
Anti-TNF Biologicals Enhance the Anti-Inflammatory Properties of IgG N-Glycome in Crohn’s Disease
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Samples and Ethical Considerations
2.2. IgG N-Glycan Sample Preparation
2.3. HILIC-UHPLC-FLD Analysis of 2-AB Labelled IgG N-Glycans
2.4. Statistical Analysis
3. Results
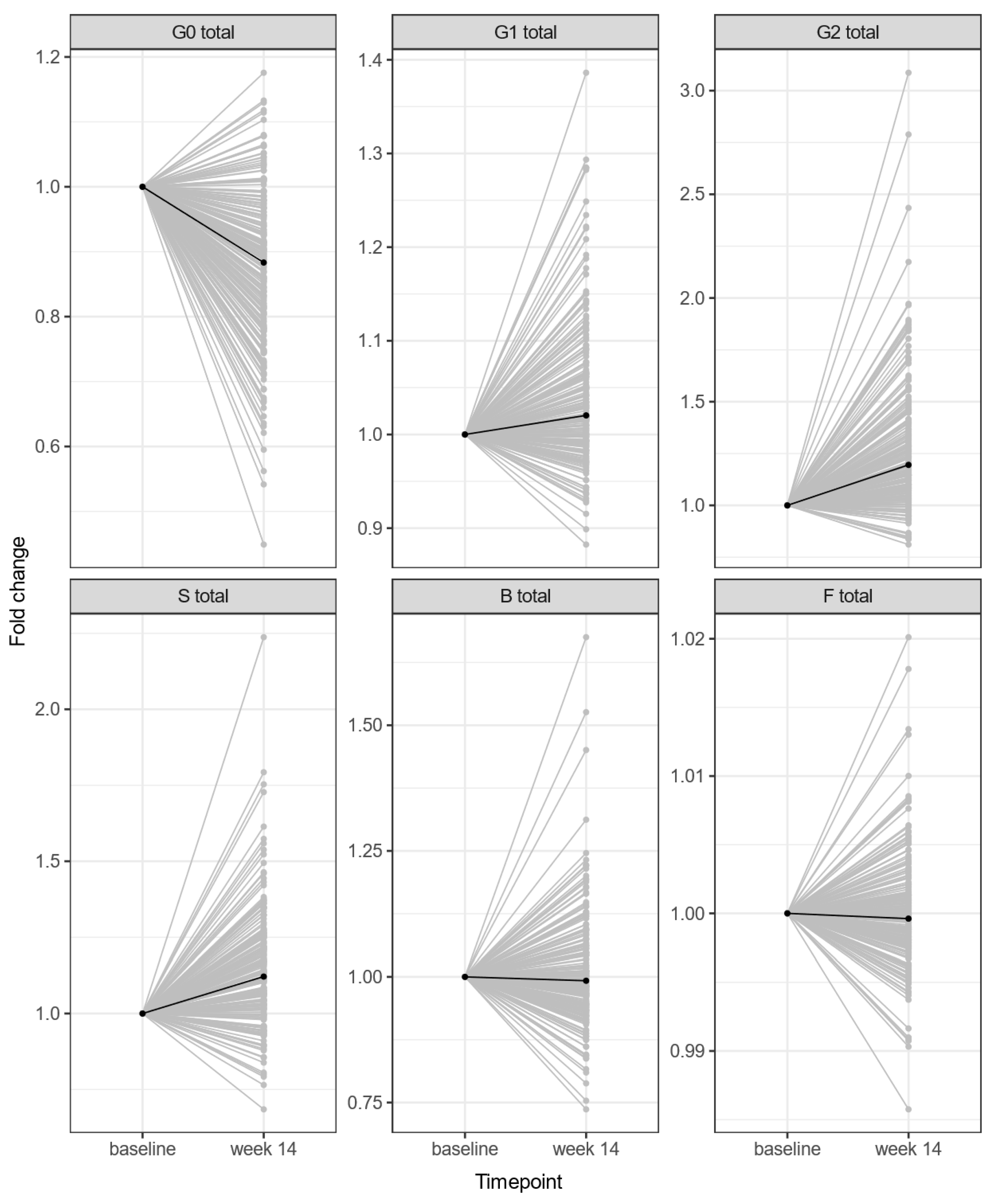
3.1. Anti-TNF Therapy Changes the IgG N-Glycome Composition of Crohn’s Disease Patients
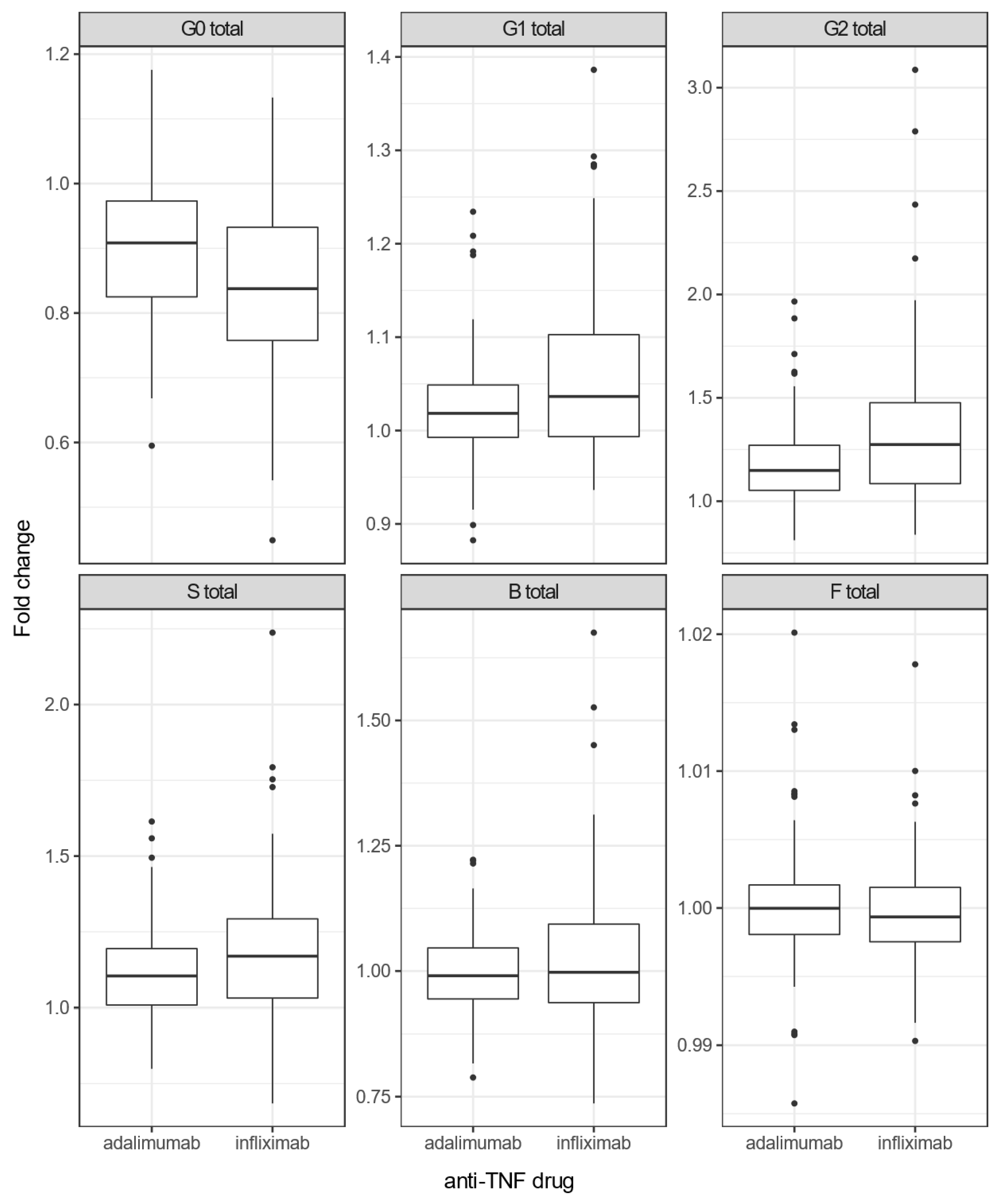
3.2. Infliximab Causes More Pronounced Changes in IgG N-Glycome than Adalimumab in Crohn’s Disease Patients
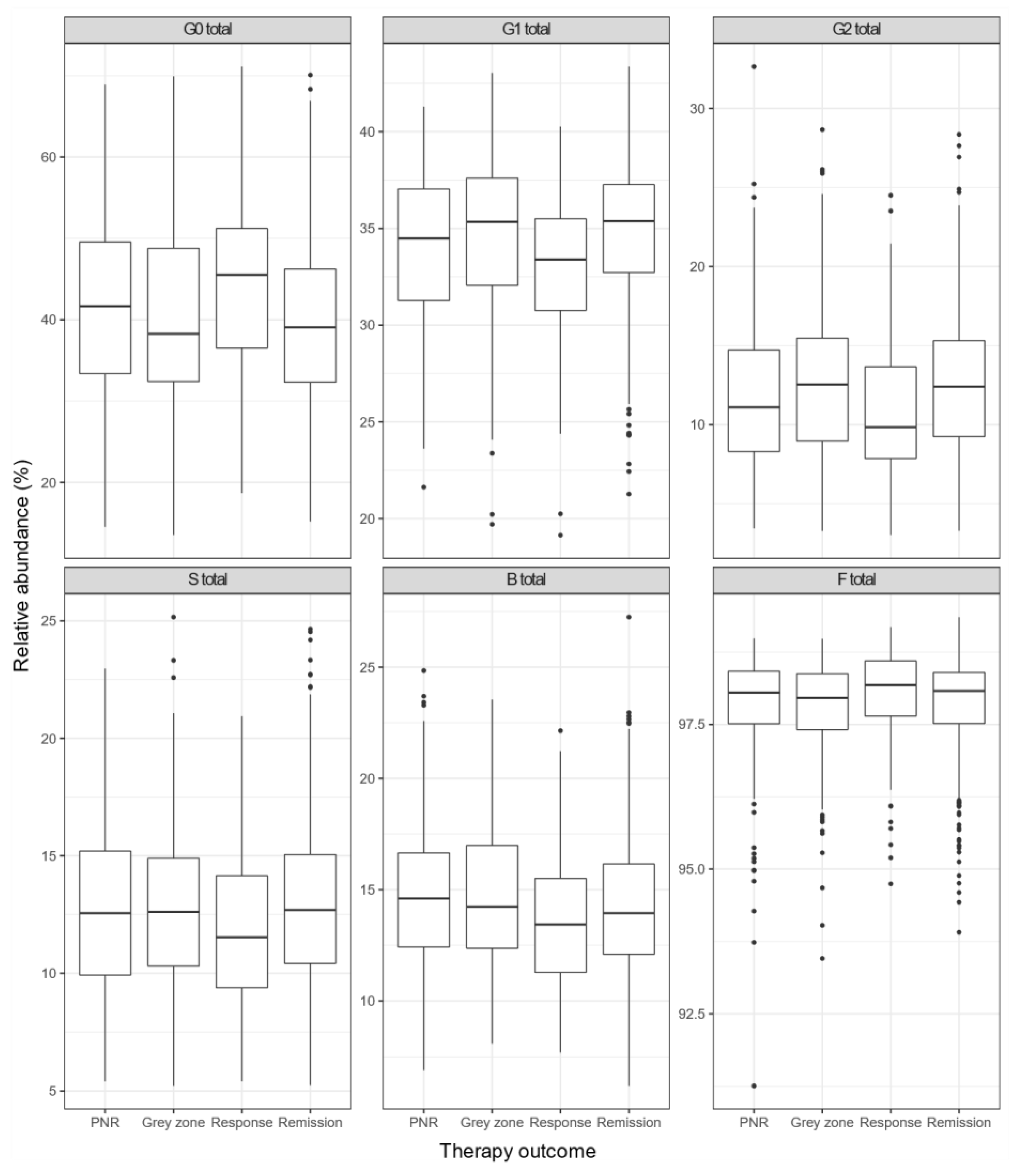
3.3. Composition of IgG N-Glycome Differs between Groups with Different Anti-TNF Therapy Outcomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2017 Inflammatory Bowel Disease Collaborators The Global, Regional, and National Burden of Inflammatory Bowel Disease in 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2020, 5, 17–30. [CrossRef] [Green Version]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide Incidence and Prevalence of Inflammatory Bowel Disease in the 21st Century: A Systematic Review of Population-Based Studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Peyrin-Biroulet, L.; Eisenhut, M.; Shin, J.I. IBD Immunopathogenesis: A Comprehensive Review of Inflammatory Molecules. Autoimmun. Rev. 2017, 16, 416–426. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N.; Bernstein, C.N.; Iliopoulos, D.; Macpherson, A.; Neurath, M.F.; Ali, R.A.R.; Vavricka, S.R.; Fiocchi, C. Environmental Triggers in IBD: A Review of Progress and Evidence. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, L.; Xie, R.; Wang, B.; Jiang, K.; Cao, H. Stress Triggers Flare of Inflammatory Bowel Disease in Children and Adults. Front. Pediatr. 2019, 7, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirkov, M.U.; Verstockt, B.; Cleynen, I. Genetics of Inflammatory Bowel Disease: Beyond NOD2. Lancet Gastroenterol. Hepatol. 2017, 2, 224–234. [Google Scholar] [CrossRef]
- Theodoratou, E.; Campbell, H.; Ventham, N.T.; Kolarich, D.; Pučić-Baković, M.; Zoldoš, V.; Fernandes, D.; Pemberton, I.K.; Rudan, I.; Kennedy, N.A.; et al. The Role of Glycosylation in IBD. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 588–600. [Google Scholar] [CrossRef] [PubMed]
- Hanić, M.; Trbojević-Akmačić, I.; Lauc, G. Inflammatory Bowel Disease—Glycomics Perspective. Biochim. Biophys. Acta-Gen. Subj. 2019, 1863, 1595–1601. [Google Scholar] [CrossRef]
- Verhelst, X.; Dias, A.M.; Colombel, J.-F.; Vermeire, S.; Van Vlierberghe, H.; Callewaert, N.; Pinho, S.S. Protein Glycosylation as a Diagnostic and Prognostic Marker of Chronic Inflammatory Gastrointestinal and Liver Diseases. Gastroenterology 2020, 158, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Gudelj, I.; Lauc, G.; Pezer, M. Immunoglobulin G Glycosylation in Aging and Diseases. Cell. Immunol. 2018, 333, 65–79. [Google Scholar] [CrossRef]
- Radovani, B.; Gudelj, I. N-Glycosylation and Inflammation; the Not-So-Sweet Relation. Front. Immunol. 2022, 13, 893365. [Google Scholar] [CrossRef]
- Li, T.; DiLillo, D.J.; Bournazos, S.; Giddens, J.P.; Ravetch, J.V.; Wang, L.X. Modulating IgG Effector Function by Fc Glycan Engineering. Proc. Natl. Acad. Sci. USA 2017, 114, 3485–3490. [Google Scholar] [CrossRef] [Green Version]
- Quast, I.; Peschke, B.; Lünemann, J.D. Regulation of Antibody Effector Functions through IgG Fc N-Glycosylation. Cell. Mol. Life Sci. 2017, 74, 837–847. [Google Scholar] [CrossRef] [PubMed]
- van de Bovenkamp, F.S.; Hafkenscheid, L.; Rispens, T.; Rombouts, Y. The Emerging Importance of IgG Fab Glycosylation in Immunity. J. Immunol. 2016, 196, 1435–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trbojevic Akmacic, I.; Ventham, N.T.; Theodoratou, E.; Vučković, F.; Kennedy, N.A.; Krištić, J.; Nimmo, E.R.; Kalla, R.; Drummond, H.; Štambuk, J.; et al. Inflammatory Bowel Disease Associates with Proinflammatory Potential of the Immunoglobulin G Glycome. Inflamm. Bowel Dis. 2015, 21, 1237–1247. [Google Scholar] [CrossRef] [Green Version]
- Šimurina, M.; de Haan, N.; Vučković, F.; Kennedy, N.A.; Štambuk, J.; Falck, D.; Trbojević-Akmačić, I.; Clerc, F.; Razdorov, G.; Khon, A.; et al. Glycosylation of Immunoglobulin G Associates with Clinical Features of Inflammatory Bowel Diseases. Gastroenterology 2018, 154, 1320–1333. [Google Scholar] [CrossRef]
- Adegbola, S.O.; Sahnan, K.; Warusavitarne, J.; Hart, A.; Tozer, P. Anti-TNF Therapy in Crohn’s Disease. Int. J. Mol. Sci. 2018, 19, 2244. [Google Scholar] [CrossRef] [Green Version]
- Cushing, K.; Higgins, P.D.R. Management of Crohn Disease: A Review. JAMA 2021, 325, 69. [Google Scholar] [CrossRef] [PubMed]
- Melsheimer, R.; Geldhof, A.; Apaolaza, I.; Schaible, T. Remicade® (Infliximab): 20 Years of Contributions to Science and Medicine. Biologics 2019, 13, 139–178. [Google Scholar] [CrossRef] [Green Version]
- Mitoma, H.; Horiuchi, T.; Tsukamoto, H.; Ueda, N. Molecular Mechanisms of Action of Anti-TNF-α Agents—Comparison among Therapeutic TNF-α Antagonists. Cytokine 2018, 101, 56–63. [Google Scholar] [CrossRef]
- Lee, C.; Jeong, M.; Lee, J.A.J.; Seo, S.; Cho, S.C.; Zhang, W.; Jaquez, O. Glycosylation Profile and Biological Activity of Remicade® Compared with Flixabi® and Remsima®. mAbs 2017, 9, 968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tebbey, P.W.; Varga, A.; Naill, M.; Clewell, J.; Venema, J. Consistency of Quality Attributes for the Glycosylated Monoclonal Antibody Humira® (Adalimumab). mAbs 2015, 7, 805. [Google Scholar] [CrossRef] [PubMed]
- Štambuk, J.; Vučković, F.; Habazin, S.; Hanić, M.; Novokmet, M.; Nikolaus, S.; Tran, F.; Schreiber, S.; Franke, A.; Rosenstiel, P.; et al. Distinct Longitudinal Changes in Immunoglobulin G N-Glycosylation Associate with Therapy Response in Chronic Inflammatory Diseases. Int. J. Mol. Sci. 2022, 23, 8473. [Google Scholar] [CrossRef] [PubMed]
- Collins, E.S.; Galligan, M.C.; Saldova, R.; Adamczyk, B.; Abrahams, J.L.; Campbell, M.P.; Ng, C.-T.; Veale, D.J.; Murphy, T.B.; Rudd, P.M.; et al. Glycosylation Status of Serum in Inflammatory Arthritis in Response to Anti-TNF Treatment. Rheumatology 2013, 52, 1572–1582. [Google Scholar] [CrossRef] [Green Version]
- Atreya, R.; Neurath, M.F.; Siegmund, B. Personalizing Treatment in IBD: Hype or Reality in 2020? Can We Predict Response to Anti-TNF? Front. Med. 2020, 7, 517. [Google Scholar] [CrossRef]
- Roda, G.; Jharap, B.; Neeraj, N.; Colombel, J.-F. Loss of Response to Anti-TNFs: Definition, Epidemiology, and Management. Clin. Transl. Gastroenterol. 2016, 7, e135. [Google Scholar] [CrossRef]
- Pereira, M.S.; Maia, L.; Azevedo, L.F.; Campos, S.; Carvalho, S.; Dias, A.M.; Albergaria, A.; Lima, J.; Marcos-Pinto, R.; Lago, P.; et al. A [Glyco]Biomarker That Predicts Failure to Standard Therapy in Ulcerative Colitis Patients. J. Crohn’s Colitis 2019, 13, 39–49. [Google Scholar] [CrossRef]
- Trbojević-Akmačić, I.; Ugrina, I.; Lauc, G. Comparative Analysis and Validation of Different Steps in Glycomics Studies. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2017; Volume 586, pp. 37–55. [Google Scholar]
- Hanić, M.; Lauc, G.; Trbojević-Akmačić, I. N-Glycan Analysis by Ultra-Performance Liquid Chromatography and Capillary Gel Electrophoresis with Fluorescent Labeling. Curr. Protoc. Protein Sci. 2019, 97, e95. [Google Scholar] [CrossRef]
- Agakova, A.; Vučković, F.; Klarić, L.; Lauc, G.; Agakov, F. Automated Integration of a UPLC Glycomic Profile. Methods Mol. Biol. 2017, 1503, 217–233. [Google Scholar] [CrossRef]
- Pučić, M.; Knežević, A.; Vidič, J.; Adamczyk, B.; Novokmet, M.; Polašek, O.; Gornik, O.; Šupraha-Goreta, S.; Wormald, M.R.; Redžić, I.; et al. High Throughput Isolation and Glycosylation Analysis of IgG–Variability and Heritability of the IgG Glycome in Three Isolated Human Populations. Mol. Cell. Proteom. 2011, 10, M111.010090. [Google Scholar] [CrossRef] [Green Version]
- Torres, J.; Bonovas, S.; Doherty, G.; Kucharzik, T.; Gisbert, J.P.; Raine, T.; Adamina, M.; Armuzzi, A.; Bachmann, O.; Bager, P.; et al. ECCO Guidelines on Therapeutics in Crohn’s Disease: Medical Treatment. J. Crohn’s Colitis 2020, 14, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, N.A.; Heap, G.A.; Green, H.D.; Hamilton, B.; Bewshea, C.; Walker, G.J.; Thomas, A.; Nice, R.; Perry, M.H.; Bouri, S.; et al. Predictors of Anti-TNF Treatment Failure in Anti-TNF-Naive Patients with Active Luminal Crohn’s Disease: A Prospective, Multicentre, Cohort Study. Lancet Gastroenterol. Hepatol. 2019, 4, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Shinzaki, S.; Iijima, H.; Nakagawa, T.; Egawa, S.; Nakajima, S.; Ishii, S.; Irie, T.; Kakiuchi, Y.; Nishida, T.; Yasumaru, M.; et al. IgG Oligosaccharide Alterations Are a Novel Diagnostic Marker for Disease Activity and the Clinical Course of Inflammatory Bowel Disease. Am. J. Gastroenterol. 2008, 103, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Peyrin-Biroulet, L.; Lémann, M. Review Article: Remission Rates Achievable by Current Therapies for Inflammatory Bowel Disease. Aliment. Pharmacol. Ther. 2011, 33, 870–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Fumery, M.; Sandborn, W.J.; Murad, M.H. Systematic Review and Network Meta-Analysis: First- and Second-Line Biologic Therapies for Moderate-Severe Crohn’s Disease. Aliment. Pharmacol. Ther. 2018, 48, 394–409. [Google Scholar] [CrossRef]
- Thorlund, K.; Druyts, E.; Mills, E.J.; Fedorak, R.N.; Marshall, J.K. Adalimumab versus Infliximab for the Treatment of Moderate to Severe Ulcerative Colitis in Adult Patients Naïve to Anti-TNF Therapy: An Indirect Treatment Comparison Meta-Analysis. J. Crohns Colitis 2014, 8, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croce, A.; Firuzi, O.; Altieri, F.; Eufemi, M.; Agostino, R.; Priori, R.; Bombardieri, M.; Alessandri, C.; Valesini, G.; Saso, L. Effect of Infliximab on the Glycosylation of IgG of Patients with Rheumatoid Arthritis. J. Clin. Lab. Anal. 2007, 21, 303–314. [Google Scholar] [CrossRef]
- Van de Geijn, F.E.; Wuhrer, M.; Selman, M.H.J.; Willemsen, S.P.; de Man, Y.A.; Deelder, A.M.; Hazes, J.M.W.; Dolhain, R.J.E.M. Immunoglobulin G Galactosylation and Sialylation Are Associated with Pregnancy-Induced Improvement of Rheumatoid Arthritis and the Postpartum Flare: Results from a Large Prospective Cohort Study. Arthritis Res. Ther. 2009, 11, R193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondt, A.; Selman, M.H.J.; Deelder, A.M.; Hazes, J.M.W.; Willemsen, S.P.; Wuhrer, M.; Dolhain, R.J.E.M. Association between Galactosylation of Immunoglobulin G and Improvement of Rheumatoid Arthritis during Pregnancy Is Independent of Sialylation. J. Proteome Res. 2013, 12, 4522–4531. [Google Scholar] [CrossRef]
- Quast, I.; Keller, C.W.; Maurer, M.A.; Giddens, J.P.; Tackenberg, B.; Wang, L.-X.; Münz, C.; Nimmerjahn, F.; Dalakas, M.C.; Lünemann, J.D. Sialylation of IgG Fc Domain Impairs Complement-Dependent Cytotoxicity. J. Clin. Investig. 2015, 125, 4160–4170. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Week 0 (Baseline) | Week 14 | |
|---|---|---|
| Number of samples (N) | 1315 | 198 |
| Median age at first dose [IQR], yr | 33 [23–47] | 33 [24–46] |
| Number of females (%) | 673 (51.2) | 86 (43.4) |
| Median disease duration at first dose [IQR], yr | 2.5 [0.7–9.0] | 2.0 [0.6–10.1] |
| Anti-TNFα treatment (n) | ||
| Infliximab (IFX) | 820 | 100 |
| Adalimumab (ADA) | 495 | 98 |
| Therapy outcome (n) | ||
| Primary non-response (PNR) | 258 | 33 |
| Response | 180 | 165 |
| Grey zone | 231 | / |
| Remission | 515 | / |
| N/A | 131 | / |
| Derived Trait | Effect | Standard Error | p-Value | Adjusted p-Value |
|---|---|---|---|---|
| G0 | −0.55 | 0.04 | 1.34 × 10−29 | 4.03 × 10−29 |
| G1 | 0.35 | 0.05 | 1.74 × 10−12 | 2.61 × 10−12 |
| G2 | 0.60 | 0.04 | 1.51 × 10−32 | 9.07 × 10−32 |
| S | 0.46 | 0.04 | 1.88 × 10−21 | 3.75 × 10−21 |
| B | 0.02 | 0.04 | 6.25 × 10−1 | 6.25 × 10−1 |
| F | −0.02 | 0.04 | 5.77 × 10−1 | 6.25 × 10−1 |
| Derived Trait | Effect | Standard Error | p-Value | Adjusted p-Value |
|---|---|---|---|---|
| G0 | −0.24 | 0.08 | 1.73 × 10−3 | 3.46 × 10−3 |
| G1 | 0.28 | 0.09 | 1.08 × 10−3 | 3.23 × 10−3 |
| G2 | 0.26 | 0.08 | 8.85 × 10−4 | 3.23 × 10−3 |
| S | 0.18 | 0.08 | 2.34 × 10−2 | 3.51 × 10−2 |
| B | 0.05 | 0.07 | 4.88 × 10−1 | 4.88 × 10−1 |
| F | −0.06 | 0.08 | 4.73 × 10−1 | 4.88 × 10−1 |
| Derived Trait | p-Value | Adjusted p-Value |
|---|---|---|
| G0 | 9.81 × 10−8 | 1.96 × 10−7 |
| G1 | 2.95 × 10−8 | 1.77 × 10−7 |
| G2 | 7.98 × 10−8 | 1.96 × 10−7 |
| S | 1.31 × 10−3 | 1.97 × 10−3 |
| B | 9.16 × 10−2 | 9.16 × 10−2 |
| F | 7.41 × 10−2 | 8.89 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanić, M.; Vučković, F.; Deriš, H.; Bewshea, C.; Lin, S.; Goodhand, J.R.; Ahmad, T.; Trbojević-Akmačić, I.; Kennedy, N.A.; Lauc, G.; et al. Anti-TNF Biologicals Enhance the Anti-Inflammatory Properties of IgG N-Glycome in Crohn’s Disease. Biomolecules 2023, 13, 954. https://doi.org/10.3390/biom13060954
Hanić M, Vučković F, Deriš H, Bewshea C, Lin S, Goodhand JR, Ahmad T, Trbojević-Akmačić I, Kennedy NA, Lauc G, et al. Anti-TNF Biologicals Enhance the Anti-Inflammatory Properties of IgG N-Glycome in Crohn’s Disease. Biomolecules. 2023; 13(6):954. https://doi.org/10.3390/biom13060954
Chicago/Turabian StyleHanić, Maja, Frano Vučković, Helena Deriš, Claire Bewshea, Simeng Lin, James R. Goodhand, Tariq Ahmad, Irena Trbojević-Akmačić, Nicholas A. Kennedy, Gordan Lauc, and et al. 2023. "Anti-TNF Biologicals Enhance the Anti-Inflammatory Properties of IgG N-Glycome in Crohn’s Disease" Biomolecules 13, no. 6: 954. https://doi.org/10.3390/biom13060954