

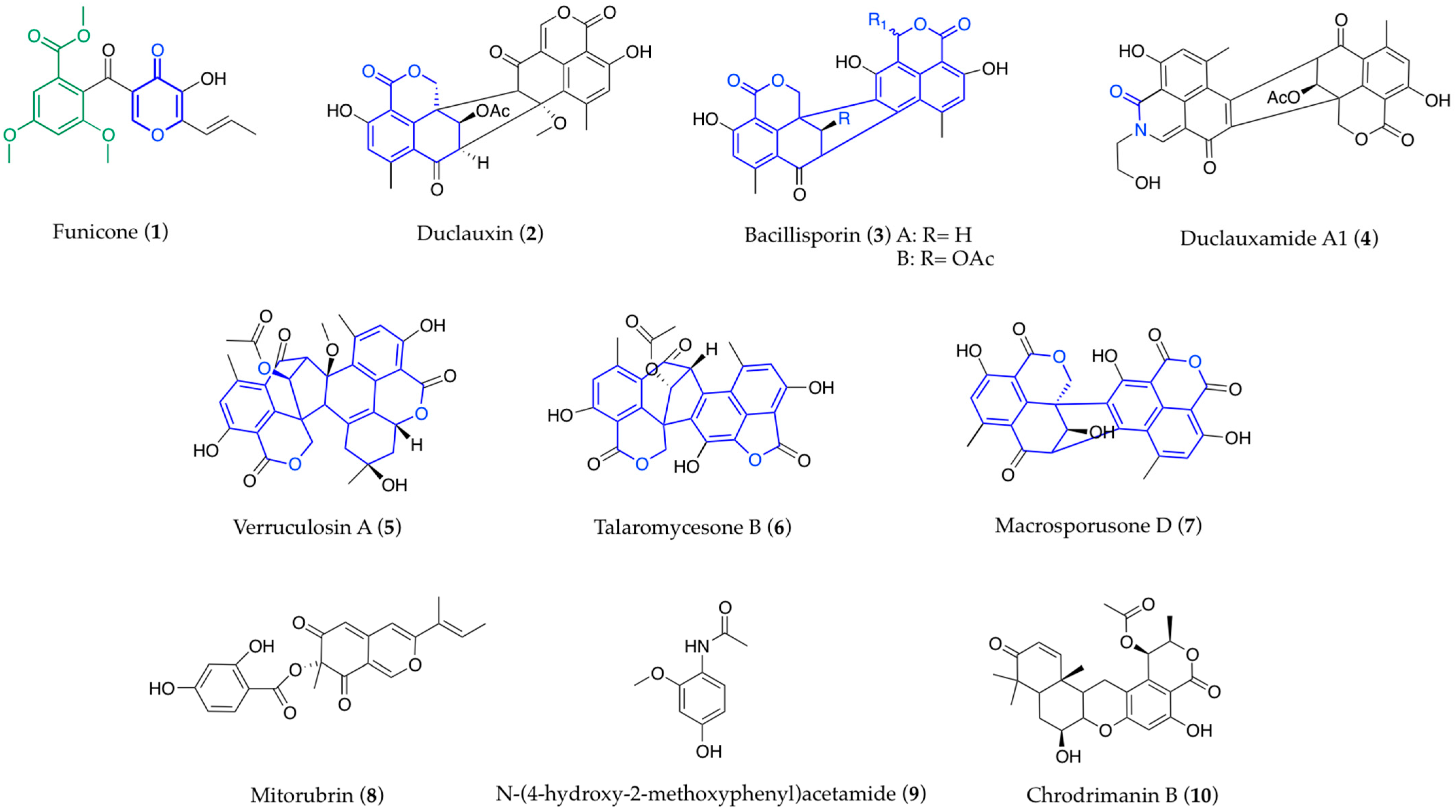
The Outstanding Chemodiversity of Marine-Derived Talaromyces


Abstract
:1. Introduction
2. Occurrence of Talaromyces in the Marine Environment
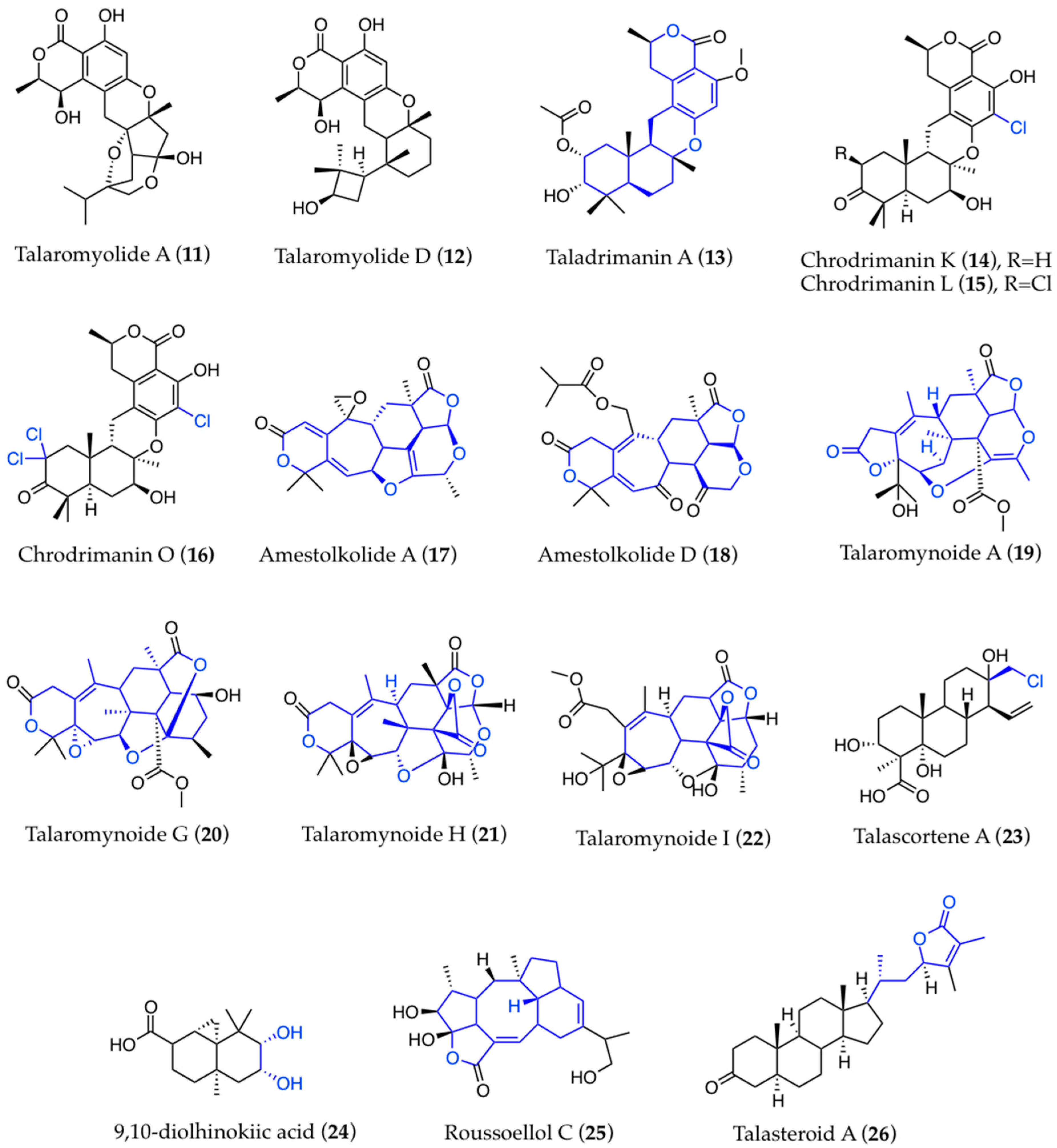
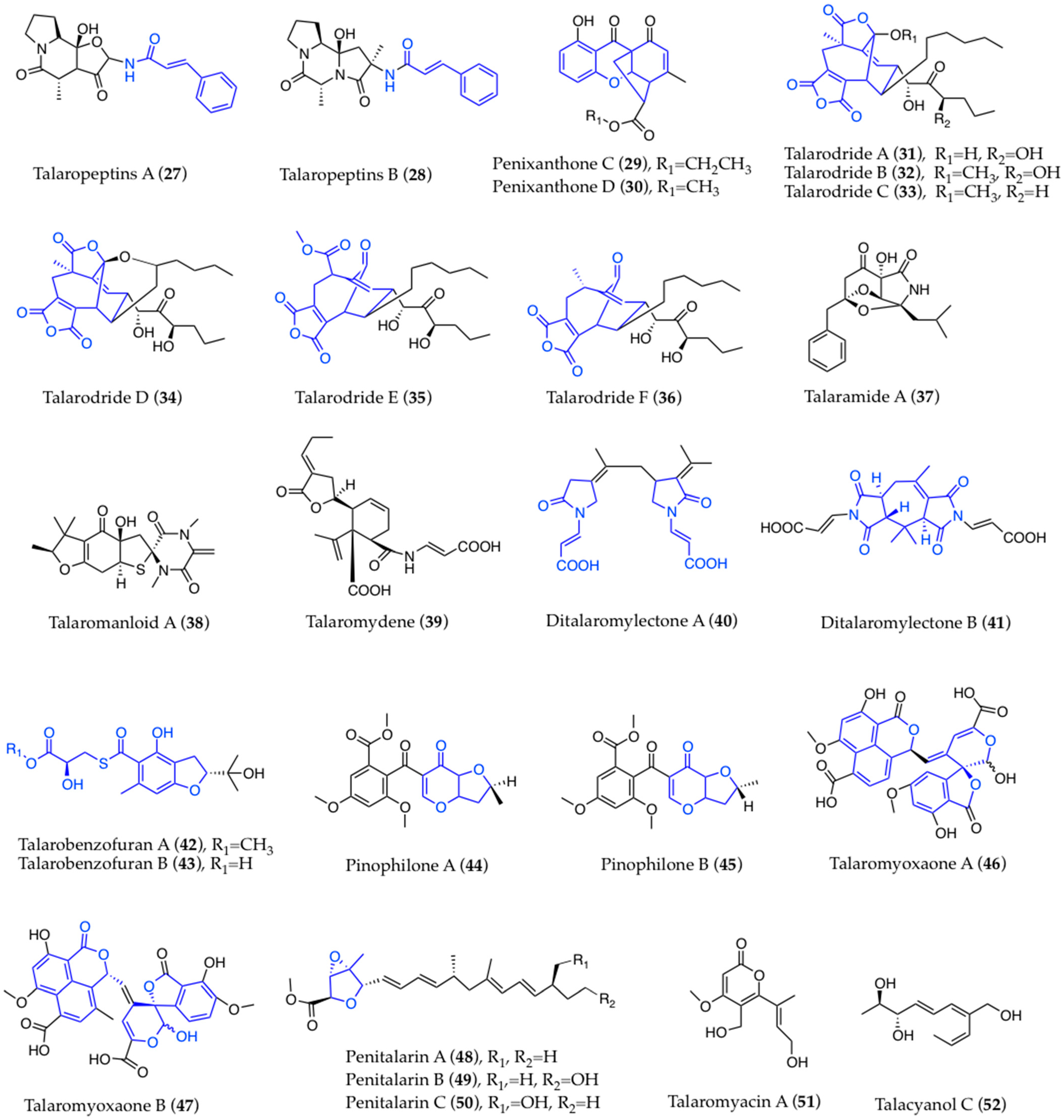
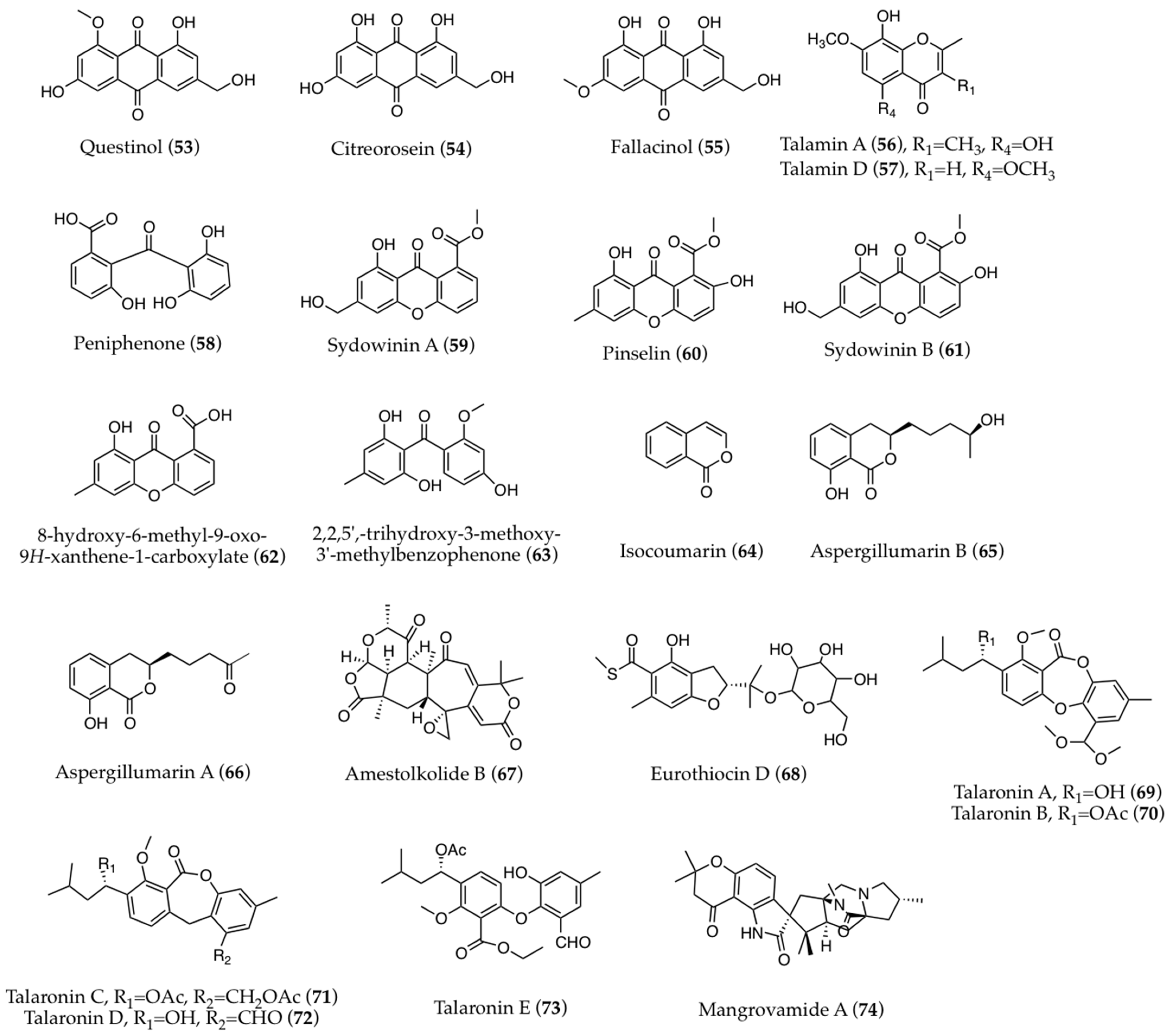
3. Structural Aspects
3.1. Biogenesis and Structure-Activity Relationships
3.2. Other Biological Sources of the Known Compounds
4. Biological Properties
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benjamin, C.R. Ascocarps of Aspergillus and Penicillium. Mycologia 1955, 47, 669–687. [Google Scholar] [CrossRef]
- Adams, P.B. The potential of mycoparasites for biological control of plant diseases. Ann. Rev. Phytopathol. 1990, 28, 59–72. [Google Scholar] [CrossRef]
- Nicoletti, R.; Andolfi, A.; Salvatore, M.M. Endophytic fungi of the genus Talaromyces and plant health. In Microbial Endophytes and Plant Growth; Solanki, M.K., Yadav, M.K., Singh, B.P., Gupta, V.K., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 183–213. [Google Scholar]
- Nicoletti, R.; Becchimanzi, A. Talaromyces–insect relationships. Microorganisms 2022, 10, 45. [Google Scholar] [CrossRef]
- Kempken, F.; Rohlfs, M. Fungal secondary metabolite biosynthesis–a chemical defence strategy against antagonistic animals? Fungal Ecol. 2010, 3, 107–114. [Google Scholar] [CrossRef]
- Devi, R.; Kaur, T.; Kour, D.; Rana, K.L.; Yadav, A.; Yadav, A.N. Beneficial fungal communities from different habitats and their roles in plant growth promotion and soil health. Microb. Biosyst. 2020, 5, 21–47. [Google Scholar] [CrossRef]
- Frisvad, J.C. Taxonomy, chemodiversity, and chemoconsistency of Aspergillus, Penicillium, and Talaromyces species. Front. Microbiol. 2015, 5, 773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, M.M.; Li, J.; Jiang, C.X.; Shi, Y.P.; Di, D.L.; Crews, P.; Wu, Q.X. The bioactive secondary metabolites from Talaromyces species. Nat. Prod. Bioprosp. 2016, 6, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Nicoletti, R.; Salvatore, M.M.; Andolfi, A. Secondary metabolites of mangrove-associated strains of Talaromyces. Mar. Drugs 2018, 16, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proksa, B. Talaromyces flavus and its metabolites. Chem. Pap. 2010, 64, 696–714. [Google Scholar] [CrossRef]
- Nicoletti, R.; Trincone, A. Bioactive compounds produced by strains of Penicillium and Talaromyces of marine origin. Mar. Drugs 2016, 14, 37. [Google Scholar] [CrossRef] [Green Version]
- Pang, K.L.; Overy, D.P.; Jones, E.G.; da Luz Calado, M.; Burgaud, G.; Walker, A.K.; Johnson, J.A.; Kerr, R.G.; Cha, H.J.; Bills, G.F. ‘Marine fungi’ and ‘marine-derived fungi’ in natural product chemistry research: Toward a new consensual definition. Fungal Biol. Rev. 2016, 30, 163–175. [Google Scholar] [CrossRef]
- Gladfelter, A.S.; James, T.Y.; Amend, A.S. Marine fungi. Curr. Biol. 2019, 29, R191–R195. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef] [Green Version]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 96, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Han, P.J.; Sun, J.Q.; Wang, L. Two new sexual Talaromyces species discovered in estuary soil in China. J. Fungi 2022, 8, 36. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.; Liu, W.; Liu, Y.; Huang, X.; She, Z. Polyketides with immunosuppressive activities from mangrove endophytic fungus Penicillium sp. ZJ-SY2. Mar. Drugs 2016, 14, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Cui, C.B.; Li, C.W. A new cyclic dipeptide penicimutide: The activated production of cyclic dipeptides by introduction of neomycin-resistance in the marine-derived fungus Penicillium purpurogenum G59. Arch. Pharm. Res. 2016, 39, 762–770. [Google Scholar] [CrossRef]
- Wu, C.; Zhao, Y.; Chen, R.; Liu, D.; Liu, M.; Proksch, P.; Guo, P.; Lin, W. Phenolic metabolites from mangrove-associated Penicillium pinophilum fungus with lipid-lowering effects. RSC Adv. 2016, 6, 21969–21978. [Google Scholar] [CrossRef]
- Huang, H.; Liu, T.; Wu, X.; Guo, J.; Lan, X.; Zhu, Q.; Zheng, X.; Zhang, K. A new antibacterial chromone derivative from mangrove-derived fungus Penicillium aculeatum (No. 9EB). Nat. Prod. Res. 2017, 31, 2593–2598. [Google Scholar] [CrossRef]
- Kong, F.D.; Ma, Q.Y.; Huang, S.Z.; Wang, P.; Wang, J.F.; Zhou, L.M.; Yuan, J.Z.; Dai, H.F.; Zhao, Y.X. Chrodrimanins K–N and related meroterpenoids from the fungus Penicillium sp. SCS-KFD09 isolated from a marine worm, Sipunculus nudus. J. Nat. Prod. 2017, 80, 1039–1047. [Google Scholar] [CrossRef]
- Sumilat, D.A.; Yamazaki, H.; Endo, K.; Rotinsulu, H.; Wewengkang, D.S.; Ukai, K.; Namikoshi, M. A new biphenyl ether derivative produced by Indonesian ascidian-derived Penicillium albobiverticillium. J. Nat. Med. 2017, 71, 776–779. [Google Scholar] [CrossRef]
- Tao, H.; Wei, X.; Lin, X.; Zhou, X.; Dong, J.; Yang, B. Penixanthones A and B, two new xanthone derivatives from fungus Penicillium sp. SYFz-1 derived of mangrove soil sample. Nat. Prod. Res. 2017, 31, 2218–2222. [Google Scholar] [CrossRef]
- Zhang, Z.; He, X.; Zhang, G.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Inducing secondary metabolite production by combined culture of Talaromyces aculeatus and Penicillium variabile. J. Nat. Prod. 2017, 80, 3167–3171. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Tao, H.; Lin, X.; Wang, J.; Liao, S.; Dong, J.; Zhou, X.; Liu, Y. Prenylated indole alkaloids and chromone derivatives from the fungus Penicillium sp. SCSIO041218. Tetrahedron 2018, 74, 77–82. [Google Scholar] [CrossRef]
- He, F.; Li, X.; Yu, J.H.; Zhang, X.; Nong, X.; Chen, G.; Zhu, K.; Wang, Y.Y.; Bao, J.; Zhang, H. Secondary metabolites from the mangrove sediment-derived fungus Penicillium pinophilum SCAU037. Fitoterapia 2019, 136, 104177. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Ge, H.; Yi, W.; Wu, B.; Zhang, Z. Bioactive drimane sesquiterpenoids and isocoumarins from the marine-derived fungus Penicillium minioluteum ZZ1657. Tetr. Lett. 2020, 61, 151504. [Google Scholar] [CrossRef]
- Wang, B.; Chen, Q.H.; Jiang, T.; Cai, Y.W.; Huang, G.L.; Sun, X.P.; Zheng, C.J. Secondary metabolites from the mangrove-derived fungus Penicillium verruculosum and their bioactivities. Chem. Nat. Comp. 2021, 57, 588–591. [Google Scholar] [CrossRef]
- Hawas, U.W.; Abou El-Kassem, L.T.; Ahmed, E.F.; Alghamdi, R.A. Bioactive sulfonyl metabolites from the Red Sea endophytic fungus Penicillium aculeatum. Nat. Prod. Res. 2022, 36, 2713–2721. [Google Scholar] [CrossRef]
- Mei, R.Q.; Wang, B.; Zeng, W.N.; Huang, G.L.; Chen, G.Y.; Zheng, C.J. Bioactive isocoumarins isolated from a mangrove-derived fungus Penicillium sp. MGP11. Nat. Prod. Res. 2022, 36, 1260–1265. [Google Scholar] [CrossRef]
- Fouillaud, M.; Venkatachalam, M.; Llorente, M.; Magalon, H.; Cuet, P.; Dufossé, L. Biodiversity of pigmented fungi isolated from marine environment in La Réunion Island, Indian Ocean: New resources for colored metabolites. J. Fungi 2017, 3, 36. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, Y.; Liu, Z.; Cai, R.; Lu, Y.; Huang, X.; She, Z. Isocoumarins and benzofurans from the mangrove endophytic fungus Talaromyces amestolkiae possess α-glucosidase inhibitory and antibacterial activities. RSC Adv. 2016, 6, 26412–26420. [Google Scholar] [CrossRef]
- Huang, L.J.; Li, X.A.; Jin, M.Y.; Guo, W.X.; Lei, L.R.; Liu, R.; Zhang, M.Z.; Guo, D.L.; Wang, D.; Zhou, Y.; et al. Two previously undescribed phthalides from Talaromyces amestolkiae, a symbiotic fungus of Syngnathus acus. J. Asian Nat. Prod. Res. 2023, 25, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Zhou, X.M.; Yang, X.; Tang, M.M.; Liao, Q.Y.; Meng, B.Z.; Liao, S.; Chen, G.Y. Three new bioactive natural products from the fungus Talaromyces assiutensis JTY2. Bioorg. Chem. 2020, 94, 103362. [Google Scholar] [CrossRef]
- Mishra, R.C.; Kalra, R.; Dilawari, R.; Deshmukh, S.K.; Barrow, C.J.; Goel, M. Characterization of an endophytic strain Talaromyces assiutensis, CPEF04 with evaluation of production medium for extracellular red pigments having antimicrobial and anticancer properties. Front. Microbiol. 2021, 12, 665702. [Google Scholar] [CrossRef] [PubMed]
- Heydari, H.; Koc, A.; Simsek, D.; Gozcelioglu, B.; Altanlar, N.; Konuklugil, B. Isolation, identification and bioactivity screening of Turkish marine-derived fungi. Farmacia 2019, 67, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhou, L.; Wang, M.; Wei, L.; Qu, H.; Ma, J.; Ju, J.; Han, Z. Chemical constituents from marine derived fungus Talaromyces cellulolyticus SHJ-3 and its chemotaxonomic significance. Biochem. Syst. Ecol. 2022, 100, 104377. [Google Scholar] [CrossRef]
- Shin, H.J.; Anh, C.V.; Cho, D.Y.; Choi, D.K.; Kang, J.S.; Trinh, P.T.H.; Choi, B.K.; Lee, H.S. New polyenes from the marine-derived fungus Talaromyces cyanescens with anti-neuroinflammatory and cytotoxic activities. Molecules 2021, 26, 836. [Google Scholar] [CrossRef]
- Anand, B.G.; Thomas, C.N.; Prakash, S. In vitro cytotoxicity and antimicrobial activity of Talaromyces flavus SP5 inhabited in the marine sediment of Southern Coast of India. Chin. J. Nat. Med. 2016, 14, 913–921. [Google Scholar] [CrossRef]
- Cai, J.; Zhu, X.C.; Zeng, W.N.; Wang, B.; Luo, Y.P.; Liu, J.; Chen, M.J.; Li, G.Y.; Huang, G.L.; Chen, G.Y.; et al. Talaromarins A–F: Six new isocoumarins from mangrove-derived fungus Talaromyces flavus TGGP35. Mar. Drugs 2022, 20, 361. [Google Scholar] [CrossRef] [PubMed]
- Chalearmsrimuang, T.; Suasa-ard, S.; Jantasorn, A.; Dethoup, T. Effects of marine antagonistic fungi against plant pathogens and rice growth promotion activity. J. Pure Appl. Microbiol. 2022, 16, 402–419. [Google Scholar] [CrossRef]
- Liu, C.H.; Huang, X.; Xie, T.N.; Duan, N.; Xue, Y.R.; Zhao, T.X.; Lever, M.A.; Hinrichs, K.U.; Inagaki, F. Exploration of cultivable fungal communities in deep coal-bearing sediments from~ 1.3 to 2.5 km below the ocean floor. Environ. Microbiol. 2017, 19, 803–818. [Google Scholar] [CrossRef]
- Nie, Y.; Yang, W.; Liu, Y.; Yang, J.; Lei, X.; Gerwick, W.H.; Zhang, Y. Acetylcholinesterase inhibitors and antioxidants mining from marine fungi: Bioassays, bioactivity coupled LC–MS/MS analyses and molecular networking. Mar. Life Sci. Technol. 2020, 2, 386–397. [Google Scholar] [CrossRef]
- Zahri, M.Z.C.; Rehan, M.M.; Kamarudin, K.R.; Lufti, F.N.M.; Radzi, S.M.; Rehan, A.M. Checklist of marine fungi and yeasts associated with Holothuria (Mertensiothuria) leucospilota from Pangkor Island. J. Sust. Nat. Res. 2020, 1, 29–36. [Google Scholar]
- Wang, Y.; Li, X.; Lü, F.; Yang, S.; Meng, L.; Wang, B. Discovery of prenylated indole alkaloid and natural xanthone from cold-seep sediment derived fungus Talaromyces funiculosus SD-523. J. Oceanol. Limnol. 2023. [Google Scholar] [CrossRef]
- Shi, C.; Chen, J.; Ge, Q.; Sun, J.; Guo, W.; Wang, J.; Peng, L.; Xu, Q.; Fan, G.; Zhang, W.; et al. Draft genomes and comparative analysis of seven mangrove rhizosphere-associated fungi isolated from Kandelia obovata and Acanthus ilicifolius. Front. Fungal Biol. 2021, 2, 626904. [Google Scholar] [CrossRef]
- Cong, M.; Zhang, Y.; Feng, X.; Pang, X.; Liu, Y.; Zhang, X.; Yang, Z.; Wang, J. Anti-inflammatory alkaloids from the cold-seep-derived fungus Talaromyces helicus SCSIO41311. 3 Biotech 2022, 12, 161. [Google Scholar] [CrossRef]
- Li, M.; Li, S.; Hu, J.; Gao, X.; Wang, Y.; Liu, Z.; Zhang, W. Thioester-containing benzoate derivatives with α-glucosidase inhibitory activity from the deep-sea-derived fungus Talaromyces indigoticus FS688. Mar. Drugs 2022, 20, 33. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Li, X.M.; Liu, H.; Meng, L.H.; Wang, B.G. Two new diphenylketones and a new xanthone from Talaromyces islandicus EN-501, an endophytic fungus derived from the marine red alga Laurencia okamurai. Mar. Drugs 2016, 14, 223. [Google Scholar] [CrossRef] [Green Version]
- Heo, Y.M.; Lee, H.; Kim, K.; Kwon, S.L.; Park, M.Y.; Kang, J.E.; Kim, G.H.; Kim, B.S.; Kim, J.J. Fungal diversity in intertidal mudflats and abandoned solar salterns as a source for biological resources. Mar. Drugs 2019, 17, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Zhang, X.; Lin, R.; Yang, H.; Song, F.; Xu, X.; Wang, L. New secondary metabolites from the marine-derived fungus Talaromyces mangshanicus BTBU20211089. Mar. Drugs 2022, 20, 79. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Yang, S.Q.; Li, X.M.; Hu, X.Y.; Li, X.; Wang, B.G. Aromatic polyketides from the deep-sea cold-seep mussel associated endozoic fungus Talaromyces minioluteus CS-138. Mar. Drugs 2022, 20, 529. [Google Scholar] [CrossRef]
- Muwawa, E.M.; Makonde, H.M.; Obieze, C.C.; Jefwa, J.M.; Kahindi, J.H.P.; Khasa, D.P. Isolation, characterization and biotechnological potential of tropical culturable rhizospheric fungi from four mangrove species in Kenya. Afr. J. Microbiol. Res. 2020, 14, 541–554. [Google Scholar]
- Machado, F.P.; Rodrigues, I.C.; Georgopolou, A.; Gales, L.; Pereira, J.A.; Costa, P.M.; Mistry, S.; Ghoran, S.H.; Silva, A.M.S.; Dethoup, T.; et al. New hybrid phenalenone dimer, highly conjugated dihydroxylated C28 steroid and azaphilone from the culture extract of a marine sponge-associated fungus, Talaromyces pinophilus KUFA 1767. Mar. Drugs 2023, 21, 194. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Taritla, S.; Sharma, A.; Jayabaskaran, C. Antiproliferative and antioxidative bioactive compounds in extracts of marine-derived endophytic fungus Talaromyces purpureogenus. Front. Microbiol. 2018, 9, 1777. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Li, C.W.; Cui, C.B.; Cai, B.; Xu, L.L.; Zhu, H.J. Four new antitumor metabolites isolated from a mutant 3-f-31 strain derived from Penicillium purpurogenum G59. Eur. J. Med. Chem. 2018, 158, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, T.R.; Santos, G.S.; Turatti, I.C.C.; Paziani, M.H.; von Zeska Kress, M.R.; Colepicolo, P.; Debonsi, H.M. Characterization of the lipid profile of Antarctic brown seaweeds and their endophytic fungi by gas chromatography–mass spectrometry (GC–MS). Polar Biol. 2019, 42, 1431–1444. [Google Scholar] [CrossRef]
- Cao, X.; Shi, Y.; Wu, X.; Wang, K.; Huang, S.; Sun, H.; Dickschat, J.S.; Wu, B. Talaromyolides A–D and talaromytin: Polycyclic meroterpenoids from the fungus Talaromyces sp. CX11. Org. Lett. 2019, 21, 6539–6542. [Google Scholar] [CrossRef]
- Huang, Z.H.; Liang, X.; Li, C.J.; Gu, Q.; Ma, X.; Qi, S.H. Talaromynoids A–I, highly oxygenated meroterpenoids from the marine-derived fungus Talaromyces purpureogenus SCSIO 41517 and their lipid accumulation inhibitory activities. J. Nat. Prod. 2021, 84, 2727–2737. [Google Scholar] [CrossRef]
- Liang, X.; Huang, Z.H.; Shen, W.B.; Lu, X.H.; Zhang, X.X.; Ma, X.; Qi, S.H. Talaromyoxaones A and B: Unusual oxaphenalenone spirolactones as phosphatase inhibitors from the marine-derived fungus Talaromyces purpureogenus SCSIO 41517. J. Org. Chem. 2021, 86, 12831–12839. [Google Scholar] [CrossRef]
- Taritla, S.; Kumari, M.; Kamat, S.; Bhat, S.G.; Jayabaskaran, C. Optimization of physicochemical parameters for production of cytotoxic secondary metabolites and apoptosis induction activities in the culture extract of a marine algal–derived endophytic fungus Aspergillus sp. Front. Pharm. 2021, 12, 542891. [Google Scholar] [CrossRef]
- Ahmed, N.E.; Salem, S.S.; Hashem, A.H. Statistical optimization, partial purification, and characterization of phytase produced from Talaromyces purpureogenus NSA20 using potato peel waste and its application in dyes de-colorization. Biointerface Res. Appl. Chem. 2022, 12, 4417–4431. [Google Scholar]
- Küppers, L.; Ebrahim, W.; El-Neketi, M.; Özkaya, F.C.; Mándi, A.; Kurtán, T.; Orfali, R.S.; Müller, W.E.G.; Hartmann, R.; Lin, W.; et al. Lactones from the sponge-derived fungus Talaromyces rugulosus. Mar. Drugs 2017, 15, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.H.; Li, X.M.; Zhang, F.Z.; Wang, Y.N.; Wang, B.G. Talascortenes A–G, highly oxygenated diterpenoid acids from the sea-anemone-derived endozoic fungus Talaromyces scorteus AS-242. J. Nat. Prod. 2020, 83, 2528–2536. [Google Scholar] [CrossRef] [PubMed]
- Dewapriya, P.; Prasad, P.; Damodar, R.; Salim, A.A.; Capon, R.J. Talarolide A, a cyclic heptapeptide hydroxamate from an Australian marine tunicate-associated fungus, Talaromyces sp. (CMB-TU011). Org. Lett. 2017, 19, 2046–2049. [Google Scholar] [CrossRef] [PubMed]
- Paranetharan, M.S.; Thirunavukkarasu, N.; Rajamani, T.; Murali, T.S.; Suryanarayanan, T.S. Salt-tolerant chitin and chitosan modifying enzymes from Talaromyces stipitatus, a mangrove endophyte. Mycosphere 2018, 9, 215–226. [Google Scholar] [CrossRef]
- Lumbreras-Martínez, H.; Espinoza, C.; Fernández, J.J.; Norte, M.; Lagunes, I.; Padrón, J.M.; Lopez-Portillo, J.; Trigos, Á. Bioprospecting of fungi with antiproliferative activity from the mangrove sediment of the Tampamachoco coastal lagoon, Veracruz, Mexico. Sci. Fungorum 2018, 48, 53–60. [Google Scholar] [CrossRef]
- Liu, H.; Yan, C.; Li, C.; You, T.; She, Z. Naphthoquinone derivatives with anti-inflammatory activity from mangrove-derived endophytic fungus Talaromyces sp. SK-S009. Molecules 2020, 25, 576. [Google Scholar] [CrossRef] [Green Version]
- Rahaman, M.S.; Siraj, M.A.; Sultana, S.; Seidel, V.; Islam, M.A. Molecular phylogenetics and biological potential of fungal endophytes from plants of the Sundarbans mangrove. Front. Microbiol. 2020, 11, 570855. [Google Scholar] [CrossRef]
- Ma, M.; Yi, W.; Qin, L.; Lian, X.Y.; Zhang, Z. Talaromydien a and talaroisocoumarin A, new metabolites from the marine-sourced fungus Talaromyces sp. ZZ1616. Nat. Prod. Res. 2021, 36, 460–465. [Google Scholar] [CrossRef]
- Luo, M.; Chen, Y.; He, J.; Tang, X.; Wu, X.; Xu, C. Identification of a new Talaromyces strain DYM25 isolated from the Yap Trench as a biocontrol agent against Fusarium wilt of cucumber. Microbiol. Res. 2021, 251, 126841. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, C.; Huang, L.; Zhang, X.; Zhang, G.; Che, Q.; Li, D.; Zhu, T. Talarodrides A–F, nonadrides from the Antarctic sponge-derived fungus Talaromyces sp. HDN1820200. J. Nat. Prod. 2021, 84, 3011–3019. [Google Scholar] [CrossRef]
- Liu, G.; Huo, R.; Niu, S.; Song, F.; Liu, L. Two new cytotoxic decalin derivatives from marine-derived fungus Talaromyces sp. Chem. Biodiver. 2022, 19, e202100990. [Google Scholar] [CrossRef]
- Song, F.; Dong, Y.; Wei, S.; Zhang, X.; Zhang, K.; Xu, X. New antibacterial secondary metabolites from a marine-derived Talaromyces sp. strain BTBU20213036. Antibiotics 2022, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, C.; Fang, T.; Wu, L.; Liu, W.; Tang, J.; Long, Y. New steroid and isocoumarin from the mangrove endophytic fungus Talaromyces sp. SCNU-F0041. Molecules 2022, 27, 5766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, J.; Huo, R.; Xue, Y.; Hong, K.; Liu, L. Sulfur-containing benzofurans and α-pyrones from the mangrove-derived fungus Talaromyces sp. WHUF0341. Front. Mar. Sci. 2022, 9, 1034945. [Google Scholar] [CrossRef]
- Lv, H.; Su, H.; Xue, Y.; Jia, J.; Bi, H.; Wang, S.; Zhang, J.; Zhu, M.; Emam, M.; Wang, H.; et al. Polyketides with potential bioactivities from the mangrove-derived fungus Talaromyces sp. WHUF0362. Mar. Life Sci. Technol. 2023, 3, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Guan, X.; Lai, Q.; Yu, D.; Chen, Z.; Fu, X.; Zhang, B.; Chen, C.; Shao, Z.; Xia, J.; et al. Characterization of a bioactive meroterpenoid isolated from the marine-derived fungus Talaromyces sp. Appl. Microbiol. Biotechnol. 2022, 106, 2927–2935. [Google Scholar] [CrossRef]
- Hong, X.; Tang, L.; Chen, Z.; Lai, Q.; Zhang, B.; Jiang, Y.; Wang, X.; He, R.; Lin, J.; Shao, Z.; et al. Benzoquinone and furopyridinone derivatives from the marine-derived fungus Talaromyces sp. MCCC 3A01752. Nat. Prod. Res. 2022, 17, 13–26. [Google Scholar] [CrossRef]
- Nuansri, S.; Rukachaisirikul, V.; Muanprasat, C.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Furanone, morpholinone and tetrahydrofuran derivatives from the marine-derived fungus Talaromyces sp. PSU-MF07. Nat. Prod. Res. 2022, 3, 267–286. [Google Scholar] [CrossRef]
- Sun, C.; Liu, Q.; Shah, M.; Che, Q.; Zhang, G.; Zhu, T.; Zhou, J.; Rong, X.; Li, D. Talaverrucin A, heterodimeric oxaphenalenone from antarctica sponge-derived fungus Talaromyces sp. HDN151403, inhibits Wnt/β-catenin signaling pathway. Org. Lett. 2022, 24, 3993–3997. [Google Scholar] [CrossRef]
- Cai, R.; Chen, S.; Long, Y.; She, Z. Depsidones from Talaromyces stipitatus SK-4, an endophytic fungus of the mangrove plant Acanthus ilicifolius. Phytochem. Lett. 2017, 20, 196–199. [Google Scholar] [CrossRef]
- Noinart, J.; Buttachon, S.; Dethoup, T.; Gales, L.; Pereira, J.A.; Urbatzka, R.; Freitas, S.; Lee, M.; Silva, A.M.S.; Pinto, M.M.M.; et al. A new ergosterol analog, a new bis-anthraquinone and anti-obesity activity of anthraquinones from the marine sponge-associated fungus Talaromyces stipitatus KUFA 0207. Mar. Drugs 2017, 15, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.H.; Zhao, Y.J.; Qi, L.; Du, H.F.; Cao, F.; Wang, C.Y. Talasteroid, a new withanolide from the marine-derived fungus Talaromyces stollii. Nat. Prod. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Castro-Carvalho, B.; Ramos, A.A.; Prata-Sena, M.; Malhão, F.; Moreira, M.; Gargiulo, D.; Dethoup, T.; Buttachon, S.; Kijjoa, A.; Rocha, E. Marine-derived fungi extracts enhance the cytotoxic activity of doxorubicin in nonsmall cell lung cancer cells A459. Pharmacogn. Res. 2017, 9, S92–S98. [Google Scholar]
- Buttachon, S.; Zin, W.W.M.; Dethoup, T.; Gales, L.; Pereira, J.A.; Silva, A.M.; Kijjoa, A. Secondary metabolites from the culture of the marine sponge-associated fungi Talaromyces tratensis and Sporidesmium circinophorum. Planta Med. 2016, 82, 888–896. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Yang, L.; Feng, L.; Hu, F.; Zhang, F.; Ren, J.; Qiu, Y.; Wang, Z. Verruculosins A–B, new oligophenalenone dimers from the soft coral-derived fungus Talaromyces verruculosus. Mar. Drugs 2019, 17, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, Y.; Yu, Z.; Liang, X.; Qi, S. Phylogenetic diversity and bioactivity of culturable deep-sea-derived fungi from Okinawa Trough. J. Oceanol. Limnol. 2021, 39, 892–902. [Google Scholar] [CrossRef]
- Li, J.X.; Lei, X.X.; Tan, Y.H.; Liu, Y.H.; Yang, B.; Li, Y.Q. Two new bioactive polyphenols from the soft coral-derived fungus Talaromyces sp. SCSIO 041201. Nat. Prod. Res. 2021, 35, 5778–5785. [Google Scholar] [CrossRef]
- Huang, J.; She, J.; Yang, X.; Liu, J.; Zhou, X.; Yang, B. A new macrodiolide and two new polycyclic chromones from the fungus Penicillium sp. SCSIO041218. Molecules 2019, 24, 1686. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; He, L.; Chen, D.; Cai, R.; Long, Y.; Lu, Y.; She, Z. Talaramide A, an unusual alkaloid from the mangrove endophytic fungus Talaromyces sp. (HZ-YX1) as an inhibitor of mycobacterial PknG. New J. Chem. 2017, 41, 4273–4276. [Google Scholar] [CrossRef]
- Wang, B.; Nong, X.H.; Zeng, W.N.; Li, S.S.; Li, G.Y.; Liu, J.; Huang, G.L.; Zheng, C.J. Study on bioactive secondary metabolites from the mangrove-derived fungus Penicillium verruculosum TGM14. Chem. Nat. Comp. 2022, 58, 812–815. [Google Scholar] [CrossRef]
- Li, H.L.; Li, X.M.; Li, X.; Wang, C.Y.; Liu, H.; Kassack, M.U.; Meng, L.H.; Wang, B.G. Antioxidant hydroanthraquinones from the marine algal-derived endophytic fungus Talaromyces islandicus EN-501. J. Nat. Prod. 2017, 80, 162–168. [Google Scholar] [CrossRef]
- Liu, Z.; Kou, J.; Li, M.; Chen, Y.; Li, S.; Li, H.; Zhang, W. Molecular networking-based discovery of new polyketides from the deep-sea-derived fungus Talaromyces indigoticus FS688. Tetrahedron 2023, 138, 133410. [Google Scholar] [CrossRef]
- Cao, X.; Ge, Y.; Lan, D.; Wu, X.; Wu, B. Spirocyclic polyketides from the marine fungus Talaromyces sp. CX11. Fitoterapia 2023, 164, 105359. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Huang, H.; Chen, Y.; Li, S.; Liu, Z.; Wang, Y.; Zhang, W.; Gao, X. Polyketides from the deep-sea-derived fungus Talaromyces indigoticus FS688 and their cytotoxicites. Chin. J. Org. Chem. 2022, 42, 2975–2980. [Google Scholar] [CrossRef]
- Venkatachalam, M.; Magalon, H.; Dufossé, L.; Fouillaud, M. Production of pigments from the tropical marine-derived fungi Talaromyces albobiverticillius: New resources for natural red-colored metabolites. J. Food Comp. Anal. 2018, 70, 35–48. [Google Scholar] [CrossRef]
- Wu, C.J.; Qiao, H.; Cui, C.B.; Yang, Y.; Xu, R.; Sun, C.X.; Li, D.H.; Li, C.W. Two new polyketides isolated from a diethyl sulphate mutant of marine-derived Penicillium purpurogenum G59. Nat. Prod. Res. 2019, 33, 2977–2981. [Google Scholar] [CrossRef]
- Xia, M.W.; Yang, Y.; Xu, R.; Li, C.W.; Cui, C.B. A new polyketide purpurogenic acid: The activated production of polyketides by the diethyl sulphate mutagenesis of marine-derived Penicillium purpurogenum G59. Nat. Prod. Res. 2019, 33, 89–94. [Google Scholar] [CrossRef]
- Chen, S.; Ding, M.; Liu, W.; Huang, X.; Liu, Z.; Lu, Y.; Liu, H.; She, Z. Anti-inflammatory meroterpenoids from the mangrove endophytic fungus Talaromyces amestolkiae YX1. Phytochemistry 2018, 146, 8–15. [Google Scholar] [CrossRef]
- Kong, F.D.; Zhang, R.S.; Ma, Q.Y.; Xie, Q.Y.; Wang, P.; Chen, P.W.; Zhou, L.M.; Dai, H.F.; Luo, D.Q.; Zhao, Y.X. Chrodrimanins O–S from the fungus Penicillium sp. SCS-KFD09 isolated from a marine worm, Sipunculus nudus. Fitoterapia 2017, 122, 1–6. [Google Scholar] [CrossRef]
- Yi, L.; Cui, C.B.; Li, C.W.; Peng, J.X.; Gu, Q.Q. Chromosulfine, a novel cyclopentachromone sulfide produced by a marine-derived fungus after introduction of neomycin resistance. RSC Adv. 2016, 6, 43975–43979. [Google Scholar] [CrossRef]
- Cao, X.; Shi, Y.; Wu, S.; Wu, X.; Wang, K.; Sun, H.; He, S.; Dickschat, J.S.; Wu, B. Polycyclic meroterpenoids, talaromyolides E−K for antiviral activity against pseudorabies virus from the endophytic fungus Talaromyces purpureogenus. Tetrahedron 2020, 76, 131349. [Google Scholar] [CrossRef]
- Li, C.W.; Xia, M.W.; Cui, C.B.; Peng, J.X.; Li, D.H. A novel oxaphenalenone, penicimutalidine: Activated production of oxaphenalenones by the diethyl sulphate mutagenesis of marine-derived fungus Penicillium purpurogenum G59. RSC Adv. 2016, 6, 82277–82281. [Google Scholar] [CrossRef]
- Li, C.W.; Wu, C.J.; Cui, C.B.; Xu, L.L.; Cao, F.; Zhu, H.J. Penicimutamides A–C: Rare carbamate-containing alkaloids from a mutant of the marine-derived Penicillium purpurogenum G59. RSC Adv. 2016, 6, 73383–73387. [Google Scholar] [CrossRef]
- Wu, C.J.; Li, C.W.; Gao, H.; Huang, X.J.; Cui, C.B. Penicimutamides D–E: Two new prenylated indole alkaloids from a mutant of the marine-derived Penicillium purpurogenum G59. RSC Adv. 2017, 7, 24718–24722. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Dong, Y.; Yang, Y.; Xu, R.; Li, C.W.; Cui, C.B. Penicimutanin C, a new alkaloidal compound, isolated from a neomycin-resistant mutant 3-f-31 of Penicillium purpurogenum G59. Chem. Biodivers. 2020, 17, e2000241. [Google Scholar] [CrossRef]
- Zhou, C.; Cao, X.; Ge, Y.; Wu, X.; Zhang, Z.; Ma, Y.; Dickschat, J.S.; Wu, B. Talaropeptins A and B, tripeptides with an N-trans-cinnamoyl moiety from the marine-derived fungus Talaromyces purpureogenus CX11. J. Nat. Prod. 2022, 85, 2620–2625. [Google Scholar] [CrossRef]
- Wang, W.; Wan, X.; Liu, J.; Wang, J.; Zhu, H.; Chen, C.; Zhang, Y. Two new terpenoids from Talaromyces purpurogenus. Mar. Drugs 2018, 16, 150. [Google Scholar] [CrossRef] [Green Version]
- Lebeau, J.; Petit, T.; Fouillaud, M.; Dufossé, L.; Caro, Y. Alternative extraction and characterization of nitrogen-containing azaphilone red pigments and ergosterol derivatives from the marine-derived fungal Talaromyces sp. 30570 strain with industrial relevance. Microorganisms 2020, 8, 1920. [Google Scholar] [CrossRef]
- Lebeau, J.; Venkatachalam, M.; Fouillaud, M.; Petit, T.; Vinale, F.; Dufossé, L.; Caro, Y. Production and new extraction method of polyketide red pigments produced by ascomycetous fungi from terrestrial and marine habitats. J. Fungi 2017, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Nicoletti, R.; Manzo, E.; Ciavatta, M.L. Occurence and bioactivities of funicone-related compounds. Int. J. Mol. Sci. 2009, 10, 1430–1444. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, M.M.; DellaGreca, M.; Andolfi, A.; Nicoletti, R. New insights into chemical and biological properties of funicone-like compounds. Toxins 2022, 14, 466. [Google Scholar] [CrossRef] [PubMed]
- Shahid, H.; Cai, T.; Wang, Y.; Zheng, C.; Yang, Y.; Mao, Z.; Ding, P.; Shan, T. Duclauxin derivatives from fungi and their biological activities. Front. Microbiol. 2021, 12, 766440. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Hosoe, T.; Nozawa, K.; Yaguchi, T.; Udagawa, S.I.; Kawai, K.I. Mitorubrin derivatives on ascomata of some Talaromyces species of ascomycetous fungi. J. Nat. Prod. 1999, 62, 1328–1329. [Google Scholar] [CrossRef] [PubMed]
- Tammam, M.A.; El-Din, M.I.G.; Abood, A.; El-Demerdash, A. Recent advances in the discovery, biosynthesis, and therapeutic potential of isocoumarins derived from fungi: A comprehensive update. RSC Adv. 2023, 13, 8049–8089. [Google Scholar] [CrossRef] [PubMed]
- Osmanova, N.; Schultze, W.; Ayoub, N. Azaphilones: A class of fungal metabolites with diverse biological activities. Phytochem. Rev. 2010, 9, 315–342. [Google Scholar] [CrossRef]
- Diaz-Munoz, G.; Miranda, I.L.; Sartori, S.K.; de Rezende, D.C.; Diaz, M.A. Anthraquinones: An overview. Stud. Nat. Prod. Chem. 2018, 58, 313–338. [Google Scholar]
- Panda, S.S.; Chand, M.; Sakhuja, R.; Jain, S.C. Xanthones as potential antioxidants. Curr. Med. Chem. 2013, 20, 4481–4507. [Google Scholar] [CrossRef]
- Zhao, M.; Tang, Y.; Xie, J.; Zhao, Z.; Cui, H. Meroterpenoids produced by fungi: Occurrence, structural diversity, biological activities, and their molecular targets. Eur. J. Med. Chem. 2021, 209, 112860. [Google Scholar] [CrossRef]
- Stierle, D.B.; Stierle, A.A.; Bugni, T. Sequoiamonascins A−D: Novel anticancer metabolites isolated from a redwood endophyte. J. Org. Chem. 2003, 68, 4966–4969. [Google Scholar] [CrossRef]
- Zimowska, B.; Bielecka, M.; Abramczyk, B.; Nicoletti, R. Bioactive products from endophytic fungi of sages (Salvia spp.). Agriculture 2020, 10, 543. [Google Scholar] [CrossRef]
- Nonaka, K.; Chiba, T.; Suga, T.; Asami, Y.; Iwatsuki, M.; Masuma, R.; Omura, S.; Shiomi, K. Coculnol, a new penicillic acid produced by a coculture of Fusarium solani FKI-6853 and Talaromyces sp. FKA-65. J. Antib. 2015, 68, 530–532. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, M.M.; DellaGreca, M.; Nicoletti, R.; Salvatore, F.; Vinale, F.; Naviglio, D.; Andolfi, A. Talarodiolide, a new 12-membered macrodiolide, and GC/MS investigation of culture filtrate and mycelial extracts of Talaromyces pinophilus. Molecules 2018, 23, 950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, 201–241. [Google Scholar]
- Frisvad, J.C.; Larsen, T.O. Chemodiversity in the genus Aspergillus. Appl. Microbiol. Biotechnol. 2015, 99, 7859–7877. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Rank, C.; Nielsen, K.F.; Larsen, T.O. Metabolomics of Aspergillus fumigatus. Med. Mycol. 2009, 47, S53–S71. [Google Scholar] [CrossRef] [PubMed]
- Medentsev, A.G.; Arinbasarova, A.Y.; Akimenko, V.K. Biosynthesis of naphthoquinone pigments by fungi of the genus Fusarium. Appl. Biochem. Microbiol. 2005, 41, 503–507. [Google Scholar] [CrossRef]
- Tammam, M.A.; Sebak, M.; Greco, C.; Kijjoa, A.; El-Demerdash, A. Chemical diversity, biological activities and biosynthesis of fungal naphthoquinones and their derivatives: A comprehensive update. J. Mol. Struct. 2022, 1268, 133711. [Google Scholar] [CrossRef]
- Chen, D.; Liu, L.; Lu, Y.; Chen, S. Identification of fusarielin M as a novel inhibitor of Mycobacterium tuberculosis protein tyrosine phosphatase B (MptpB). Bioorg. Chem. 2021, 106, 104495. [Google Scholar] [CrossRef]
- Ishii, K.; Sakai, K.; Ueno, Y.; Tsunoda, H.; Enomoto, M. Solaniol, a toxic metabolite of Fusarium solani. Appl. Microbiol. 1971, 22, 718–720. [Google Scholar] [CrossRef]
- Lou, J.; Fu, L.; Peng, Y.; Zhou, L. Metabolites from Alternaria fungi and their bioactivities. Molecules 2013, 18, 5891–5935. [Google Scholar] [CrossRef] [PubMed]
- Hamed, S.R.; Abdel-Azeem, A.M.; Dar, P.M. Recent advancements on the role of biologically active secondary metabolites from Chaetomium. In Recent Developments on Genus Chaetomium; Abdel-Azeem, A.M., Ed.; Springer: Cham, Switzerland, 2020; pp. 177–204. [Google Scholar]
- Stodola, F.H.; Cabot, C.; Benjamin, C.R. Structure of ramulosin, a metabolic product of the fungus Pestalotia ramulosa. Biochem. J. 1964, 9, 92–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Deng, Z.; Guo, Z.; Peng, Y.; Huang, N.; He, H.; Tu, X.; Zou, K. Effect of culture conditions on metabolite production of Xylaria sp. Molecules 2015, 20, 7940–7950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Liu, S.; Sun, B.; Niu, S.; Li, E.; Liu, X.; Che, Y. Polyketides from the ascomycete fungus Leptosphaeria sp. J. Nat. Prod. 2010, 73, 905–910. [Google Scholar] [CrossRef] [PubMed]
- Sassa, T.; Aoki, H.; Namiki, M.; Munakata, K. Plant growth promoting metabolites of Sclerotinia sclerotiorum: Part I. Isolation and structures of sclerotinin A and B. Agr. Biol. Chem. 1968, 32, 1432–1439. [Google Scholar]
- Ding, Y.; Greshock, T.J.; Miller, K.A.; Sherman, D.H.; Williams, R.M. Premalbrancheamide: Synthesis, isotopic labeling, biosynthetic incorporation, and detection in cultures of Malbranchea aurantiaca. Org. Lett. 2008, 10, 4863–4866. [Google Scholar] [CrossRef] [Green Version]
- Hauser, V.D.; Sigg, H.P. Isolierung und abbau von sordarin. 1. Mitteilung über sordarin. Helv. Chim. Acta 1971, 54, 1178–1190. [Google Scholar] [CrossRef]
- Camarda, L.; Merlini, L.; Nasini, G. Metabolites of Cercospora. Taiwapyrone, an α-pyrone of unusual structure from Cercospora taiwanensis. Phytochemistry 1976, 15, 537–539. [Google Scholar] [CrossRef]
- Li, X.; Gao, J.; Li, M.; Cui, H.; Jiang, W.; Tu, Z.C.; Yuan, T. Aromatic cadinane sesquiterpenoids from the fruiting bodies of Phellinus pini block SARS-CoV-2 Spike–ACE2 interaction. J. Nat. Prod. 2021, 84, 2385–2389. [Google Scholar] [CrossRef]
- Ter Heide, R.; Provatoroff, N.; Traas, P.C.; De Valois, P.J.; Van der Plasse, N.; Wobben, H.J.; Timmer, R. Qualitative analysis of the odoriferous fraction of oakmoss (Evernia prunastri). J. Agric. Food Chem. 1975, 23, 950–957. [Google Scholar] [CrossRef]
- Zhao, Q.; Wang, C.X.; Yu, Y.; Wang, G.Q.; Zheng, Q.C.; Chen, G.D.; Lian, Y.Y.; Lin, F.; Guo, L.D.; Gao, H. Nodulisporipyrones A–D, new bioactive α-pyrone derivatives from Nodulisporium sp. J. Asian Nat. Prod. Res. 2015, 17, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.F.; Yu, R.J.; Li, X.X.; Gao, H.; Guo, L.D.; Tang, J.S.; Yao, X.S. ¹H and ¹³C NMR spectral assignments of 2-pyrone derivatives from an endophytic fungus of Sarcosomataceae. Magn. Reson. Chem. 2015, 53, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Carter, G.T. Remisporine B, a novel dimeric chromenone derived from spontaneous Diels–Alder reaction of remisporine A. Tetr. Lett. 2003, 44, 3119–3122. [Google Scholar] [CrossRef]
- Krick, A.; Kehraus, S.; Gerhäuser, C.; Klimo, K.; Nieger, M.; Maier, A.; Fiebig, H.H.; Atodiresei, I.; Raabe, G.; Fleischhauser, J.; et al. Potential cancer chemopreventive in vitro activities of monomeric xanthone derivatives from the marine algicolous fungus Monodictys putredinis. J. Nat. Prod. 2007, 70, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Arunpanichlert, J.; Rukachaisirikul, V.; Phongpaichit, S.; Supaphon, O.; Sakayaroj, J. Meroterpenoid, isocoumarin, and phenol derivatives from the seagrass-derived fungus Pestalotiopsis sp. PSU-ES194. Tetrahedron 2015, 71, 882–888. [Google Scholar] [CrossRef]
- Liu, X.J.; Li, H.J.; Ma, W.Z.; Zhang, F.M.; Xu, M.Y.; Mahmud, T.; Lan, W.J. Phomaligols F–I, polyoxygenated cyclohexenone derivatives from marine-derived fungus Aspergillus flavus BB1. Bioorg. Chem. 2021, 115, 105269. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Kwon, T.O.; Gloer, J.B.; Marvanová, L.; Shearer, C.A. Tenellic acids A−D: New bioactive diphenyl ether derivatives from the aquatic fungus Dendrospora tenella. J. Nat. Prod. 1999, 62, 580–583. [Google Scholar] [CrossRef] [PubMed]
- Walton, J.D. Horizontal gene transfer and the evolution of secondary metabolite gene clusters in fungi: An hypothesis. Fungal Gen. Biol. 2000, 30, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Rosewich, U.L.; Kistler, H.C. Role of horizontal gene transfer in the evolution of fungi. Ann. Rev. Phytopathol. 2000, 38, 325–363. [Google Scholar] [CrossRef] [Green Version]
- Husnik, F.; McCutcheon, J.P. Functional horizontal gene transfer from bacteria to eukaryotes. Nat. Rev. Microbiol. 2018, 16, 67–79. [Google Scholar] [CrossRef]
- Barreiro, C.; Gutiérrez, S.; Olivera, E.R. Fungal horizontal gene transfer: A history beyond the phylogenetic kingdoms. In Horizontal Gene Transfer: Breaking Borders between Living Kingdoms; Villa, G.T., Viñas, M., Eds.; Springer: Cham, Switzerland, 2019; pp. 315–336. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Magno, S.; Mangoni, A.; Pansini, M. Incisterols, a new class of highly degraded sterols from the marine sponge Dictyonella incisa. J. Am. Chem. Soc. 1990, 112, 3505–3509. [Google Scholar] [CrossRef]
- Sathiyabama, M.; Parthasarathy, R. Withanolide production by fungal endophyte isolated from Withania somnifera. Nat. Prod. Res. 2018, 32, 1573–1577. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.Y.; Cao, S.J.; Chen, L.X.; Qiu, F. Natural withanolides, an update. Nat. Prod. Rep. 2022, 39, 784–813. [Google Scholar] [CrossRef] [PubMed]
- Eakjamnong, W.; Keawsalong, N.; Dethoup, T. Novel ready-to-use dry powder formulation of Talaromyces tratensis KUFA0091 to control dirty panicle disease in rice. Biol. Control 2021, 152, 104454. [Google Scholar] [CrossRef]
- Paranetharan, M.S.; Thirunavukkarasu, N.; Rajamani, T.; Suryanarayanan, T.S. Biomass destructuring enzymes of fungal endophytes of mangrove roots. Curr. Res. Environ. Appl. Mycol. 2022, 12, 259–267. [Google Scholar] [CrossRef]
- Dewapriya, P.; Khalil, Z.G.; Prasad, P.; Salim, A.A.; Cruz-Morales, P.; Marcellin, E.; Capon, R.J. Talaropeptides AD: Structure and biosynthesis of extensively N-methylated linear peptides from an Australian marine tunicate-derived Talaromyces sp. Front. Chem. 2018, 6, 394. [Google Scholar] [CrossRef]
- Dethoup, T.; Kaewsalong, N.; Songkumorn, P.; Jantasorn, A. Potential application of a marine-derived fungus, Talaromyces tratensis KUFA 0091 against rice diseases. Biol. Control 2018, 119, 1–6. [Google Scholar] [CrossRef]
- Hiremath, I.S.; Goel, A.; Warrier, S.; Kumar, A.P.; Sethi, G.; Garg, M. The multidimensional role of the Wnt/β-catenin signaling pathway in human malignancies. J. Cell Physiol. 2022, 237, 199–238. [Google Scholar] [CrossRef]
- Stammati, A.; Nicoletti, R.; De Stefano, S.; Zampaglioni, F.; Zucco, F. Cytostatic properties of a novel compound derived from Penicillium pinophilum: An in vitro study. Altern. Lab. Anim. 2002, 30, 69–75. [Google Scholar] [CrossRef]
- Buommino, E.; Nicoletti, R.; Gaeta, G.M.; Orlando, M.; Ciavatta, M.L.; Baroni, A.; Tufano, M.A. 3-O-methylfunicone, a secondary metabolite produced by Penicillium pinophilum, induces growth arrest and apoptosis in HeLa cells. Cell Prolif. 2004, 37, 413–426. [Google Scholar] [CrossRef]
- Baroni, A.; De Luca, A.; De Filippis, A.; Petrazzuolo, M.; Manente, L.; Nicoletti, R.; Tufano, M.A.; Buommino, E. 3-O-methylfunicone, a metabolite of Penicillium pinophilum, inhibits proliferation of human melanoma cells by causing G2+ M arrest and inducing apoptosis. Cell Prolif. 2009, 42, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; Paoletti, I.; De Filippis, A.; Nicoletti, R.; Ciavatta, M.L.; Menegozzo, S.; Menegozzo, M.; Tufano, M.A. 3-O-Methylfunicone, a metabolite produced by Penicillium pinophilum, modulates ERK1/2 activity, affecting cell motility of human mesothelioma cells. Cell Prolif. 2010, 43, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; Tirino, V.; De Filippis, A.; Silvestri, F.; Nicoletti, R.; Ciavatta, M.L.; Pirozzi, G.; Tufano, M.A. 3-O-methylfunicone, from Penicillium pinophilum, is a selective inhibitor of breast cancer stem cells. Cell Prolif. 2011, 44, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; De Filippis, A.; Nicoletti, R.; Menegozzo, M.; Menegozzo, S.; Ciavatta, M.L.; Rizzo, A.; Brancato, V.; Tufano, M.A.; Donnarumma, G. Cell-growth and migration inhibition of human mesothelioma cells induced by 3-O-methylfunicone from Penicillium pinophilum and cisplatin. Investig. New Drugs 2012, 30, 1343–1351. [Google Scholar] [CrossRef]
- Fiorito, F.; Cerracchio, C.; Salvatore, M.M.; Serra, F.; Pucciarelli, A.; Amoroso, M.G.; Nicoletti, R.; Andolfi, A. Antiviral property of the fungal metabolite 3-O-methylfunicone in bovine herpesvirus 1 infection. Microorganisms 2022, 10, 188. [Google Scholar] [CrossRef]
- Cerracchio, C.; Iovane, V.; Salvatore, M.M.; Amoroso, M.G.; Dakroub, H.; DellaGreca, M.; Nicoletti, R.; Andolfi, A.; Fiorito, F. Effectiveness of the fungal metabolite 3-O-methylfunicone towards canine coronavirus in a canine fibrosarcoma cell line (A72). Antibiotics 2022, 11, 1594. [Google Scholar] [CrossRef]
- Wang, C.; Tang, S.; Cao, S. Antimicrobial compounds from marine fungi. Phytochem. Rev. 2021, 20, 85–117. [Google Scholar] [CrossRef]
- Bellavita, R.; Maione, A.; Merlino, F.; Siciliano, A.; Dardano, P.; De Stefano, L.; Galdiero, S.; Galdiero, E.; Grieco, P.; Falanga, A. Antifungal and antibiofilm activity of cyclic temporin L peptide analogues against albicans and non-albicans Candida species. Pharmaceutics 2022, 14, 454. [Google Scholar] [CrossRef]
- Youssef, F.S.; Ashour, M.L.; Singab, A.N.B.; Wink, M. A comprehensive review of bioactive peptides from marine fungi and their biological significance. Mar. Drugs 2019, 17, 559. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Furutani, S.; Ihara, M.; Ling, Y.; Yang, X.; Kai, K.; Hayashi, H.; Matsuda, K. Meroterpenoid chrodrimanins are selective and potent blockers of insect GABA-gated chloride channels. PLoS ONE 2015, 10, e0122629. [Google Scholar] [CrossRef]
- Noor, A.O.; Almasri, D.M.; Bagalagel, A.A.; Abdallah, H.M.; Mohamed, S.G.A.; Mohamed, G.A.; Ibrahim, S.R.M. Naturally occurring isocoumarins derivatives from endophytic fungi: Sources, isolation, structural characterization, biosynthesis, and biological activities. Molecules 2020, 25, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Source | Location | Reference |
|---|---|---|---|
| T. aculeatus | mangrove (Kandelia candel, leaf) | Guangdong (China) | [20] * |
| deep sea sediment | Indian Ocean | [24] * | |
| red alga (Laurencia obtusa) | Suez Gulf (Egypt) | [29] * | |
| T. albobiverticillius | coral rubble, sediment | La Reunion Island | [31] * |
| unidentified ascidian | Manado (Indonesia) | [22] * | |
| T. amestolkiae | mangrove (Kandelia obovata, leaf) | Guangdong (China) | [32] * |
| pipefish (Syngnathus acus) † | Hainan (China) | [33] * | |
| T. assiutensis | mangrove (Ceriops tagal, leaf) | South China Sea | [34] * |
| mangrove (Avicennia marina, root) | Maharashtra (India) | [35] | |
| T. brunneus | sponge (Axinella polypoides) | Marmara (Turkey) | [36] |
| T. cellulolyticus | coral | South China Sea | [37] * |
| T. cyanescens | green alga (Caulerpa sp.) | Da Nang (Vietnam) | [38] * |
| T. flavus | sediment | Kanyakumari district (India) | [39] |
| mangrove (Acanthus ilicifolius, stem) | Hainan (China) | [40] * | |
| sponge (Mycale sp.) | Samaesarn Island (Thailand) | [41] | |
| T. funiculosus | deep sea sediment | Shimokita Peninsula (Japan) | [42] |
| mangrove sediment | Hainan (China) | [23,25] * | |
| coral (Porites compressa) | Zhanjiang (China) | [43] | |
| sea cucumber (Holothuria leucospilota) | Pangkor Island (Malaysia) | [44] | |
| deep sea sediment | South China Sea | [45] * | |
| T. fuscoviridis | mangrove rhizosphere | Hainan (China) | [46] |
| T. haitouensis | mudflat in estuary | Jiangsu (China) | [16] |
| T. helicus | deep sea sediment | South China Sea | [47] * |
| T. indigoticus | deep sea sediment | South China Sea | [48] * |
| T. islandicus | red alga (Laurencia okamurai) | Qingdao (China) | [49] * |
| T. liani | mudflat in intertidal zone | Yongyudo (South Korea) | [50] |
| T. mangshanicus | sediment | South China Sea | [51] * |
| T. minioluteus | sediment | East China Sea | [27] * |
| mussel (Gigantidas platifrons) | South China Sea | [52] * | |
| T. pinophilus | mangrove sediment | Xiamen (China) | [19] * |
| mangrove rhizosphere | Techeng Isle (China) | [26] * | |
| mangrove (A. marina) rhizosphere | Gazi Bay (Kenya) | [53] | |
| sponge (Mycale sp.) | Samaesarn Island (Thailand) | [54] * | |
| T. purpureogenus | mudflat in intertidal zone | Tianjin (China) | [18] * |
| brown alga | Kovalam (India) | [55] | |
| mud at the coastline | Hebei (China) | [56] * | |
| brown alga (Phaeurus antarcticus) | Half Moon Island (Antarctica) | [57] | |
| red alga (Grateloupia filicina) | Zhejiang (China) | [58] * | |
| soft coral | Nansha islands (China) | [59] * | |
| soft coral | South China Sea | [60] * | |
| brown alga (Sargassum muticum) | Kerala (India) | [61] | |
| water | Sharm El-Sheikh governorate (Egypt) | [62] | |
| T. rotundus | reef water | La Reunion Island | [31] |
| T. rugulosus | sponge (Axinella cannabina) | Sığaçık-İzmir (Turkey) | [63] * |
| T. scorteus | sea anemone (Cerianthus sp.) | Magellan Sea Mounts | [64] * |
| Talaromyces sp. | mangrove (Sonneratia apetala, leaf) | Guangdong (China) | [17] * |
| unidentified tunicate | Tweed Heads (Australia) | [65] * | |
| annellid (Sipunculus nudus) | Haikou Bay (China) | [21] * | |
| mangrove (Rhizophora mucronata, root) | Andaman Islands (India) | [66] | |
| mangrove (Laguncularia racemosa) rhizosphere | Vera Cruz (Mexico) | [67] | |
| abandoned saltern mudflat in intertidal zone | Yubudo (South Korea) Gopado, Yongyudo (South Korea) | [50] | |
| mangrove (R. mucronata) rhizosphere | Gazi Bay, Mida Creek (Kenya) | [53] | |
| coral (Porites pukoensis) | Zhanjiang (China) | [43] | |
| mangrove (K. obovata, fruit) | Guangxi (China) | [68] * | |
| mangrove (Brownlowia tersa, stem) | Sundarbans (Bangladesh) | [69] | |
| mangrove (Ceriops decandra, bark) | |||
| mangrove (Heritiera fomes, bark) | |||
| mangrove (Xylocarpus granatum, bark) | |||
| mangrove (Xylocarpus moluccensis, bark) | |||
| halibut (Hippoglossus sp.) | Zhejiang (China) | [70] * | |
| water | Yap Trench | [71] | |
| sponge | Weddell Sea (Antarctica) | [72] * | |
| mangrove (X. granatum, root) | Hainan (China) | [30] * | |
| sediment | Zhejiang (China) | [73] * | |
| mudflat in intertidal zone | Qingdao (China) | [74] * | |
| mangrove (Kandelia sp., leaf) | Guangdong (China) | [75] * | |
| mangrove soil | Hainan (China) | [76,77] * | |
| water | Dongshan Island (China) | [78,79] * | |
| unidentified sponge | Bulon Island (Thailand) | [80] * | |
| unidentified sponge | Prydz Bay (Antarctica) | [81] * | |
| T. stipitatus | mangrove (A. ilicifolius, leaf) | Guanxi (China) | [82] * |
| sponge (Stylissa flabelliformis) | Samaesarn Island (Thailand) | [83] * | |
| mangrove (A. marina, root) | Tamil Nadu (India) | [66] | |
| mudflat in intertidal zone | Yongyudo (South Korea) | [50] | |
| mangrove (R. mucronata) rhizosphere | Gazi Bay (Kenya) | [53] | |
| brown alga (S. muticum) | Kerala (India) | [61] | |
| T. stollii | unknown | Bohai Sea (China) | [84] * |
| T. trachyspermus | sponge (Clathria reinwardti) | Kram Island (Thailand) | [85] |
| T. tratensis | sponge (Mycale sp.) | Samaesarn Island (Thailand) | [86] * |
| T. variabilis | mangrove rhizosphere | Fujian (China) | [24] * |
| T. verruculosus | reef water | La Reunion Island | [31] |
| soft coral (Goniopora sp.) | Hainan (China) | [87] * | |
| mangrove (A. marina) rhizosphere | Mida Creek (Kenya) | [53] | |
| mangrove (C. tagal) rhizosphere | Gazi Bay (Kenya) | ||
| mangrove (X. moluccensis, pneumatophores) | Sundarbans (Bangladesh) | [69] | |
| deep-sea sediment | Okinawa Trough | [88] | |
| mangrove (X. granatum) | South China Sea | [28] * | |
| T. versatilis | soft coral | Yongxing Island (China) | [89] * |
| T. zhenhaiensis | mudflat in estuary | Zhejiang (China) | [16] |
| Compound | Species | Reference |
|---|---|---|
| Alkaloids | ||
| Chaetominine B | T. helicus | [47] |
| Ditalaromylectones A–B | T. mangshanicus | [51] |
| 11,17-epi-Mangrovamide A | T. funiculosus | [43] |
| Mangrovamides D–K | T. funiculosus | [25] |
| Mangrovlide A | T. funiculosus | [90] |
| Talaramide A | T. amestolkiae | [91] |
| Talaromanloid A | T. mangshanicus | [51] |
| Amides | ||
| Penicimumide | T. purpureogenus | [56] |
| Talaromydene | T. mangshanicus | [51] |
| Talaromydien A | Talaromyces sp. T. verruculosus | [70] [92] |
| Talaromylectone | T. mangshanicus | [51] |
| Anthraquinones | ||
| 4-8-Dihydroxyconiothyrinone B, 8-11-dihydroxyconiothyrinone B, 8-hydroxyconiothyrinone B, 8-dihydroxy-10-O-methyldendryol E | T. islandicus | [93] |
| 2,2′-bis-(7-Methyl-1,4,5-trihydroxy-anthracene-9,10-dione) | T. stipitatus | [83] |
| Rugulosin D | Talaromyces sp. | [74] |
| Azaphilones | ||
| Azaphilone compounds 1–3 and 5 | T. indigoticus | [94] |
| 7-epi-Pinazaphilone B | T. pinophilus | [54] |
| Talaromyacins A–C | T. purpureogenus | [95] |
| Benzofurans | ||
| Eurothiocins C–H | T. indigoticus | [48] |
| 1-(5-Hydroxy-7-methoxybenzofuran-3-yl)ethan-1-one, 5-hydroxy-7-methoxy-2-methylbenzofuran-3-carboxylic acid | T. amestolkiae | [32] |
| (2-Hydroxypropan-2-yl)-6-methyl-2,3-dihydrobenzofuran-4-ol | T. indigoticus | [96] |
| Isoprenyl-benzofuran derivative | T. indigoticus | [94] |
| Talabenzofurans A–C | Talaromyces sp. | [76] |
| Talarominine A | T. minioluteus | [52] |
| Benzophenones | ||
| 2,2′,5′-Trihydroxy-3-methoxy-3′-methylbenzophenone, 2,2′,3,5-tetrahydroxy-3′-methylbenzophenone | T. islandicus | [49] |
| Benzoquinones | ||
| Anserinone C | Talaromyces sp. | [79] |
| Chromones | ||
| 2-(2′-Hydroxypropyl)-5-methyl-7,8-dihydroxychromone | T. aculeatus | [20] |
| Decalins | ||
| Fusarielins O–P | Talaromyces sp. | [73] |
| Depsidones | ||
| 5′-Hydroxypenicillide | T. pinophilus | [19] |
| Talamins A–D | T. minioluteus | [52] |
| Talaromyones A–B | T. stipitatus Talaromyces sp. | [82] [77] |
| Talaronins A–H | Talaromyces sp. | [77] |
| Diphenyl ethers | ||
| 2-Hydroxy-6-(2′-hydroxy-3′-hydroxymethyl-5-methylphenoxy)-benzoic acid | T. albobiverticillius | [22] |
| Funicones | ||
| Pinophilones A–E | T. pinophilus | [26] |
| Furans | ||
| Talarofuranone, talarotetrahydrofuran | Talaromyces sp. | [80] |
| Indenes | ||
| 1,2-Indandiol | T. funiculosus | [23] |
| Isocoumarins | ||
| Aspergillumarin C | Talaromycs sp. | [75] |
| 5,6-Dihydroxy-3-(4-hydroxypentyl)-isochroman-1-one, 6,8-dihydroxy-5-methoxy-3-methyl-isochromen-1-one, 5-hydroxy-4-(1-hydroxyethyl)-8-methoxyisocoumarin, 6-hydroxy-8-methoxy-3,4-dimethylisocoumarin, isobutyric acid 5,7-dihydroxy-2-methyl-4-oxo-3,4-dihydro-naphththalen-1-yl methyl ester | T. amestolkiae | [32] |
| 3-(4,5-Dihydroxy-pentyl)-8-hydroxy-isochroman-1-one | T. amestolkiae T. flavus | [32] [40] |
| Penicimarins L–M | Talaromyces sp. | [30] |
| Peniisocoumarin H | T. minioluteus | [27] |
| Talaroisocoumarin A | Talaromyces sp. | [70] |
| Talaromarins A–F | T. flavus | [40] |
| Talumarins A–B | T. rugulosus | [63] |
| Tratenopyrone | T. tratensis | [86] |
| Ketones | ||
| 6-(2-Carboxyvinyl)-N-GABA-PP-V | T. albobiverticillius | [97] |
| Penicillquei C | T. verruculosus | [92] |
| Penicimutanolones A–B, penicimutanolone A methyl ether | T. purpureogenus | [56] |
| Penitalarins A–C | T. aculeatus/T. variabilis | [24] |
| 2-Prop-1-en-1-yl-oct-4-ene-1,6,7-triol | T. indigoticus | [96] |
| Purpurofuranone | T. purpureogenus | [98] |
| Purpurogenic acid | T. purpureogenus | [99] |
| Purpuropyranone | T. purpureogenus | [98] |
| Talarocyclopenta A–C | T. assiutensis | [34] |
| Lactones | ||
| Lactone acid n-butyl ester, lactone diacid 7-O-n-butyl ester, 4-methoxylactone acid n-butyl ester | T. rugulosus | [40] |
| 5-Methylhexahydrofuro[2,3-b]furan-2-yl-ethanol | T. indigoticus | [96] |
| Nafuredin B | T. aculeatus/T. variabilis | [24] |
| cis-Resorcylide, 7-O-n-butylresorcylides, 7-hydroxyresorcylides, 7-methoxyresorcylides | T. rugulosus | [40] |
| Talarodilactones A–B | T. rugulosus | [40] |
| Meroterpenoids | ||
| Amestolkolides A–D | T. amestolkiae | [100] |
| Chrodrimanins K–S | Talaromyces sp. | [21,101] |
| Chromosulfine | T. purpureogenus | [102] |
| Taladrimanin A | Talaromyces sp. | [78] |
| Talaromynoids A–I | T. purpureogenus | [59] |
| Talaromyolides A–K | T. purpureogenus | [58,103] |
| Talaromytin | T. purpureogenus | [58] |
| Morpholinones | ||
| Talaromorpholinone | Talaromyces sp. | [80] |
| Naphthoquinones | ||
| Talanaphthoquinones A–B | Talaromyces sp. | [68] |
| Nonadrides | ||
| Talarodrides A–F | Talaromyces sp. | [72] |
| Phenalenones | ||
| Abeopyrenulin, 11-apopyrenulin | T. purpureogenus | [60] |
| Amestolkins A–B | T. amestolkiae | [33] |
| Bacillisporins K–L | Talaromyces sp. | [74] |
| Dihydroxy-ergosta-4,6,8(14)-tetraen-3-one | T. pinophilus | [54] |
| Penicimutalidine | T. purpureogenus | [104] |
| Penicimutamides A–E | T. purpureogenus | [105,106] |
| Penicimutanin C | T. purpureogenus | [107] |
| Talaromyoxaones A–B | T. purpureogenus | [60] |
| Talaropinophilide, talaropinophilone | T. pinophilus | [54] |
| Talaverrucin A | Talaromyces sp. | [81] |
| Verruculosins A–B | T. verruculosus | [87] |
| Peptides | ||
| Penicimutide | T. purpureogenus | [18] |
| Talaropeptins A–B | T. purpureogenus | [108] |
| Polyenes | ||
| Talacyanols A–C | T. cyanescens | [38] |
| Polyphenols | ||
| Talaversatilis A–B | T. versatilis | [89] |
| Pyrones | ||
| Talapyrones A–B | Talaromyces sp. | [76] |
| Pyrroles | ||
| (R)-3-Hydroxy-2,7-dimethylfuro[3,4-b]pyridin-5(7H)-one | Talaromyces sp. | [79] |
| 10-Hydroxy-8-demethyltalaromydine, 11-hydroxy-8-demethyltalaromydine | T. mangshanicus | [51] |
| Sterols | ||
| Cyclosecosteroid A | Talaromyces sp. | [75] |
| Talarosterone | T. stipitatus | [83] |
| Talasteroid | T. stollii | [84] |
| Sulfones | ||
| Pensulfonamide, pensulfonoxy | T. aculeatus | [29] |
| Terpenes | ||
| Dihydroxyisocupressic acid | T. scorteus | [64] |
| 9,10-Diolhinokiic acid | T. purpureogenus | [109] |
| Purpurides E–G | T. minioluteus | [27] |
| Roussellol C | T. purpureogenus | [109] |
| Talascortenes A–G | T. scorteus | [64] |
| Verruculides B2–B3 | Talaromyces sp. | [21] |
| Xanthones | ||
| Penixanthones A–D | T. funiculosus | [23,90] |
| 1,4,7-Trihydroxy-6-methylxanthone | T. islandicus | [49] |
| Compound | Species | Reference |
|---|---|---|
| Acids | ||
| Asperitaconic acid B, butylitaconic acid | T. assiutensis | [34] |
| Bromothiobenzoic acid | T. aculeatus | [29] |
| Coculnol, acetylcoculnol | Talaromycs sp. | [79] |
| Hydroxybenzoic acid | T. versatilis | [89] |
| 8-Hydroxy-carboxy-methylenenonanoic acid, 9-hydroxy-carboxy-methylenenonanoic acid | T. assiutensis | [34] |
| Isocyclopaldic acid | T. funiculosus | [45] |
| Methylcurvulinate | T. minioluteus | [27] |
| Methylorsellinate | T. indigoticus | [96] |
| Alcohols | ||
| bis-Methoxybenzyl-butanediol | T. tratensis | [86] |
| Alkaloids | ||
| Alantrypinone | T. verruculosus | [28] |
| Chaetominine | T. helicus | [47] |
| Cyclotryprostatin B | T. helicus | [47] |
| Cyclotryprostatin E | T. purpureogenus | [109] |
| Dihydroxyfumitremorgin C | T. helicus | [47] |
| Fructigenines A–B | T. purpureogenus | [107] |
| Fumigaclavine C, fumigatin oxide, fumiquinazolines F, G, J | T. helicus | [47] |
| Mangrovamides A, C, G, I | T. funiculosus | [25,45] |
| Methoxyspirotryprostatin B | T. purpureogenus T. helicus | [109] [47] |
| Methyl-hexahydro-pyrazino-pyrido-indole-dione | T. purpureogenus | [92] |
| Penicimutanin A | T. purpureogenus | [107] |
| Premalbrancheamide | T. purpureogenus | [106] |
| Pseurotin A, F1, methylpseurotin A, norpseurotin A | T. helicus | [47] |
| Rugulosuvine A | T. purpureogenus | [107] |
| Spiro-dipyrrolo-pyrazine-indole-trione | T. helicus | [47] |
| Tryptoquivalines F, J, isotryptoquivaline F | T. helicus | [47] |
| Amides | ||
| Hydroxy-methoxyphenyl-acetamide | T. cellulolyticus | [37] |
| Hydroxy-methyl-oxobutyl-butanamide | Talaromyces sp. | [80] |
| Anthraquinones | ||
| Acetylquestinol | T. pinophilus | [34] |
| Citrorosein | T. stipitatus T. minioluteus | [83] [27] |
| Dihydroxy-methoxy-methyl-anthracene-dione | T. funiculosus | [23] |
| Emodin, fallacinol, questinol, rheoemodin | T. stipitatus | [83] |
| Questin | T. funiculosus | [45] |
| Rugulosin A | Talaromyces sp. | [74] |
| Azaphilones | ||
| FK17-P2b1 | T. minioluteus Talaromyces sp. | [27] [78] |
| Glutarylmonascorubraminic acid, hydroxyethyl-monascorubramin, threonine-monascorubramine, threonine-rubropunctamine, GABA-rubropunctatin | T. albobiverticillius | [110] |
| Mitorubrin | T. purpureogenus Talaromyces sp. | [99] [73] |
| Mitorubrinol | T. purpureogenus | [99] |
| Monascorubramine, glutarylrubropunctamine, glycylrubropunctatin | T. albobiverticillius | [111] |
| Peniazaphilin B | Talaromyces sp. | [76,79] |
| Pinazaphilone B | Talaromyces sp. | [73] |
| Pinophilin | Talaromyces sp. | [73] |
| Pinophilins B, G | T. pinophilus | [26] |
| Purpurquinone A | T. minioluteus | [27] |
| Sch1385568 | T. pinophilus | [34] |
| Sch725680 | T. pinophilus | [26] |
| Sequoiamonascin C | Talaromyces sp. | [73] |
| Wortmin | T. tratensis | [86] |
| Benzaldehydes | ||
| Dihydroxybenzaldehyde | Talaromyces sp. | [77] |
| Ethyl-dihydroxy-methylbenzaldehyde | Talaromyces sp. | [78] |
| Hydroxybenzaldehyde, hydroxy-methylbutenyl-benzaldehyde | Talaromyces sp. | [79] |
| Benzofurans | ||
| Carboxy-methyl-butenyl-octahydro-methoxycarbonyl-3-methyl-methylene-oxo-benzofuranacetic acid | Talaromyces sp. | [80] |
| Dihydroxy-dimethyl-dibenzofuran | T. versatilis | [89] |
| Eurothiocin A | T. cyanescens T. indigoticus Talaromyces sp. | [38] [48] [76] |
| Purpuresters A–B | T. minioluteus | [27] |
| Trypacidin | T. helicus | [47] |
| Benzoquinones | ||
| Fumiquinone B | T. helicus | [47] |
| Cerebrosides | ||
| Flavuside B | T. verruculosus | [28] |
| Cyclopentenones, Cyclohexenones | ||
| Phomaligol A | T. funiculosus | [45] |
| Terrein | T. verruculosus | [92] |
| Decalins | ||
| Fusarielin M | Talaromyces sp. | [73] |
| Depsidones | ||
| Dehydroisopenicillide, dehydroxypenicillide, purpactin C | T. pinophilus | [19] |
| Isopenicillide | T. pinophilus | [19,26] |
| Methyldehydroisopenicillide | T. pinophilus | [26] |
| Penicillide | T. pinophilus T. funiculosus T. stipitatus T. verruculosus Talaromyces sp. | [19,26] [23] [82] [28] [74] |
| Purpactin A (=vermixocin B) | T. pinophilus T. stipitatus Talaromyces sp. | [19] [82] [77] |
| Purpactin C’ | Talaromyces sp. | [77] |
| Secopenicillide A | T. pinophilus Talaromyces sp. | [19] [77] |
| Secopenicillide B | T. stipitatus Talaromyces sp. | [82] [77] |
| Diphenyl ethers | ||
| Diorcinol, methyldiorcinol, methoxycarbonyldiorcinol | T. versatilis | [89] |
| Methoxy-methyl-biphenyltriol | T. mangshanicus | [51] |
| Methyl tenellate | T. pinophilus | [19] |
| Tenellic acid A | T. stipitatus | [82] |
| Tenellic acid C | T. stipitatus Talaromyces sp. | [82] [77] |
| Esters | ||
| Ethyl everninate | T. indigoticus | [96] |
| Methyl-hydroxy-methylhexenoate, methyl-hydroxyphenyl-acetate | T. minioluteus | [27] |
| Funicones | ||
| Demethylvermistatin, epi-hydroxydihydrovermistatin, methyldihydrovermistatin, penisimplicissin, demethylpenisimplicissin, penicidones C–D | T. pinophilus | [26] |
| Dihydrovermistatin | T. pinophilus Talaromyces sp. | [26] [78] |
| Funicone, deoxyfunicone | T. pinophilus | [19] |
| Methylfunicone, hydroxyvermistatin, methoxyvermistatin | T. pinophilus | [19,26] |
| Vermistatin | T. pinophilus Talaromyces sp. | [19,26] [73,78] |
| Furans | ||
| Azaspirofuran A | T. helicus | [47] |
| Cillifuranone | T. purpureogenus | [98] |
| Glycosides | ||
| Carnemycins B, E | T. verruculosus | [28] |
| Isocoumarins | ||
| Aspergillumarin A | T. amestolkiae T. flavus T. rugulosus T. verruculosus Talaromyces sp. | [32] [40] [63] [92] [30,70,75] |
| Aspergillumarin B | T. amestolkiae T. verruculosus Talaromyces sp. | [32] [92] [70,75] |
| Dihydroxy-2-hydroxypropyl-methylisochromenone, dihydroxy-2S-hydroxypropyl-methylisochromenone | T. flavus Talaromyces sp. | [40] [78] |
| Dihydroxyl-oxoisochromanyl-propanoic acid | Talaromyces sp. | [75] |
| Dihydroxymellein | Talaromyces sp. | [77] |
| Dihydroxy-trimethylisochromanone, dihydroxy-trimethylisochroman | Talaromyces sp. | [79] |
| Dimethyl-dihydroxyisocoumarin | T. amestolkiae | [32] |
| Hydroxy-hydroxymethyl-methoxy-methylisocoumarin | T. amestolkiae | [32] |
| Hydroxy-hydroxypropyl-methoxyisochromanone | T. flavus | [40] |
| Hydroxymellein | T. cellulolyticus Talaromyces sp. | [37] [78,79] |
| Hydroxy-methoxy-dimethylchromone | T. minioluteus | [52] |
| Hydroxy-methoxy-methylphthalide | T. funiculosus | [90] |
| Hydroxy-methyl-dimethoxycoumarin | Talaromyces sp. | [70] |
| Hydroxypropyl-hydroxy-dihydroisocoumarin | T. flavus | [40] |
| Hydroxyramulosin | Talaromyces sp. | [76] |
| Orthosporin | T. minioluteus | [27] |
| Penicifuran A | Talaromyces sp. | [70] |
| Peniciisocoumarins A–G | T. flavus | [40] |
| Peniciisocoumarin D | Talaromyces sp. | [70] |
| Peniciisocoumarins E–F | Talaromyces sp. | [30] |
| Penicilloxalone B | Talaromyces sp. | [30,70] |
| Penicimarin B | T. amestolkiae | [32] |
| Penicimarin C | T. amestolkiae T. flavus | [32] [40] |
| Penicimarin G | T. flavus Talaromyces sp. T. verruculosus | [40] [30] [92] |
| Penicimarin H | T. flavus Talaromyces sp. | [40] [30] |
| Penicimarin I | Talaromyces sp. | [30] |
| Penicimarin N | T. flavus | [40] |
| Pestalotiorin | T. flavus Talaromycs sp. | [40] [79] |
| Ramulosin | T. cyanescens Talaromyces sp. | [38] [76] |
| Sclerotinin A | Talaromyces sp. | [78,79] |
| Sclerotinin B | Talaromyces sp. | [79] |
| Sescandelin | T. amestolkiae Talaromyces sp. | [32] [70] |
| Sescandelin B | T. amestolkiae | [32] |
| Trihydroxy-hydroxyethylisocoumarin | T. amestolkiae Talaromyces sp. | [32] [70] |
| Ketones | ||
| Dihydro-hydroxy-hydroxymethyl-methoxy-methylnaphtho-furandione | Talaromyces sp. | [68] |
| Methyl-dihydropyranone | Talaromyces sp. | [78] |
| Penicillquei A | T. verruculosus | [92] |
| Lactones | ||
| Aspergilactone B | T. verruculosus | [92] |
| Carboxyphthalide | T. aculeatus | [20] |
| Corymbiferan lactone A | T. purpureogenus | [104] |
| Dehydromevalonic lactone, mevalonolactone | T. funiculosus | [90] |
| Deoxyrubralactone | T. pinophilus | [34] |
| Lactone acid, lactone diacid | T. rugulosus | [63] |
| Nafuredin A | T. aculeatus/T. variabilis T. mangshanicus | [24] [51] |
| Meroterpenoids | ||
| Austinolide | T. purpureogenus T. mangshanicus T. stollii | [103] [51] [84] |
| Austin, austinol, dehydroaustin | T. stollii | [84] |
| Berkeleyacetal, berkeleyacetal A, epoxyberkeleydione | T. purpureogenus | [59] |
| Chrodrimanins A–B | T. amestolkiae Talaromyces sp. T. cellulolyticus T. stollii | [100] [21,78] [37] [84] |
| Chrodrimanin C | T. cellulolyticus T. stollii | [37] [84] |
| Chrodrimanin E | Talaromyces sp. | [101] |
| Chrodrimanin F | Talaromyces sp. T. cellulolyticus | [101] [37] |
| Chrodrimanin H | Talaromyces sp. T. cellulolyticus | [21,78] [37] |
| Dehydroaustinol | T. mangshanicus T. stollii | [51] [84] |
| Hydroxypentacecilide A | Talaromyces sp. | [101] |
| Miniolutelide C | T. purpureogenus | [59] |
| Preaustinoid | T. purpureogenus | [103] |
| Purpurogenolide E | T. amestolkiae | [100] |
| Territrem B | T. verruculosus | [92] |
| Thailandolide B | Talaromyces sp. | [79] |
| Verruculide A | T. cellulolyticus | [37] |
| Verruculide B | Talaromyces sp. | [101] |
| Naphthoquinones | ||
| Acetonyl-methyl-hydroxy-methoxy-naphthazarin, acetyloxyethyl-hydroxy-dimethoxy-naphthalenedione, hydroxy-hydroxyethyl-dimethoxy-naphthalenedione | Talaromyces sp. | [68] |
| Anhydrofusarubin | Talaromyces sp. | [68] |
| Ethyl-dimethoxyjuglone | Talaromyces sp. | [68] |
| Javanicin, anhydrojavanicin | Talaromyces sp. | [68] |
| Peptides | ||
| Cyclo(l-Val- l-Pro), cyclo(l-Ile- l-Pro), cyclo(l-Leu- l-Pro), cyclo(l-Phe- l-Pro) | T. purpureogenus | [18] |
| Phenalenones | ||
| Bacillisporin A | Talaromyces sp. T. pinophilus | [81] [34] |
| Bacillisporin B | T. aculeatus Talaromyces sp. T. pinophilus | [20] [74] [34] |
| Bacillisporin C | T. aculeatus T. purpureogenus Talaromyces sp. | [20] [104] [77] |
| Bacillisporin F | T. verruculosus | [87] |
| Dihydroxy-hydroxybenzylidene-methylbutenyl-indane-carboxylic acid methyl ester | T. verruculosus | [28] |
| Duclauxin, xenoclauxin | T. verruculosus | [87] |
| Macrosporusone D | Talaromyces sp. | [74] |
| SF226 | T. purpureogenus | [104] |
| Phenols | ||
| Acetamidophenol | Talaromyces sp. | [70] |
| Alternaphenol B | Talaromyces sp. | [77] |
| Altenusin | T. mangshanicus Talaromyces sp. | [51] [73] |
| Expansols C–F | T. versatilis | [89] |
| Hydroxymethyl-methyl-heptenylphenol | T. versatilis | [89] |
| Methyl-hydroxy-trimethylphenylpropionate | T. funiculosus | [90] |
| Pyrocatechol | Talaromyces sp. | [79] |
| Talaromycin C, deacetyltalaromycin C | T. pinophilus Talaromyces sp. | [19] [77] |
| Trihydroxybutyl-hydroxy-hydroxy-methylphenoxy-methylphenylacetate | T. versatilis | [89] |
| Tyrosol | T. verruculosus | [28] |
| Phenones | ||
| Isomonodictyphenone | T. versatilis | [89] |
| Monodictyphenone | T. albobiverticillius | [22] |
| Pyridines | ||
| Aminopyridine | T. verruculosus | [28] |
| Pyrones | ||
| Dihydroaspyrone | T. indigoticus | [96] |
| Fonsecinone A | T. aculeatus | [29] |
| Nodulisporipyrone A | Talaromyces sp. | [76] |
| Scirpyrone H, xylapyrone E | T. indigoticus | [96] |
| Similanpyrone B, hydroxy-dimethylpyrone | Talaromyces sp. | [77] |
| Taiwapyrone | T. purpureogenus | [98] |
| Pyrrolidines | ||
| Dioxo-propanylidene-pyrrolidinyl- acrylic acid, propanylidene-pyrrolidine-dione | T. mangshanicus | [51] |
| Sterols | ||
| Cerevisterol | Talaromycs sp. | [75] |
| Cyathisterone | T. stipitatus | [83] |
| Dankasterone | T. purpureogenus | [109] |
| Dankasterone B | T. funiculosus | [25] |
| Epidioxyergostadienol | T. verruculosus Talaromyces sp. | [28] [75] |
| Ergostatrienol | T. aculeatus | [29] |
| Ergosterol, ergostadienetetraol, ergostadienetriol | T. albobiverticillius T. verruculosus Talaromyces sp. | [111] [28] [75] |
| Ergosterol-endoperoxide, ergostatetraenone | T. stipitatus | [83] |
| Ganodermaside A | T. verruculosus | [28] |
| Helvolic acid | T. aculeatus | [29] |
| Hydroxy-ergostatrienone | T. stollii | [84] |
| Methylincisterol, dimethylincisterol A3 | T. versatilis | [89] |
| Terpenes | ||
| Berkedrimane B | T. minioluteus | [27] |
| Hydroxyconfertifolin | T. minioluteus | [27] |
| Penioxalicin | Talaromyces sp. | [80] |
| Piniterpenoid D | T. pinophilus | [34] |
| Sordarin | Talaromyces sp. | [65] |
| Solaniol | Talaromyces sp. | [68] |
| Xanthones | ||
| Conioxanthone A | Talaromycs sp. | [17] |
| Dihydroxymethyl-hydroxymethylxanthone | T. funiculosus | [45] |
| Leptosphaerin G | T. funiculosus | [25] |
| Pinselin, methyl-hydroxy-methyl-oxo-xanthene-carboxylate, sydowinins A–B | Talaromycs sp. | [17] |
| Remisporine B, epi-remisporine B | Talaromycs sp. | [17] |
| Secalonic acid A | T. stipitatus | [83] |
| Secalonic acid D | Talaromyces sp. | [77] |
| Trihydroxymethylxanthone | T. islandicus | [49] |
| Compound Name | Reported Bioactivities 1 | References |
|---|---|---|
| Acetonyl-methyl-hydroxy-methoxy-naphthazarin, acetyloxyethyl-hydroxy-dimethoxy-naphthalenedione | anti-inflammatory, cytotoxic (RAW 264.7) | [68] |
| Alantrypinone | α-glucosidase inhibitor | [28] |
| Altenusin | antioxidant, cytotoxic (B16, MCF-7, HepG2) antibacterial (S. aureus), antifungal (C. albicans) | [73] [51] |
| Amestolkolides A–B | anti-inflammatory | [100] |
| Amestolkines A–B | anti-inflammatory | [33] |
| Anhydrofusarubin, anhydrojavanicin | anti-inflammatory | [68] |
| Anserinone C | antibacterial (S. aureus), cytotoxic (MKN1) | [79] |
| Aspergillumarin A | α-glucosidase inhibitor antibacterial (E. coli, MRSA), antifungal (C. albicans) antioxidant | [32] [70] [30] |
| Aspergillumarin B | α-glucosidase inhibitor | [32] |
| Asperitaconic acid B | anti-inflammatory | [34] |
| Austin, austinol, austinolide, dehydroaustin, dehydroaustinol | antioxidant | [84] |
| Azaspirofuran A | anti-inflammatory | [47] |
| Bacillisporin A | α-glucosidase inhibitor, antibacterial (B. subtilis) antibacterial (S. aureus, MRSA) | [20] [54] |
| Bacillisporin B | α-glucosidase inhibitor, antibacterial (B. subtilis) antibacterial (E. faecalis, S.aureus, MRSA) antibacterial (S. aureus) | [20] [54] [74] |
| Bacillisporin C | antiproliferative (K562, HL-60, BGC-823, HeLa) | [104] |
| Bacillisporin F | protein tyrosine phosphatase inhibitor | [87] |
| Bacillisporins K–L | antibacterial (S. aureus) | [74] |
| Bromothiobenzoic acid | antibacterial (E. coli, K. pneumoniae, S. aureus), cytotoxic (HCT 116, HepG2 MCF-7) | [29] |
| Chrodrimanins A, C | antioxidant | [84] |
| Chrodrimanin B | protein tyrosine phosphatase inhibitor antioxidant | [101] [84] |
| Chrodrimanins K, N | antiviral (H1N1) | [21] |
| Chrodrimanins O, R–S | protein tyrosine phosphatase inhibitor | [101] |
| Chromosulfine | antiproliferative-proapoptotic (MCF-7, K562, HL-60, HeLa, BGC-823) | [102] |
| Citrorosein, questinol | lipid lowering | [83] |
| Conioxanthone A | immunosuppressive | [17] |
| Corymbiferan lactone A | antiproliferative (HL-60, BGC-823, HeLa) | [104] |
| Cyclosecosteroid A | acetylcholineterase inhibitor | [75] |
| Cyclotryprostatin B | anti-inflammatory | [47] |
| Dankasterone | antiproliferative (HL-60, A549, MCF-7, SW480) | [109] |
| Dehydroisopenicillide | anticholesterol, lipid lowering | [19] |
| Dihydro-hydroxy-hydroxymethyl-methoxy-methyl-naphthofurandione | anti-inflammatory | [68] |
| Dihydroxyconiothyrinone B | antibacterial (E. coli, E. tarda, S. aureus), antioxidant | [93] |
| Dihydroxy-dimethyl-dibenzofuran | antibacterial (E. coli, E. faecalis, MRSA, S. aureus), antifouling (B. neritina) | [89] |
| Dihydroxyfumitremorgin C | anti-inflammatory | [47] |
| Dihydroxy-hydroxybenzylidene-methylbutenyl-indane-carboxylic acid methyl ester | antibacterial (B. cereus, S. albus, S. aureus) | [28] |
| Dihydroxy-hydroxypentyl-isochromanone | α-glucosidase inhibitor α-glucosidase inhibitor, antioxidant | [32] [40] |
| Dihydroxy-hydroxypropyl-methyl-isochromenone, hydroxy-hydroxypropyl-methoxyisochromanone, hydroxypropyl-hydroxy-dihydroisocoumarin | antioxidant | [40] |
| Dihydroxyisocupressic acid | antibacterial (V. parahemolyticus) | [72] |
| Dihydroxy-methoxy-methylisochromenone, dihydroxy-pentyl-hydroxy-isochromanone, dimethyl-dihydroxyisocoumarin, hydroxy-hydroxyethyl-methoxyisocoumarin, hydroxy-hydroxymethyl-methoxy-methylisocoumarin, hydroxy-methoxy-dimethylisocoumarin | α-glucosidase inhibitors | [32] |
| Dihydroxy-methyldendryol E | antibacterial (S. aureus), antioxidant | [93] |
| Dihydroxy-methyl-hydroxymethyl-xanthone | antibacterial (A. hydrophila) | [45] |
| Diolhinokiic acid | antiproliferative (HL-60, A549) | [109] |
| Diorcinol, methoxycarbonyldiorcinol | antibacterial (E. coli, E. faecalis, MRSA, S. aureus), antifouling (B. neritina) | [89] |
| Ditalaromylectone A | antifungal (C. albicans) | [51] |
| Epoxyberkeleydione | lipid lowering | [59] |
| Ergosta-trienol | cytotoxic (HepG2, MCF-7) | [29] |
| Ethyl-dimethoxyjuglone | anti-inflammatory | [68] |
| Eurothiocin A | α-glucosidase inhibitor anti-inflammatory | [76] [38] |
| Eurothiocins D, F, G | α-glucosidase inhibitors | [48] |
| Expansols E–F | antifouling (B. neritina) | [89] |
| Fructigenines A–B | antiproliferative (K562, HeLa, HL-60, BGC-823, MCF-7) | [107] |
| Fumigaclavine C, fumigatin oxide, fumiquinazoline F, fumiquinone B | anti-inflammatory | [47] |
| Funicone, deoxyfunicone, 3-O-methylfunicone, hydroxyvermistatin, methoxyvermistatin | anticholesterol, lipid lowering | [19] |
| Fusarielins M, O, P | cytotoxic (B16) | [73] |
| Hydroxyconiothyrinone B | antibacterial (S. aureus), antioxidant | [93] |
| Hydroxy-ergosta-trienone | antioxidant | [84] |
| Hydroxy-hydroxyethyl-dimethoxy-naphthalenedione | anti-inflammatory, cytotoxic (RAW 264.7) | [68] |
| Hydroxy-hydroxy-hydroxymethyl-methylphenoxy-benzoic acid | protein tyrosine phosphatase inhibitor | [22] |
| Hydroxy-methoxy-benzofuranyl-ethanone, hydroxy-methoxy- methylbenzofuran-carboxylic acid | antibacterial (B. subtilis, E. coli, S. aureus, S. epidermidis) | [32] |
| Hydroxy-methyl-dimethoxycoumarin | antibacterial (MRSA), antifungal (C. albicans) | [70] |
| Hydroxypentacecilide A | antiviral (H1N1) | [21] |
| Hydroxypropyl-methyl-dihydroxychromone | antibacterial (Salmonella) | [20] |
| Isobutyric acid dihydroxy-methyl-oxo-dihydro-naphththalenyl methyl ester | α-glucosidase inhibitor | [32] |
| Isocyclopaldic acid | antibacterial (A. hydrophila, E. coli, M. luteus, P. aeruginosa, V. anguillarum, V. harveyi, V. parahemolyticus) | [45] |
| Isotryptoquivaline F | anti-inflammatory | [47] |
| Javanicin | anti-inflammatory, cytotoxic (RAW 264.7) | [68] |
| Macrosporusone D | antibacterial (S. aureus) | [74] |
| epi-Mangrovamide A | antibacterial (V. harveyi. V. parahaemolyticus) | [45] |
| Mangrovamide I | antibacterial (A. hydrophila, E. coli, M. luteus, P. aeruginosa, V. anguillarum, V. harveyi, V. parahemolyticus) | [45] |
| Methoxy-methyl-biphenyl-triol | antibacterial (S. aureus) | [51] |
| Methylhexahydrofuro-furanylethanol | cytotoxic (SF-268, MCF-7, HepG2, A549) | [96] |
| Methylincisterol, dimethylincisterol A3 | antifouling (B. neritina) | [89] |
| Methylpseurotin A, norpseurotin A | anti-inflammatory | [47] |
| Methyltenellate | lipid lowering | [19] |
| Monodictyphenone | protein tyrosine phosphatase inhibitor | [22] |
| Nafuredin B | cytotoxic (HeLa, MCF-7, K562, HCT 116, HL-60, A549) | [24] |
| Penicidone C | α-glucosidase inhibitor | [26] |
| Penicifuran A | antibacterial (E. coli, MRSA), antifungal (C. albicans) | [70] |
| Peniciisocoumarins C, F, G | antioxidant | [40] |
| Peniciisocoumarin D | α-glucosidase inhibitor, antioxidant antibacterial (E. coli, MRSA), antifungal (C. albicans) | [40] [70] |
| Peniciisocoumarin E | antioxidant | [30] |
| Peniciisocoumarin H | antibacterial (E. coli, MRSA), antifungal (C. albicans) | [27] |
| Penicillide | α-glucosidase inhibitor cytotoxic (H1975, HL7702, K562, MCF-7) | [26] [23] |
| Penicilloxalone B | antibacterial (E. coli, MRSA) antioxidant | [70] [30] |
| Penicimarins B–C | α-glucosidase inhibitor | [32] |
| Penicimarin G | antibacterial (B. cereus, E. coli, S. aureus), antioxidant α-glucosidase inhibitor, antioxidant | [92] [30] |
| Penicimarin H | antioxidant α-glucosidase inhibitor, antioxidant | [40] [30] |
| Penicimarin I | α-glucosidase inhibitor | [30] |
| Penicimarins L–M | antioxidant | [30] |
| Penicimarin N | α-glucosidase inhibitor, antioxidant | [40] |
| Penicimumide | antiproliferative (A549, HeLa, MCF-7, HepG2, NCI-H1975, HL-60, K562, LS180, SW480, HT29, BXPC-3, PANC-1) | [56] |
| Penicimutalidine | antiproliferative (K562, HL-60, BGC-823, HeLa) | [104] |
| Penicimutamides A–F | antiproliferative (K562, HL-60, BGC-823, HeLa) | [105,106] |
| Penicimutanines A, C | antiproliferative (K562, HeLa, HL-60, BGC-823, MCF-7) | [107] |
| Penicimutanolones A–B, penicimutanolone A methyl ether | antiproliferative (A549, HeLa, MCF-7, HCT 116, HepG2, NCI-H1975, HL-60, K562, LS180, SW480, HT29, PC-3, BXPC-3, PANC-1) | [56] |
| Penicimutide | antiproliferative (HeLa) | [18] |
| Penioxalicin | antibacterial (MRSA) | [80] |
| Peniphenone, pinselin | immunosuppressive | [17] |
| Penixanthones A–B | antiallergic cytotoxic (H1975, HL7702, K562, MCF-7) | [25] [23] |
| Penixanthones C–D | cytotoxic (K562, MCF-7, Huh7) | [90] |
| Pensulfonamide | antibacterial (E. coli, K. pneumoniae, S. aureus), antifungal (A. niger and C. albicans), cytotoxic (MCF-7, HCT 116, HepG2) | [29] |
| Pensulfonoxy | antibacterial (E. coli, K. pneumoniae, S. aureus), antifungal (A. niger), cytotoxic (HCT 116, HepG2) | [29] |
| Pestalotiorin | α-glucosidase inhibitor | [40] |
| Propenyl-octene-triol | cytotoxic (SF-268, MCF-7, HepG2, A549) | [96] |
| Purpactin A | antibacterial (H. pylori) α-glucosidase inhibitor | [77] [82] |
| Purpurides E–F | antibacterial (E. coli, MRSA), antifungal (C. albicans) | [27] |
| Purpuride G | antibacterial (E. coli, MRSA), antifungal (C. albicans), antiproliferative (U251, U87MG) | [27] |
| Purpurogenic acid | antiproliferative (K562, HL-60, HeLa, BGC-823) | [99] |
| Roussoellol C | antiproliferative (HL-60, A549, MCF-7, SW480) | [109] |
| Rugulosin A | antibacterial (S. aureus) | [74] |
| Rugulosuvine | antiproliferative (K562, HeLa, HL-60, BGC-823, MCF-7) | [107] |
| Sch1385568 | antibacterial (MRSA, S. aureus) | [54] |
| Sch725680 | α-glucosidase inhibitor, antibacterial (M. smegmatis, S. aureus) | [26] |
| Secalonic acid D | antibacterial (H. pylori), cytotoxic (Bel-7402, HCT 116) | [77] |
| Secopenicillide A | lipid lowering | [19] |
| Secopenicillide B | antibacterial (H. pylori) | [77] |
| Sequoiamonascin C | cytotoxic (B16, MCF-7) | [73] |
| Sescandelin | α-glucosidase inhibitor antibacterial (E. coli, MRSA), antifungal (C. albicans) | [32] [70] |
| Sescandelin B | α-glucosidase inhibitor | [32] |
| SF226 | antiproliferative (K562, HL-60, BGC-823, HeLa) | [104] |
| Solaniol | anti-inflammatory, cytotoxic (RAW 264.7) | [68] |
| Sydowinin A | immunosuppressive | [17] |
| Talabenzofuran C | α-glucosidase inhibitor | [76] |
| Talacyanol A | anti-inflammatory, cytotoxic (HCT-15, NUGC-3, MDA-MB-231, PC-3, NCI-H23, ACHN) | [38] |
| Taladrimanin A | antibacterial (S. aureus), antiproliferative-proapoptotic (MGC803, MKN28) | [78] |
| Talamin A | antibacterial (V. vulnificus), antioxidant | [52] |
| Talamin B | antibacterial (MRSA, V. vulnificus) | [52] |
| Talamin D | antioxidant | [52] |
| Talanaphthoquinone A | anti-inflammatory, cytotoxic (RAW 264.7) | [68] |
| Talaramide | mycobacterial PknG kinase inhibitor | [91] |
| Talarocyclopenta A | antibacterial (E. coli, S. aureus), anti-inflammatory | [34] |
| Talarocyclopenta B | antibacterial (B. cereus, B. subtilis, E. coli, M. tetragenus, S. albus, S. aureus), anti-inflammatory | [34] |
| Talarocyclopenta C | anti-inflammatory | [34] |
| Talarodilactones A–B | cytotoxic (L5178Y) | [63] |
| Talarodrides A–B | antibacterial (P. mirabilis, V. parahemolyticus) | [72] |
| Talaroisocoumarin A | antibacterial (E. coli, MRSA), antifungal (C. albicans) | [70] |
| Talaromarin F | antioxidant | [40] |
| Talarominine A | antibacterial (MRSA, M. luteus, P. aeruginosa, V. harveyi, V. vulnificus), antioxidant | [52] |
| Talaromynoid E | protein tyrosine phosphatase inhibitor | [59] |
| Talaromynoids G–I | lipid lowering | [59] |
| Talaromyolides D, I, K | antiviral (PRV) | [58,103] |
| Talaromyone A | antibacterial (H. pylori) | [77] |
| Talaromyone B | antibacterial (B. subtilis), α-glucosidase inhibitor | [82] |
| Talaromyoxaones A–B | protein tyrosine phosphatase inhibitors | [60] |
| Talaronin E | antibacterial (H. pylori) | [77] |
| Talaropeptins A–B | antifungal (F. oxysporum) | [108] |
| Talascortenes | antibacterial (A. hydrophila, E. coli, E. tarda, M. luteus, P. aeruginosa, V. harveyi, V. parahemolyticus), antifungal (C. gloeosporioides, F. oxysporum, G. graminis, R. cerealis) | [64] |
| Talasteroid | antioxidant | [84] |
| Talaverrucin A | Wnt/β-catenin pathway inhibitor | [81] |
| Tenellic acid A | α-glucosidase inhibitor | [82] |
| Tetrahydroxymethylbenzophenone, trihydroxymethylxanthone | antibacterial (E. coli, P. aeruginosa, S. aureus, V. alginolyticus, V. harveyi, V. parahaemolyticus), antioxidant | [49] |
| Trihydroxybutyl-hydroxy-hydroxy-methylphenoxy-methylphenylacetate | antibacterial (E. coli, E. faecalis, MRSA, S. aureus) | [89] |
| Trihydroxy-hydroxyethyl-isocoumarin | α-glucosidase inhibitor | [32] |
| Trihydroxy-methoxymethylbenzophenone | antioxidant | [49] |
| Vermistatin | anticholesterol, lipid lowering cytotoxic (B16) | [19] [73] |
| Verruculide B2 | antibacterial (S. aureus) | [21] |
| Verruculosin A, xenoclauxin | protein tyrosine phosphatase inhibitor | [87] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicoletti, R.; Bellavita, R.; Falanga, A. The Outstanding Chemodiversity of Marine-Derived Talaromyces. Biomolecules 2023, 13, 1021. https://doi.org/10.3390/biom13071021
Nicoletti R, Bellavita R, Falanga A. The Outstanding Chemodiversity of Marine-Derived Talaromyces. Biomolecules. 2023; 13(7):1021. https://doi.org/10.3390/biom13071021
Chicago/Turabian StyleNicoletti, Rosario, Rosa Bellavita, and Annarita Falanga. 2023. "The Outstanding Chemodiversity of Marine-Derived Talaromyces" Biomolecules 13, no. 7: 1021. https://doi.org/10.3390/biom13071021