

Inhibitory Potential of Synthetic Amino Acid Derivatives against Digestive Enzymes as Promising Hypoglycemic and Anti-Obesity Agents
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
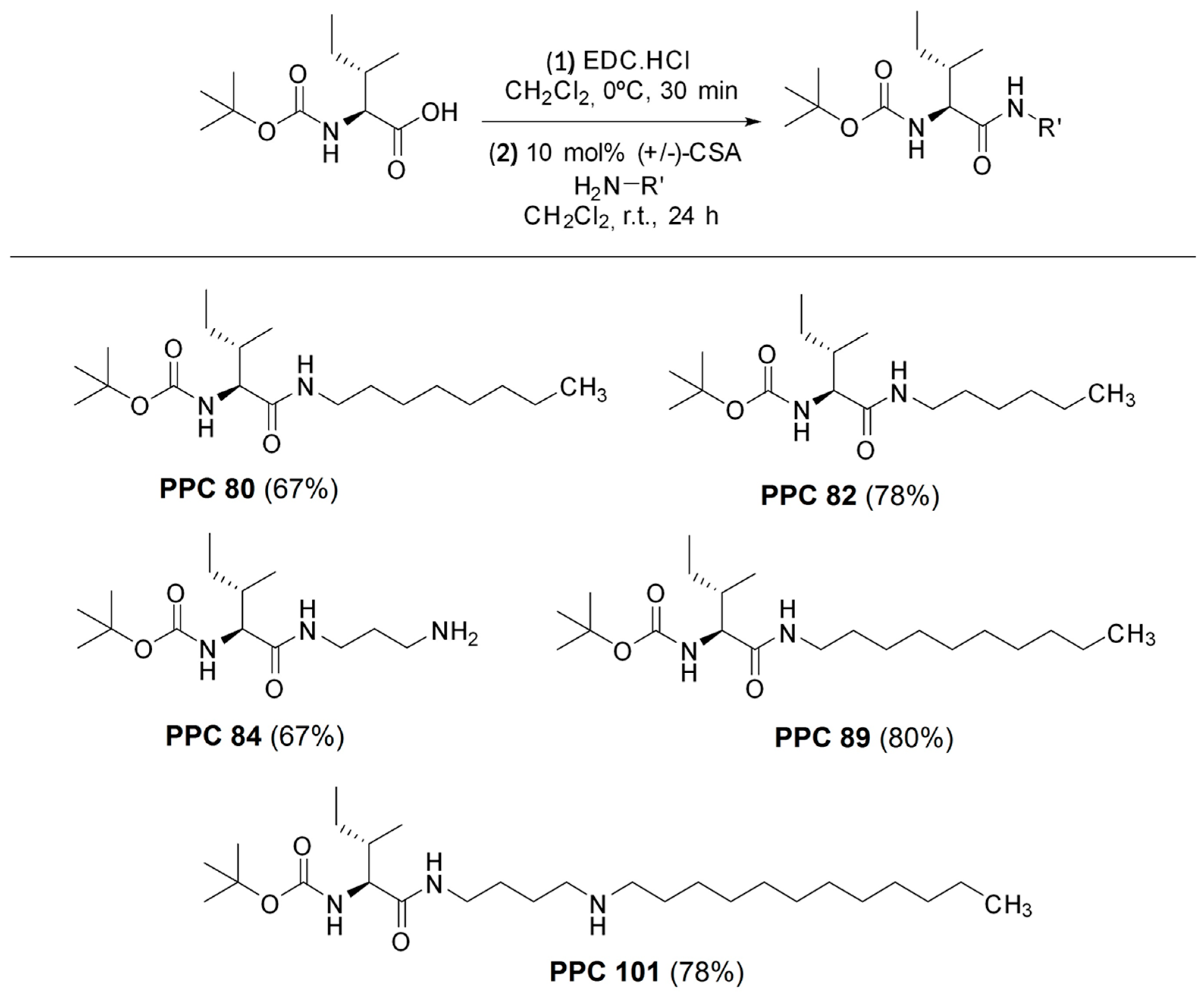
2.1. Synthesis of Amino Acid Derivatives
2.2. Chemicals
2.3. Inhibitory Activity on Digestive Enzymes
2.3.1. Pancreatic Lipase Inhibition Assay
2.3.2. Pancreatic α-Amylase Inhibition Assay
2.3.3. α-Glucosidase Inhibition Assay
2.3.4. Determination of the Inhibitory Effect and IC50
2.3.5. Determination of Kinetic Parameters
2.4. Statistical Analysis
3. Results
3.1. Synthesis of Protected Amino Acid Derivatives
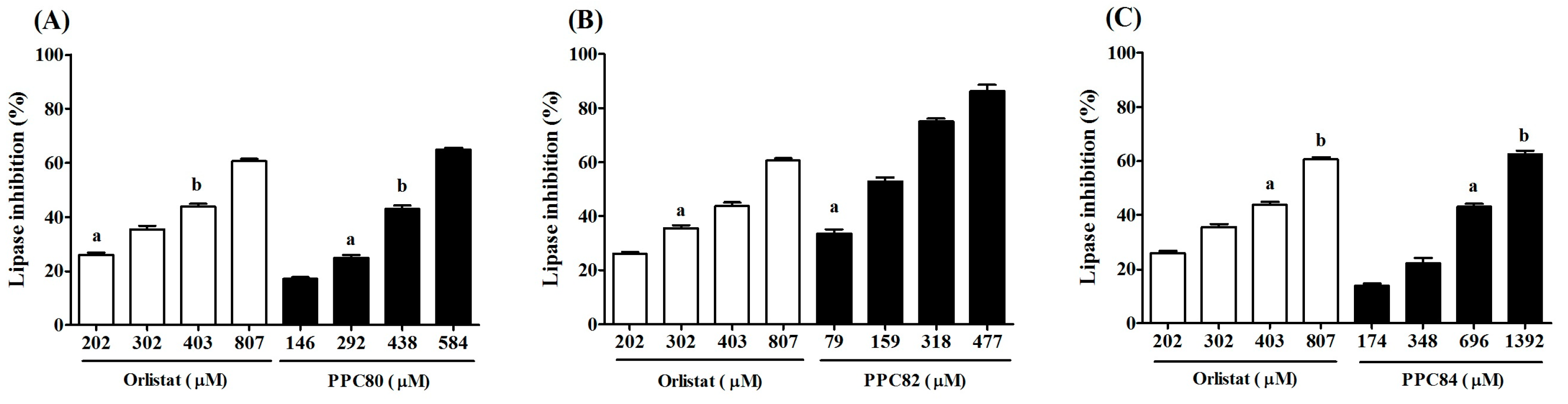
3.2. Inhibitory Effect of Amino Acid Derivatives on Pancreatic Lipase
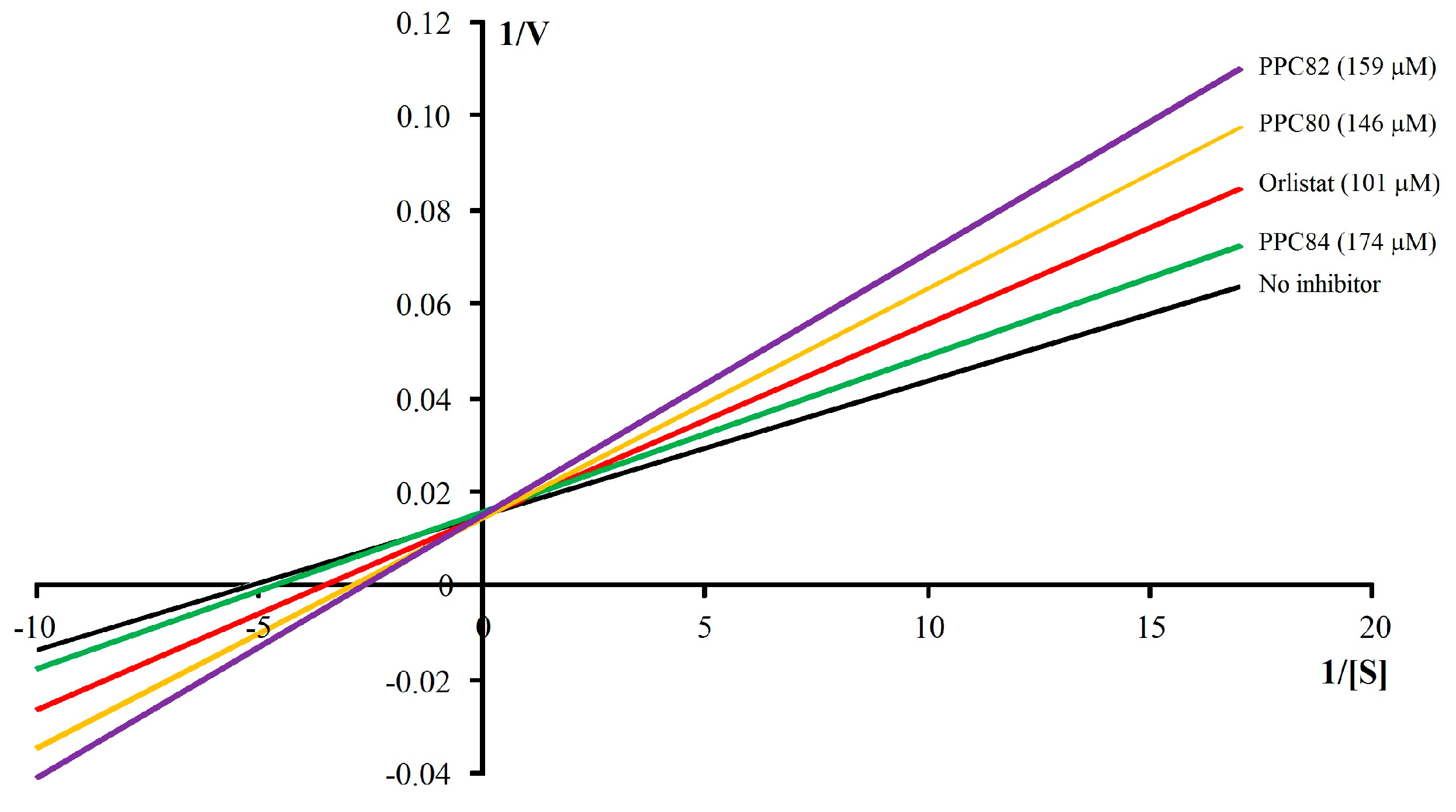
3.3. Kinetic Parameters on Pancreatic Lipase Activity
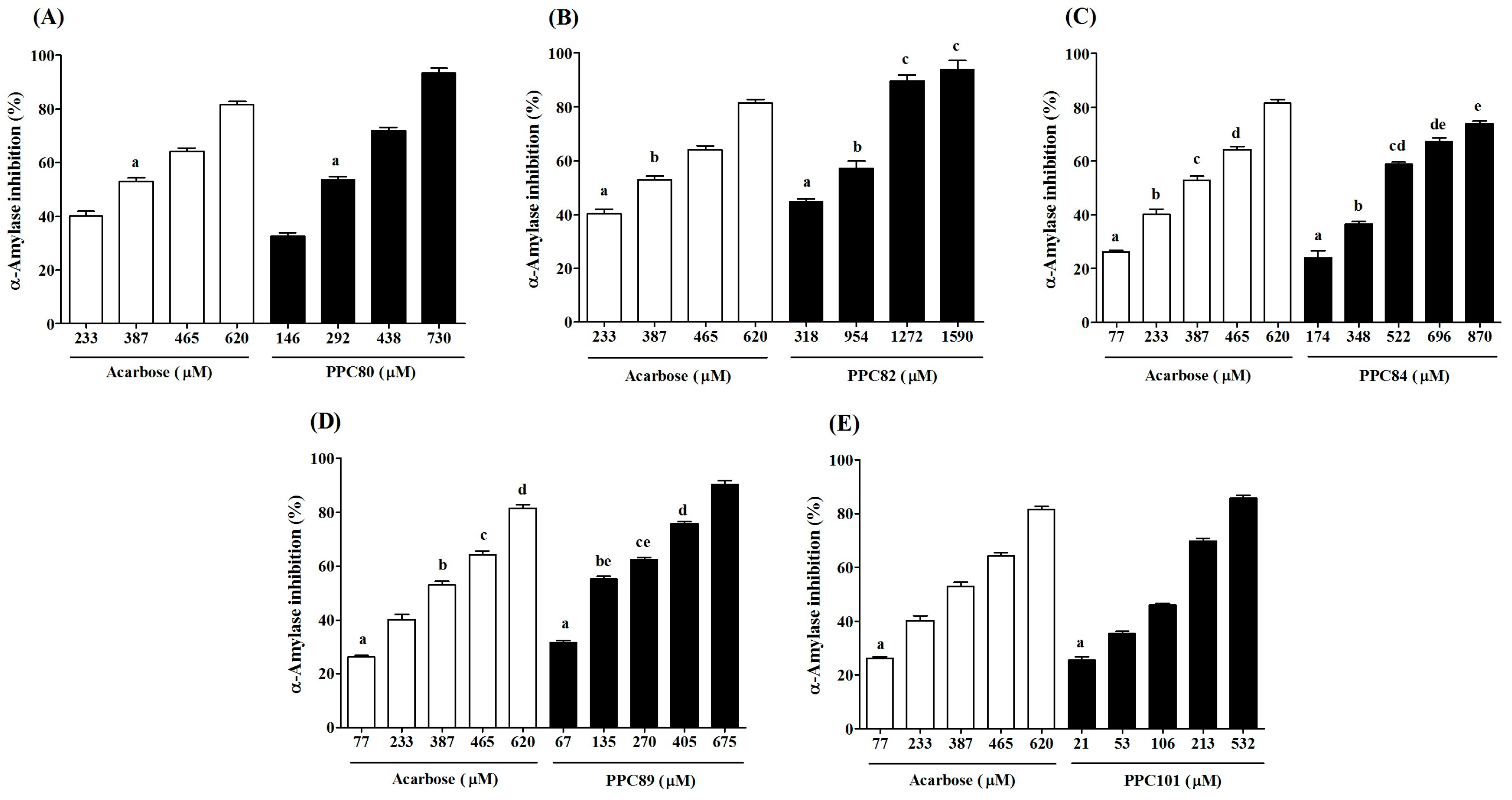
3.4. Inhibitory Effect of Amino Acid Derivatives on Pancreatic α-Amylase
3.5. Kinetic Parameters against Pancreatic α-Amylase
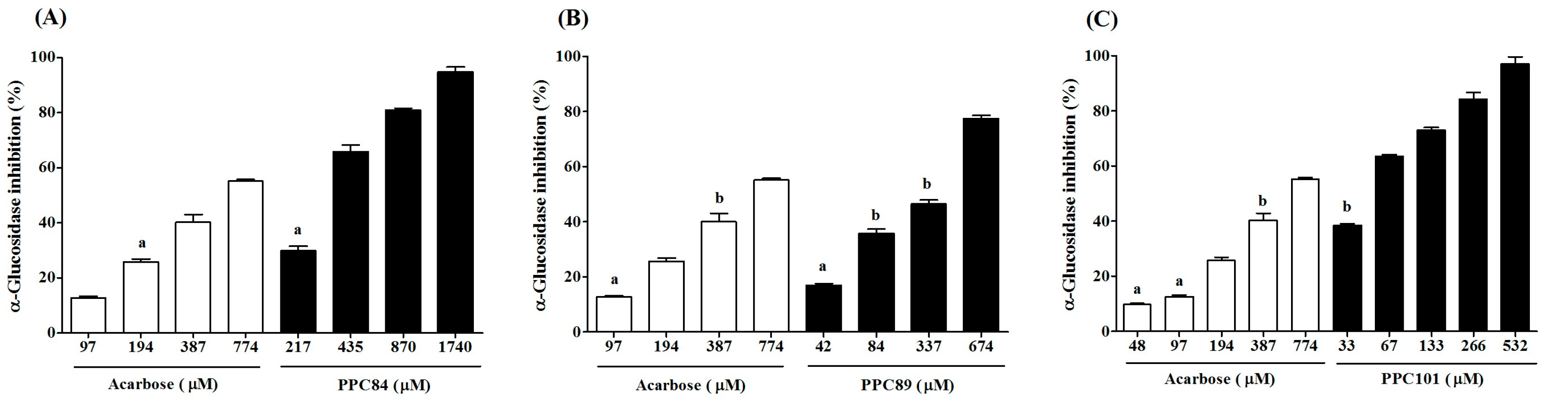
3.6. Inhibitory Effect of Amino Acid Derivatives on α-Glucosidase
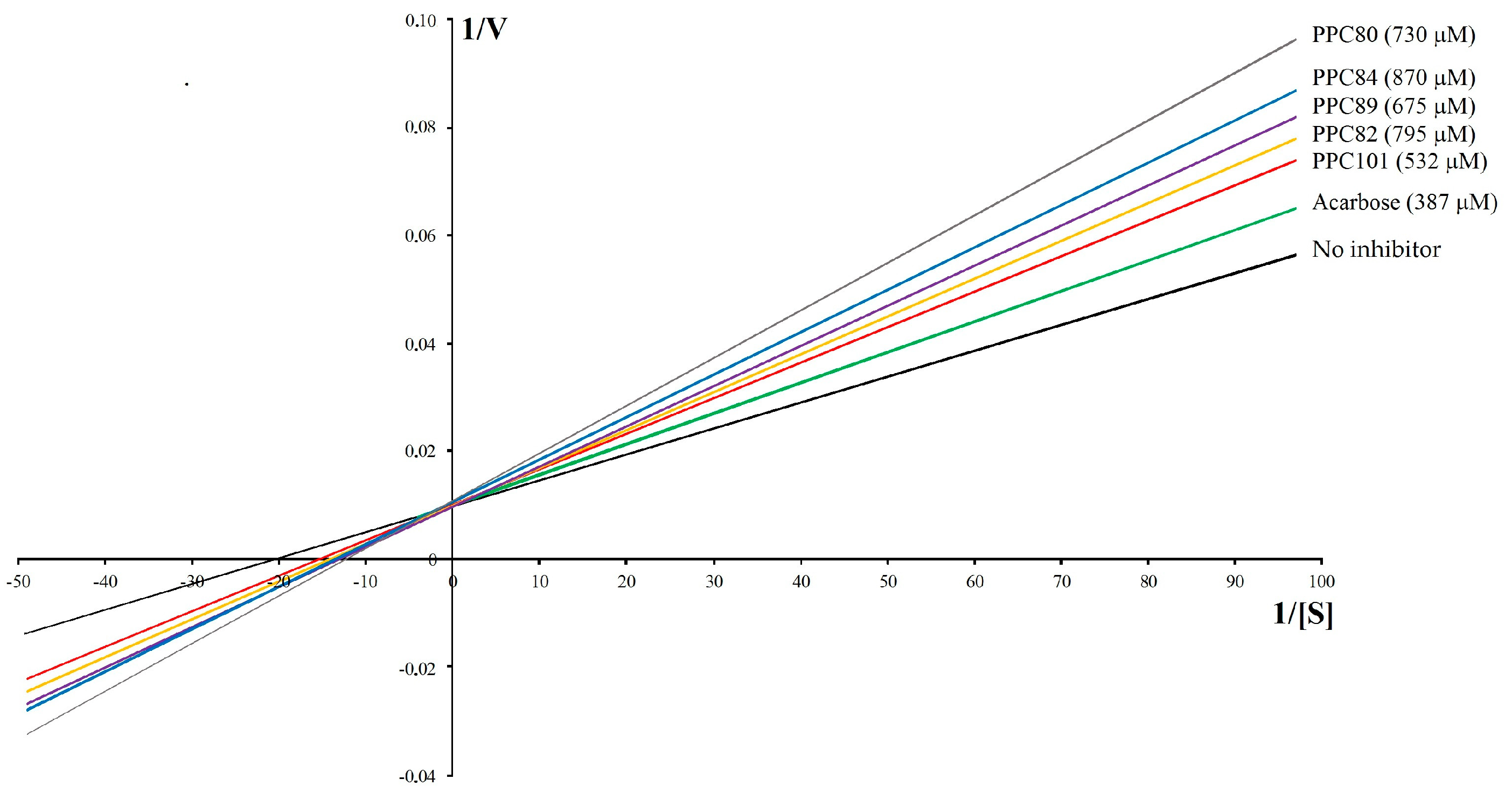
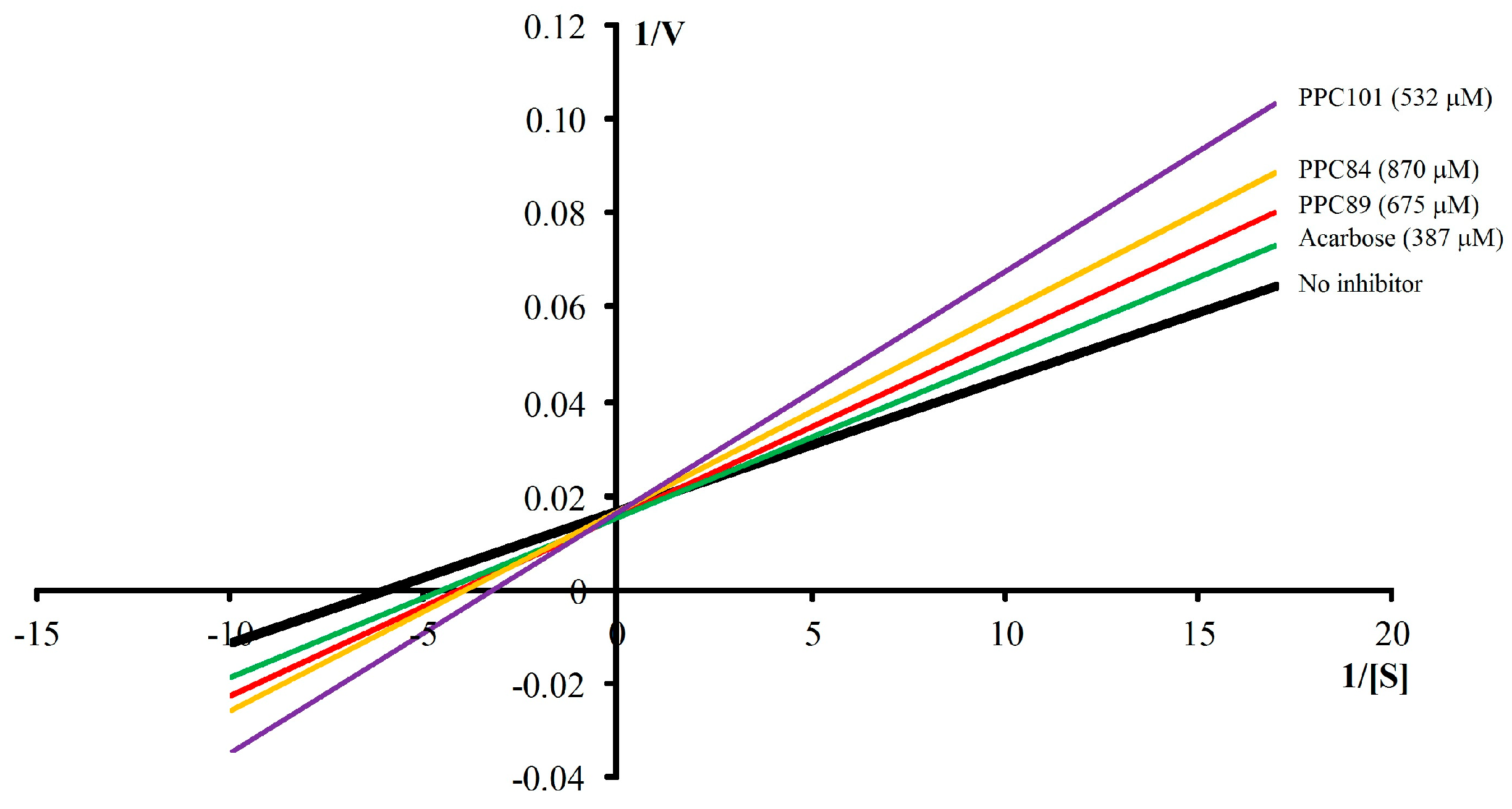
3.7. Kinetic Parameters of α-Glucosidase
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sensoy, I. A review on the food digestion in the digestive tract and the used in vitro models. Curr. Res. Food Sci. 2021, 4, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Soni, N.K.; Trivedi, H.H.; Kumar, S.; Prakash, A.; Roy, S.; Qamra, A.; Mukherjee, S. A review of digestive enzyme and probiotic supplementation for functional gastrointestinal disorders. Indian Pract. 2020, 73, 35–39. [Google Scholar]
- Hosseini, F.; Jayedi, A.; Khan, T.A.; Shab-Bidar, S. Dietary carbohydrate and the risk of type 2 diabetes: An updated systematic review and dose–response meta-analysis of prospective cohort studies. Sci. Rep. 2022, 12, 2491. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.-C.; Shamay, A.; Argov-Argaman, N. Lipid metabolism in mammary epithelial cells-A comparison of common in vitro models. Adv. Diary Res. 2018, 6, 1–9. [Google Scholar] [CrossRef]
- Saklayen, M.G. The global epidemic of the metabolic syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinicolantonio, J.J.; Bhutani, J.; O’keefe, J.H. Acarbose: Safe and effective for lowering postprandial hyperglycaemia and improving cardiovascular outcomes. Open Heart 2015, 2, e000327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoso, M.; Ong, L.L.; Aijijiyah, N.P.; Wati, F.A.; Azminah, A.; Annuur, R.M.; Fadlan, A.; Judeh, Z.M.A. Synthesis, α-glucosidase inhibition, α-amylase inhibition, and molecular docking studies of 3,3-di(indolyl)indolin-2-ones. Heliyon 2021, 8, e09045. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, K.; Sharma, A.; Kaur, K.; Kaur, K.; Chadha, R.; Bedi, P.M.S. Recent developments in synthetic α-glucosidase inhibitors: A comprehensive review with structural and molecular insight. J. Mol. Struct. 2023, 1281, 135115. [Google Scholar] [CrossRef]
- Lin, X.; Li, H. Obesity: Epidemiology, pathophysiology, and therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Liu, T.-T.; Liu, X.-T.; Chen, Q.-X.; Shi, Y. Lipase inhibitors for obesity: A review. Biomed. Pharmacother. 2020, 128, 110314. [Google Scholar] [CrossRef]
- Kushner, R.F. Weight loss strategies for treatment of obesity. Prog. Cardiovasc. Dis. 2014, 56, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Rodríguez, J.A.; Lévuok-Mena, K.P.; Cardozo-Muñoz, J.C.; Parra-Amin, J.E.; Lopez-Vallejo, F.; Cuca-Suárez, L.E.; Patiño-Ladino, O.J. In vitro and in silico study of the α-glucosidase and lipase inhibitory activities of chemical constituents from Piper cumanense (Piperaceae) and synthetic analogs. Plants 2022, 11, 2188. [Google Scholar] [CrossRef] [PubMed]
- Blaskovich, M.A.T. Unusual amino acids in medicinal chemistry. J. Med. Chem. 2016, 59, 10807–10836. [Google Scholar] [CrossRef] [PubMed]
- De Castro, P.P.; Campos, D.L.; Pavan, F.R.; Amarante, G.W. Dual-protected amino acid derivatives as new antitubercular agents. Chem. Biol. Drug. Des. 2018, 92, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- De Castro, P.P.; Siqueira, R.; Conforte, L.; Franco, C.; Bressan, G.; Amarante, G.W. Cytotoxic Activity of Synthetic Chiral Amino Acid Derivatives. J. Braz. Chem. Soc. 2020, 31, 193–200. [Google Scholar] [CrossRef]
- Oliva, R.; Chino, M.; Pane, K.; Pistorio, V.; Santis, A.; Pizzo, E.; D’errico, G.; Pavone, V.; Lombardi, A.; Vecchio, P.; et al. Exploring the role of unnatural amino acids in antimicrobial peptides. Sci. Rep. 2018, 8, 8888. [Google Scholar] [CrossRef] [Green Version]
- Castro, P.P.; Rimulo, I.M.R.; Almeida, A.M.; Diniz, R.; Amarante, G.W. Brønsted acid-catalyzed epimerization-free preparation of dual-protected amino acid derivatives. ACS Omega 2017, 2, 2967–2976. [Google Scholar] [CrossRef]
- Santos, B.C.S.; Pires, A.S.; Yamamoto, C.H.; Couri, M.R.C.; Taranto, A.G.; Alves, M.S.; Araújo, A.L.S.M.; Sousa, O.V. Methyl chavicol and its synthetic analogue as possible antioxidant and antilipase agents based on the in vitro and in silico assays. Oxid. Med. Cell. Longev. 2018, 2018, 2189348. [Google Scholar] [CrossRef] [Green Version]
- Freitas, T.C.; Oliveira, R.J.; Mendonça, R.J.; Candido, P.A.; Pereira, L.S.S.; Devienne, K.F.; Silva, A.C.; Pereira, C.A. Identification of bioactive compounds and analysis of inhibitory potential of the digestive enzymes from Syzygium sp. extracts. J. Chem. 2019, 2019, 3410953. [Google Scholar] [CrossRef] [Green Version]
- Chelladurai, G.R.M.; Chinnachamy, C. Alpha amylase and alpha glucosidase inhibitory effects of aqueous stem extract of Salacia oblonga and its GC-MS analysis. Braz. J. Pharm. Sci. 2018, 54, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Robin, T.; Reuveni, S.; Urbakh, M. Single-molecule theory of enzymatic inhibition. Nat. Commun. 2018, 9, 779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awosika, T.O.; Aluko, R.E. Inhibition of the in vitro activities of α-amylase, α-glucosidase and pancreatic lipase by yellow field pea (Pisum sativum L.) protein hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Fontes, R.; Ribeiro, J.M.; Sillero, A. Inhibition and activation of enzymes. The effect of a modifier on the reaction rate and on kinetic parameters. Acta Biochim. Pol. 2000, 47, 233–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngoh, Y.; Gan, C. Enzyme-assisted extraction and identification of antioxidativeanda-amylase inhibitory peptides from Pinto beans (Phaseolus vulgaris cv. Pinto). Food Chem. 2015, 190, 331–337. [Google Scholar] [CrossRef]
- Kido, Y.; Hiramoto, S.; Murao, M.; Horio, Y.; Toshiyuki, M.; Kodama, T.; Nokabou, Y. ε-Polylysine inhibits pancreatic lipase activity and suppresses postprandial hypertriacylglyceridemia in rats. J. Nutr. 2003, 133, 1887–1891. [Google Scholar] [CrossRef] [Green Version]
- Lunder, M.; Bratkovic, T.; Kreft, S.; Strukelj, B. Peptide inhibitor of pancreatic lipase selected by phage display using different elution strategies. J. Lip. Res. 2005, 46, 1512–1516. [Google Scholar] [CrossRef] [Green Version]
- Lunagariya, N.A.; Patel, N.K.; Jagtap, S.C.; Bhutani, K.K. Inhibitors of pancreatic lipase: State of the art and clinical perspectives. EXCLI J. 2014, 13, 897–921. [Google Scholar]
- Priyadharshini, A.; Ahalya, S.P.; Vaishnavi, P.; Pavithra, S.; Rosario, A.R. A review on benefits and toxicity of orlistat therapy. Drug Invent. Today 2019, 12, 550–553. [Google Scholar]
- Rani, K.; Rana, R.; Datt, S. Review on characteristics and application of amylases. Int. J. Microbiol. Bioinform. 2015, 5, 1–5. [Google Scholar]
- Butterworth, P.J.; Warren, F.J.; Ellis, P.R. Humana-amylase and starch digestion: An interesting marriage. Starch Stärke 2011, 63, 395–405. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, A.A.M.; Yang, R.; Zhao, W. Identification of bioactive peptides with α-amylase inhibitory potential from enzymatic protein hydrolysates of red seaweed (Porphyra spp). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef] [PubMed]
- González-Montoya, M.; Hernández-Ledesma, B.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Bioactive peptides from germinated soybean with anti-diabetic potential by inhibition of dipeptidyl peptidase-IV, α-amylase, and α-Glucosidase enzymes. Int. J. Mol. Sci. 2019, 19, 2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afifah, N.H.; Gan, C.-Y. Antioxidative and amylase inhibitor peptides from Basil Seeds. Int. J. Pept. Res. Ther. 2015, 22, 3–10. [Google Scholar] [CrossRef]
- Mohamed, E.A.H.; Siddiqui, M.J.A.; Ang, L.F.; Sadikun, A.; Chan, S.H.; Tan, S.C.; Asmawi, M.Z.; Yam, M.F. Potent α-glucosidase and α-amylase inhibitory activities of standardized 50% ethanolic extracts and sinensetin from Orthosiphon stamineus Benth as anti-diabetic mechanism. BMC Complement. Altern. Med. 2012, 12, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraj, S.; Suresh, S.; Kadeppagari, R.-K. Amylase inhibitors and their biomedical applications. Starch/Stärke 2013, 65, 535–542. [Google Scholar] [CrossRef]
- Bays, H.E.; Fitch, A.; Christensen, S.; Burridge, K.; Tondt, J. Anti-obesity medications and investigational agents: An Obesity Medicine Association (OMA) Clinical Practice Statement (CPS) 2022. Obesity Pillars 2022, 2, 100018. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Martínez-Villalueng, C.; Hernández-Ledesma, B. Release of dipeptidyl peptidase IV, α-amylase and α-glucosidase inhibitory peptides from quinoa (Chenopodium quinoa Willd.) during in vitro simulated gastrointestinal digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Kaur, A. Antidiabetic potential of a peptide isolated from an endophytic Aspergillus awamori. J. Appl. Microbiol. 2016, 120, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Yin, Y.; Zhao, W.; Liu, J.; Chen, F. Anti-diabetic activity peptides from albumin against α-glucosidase and α-amylase. Food Chem. 2012, 135, 2078–2085. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | IC50 (µM) | ||
|---|---|---|---|
| Pancreatic Lipase | Pancreatic α-Amylase | α-Glucosidase | |
| Orlistat | 587.70 ± 14.90 | - | - |
| Acarbose | - | 326.00 ± 3.21 a | 639.00 ± 4.62 |
| PPC80 | 475.30 ± 8.25 | 275.70 ± 5.21 a | - |
| PPC82 | 167.00 ± 6.25 | 519.00 ± 19.97 b | - |
| PPC84 | 1023.00 ± 20.34 | 493.00 ± 10.97 b | 321.30 ± 2.03 |
| PPC89 | - | 171.30 ± 13.57 c | 353.00 ± 6.03 |
| PPC101 | - | 162.00 ± 1.73 c | 51.00 ± 1.73 |
| Group | Concentration (µM) | Km (mM) | Vmax (µM/min) | Slope (min−1) |
|---|---|---|---|---|
| No inhibitor | - | 0.19 ± 0.006 a | 68.65 ± 0.41 a | 2.77 |
| Orlistat | 101 | 0.14 ± 0.001 | 68.34 ± 0.40 a | 4.10 |
| PPC80 | 146 | 0.16 ± 0.004 | 68.82 ± 0.57 a | 4.65 |
| PPC82 | 159 | 0.20 ± 0.004 a | 69.13 ± 0.57 a | 5.79 |
| PPC84 | 174 | 0.10 ± 0.003 | 62.76 ± 0.35 | 3.19 |
| Group | Concentration (µM) | Km (mM) | Vmax (µM/min) | Slope (min−1) |
|---|---|---|---|---|
| No inhibitor | - | 0.060 ± 0.006 | 100.70 ± 0.34 a | 0.59 |
| Acarbose | 387 | 0.033 ± 0.002 a | 100.30 ± 0.89 a | 0.66 |
| PPC80 | 730 | 0.041 ± 0.001 a | 91.75 ± 0.48 | 0.89 |
| PPC82 | 795 | 0.036 ± 0.001 a | 99.70 ± 1.20 a | 0.72 |
| PPC84 | 870 | 0.038 ± 0.003 a | 94.35 ± 0.51 | 0.80 |
| PPC89 | 675 | 0.038 ± 0.003 a | 99.73 ± 1.77 a | 0.76 |
| PPC101 | 532 | 0.035 ± 0.002 a | 100.00 ± 1.55 a | 0.70 |
| Group | Concentration (µM) | Km (mM) | Vmax (µM/min) | Slope (min−1) |
|---|---|---|---|---|
| No inhibitor | - | 0.183 ± 0.009 | 62.42 ± 1.14 a | 2.93 |
| Acarbose | 387 | 0.106 ± 0.003 | 63.35 ± 1.43 a | 3.35 |
| PPC84 | 870 | 0.135 ± 0.007 | 62.90 ± 0.60 a | 4.29 |
| PPC89 | 675 | 0.128 ± 0.001 | 62.30 ± 1.33 a | 4.11 |
| PPC101 | 532 | 0.156 ± 0.002 | 63.09 ± 1.45 a | 4.94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, F.C.d.; Santos, B.C.S.; Castro, P.P.d.; Amarante, G.W.; Sousa, O.V.d. Inhibitory Potential of Synthetic Amino Acid Derivatives against Digestive Enzymes as Promising Hypoglycemic and Anti-Obesity Agents. Biomolecules 2023, 13, 953. https://doi.org/10.3390/biom13060953
Silva FCd, Santos BCS, Castro PPd, Amarante GW, Sousa OVd. Inhibitory Potential of Synthetic Amino Acid Derivatives against Digestive Enzymes as Promising Hypoglycemic and Anti-Obesity Agents. Biomolecules. 2023; 13(6):953. https://doi.org/10.3390/biom13060953
Chicago/Turabian StyleSilva, Franciane Campos da, Bruna Celeida Silva Santos, Pedro Pôssa de Castro, Giovanni Wilson Amarante, and Orlando Vieira de Sousa. 2023. "Inhibitory Potential of Synthetic Amino Acid Derivatives against Digestive Enzymes as Promising Hypoglycemic and Anti-Obesity Agents" Biomolecules 13, no. 6: 953. https://doi.org/10.3390/biom13060953