombine in Wheat in the Absence of the Ph1 Locus. Agron 2021, 11, 147. [Google Scholar] [CrossRef]
- Martín, A.C.; Rey, M.D.; Shaw, P.; Moore, G. Dual Effect of the Wheat Ph1 Locus on Chromosome Synapsis and Crossover. Chromosoma 2017, 126, 669–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, A.C.; Alabdullah, A.K.; Moore, G. A Separation-of-Function ZIP4 Wheat Mutant Allows Crossover between Related Chromosomes and Is Meiotically Stable. Sci. Rep. 2021, 11, 21811. [Google Scholar] [CrossRef]
- Chelysheva, L.; Gendrot, G.; Vezon, D.; Doutriaux, M.P.; Mercier, R.; Grelon, M. Zip4/Spo22 Is Required for Class I CO Formation but Not for Synapsis Completion in Arabidopsis thaliana. PLoS Genet. 2007, 3, e83. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Tang, D.; Wang, K.; Wang, M.; Huang, J.; Luo, W.; Luo, Q.; Hong, L.; Li, M.; Cheng, Z. ZIP4 in Homologous Chromosome Synapsis and Crossover Formation in Rice Meiosis. J. Cell Sci. 2012, 125, 2581–2591. [Google Scholar] [CrossRef] [Green Version]
- Tsubouchi, T.; Zhao, H.; Roeder, G.S. The Meiosis-Specific zip4 Protein Regulates Crossover Distribution by Promoting Synaptonemal Complex Formation Together with zip2. Dev. Cell 2006, 10, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Dubois, E.; De Muyt, A.; Soyer, J.L.; Budin, K.; Legras, M.; Piolot, T.; Debuchy, R.; Kleckner, N.; Zickler, D.; Espagne, E. Building Bridges to Move Recombination Complexes. Proc. Natl. Acad. Sci. USA 2019, 116, 12400–12409. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.; Kleckner, N.; Börner, G.V. Bioinformatic analyses implicate the collaborating meiotic crossover/chiasma proteins Zip2, Zip3, and Spo22/Zip4 in ubiquitin labeling. Proc. Natl. Acad. Sci. USA 2005, 102, 17594–17599. [Google Scholar] [CrossRef] [Green Version]
- Rey, M.D.; Martín, A.C.; Smedley, M.; Hayta, S.; Harwood, W.; Shaw, P.; Moore, G. Magnesium Increases Homoeologous Crossover Frequency During Meiosis in ZIP4 (Ph1 Gene) Mutant Wheat-Wild Relative Hybrids. Front. Plant Sci. 2018, 9, 509. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-González, R.H.; Borrill, P.; Lang, D.; Harrington, S.A.; Brinton, J.; Venturini, L.; Davey, M.; Jacobs, J.; Van Ex, F.; Pasha, A.; et al. The Transcriptional Landscape of Polyploid Wheat. Science 2018, 361, eaar6089. [Google Scholar] [CrossRef] [Green Version]
- Alabdullah, A.K.; Borrill, P.; Martin, A.C.; Ramirez-Gonzalez, R.H.; Hassani-Pak, K.; Uauy, C.; Shaw, P.; Moore, G. A Co-Expression Network in Hexaploid Wheat Reveals Mostly Balanced Expression and Lack of Significant Gene Loss of Homeologous Meiotic Genes Upon Polyploidization. Front. Plant Sci. 2019, 10, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Cuňado, N.; Blazquez, S.; Melchor, L.; Pradillo, M.; Santos, J.L. Understanding the Cytological Diploidization Mechanism of Polyploid Wild Wheats. Cytogenet. Genome Res. 2005, 109, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Rees, H.; Jenkins, G. Synaptonemal Complex Formation in Avena Polyploids. Heredity 1989, 63, 209–219. [Google Scholar] [CrossRef]
- Thomas, H.M.; Thomas, B.J. Synaptonemal Complex Formation in Two Allohexaploid Festuca Species and a Pentaploid Hybrid. Heredity 1993, 71, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Martinez, M.; Naranjo, T.; Cuadrado, C.; Romero, C. The Synaptic Behaviour of the Wild Forms of Triticum turgidum and T. timopheevii. Genome 2001, 44, 517–522. [Google Scholar] [CrossRef]
- Martinez, M.; Naranjo, T.; Cuadrado, C.; Romero, C. Synaptic Behaviour of the Tetraploid Wheat Triticum timopheevii. Theor. Appl. Genet. 1996, 93, 1139–1144. [Google Scholar] [CrossRef]
- Poggio, L.; González, G.E. Cytological Diploidization of Paleopolyploid Genus Zea: Divergence between Homoeologous Chromosomes or Activity of Pairing Regulator Genes? PLoS ONE 2018, 13, e0189644. [Google Scholar] [CrossRef] [Green Version]
- Jena, K.K. The Species of the Genus Oryza and Transfer of Useful Genes from Wild Species into Cultivated Rice, O. Sativa. Breed. Sci. 2010, 60, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.; Sang, T.; Lu, B.R.; Hong, D.Y. Phylogeny of Rice Genomes with Emphasis on Origins of Allotetraploid Species. Proc. Natl. Acad. Sci. USA 1999, 96, 14400–14405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dally, A.M.; Second, G. Chloroplast DNA Diversity in Wild and Cultivated Species of Rice (Genus Oryza, Section Oryza). Cladistic-Mutation and Genetic-Distance Analysis. Theor. Appl. Genet. 1990, 80, 209–222. [Google Scholar] [CrossRef]
- Kanno, A.; Hirai, A. Comparative Studies of the Structure of Chloroplast DNA from Four Species of Oryza: Cloning and Physical Maps. Theor. Appl. Genet. 1992, 83, 791–798. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Second, G.; Tanksley, S.D. Polymorphism and Phylogenetic Relationships among Species in the Genus Oryza as Determined by Analysis of Nuclear RFLPs. Theor. Appl. Genet. 1992, 83, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.K.; Brar, D.S.; Nandi, S.; Huang, N.; Khush, G.S. Phylogenetic Relationships among Oryza Species Revealed by AFLP Markers. Theor. Appl. Genet. 1999, 98, 1320–1328. [Google Scholar] [CrossRef]
- Bao, Y.; Ge, S. Origin and Phylogeny of Oryza Species with the CD Genome Based on Multiple-Gene Sequence Data. Plant Syst. Evol. 2004, 249, 55–66. [Google Scholar] [CrossRef]
- Jena, K.K.; Kochert, G. Restriction Fragment Length Polymorphism Analysis of CCDD Genome Species of the Genus Oryza L. Plant Mol. Biol. 1991, 16, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Du, Y.; Wang, X.; Wang, Q.; Zhang, B.; Chen, J.; Chen, M.; Doyle, J.J.; Ge, S. Genome Evolution in Oryza Allopolyploids of Various Ages: Insights into the Process of Diploidization. Plant J. 2021, 105, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Jenczewski, E.; Eber, F.; Grimaud, A.; Huet, S.; Lucas, M.O.; Monod, H.; Chèvre, A.M. PrBn, a Major Gene Controlling Homeologous Pairing in Oilseed Rape (Brassica Napus) Haploids. Genetics 2003, 164, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes, M.; Grandont, L.; Moore, G.; Chèvre, A.M.; Jenczewski, E. Genetic Regulation of Meiosis in Polyploid Species: New Insights into an Old Question. New Phytol. 2010, 186, 29–36. [Google Scholar] [CrossRef]
- Jenczewski, E.; Alix, K. From Diploids to Allopolyploids: The Emergence of Efficient Pairing Control Genes in Plants. CRC Crit. Rev. Plant Sci. 2004, 23, 21–45. [Google Scholar] [CrossRef]
- Nicolas, S.D.; Leflon, M.; Monod, H.; Eber, F.; Coriton, O.; Huteau, V.; Chèvre, A.M.; Jenczewski, E. Genetic Regulation of Meiotic Cross-Overs between Related Genomes in Brassica napus Haploids and Hybrids. Plant Cell 2009, 21, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Adamczyk, K.; Manzanares-Dauleux, M.; Eber, F.; Lucas, M.O.; Delourme, R.; Chèvre, A.M.; Jenczewski, E. Mapping PrBn and Other Quantitative Trait Loci Responsible for the Control of Homeologous Chromosome Pairing in Oilseed Rape (Brassica napus L.) Haploids. Genetics 2006, 174, 1583–1596. [Google Scholar] [CrossRef] [Green Version]
- Aklilu, B.B.; Soderquist, R.S.; Culligan, K.M. Genetic Analysis of the Replication Protein A Large Subunit Family in Arabidopsis Reveals Unique and Overlapping Roles in DNA Repair, Meiosis and DNA Replication. Nucleic Acids Res. 2014, 42, 3104–3118. [Google Scholar] [CrossRef] [Green Version]
- Berchowitz, L.E.; Francis, K.E.; Bey, A.L.; Copenhaver, G.P. The Role of AtMUS81 in Interference-Insensitive Crossovers in A. thaliana. PLoS Genet. 2007, 3, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Blary, A. Towards a Functional Characterization of Meiotic Recombination in Rapeseed: Analysis of the Meiotic Transcriptome and Hyper-Recombinant Mutants. Ph.D. Thesis, Université Paris-Saclay, Saint-Aubin, France, 2016. [Google Scholar]
- Sheidai, M.; Noormohamadi, Z.; Sotodeh, M. Cytogenetic Variability In Several Canola (Brassica napus) Cultivare. Caryologia 2014, 59, 267–276. [Google Scholar] [CrossRef]
- Mason, A.S.; Batley, J. Creating New Interspecific Hybrid and Polyploid Crops. Trends Biotechnol. 2015, 33, 436–441. [Google Scholar] [CrossRef] [Green Version]
- Szadkowski, E.; Eber, F.; Huteau, V.; Lodé, M.; Huneau, C.; Belcram, H.; Coriton, O.; Manzanares-Dauleux, M.J.; Delourme, R.; King, G.J.; et al. The First Meiosis of Resynthesized Brassica napus, a Genome Blender. New Phytol. 2010, 186, 102–112. [Google Scholar] [CrossRef]
- Higgins, E.E.; Howell, E.C.; Armstrong, S.J.; Parkin, I.A.P. A Major Quantitative Trait Locus on Chromosome A9, BnaPh1, Controls Homoeologous Recombination in Brassica napus. New Phytol. 2021, 229, 3281–3293. [Google Scholar] [CrossRef]
- Culligan, K.M.; Hays, J.B. Arabidopsis MutS Homologs-AtMSH2, AtMSH3, AtMSH6, and a Novel AtMSH7-Form Three Distinct Protein Heterodimers with Different Specificities for Mismatched DNA. Plant Cell 2000, 12, 991–1002. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, A.; Bomblies, K. Meiosis in Autopolyploid and Allopolyploid Arabidopsis. Curr. Opin. Plant Biol. 2016, 30, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Comai, L.; Tyagi, A.P.; Lysak, M.A. FISH Analysis of Meiosis in Arabidopsis Allopolyploids. Chromosom. Res. 2003, 11, 217–226. [Google Scholar] [CrossRef]
- Henry, I.M.; Dilkes, B.P.; Tyagi, A.; Gao, J.; Christensen, B.; Comaia, L. The Boy Named sue Quantitative Trait Locus Confers Increased Meiotic Stability to an Adapted Natural Allopolyploid of Arabidopsis. Plant Cell 2014, 26, 181–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontes, O.; Neves, N.; Silva, M.; Lewis, M.S.; Madlung, A.; Comai, L.; Viegas, W.; Pikaard, C.S. Chromosomal Locus Rearrangements Are a Rapid Response to Formation of the Allotetraploid Arabidopsis suecica Genome. Proc. Natl. Acad. Sci. USA 2004, 101, 18240–18245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, R.; Mandáková, T.; Gunis, J.; Soto-Jiménez, L.M.; Liu, C.; Lysak, M.A.; Novikova, P.Y.; Nordborg, M. Gradual Evolution of Allopolyploidy in Arabidopsis suecica. Nat. Ecol. Evol. 2021, 5, 1367–1381. [Google Scholar] [CrossRef] [PubMed]
- Yant, L.; Hollister, J.D.; Wright, K.M.; Arnold, B.J.; Higgins, J.D.; Franklin, F.C.H.; Bomblies, K. Meiotic Adaptation to Genome Duplication in Arabidopsis arenosa. Curr. Biol. 2013, 23, 2151–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.D.; Wright, K.M.; Bomblies, K.; Franklin, F.C.H. Cytological Techniques to Analyze Meiosis in Arabidopsis arenosa for Investigating Adaptation to Polyploidy. Front. Plant Sci. 2014, 4, 546. [Google Scholar] [CrossRef] [Green Version]
- Bomblies, K.; Jones, G.; Franklin, C.; Zickler, D.; Kleckner, N. The Challenge of Evolving Stable Polyploidy: Could an Increase in “Crossover Interference Distance” Play a Central Role? Chromosoma 2016, 125, 287–300. [Google Scholar] [CrossRef] [Green Version]
- Morgan, C.; White, M.A.; Franklin, F.C.H.; Zickler, D.; Kleckner, N.; Bomblies, K. Evolution of Crossover Interference Enables Stable Autopolyploidy by Ensuring Pairwise Partner Connections in Arabidopsis arenosa. Curr. Biol. 2021, 31, 4713–4726.e4. [Google Scholar] [CrossRef]
- Davies, A.; Jenkins, G.; Rees, H. Diploidisation of Lotus corniculatus L. (Fabaceae) by Elimination of Multivalents. Chromosoma 1990, 99, 289–295. [Google Scholar] [CrossRef]
- Wallace, A.J.; Callow, R.S. Meiotic Variation in an Intergenomic Autopolyploid Series. I. Chiasma Frequency. Genome 1995, 38, 122–132. [Google Scholar] [CrossRef]
- Wentworth, J.E.; Gornall, R.J. Cytogenetic Evidence for Autopolyploidy in Parnassia palustris. New Phytol. 1996, 134, 641–648. [Google Scholar] [CrossRef]
- Hollister, J.D.; Arnold, B.J.; Svedin, E.; Xue, K.S.; Dilkes, B.P.; Bomblies, K. Genetic Adaptation Associated with Genome-Doubling in Autotetraploid Arabidopsis arenosa. PLoS Genet. 2012, 8, e1003093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, K.M.; Arnold, B.; Xue, K.; Surinova, M.; O’Connell, J.; Bomblies, K. Selection on Meiosis Genes in Diploid and Tetraploid Arabidopsis arenosa. Mol. Biol. Evol. 2015, 32, 944–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, C.; Zhang, H.; Henry, C.E.; Franklin, C.F.H.; Bomblies, K. Derived Alleles of Two Axis Proteins Affect Meiotic Traits in Autotetraploid Arabidopsis arenosa. Proc. Natl. Acad. Sci. USA 2020, 117, 8980–8988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seear, P.J.; France, M.G.; Gregory, C.L.; Heavens, D.; Schmickl, R.; Yant, L.; Higgins, J.D. A Novel Allele of ASY3 Is Associated with Greater Meiotic Stability in Autotetraploid Arabidopsis lyrata. PLoS Genet. 2020, 16, e1008900. [Google Scholar] [CrossRef] [PubMed]
- Henry, I.M.; Dilkes, B.P.; Young, K.; Watson, B.; Wu, H.; Comai, L. Aneuploidy and Genetic Variation in the Arabidopsis thaliana Triploid Response. Genetics 2005, 170, 1979–1988. [Google Scholar] [CrossRef] [Green Version]
- Chao, D.Y.; Dilkes, B.; Luo, H.; Douglas, A.; Yakubova, E.; Lahner, B.; Salt, D.E. Polyploids Exhibit Higher Potassium Uptake and Salinity Tolerance in Arabidopsis. Science 2013, 341, 658–659. [Google Scholar] [CrossRef] [Green Version]
- Weiss, H.; Maluszynska, J. Chromosomal Rearrangement in Autotetraploid Plants of Arabidopsis thaliana. Hereditas 2000, 133, 255–261. [Google Scholar] [CrossRef]
- Santos, J.L.; Alfaro, D.; Sanchez-Moran, E.; Armstrong, S.J.; Franklin, F.C.H.; Jones, G.H. Partial Diploidization of Meiosis in Autotetraploid Arabidopsis thaliana. Genetics 2003, 165, 1533–1540. [Google Scholar] [CrossRef]
- Yu, Z.; Haage, K.; Streit, V.E.; Gierl, A.; Torres Ruiz, R.A. A Large Number of Tetraploid Arabidopsis thaliana Lines, Generated by a Rapid Strategy, Reveal High Stability of Neo-Tetraploids during Consecutive Generations. Theor. Appl. Genet. 2009, 118, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Parra-Nunez, P.; Pradillo, M.; Santos, J.L. Competition for Chiasma Formation between Identical and Homologous (But Not Identical) Chromosomes in Synthetic Autotetraploids of Arabidopsis thaliana. Front. Plant Sci. 2019, 9, 1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felcher, K.J.; Coombs, J.J.; Massa, A.N.; Hansey, C.N.; Hamilton, J.P.; Veilleux, R.E.; Buell, C.R.; Douches, D.S. Integration of Two Diploid Potato Linkage Maps with the Potato Genome Sequence. PLoS ONE 2012, 7, e36347. [Google Scholar] [CrossRef] [Green Version]
- Bourke, P.M.; Voorrips, R.E.; Visser, R.G.F.; Maliepaard, C. The Double-Reduction Landscape in Tetraploid Potato as Revealed by a High-Density Linkage Map. Genetics 2015, 201, 853–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, F.; Song, J.; Naess, S.K.; Helgeson, J.P.; Gebhardt, C.; Jiang, J. Development and Applications of a Set of Chromosome-Specific Cytogenetic DNA Markers in Potato. Theor. Appl. Genet. 2000, 101, 1001–1007. [Google Scholar] [CrossRef]
- Tang, X.; De Boer, J.M.; Van Eck, H.J.; Bachem, C.; Visser, R.G.F.; De Jong, H. Assignment of Genetic Linkage Maps to Diploid Solanum tuberosum Pachytene Chromosomes by BAC-FISH Technology. Chromosom. Res. 2009, 17, 899–915. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Braz, G.T.; Torres, G.A.; Jiang, J. Chromosome Painting in Meiosis Reveals Pairing of Specific Chromosomes in Polyploid Solanum Species. Chromosoma 2018, 127, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, T.; Corredor, E. Clustering of Centromeres Precedes Bivalent Chromosome Pairing of Polyploid Wheats. Trends Plant Sci. 2004, 9, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Wright, L.; Ponce, O.; Chen, J.; Prashar, A.; Sanchez-Moran, E.; Luo, Z.; Compton, L. Varietal Variation and Chromosome Behaviour during Meiosis in Solanum tuberosum. Heredity 2020, 125, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Marand, A.P.; Jansky, S.H.; Zhao, H.; Leisner, C.P.; Zhu, X.; Zeng, Z.; Crisovan, E.; Newton, L.; Hamernik, A.J.; Veilleux, R.E.; et al. Meiotic Crossovers Are Associated with Open Chromatin and Enriched with Stowaway Transposons in Potato. Genome Biol. 2017, 18, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenan-Eichler, M.; Leshkowitz, D.; Tal, L.; Noor, E.; Melamed-Bessudo, C.; Feldman, M.; Levy, A.A. Wheat Hybridization and Polyploidization Results in Deregulation of Small RNAs. Genetics 2011, 188, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Rana, K.; Xiong, Z.; Ge, X.; Li, Z.; Song, H.; Qian, W. Genetic and Epigenetic Alterations in Hybrid and Derived Hexaploids between Brassica napus and B. oleracea Revealed by SSR and MSAP Analysis. Acta Physiol. Plant. 2019, 41, 1–9. [Google Scholar] [CrossRef]
- Song, Q.; Chen, J.Z. Epigenetic and Developmental Regulation in Plant Polyploids. Curr. Opin. Plant Biol. 2015, 24, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Nieto Feliner, G.; Casacuberta, J.; Wendel, J.F. Genomics of Evolutionary Novelty in Hybrids and Polyploids. Front. Genet. 2020, 11, 1–21. [Google Scholar] [CrossRef]
- Wang, X.; Morton, J.A.; Pellicer, J.; Leitch, I.J.; Leitch, A.R. Genome Downsizing after Polyploidy: Mechanisms, Rates and Selection Pressures. Plant J. 2021, 107, 1003–1015. [Google Scholar] [CrossRef]
- Zhao, N.; Dong, Q.; Nadon, B.D.; Ding, X.; Wang, X.; Dong, Y.; Liu, B.; Jackson, S.A.; Xu, C. Evolution of Homeologous Gene Expression in Polyploid Wheat. Genes 2020, 11, 1401. [Google Scholar] [CrossRef]
- Xu, C.; Nadon, B.D.; Do Kim, K.; Jackson, S.A. Genetic and Epigenetic Divergence of Duplicate Genes in Two Legume Species. Plant Cell Environ. 2018, 41, 2033–2044. [Google Scholar] [CrossRef]
- Eilam, T.; Anikster, Y.; Millet, E.; Manisterski, J.; Feldman, M. Genome Size in Diploids, Allopolyploids, and Autopolyploids of Mediterranean Triticeae. J. Bot. 2010, 2010, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Brenchley, R.; Spannagl, M.; Pfeifer, M.; Barker, G.L.A.; D’Amore, R.; Allen, A.M.; McKenzie, N.; Kramer, M.; Kerhornou, A.; Bolser, D.; et al. Analysis of the Bread Wheat Genome Using Whole-Genome Shotgun Sequencing. Nature 2012, 491, 705–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Vega, J.M.; Feldman, M. Rapid Genomic Changes in Newly Synthesized Amphiploids of Triticum and Aegilops. II. Changes in Low-Copy Coding DNA Sequences. Genome 1998, 41, 535–542. [Google Scholar] [CrossRef]
- Benyahya, F.; Nadaud, I.; Da Ines, O.; Rimbert, H.; White, C.; Sourdille, P. SPO11.2 Is Essential for Programmed Double-Strand Break Formation during Meiosis in Bread Wheat (Triticum aestivum L.). Plant J. 2020, 104, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A. Gene Duplication as a Driver of Plant Morphogenetic Evolution. Curr. Opin. Plant Biol. 2014, 17, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Vicient, C.M.; Casacuberta, J.M. Impact of Transposable Elements on Polyploid Plant Genomes. Ann. Bot. 2017, 120, 195–207. [Google Scholar] [CrossRef]
- Glombik, M.; Bačovský, V.; Hobza, R.; Kopecký, D. Competition of Parental Genomes in Plant Hybrids. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Takahagi, K.; Inoue, K.; Mochida, K. Gene Co-Expression Network Analysis Suggests the Existence of Transcriptional Modules Containing a High Proportion of Transcriptionally Differentiated Homoeologs in Hexaploid Wheat. Front. Plant Sci. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Liu, S.L.; Adams, K.L. Dramatic Change in Function and Expression Pattern of a Gene Duplicated by Polyploidy Created a Paternal Effect Gene in the Brassicaceae. Mol. Biol. Evol. 2010, 27, 2817–2828. [Google Scholar] [CrossRef] [Green Version]
- Mason, A.S.; Wendel, J.F. Homoeologous Exchanges, Segmental Allopolyploidy, and Polyploid Genome Evolution. Front. Genet. 2020, 11, 1014. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The Genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Ren, F.; Zhang, L.; Liu, Y.; Chen, X.; Li, Y.; Zhang, L.; Zhu, B.; Zeng, P.; Li, Z.; et al. Unstable Allotetraploid Tobacco Genome Due to Frequent Homeologous Recombination, Segmental Deletion, and Chromosome Loss. Mol. Plant 2018, 11, 914–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurgobin, B.; Golicz, A.A.; Bayer, P.E.; Chan, C.K.K.; Tirnaz, S.; Dolatabadian, A.; Schiessl, S.V.; Samans, B.; Montenegro, J.D.; Parkin, I.A.P.; et al. Homoeologous Exchange Is a Major Cause of Gene Presence/Absence Variation in the Amphidiploid Brassica napus. Plant Biotechnol. J. 2018, 16, 1265–1274. [Google Scholar] [CrossRef] [Green Version]
- Samans, B.; Snowdon, R.; Mason, A.S. Homoeologous Exchanges and Gene Losses Generate Diversity and Differentiate the B. napus Genome from That of Its Ancestors. In Brassica Napus Genome; Liu, S., Snowdon, R., Chalhoub, B., Eds.; Springer: Cham, Switzerland, 2018; pp. 131–148. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Dudchenko, O.; Gao, D.; Seijo, G.; Leal-Bertioli, S.C.M.; Ren, L.; Farmer, A.D.; Pandey, M.K.; et al. The Genome Sequence of Segmental Allotetraploid Peanut Arachis hypogaea. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, W.; Chen, H.; Yang, M.; Wang, J.; Pandey, M.K.; Zhang, C.; Chang, W.C.; Zhang, L.; Zhang, X.; Tang, R.; et al. The Genome of Cultivated Peanut Provides Insight into Legume Karyotypes, Polyploid Evolution and Crop Domestication. Nat. Genet. 2019, 51, 865–876. [Google Scholar] [CrossRef]
- Sun, Y.; Wu, Y.; Yang, C.; Sun, S.; Lin, X.; Liu, L.; Xu, C.; Wendel, J.F.; Gong, L.; Liu, B. Segmental Allotetraploidy Generates Extensive Homoeologous Expression Rewiring and Phenotypic Diversity at the Population Level in Rice. Mol. Ecol. 2017, 26, 5451–5466. [Google Scholar] [CrossRef]
- Li, N.; Xu, C.; Zhang, A.; Lv, R.; Meng, X.; Lin, X.; Gong, L.; Wendel, J.F.; Liu, B. DNA Methylation Repatterning Accompanying Hybridization, Whole Genome Doubling and Homoeolog Exchange in Nascent Segmental Rice Allotetraploids. New Phytol. 2019, 223, 979–992. [Google Scholar] [CrossRef] [Green Version]
- Woodhouse, M.R.; Cheng, F.; Pires, J.C.; Lisch, D.; Freeling, M.; Wang, X. Erratum: Origin, Inheritance, and Gene Regulatory Consequences of Genome Dominance in Polyploids. Proc. Natl. Acad. Sci. USA 2014, 111, 6527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.Q.; Tang, R.H.; Jiang, J.; Xiong, F.Q.; Huang, Z.P.; Wu, H.N.; Gao, Z.K.; Zhong, R.C.; He, X.H.; Han, Z.Q. Rapid Gene Expression Change in a Novel Synthesized Allopolyploid Population of Cultivated Peanut×Arachis doigoi Cross by CDNA-SCoT and HFO-TAG Technique. J. Integr. Agric. 2017, 16, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Hu, B.; Becker, C.; Doğan, E.S.; Berendzen, K.W.; Weigel, D.; Liu, C. Altered Chromatin Compaction and Histone Methylation Drive Non-Additive Gene Expression in an Interspecific Arabidopsis Hybrid. Genome Biol. 2017, 18, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Combes, M.C.; Hueber, Y.; Dereeper, A.; Rialle, S.; Herrera, J.C.; Lashermes, P. Regulatory Divergence between Parental Alleles Determines Gene Expression Patterns in Hybrids. Genome Biol. Evol. 2015, 7, 1110–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, G.; Wendel, J.F. Cis–Trans Controls and Regulatory Novelty Accompanying Allopolyploidization. New Phytol. 2019, 221, 1691–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Wang, P.; Li, J.; Zhao, Q.; Ji, C.; Zhu, Z.; Zhai, Y.; Qin, X.; Zhou, J.; Yu, H.; et al. Whole-Genome Sequence of Synthesized Allopolyploids in Cucumis Reveals Insights into the Genome Evolution of Allopolyploidization. Adv. Sci. 2021, 8, 1–15. [Google Scholar] [CrossRef]
- Yoo, M.J.; Szadkowski, E.; Wendel, J.F. Homoeolog Expression Bias and Expression Level Dominance in Allopolyploid Cotton. Heredity 2013, 110, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicker, T.; Gundlach, H.; Spannagl, M.; Uauy, C.; Borrill, P.; Ramírez-González, R.H.; De Oliveira, R.; Mayer, K.F.X.; Paux, E.; Choulet, F. Impact of Transposable Elements on Genome Structure and Evolution in Bread Wheat. Genome Biol. 2018, 19, 1–18. [Google Scholar] [CrossRef]
- Yu, X.; Zhai, Y.; Wang, P.; Cheng, C.; Li, J.; Lou, Q.; Chen, J. Morphological, Anatomical and Photosynthetic Consequences of Artificial Allopolyploidization in Cucumis. Euphytica 2021, 217, 5. [Google Scholar] [CrossRef]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten Things You Should Know about Transposable Elements. Genome Biol. 2018, 19, 1–12. [Google Scholar] [CrossRef]
- Bariah, I.; Keidar-Friedman, D.; Kashkush, K. Where the Wild Things Are: Transposable Elements as Drivers of Structural and Functional Variations in the Wheat Genome. Front. Plant Sci. 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Venkatesh; Nandini, B. Miniature Inverted-Repeat Transposable Elements (MITEs), Derived Insertional Polymorphism as a Tool of Marker Systems for Molecular Plant Breeding. Mol. Biol. Rep. 2020, 47, 3155–3167. [Google Scholar] [CrossRef]
- Lisch, D. How Important Are Transposons for Plant Evolution? Nat. Rev. Genet. 2013, 14, 49–61. [Google Scholar] [CrossRef]
- Quesneville, H. Twenty years of transposable element analysis in the Arabidopsis thaliana genome. Mob. DNA 2020, 11, 11–13. [Google Scholar]
- Arkhipova, I.R. Neutral Theory, Transposable Elements, and Eukaryotic Genome Evolution. Mol. Biol. Evol. 2018, 35, 1332–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quadrana, L.; Etcheverry, M.; Gilly, A.; Caillieux, E.; Madoui, M.A.; Guy, J.; Bortolini Silveira, A.; Engelen, S.; Baillet, V.; Wincker, P.; et al. Transposition Favors the Generation of Large Effect Mutations That May Facilitate Rapid Adaption. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.F.; Ryan, P.R. The Role of Transposable Elements in the Evolution of Aluminium Resistance in Plants. J. Exp. Bot. 2019, 70, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, J.; Yeom, S.I.; Kim, Y.M.; Seo, E.; Kim, K.T.; Kim, M.S.; Lee, J.M.; Cheong, K.; Shin, H.S.; et al. New Reference Genome Sequences of Hot Pepper Reveal the Massive Evolution of Plant Disease-Resistance Genes by Retroduplication. Genome Biol. 2017, 18, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poretti, M.; Praz, C.R.; Meile, L.; Kälin, C.; Schaefer, L.K.; Schläfli, M.; Widrig, V.; Sanchez-Vallet, A.; Wicker, T.; Bourras, S. Domestication of High-Copy Transposons Underlays the Wheat Small RNA Response to an Obligate Pathogen. Mol. Biol. Evol. 2020, 37, 839–848. [Google Scholar] [CrossRef]
- Kent, T.V.; Uzunović, J.; Wright, S.I. Coevolution between Transposable Elements and Recombination. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160458. [Google Scholar] [CrossRef]
- Choudhury, R.R.; Rogivue, A.; Gugerli, F.; Parisod, C. Impact of Polymorphic Transposable Elements on Linkage Disequilibrium along Chromosomes. Mol. Ecol. 2019, 28, 1550–1562. [Google Scholar] [CrossRef] [Green Version]
- Kashkush, K.; Yaakov, B. Methylation, Transcription, and Rearrangements of Transposable Elements in Synthetic Allopolyploids. Int. J. Plant Genom. 2011, 2011, 569826. [Google Scholar] [CrossRef] [Green Version]
- Kashkush, K.; Feldman, M.; Levy, A.A. Transcriptional Activation of Retrotransposons Alters the Expression of Adjacent Genes in Wheat. Nat. Genet. 2003, 33, 102–106. [Google Scholar] [CrossRef]
- Cheng, H.; Sun, G.; He, S.; Gong, W.; Peng, Z.; Wang, R.; Lin, Z.; Du, X. Comparative Effect of Allopolyploidy on Transposable Element Composition and Gene Expression between Gossypium hirsutum and Its Two Diploid Progenitors. J. Integr. Plant Biol. 2019, 61, 45–59. [Google Scholar] [CrossRef] [Green Version]
- Petit, M.; Guidat, C.; Daniel, J.; Denis, E.; Montoriol, E.; Bui, Q.T.; Lim, K.Y.; Kovarik, A.; Leitch, A.R.; Grandbastien, M.A.; et al. Mobilization of Retrotransposons in Synthetic Allotetraploid Tobacco. New Phytol. 2010, 186, 135–147. [Google Scholar] [CrossRef]
- Sarilar, V.; Palacios, P.M.; Rousselet, A.; Ridel, C.; Falque, M.; Eber, F.; Chèvre, A.M.; Joets, J.; Brabant, P.; Alix, K. Allopolyploidy Has a Moderate Impact on Restructuring at Three Contrasting Transposable Element Insertion Sites in Resynthesized Brassica napus Allotetraploids. New Phytol. 2013, 198, 593–604. [Google Scholar] [CrossRef]
- Baduel, P.; Quadrana, L.; Hunter, B.; Bomblies, K.; Colot, V. Relaxed Purifying Selection in Autopolyploids Drives Transposable Element Over-Accumulation Which Provides Variants for ocal Adaptation. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das Gupta, M.; Tsiantis, M. Gene Networks and the Evolution of Plant Morphology. Curr. Opin. Plant Biol. 2018, 45, 82–87. [Google Scholar] [CrossRef]
- Wang, J.; Tian, L.; Lee, H.S.; Wei, N.E.; Jiang, H.; Watson, B.; Madlung, A.; Osborn, T.C.; Doerge, R.W.; Comai, L.; et al. Genomewide Nonadditive Gene Regulation in Arabidopsis Allotetraploids. Genetics 2006, 172, 507–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, J.A.; Ni, Z.; Scheen, A.C.; Koh, J.; Gilbert, C.A.; Lefkowitz, D.; Chen, Z.J.; Soltis, P.S.; Soltis, D.E. Evolution and Expression of Homeologous Loci in Tragopogon miscellus (Asteraceae), a Recent and Reciprocally Formed Allopolyploid. Genetics 2006, 173, 1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Combes, M.C.; Cenci, A.; Baraille, H.; Bertrand, B.; Lashermes, P. Homeologous Gene Expression in Response to Growing Temperature in a Recent Allopolyploid (Coffea arabica L.). J. Hered. 2012, 103, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Bai, Y.; Lin, X.; Zhao, N.; Hu, L.; Gong, Z.; Wendel, J.F.; Liu, B. Genome-Wide Disruption of Gene Expression in Allopolyploids but Not Hybrids of Rice Subspecies. Mol. Biol. Evol. 2014, 31, 1066–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edger, P.P.; Smith, R.; McKain, M.R.; Cooley, A.M.; Vallejo-Marin, M.; Yuan, Y.; Bewick, A.J.; Ji, L.; Platts, A.E.; Bowman, M.J.; et al. Subgenome Dominance in an Interspecific Hybrid, Synthetic Allopolyploid, and a 140-Year-Old Naturally Established Neo-Allopolyploid Monkeyflower. Plant Cell 2017, 29, 2150–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagué, V.; Just, J.; Mestiri, I.; Balzergue, S.; Tanguy, A.M.; Huneau, C.; Huteau, V.; Belcram, H.; Coriton, O.; Jahier, J.; et al. Genome-Wide Gene Expression Changes in Genetically Stable Synthetic and Natural Wheat Allohexaploids. New Phytol. 2010, 187, 1181–1194. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Li, Y.; Zhang, Z.; Li, L.; Liu, B. Transcriptome Asymmetry in Synthetic and Natural Allotetraploid Wheats, Revealed by RNA-Sequencing. New Phytol. 2016, 209, 1264–1277. [Google Scholar] [CrossRef]
- Flagel, L.; Udall, J.; Nettleton, D.; Wendel, J. Duplicate Gene Expression in Allopolyploid Gossypium Reveals Two Temporally Distinct Phases of Expression Evolution. BMC Biol. 2008, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ding, M.; Chen, Z.J. Epigenetic Perspectives on the Evolution and Domestication of Polyploid Plant and Crops. Curr. Opin. Plant Biol. 2018, 42, 37–48. [Google Scholar] [CrossRef]
- Bao, Y.; Hu, G.; Grover, C.E.; Conover, J.; Yuan, D.; Wendel, J.F. Unraveling Cis and Trans Regulatory Evolution during Cotton Domestication. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wendel, J.F.; Jackson, S.A.; Meyers, B.C.; Wing, R.A. Evolution of Plant Genome Architecture. Genome Biol. 2016, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- D’Ario, M.; Griffiths-Jones, S.; Kim, M. Small RNAs: Big Impact on Plant Development. Trends Plant Sci. 2017, 22, 1056–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crisp, P.A.; Hammond, R.; Zhou, P.; Vaillancourt, B.; Lipzen, A.; Daum, C.; Barry, K.; De Leon, N.; Buell, C.R.; Kaeppler, S.M.; et al. Variation and Inheritance of Small RNAs in Maize Inbreds and F1 Hybrids. Plant Physiol. 2020, 182, 318–331. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Liu, D.; Wu, J.; Zhao, X.; Hao, M.; Geng, S.; Yan, J.; Jiang, X.; Zhang, L.; Wu, J.; et al. MRNA and Small RNA Transcriptomes Reveal Insights into Dynamic Homoeolog Regulation of Allopolyploid Heterosis in Nascent Hexaploid Wheat. Plant Cell 2014, 26, 1878–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, F.H.; McKenzie, N.; Gardiner, L.J.; Luo, M.C.; Hall, A.; Bevan, M.W. Reduced Chromatin Accessibility Underlies Gene Expression Differences in Homologous Chromosome Arms of Diploid Aegilops tauschii and Hexaploid Wheat. Gigascience 2021, 9, 1–11. [Google Scholar] [CrossRef]
- Guo, X.; Han, F. Asymmetric Epigenetic Modification and Elimination of RDNA Sequences by Polyploidization in Wheat. Plant Cell 2014, 26, 4311–4327. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, R.J.; Earley, K.; Pontes, O.; Silva, M.; Chen, Z.J.; Neves, N.; Viegas, W.; Pikaard, C.S. A Concerted DNA Methylation/Histone Methylation Switch Regulates rRNA Gene Dosage Control and Nucleolar Dominance. Mol. Cell 2004, 13, 599–609. [Google Scholar] [CrossRef]
- Xu, Y.; Zhong, L.; Wu, X.; Fang, X.; Wang, J. Rapid Alterations of Gene Expression and Cytosine Methylation in Newly Synthesized Brassica napus Allopolyploids. Planta 2009, 229, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tian, L.; Lee, H.S.; Chen, Z.J. Nonadditive Regulation of FRI and FLC Loci Mediates Flowering-Time Variation in Arabidopsis Allopolyploids. Genetics 2006, 173, 965–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.; Zhang, T.; Stelly, D.M.; Chen, Z.J. Epigenomic and Functional Analyses Reveal Roles of Epialleles in the Loss of Photoperiod Sensitivity during Domestication of Allotetraploid Cottons. Genome Biol. 2017, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blasio, F.; Prieto, P.; Pradillo, M.; Naranjo, T. Genomic and Meiotic Changes Accompanying Polyploidization. Plants 2022, 11, 125. https://doi.org/10.3390/plants11010125
Blasio F, Prieto P, Pradillo M, Naranjo T. Genomic and Meiotic Changes Accompanying Polyploidization. Plants. 2022; 11(1):125. https://doi.org/10.3390/plants11010125
Chicago/Turabian StyleBlasio, Francesco, Pilar Prieto, Mónica Pradillo, and Tomás Naranjo. 2022. "Genomic and Meiotic Changes Accompanying Polyploidization" Plants 11, no. 1: 125. https://doi.org/10.3390/plants11010125