
Widely Targeted HPLC-MS/MS Metabolomics Analysis Reveals Natural Metabolic Insights in Insects
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Metabolite Extraction
2.3. UPLC Conditions
2.4. QTOF-MS/MS
2.5. ESI-Q TRAP-MS/MS
2.6. PCA
2.7. Hierarchical Cluster Analysis and Pearson Correlation Coefficients
2.8. Differential Metabolite Analysis
2.9. KEGG Pathway Analysis
3. Results
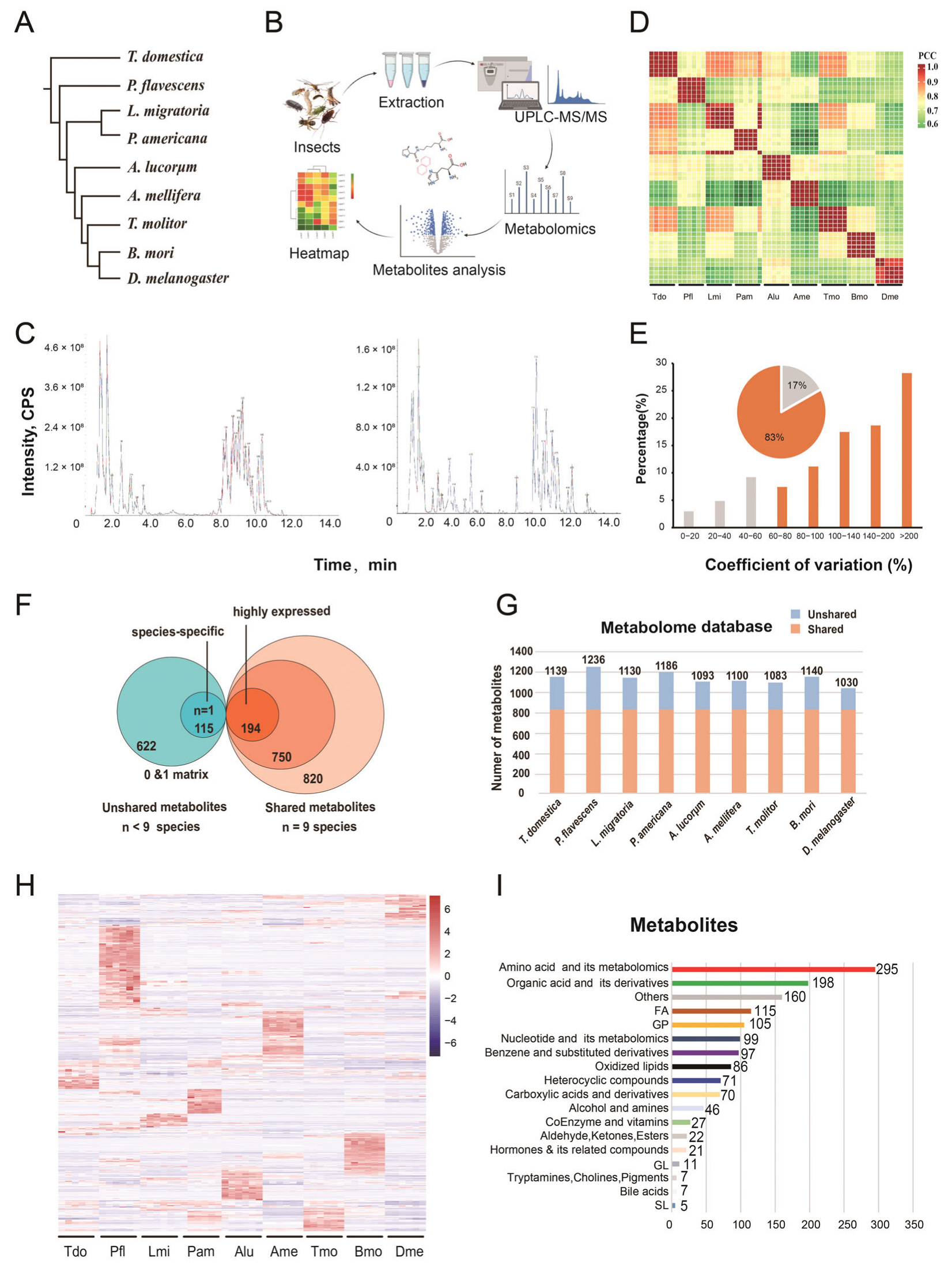
3.1. Construction of a Metabolic Dataset
3.2. The Identified Metabolites of Nine Species for the Global Overview
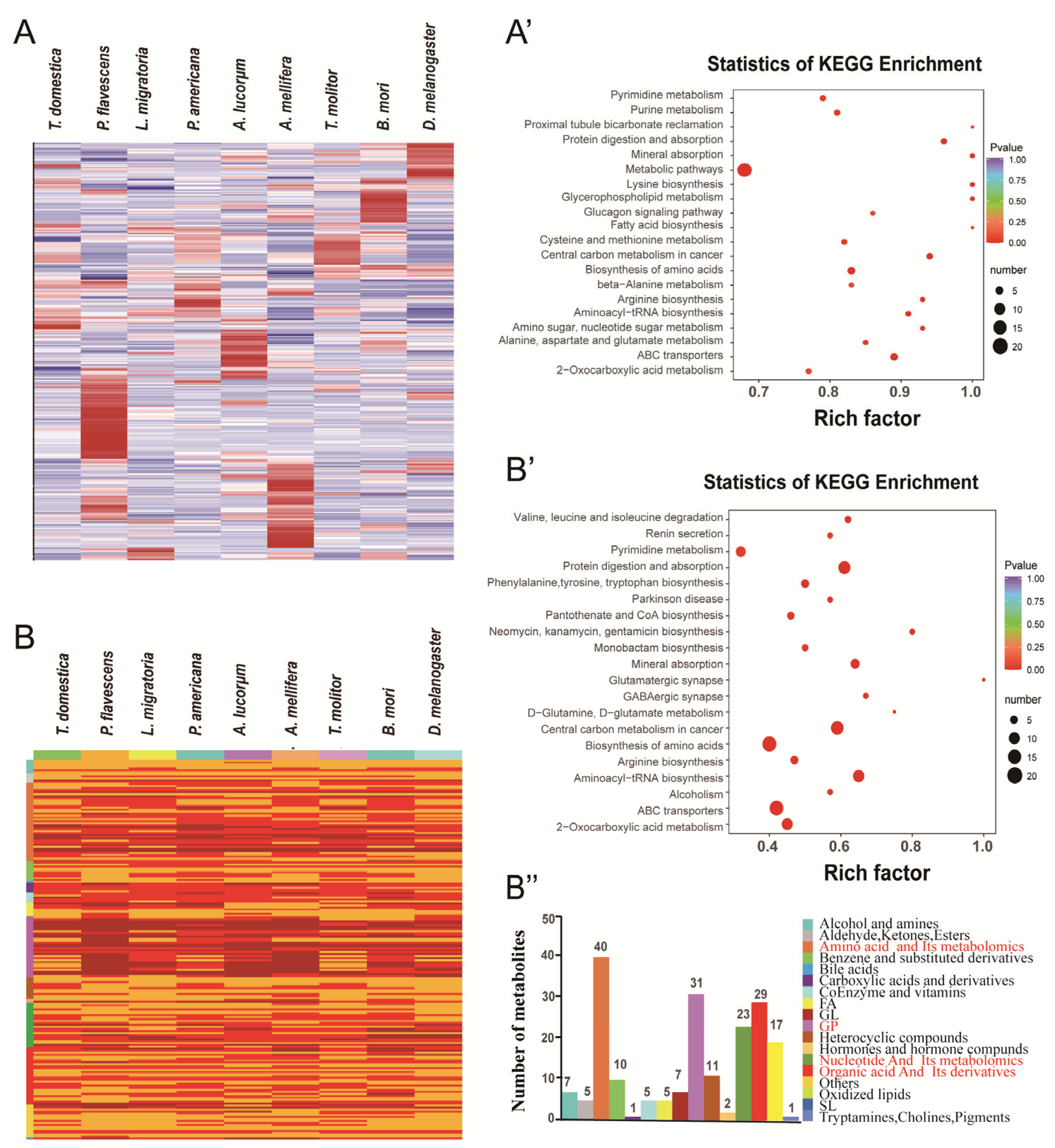
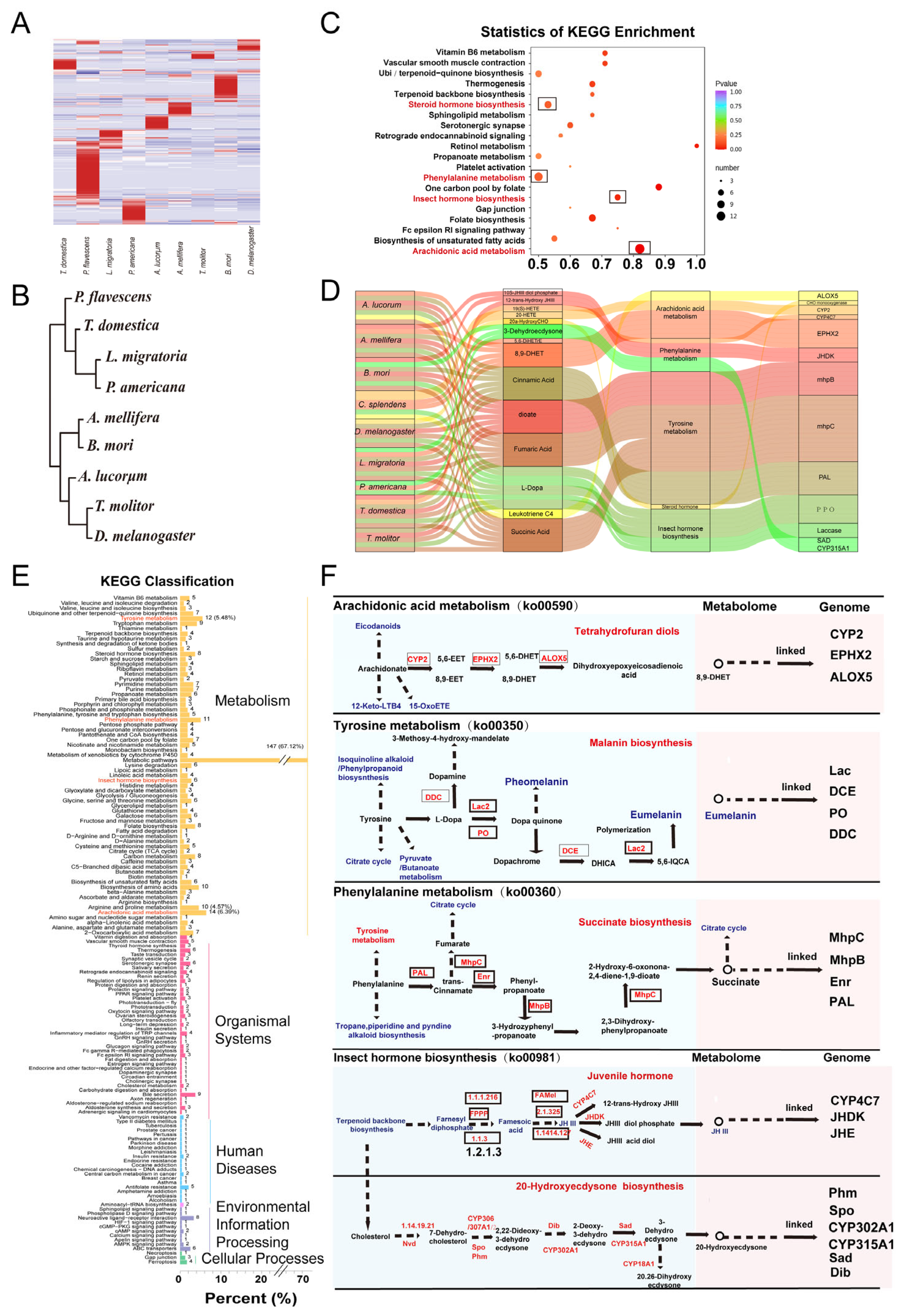
3.3. Common Metabolic Enrichments and Pathways
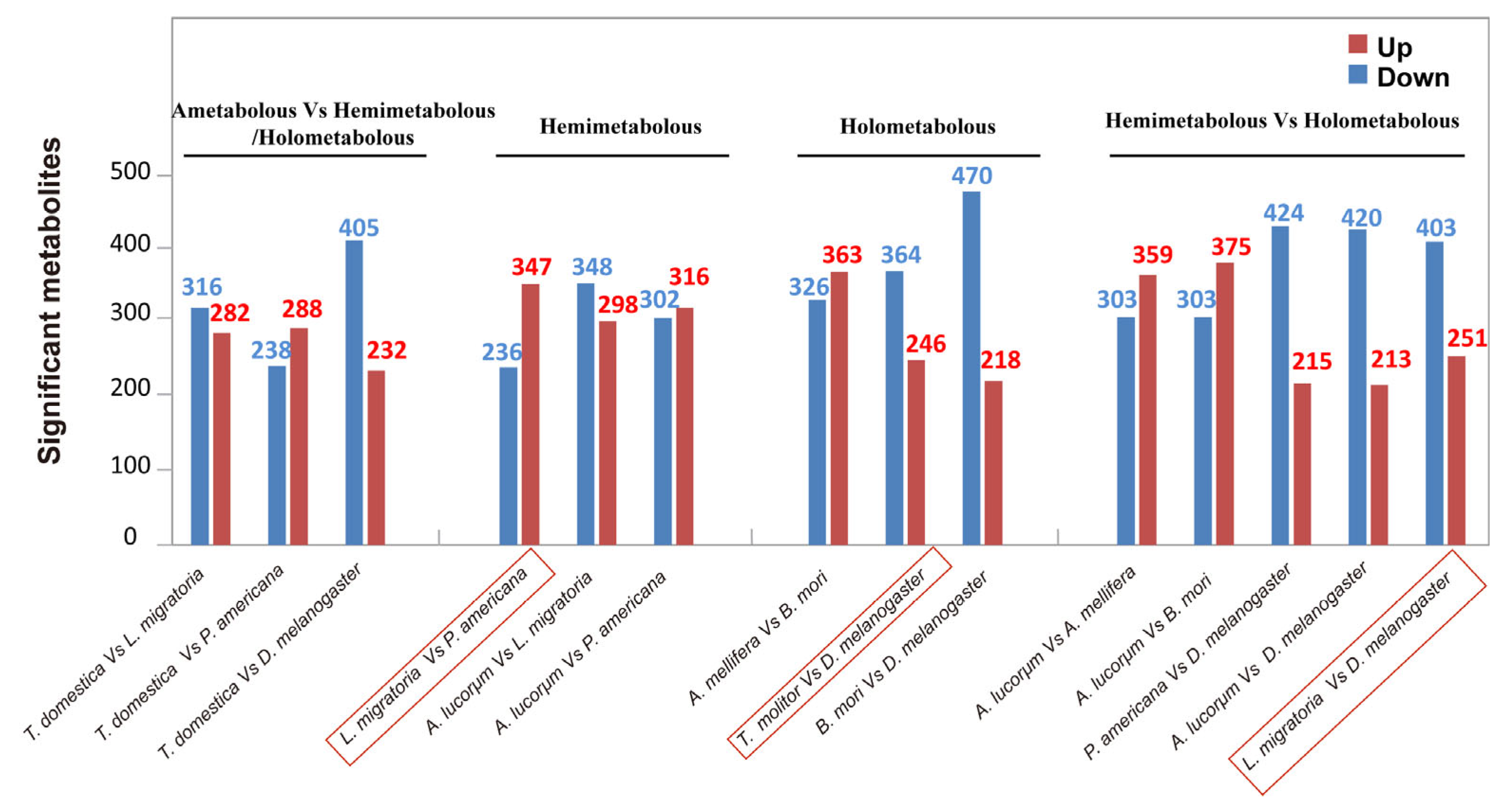
3.4. Comparison of the Metabolites between Hemimetabolous and Holometabolous Insects
3.5. Quantitative Statistics of Significantly Changed Metabolites between the Species of Interest
3.6. The Quality of Metabolites Contributes to the Cluster Relationship of Insects
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saito, K.; Matsuda, F. Metabolomics for functional genomics, systems biology, and biotechnology. Annu. Rev. Plant Biol. 2010, 61, 463–489. [Google Scholar] [CrossRef] [PubMed]
- Cromartie, R.I.T. Insect Pigments. Annu. Rev. Entomol. 1959, 4, 59–76. [Google Scholar] [CrossRef]
- Yew, J.Y.; Chung, H. Insect pheromones: An overview of function, form, and discovery. Prog. Lipid Res. 2015, 59, 88–105. [Google Scholar] [CrossRef] [PubMed]
- Truman, J.W.; Riddiford, L.M. The evolution of insect metamorphosis: A developmental and endocrine view. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20190070. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.A. The evolutionary dynamics of venom toxins made by insects and other animals. Biochem. Soc. Trans. 2020, 48, 1353–1365. [Google Scholar] [CrossRef]
- Song, J.; Zhou, S. Post-transcriptional regulation of insect metamorphosis and oogenesis. Cell. Mol. Life Sci. 2020, 77, 1893–1909. [Google Scholar] [CrossRef]
- Okude, G.; Futahashi, R. Pigmentation and color pattern diversity in Odonata. Curr. Opin. Genet. Dev. 2021, 69, 14–20. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Rivera-Perez, C.; Nouzova, M.; Lamboglia, I.; Noriega, F.G. Metabolic analysis reveals changes in the mevalonate and juvenile hormone synthesis pathways linked to the mosquito reproductive physiology. Insect Biochem. Mol. Biol. 2014, 51, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Malmendal, A.; Overgaard, J.; Bundy, J.G.; Sorensen, J.G.; Nielsen, N.C.; Loeschcke, V.; Holmstrup, M. Metabolomic profiling of heat stress: Hardening and recovery of homeostasis in Drosophila. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Colinet, H.; Renault, D.; Charoy-Guevel, B.; Com, E. Metabolic and proteomic profiling of diapause in the aphid parasitoid Praon volucre. PLoS ONE 2012, 7, e32606. [Google Scholar] [CrossRef] [Green Version]
- Kapranas, A.; Snart, C.J.; Williams, H.; Hardy, I.C.; Barrett, D.A. Metabolomics of aging assessed in individual parasitoid wasps. Sci. Rep. 2016, 6, 34848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Wu, Z.; Wang, X.; Yang, P.; Yu, D.; Zhao, C.; Xu, G.; Kang, L. Metabolomic analysis reveals that carnitines are key regulatory metabolites in phase transition of the locusts. Proc. Natl. Acad. Sci. USA 2012, 109, 3259–3263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Betel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Norbert, H.H. Transport and Utilization of Lipids in Insect Flight Muscles. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1997, 117, 475–482. [Google Scholar]
- Fiehn, O. Combining genomics, metabolome analysis, and biochemical modelling to understand metabolic networks. Comp Funct Genomics. 2001, 2, 155–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weckwerth, W. Metabolomics in systems biology. Annu. Rev. Plant Biol. 2003, 54, 669–689. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Dahuja, A.; Kumar, R.R.; Sakhare, A.; Watts, A.; Singh, B.; Goswami, S.; Sachdev, A.; Praveen, S. Role of ATP-binding cassette transporters in maintaining plant homeostasis under abiotic and biotic stresses. Physiol. Plant. 2021, 171, 785–801. [Google Scholar] [CrossRef]
- Ofori, P.A.; Mizuno, A.; Suzuki, M.; Martinoia, E.; Reuscher, S.; Aoki, K.; Shibata, D.; Otagaki, S.; Matsumoto, S.; Shiratake, K. Genome-wide analysis of ATP binding cassette (ABC) transporters in tomato. PLoS ONE 2018, 13, e0200854. [Google Scholar] [CrossRef] [Green Version]
- Mo, J.; Ma, Z.; Yan, S.; Cheung, N.K.; Yang, F.; Yao, X.; Guo, J. Metabolomic profiles in a green alga (Raphidocelis subcapitata) following erythromycin treatment: ABC transporters and energy metabolism. J. Environ. Sci. 2023, 124, 591–601. [Google Scholar] [CrossRef]
- Stottmeister, U.; Aurich, A.; Wilde, H.; Andersch, J.; Schmidt, S.; Sicker, D. White biotechnology for green chemistry: Fermentative 2-oxocarboxylic acids as novel building blocks for subsequent chemical syntheses. J. Ind. Microbiol. Biotechnol. 2005, 32, 651–664. [Google Scholar] [CrossRef]
- Xu, Y.J.; Luo, F.; Gao, Q.; Shang, Y.; Wang, C. Metabolomics reveals insect metabolic responses associated with fungal infection. Anal. Bioanal. Chem. 2015, 407, 4815–4821. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.O. Insect cuticular sclerotization: A review. Insect Biochem. Mol. Biol. 2010, 40, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Noh, M.Y.; Muthukrishnan, S.; Kramer, K.J.; Arakane, Y. Cuticle formation and pigmentation in beetles. Curr. Opin. Insect Sci. 2016, 17, 1–9. [Google Scholar] [CrossRef]
- Luo, W.; Liu, S.; Zhang, W.; Yang, L.; Huang, J.; Zhou, S.; Feng, Q.; Palli, S.R.; Wang, J.; Roth, S.; et al. Juvenile hormone signaling promotes ovulation and maintains egg shape by inducing expression of extracellular matrix genes. Proc. Natl. Acad. Sci. USA 2021, 118, e2104461118. [Google Scholar] [CrossRef] [PubMed]
- Swevers, L.; Raikhel, A.S.; Sappington, T.W.; Shirk, P.; Iatrou, K. Vitellogenesis and Post-Vitellogenic Maturation of the Insect Ovarian Follicle. In Comprehensive Molecular Insect Science; Elsevier: Amsterdam, The Netherlands, 2005; pp. 87–136. [Google Scholar]
- Treidel, L.A.; Quintanilla Ramirez, G.S.; Chung, D.J.; Menze, M.A.; Vazquez-Medina, J.P.; Williams, C.M. Selection on dispersal drives evolution of metabolic capacities for energy production in female wing-polymorphic sand field crickets, Gryllus firmus. J. Evol. Biol. 2022, 35, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Beenakkers, A.M.T.; Van der Horst, D.J.; Van Marrewijk, W.J.A. Role of lipids in energy metabolism. In Energy Metabolism in Insects; Plenum Publishing Corporotion: New York, NY, USA, 1981; pp. 53–100. [Google Scholar]
- Schilder, R.J.; Marden, J.H. Metabolic syndrome and obesity in an insect. Proc. Natl. Acad. Sci. USA 2006, 103, 18805–18809. [Google Scholar] [CrossRef] [Green Version]
- An, P.N.; Yamaguchi, M.; Bamba, T.; Fukusaki, E. Metabolome analysis of Drosophila melanogaster during embryogenesis. PLoS ONE 2014, 9, e99519. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Hou, K.; Zhang, H.; Wang, X.; Wu, W. Integrating transcriptomics and metabolomics to studies key metabolism, pathways and candidate genes associated with drought-tolerance in Carthamus tinctorius L. Under drought stress. Ind. Crops Prod. 2020, 151, 112465. [Google Scholar] [CrossRef]
- Lu, Y.; Shao, M.; Xiang, H.; Zheng, P.; Wu, T.; Ji, G. Integrative transcriptomics and metabolomics explore the mechanism of kaempferol on improving nonalcoholic steatohepatitis. Food Funct. 2020, 11, 10058–10069. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gao, Y.; Xie, W.; Gong, L.; Lu, K.; Wang, W.; Li, Y.; Liu, X.; Zhang, H.; Dong, H.; et al. Genome-wide association analyses provide genetic and biochemical insights into natural variation in rice metabolism. Nat. Genet. 2014, 46, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the Fruit Metabolome in Tomato Breeding. Cell 2018, 172, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Cheng, Y.; Chen, J.; Xu, W.; Ma, W.; Li, S.; Du, E. Widely Targeted HPLC-MS/MS Metabolomics Analysis Reveals Natural Metabolic Insights in Insects. Metabolites 2023, 13, 735. https://doi.org/10.3390/metabo13060735
Li Z, Cheng Y, Chen J, Xu W, Ma W, Li S, Du E. Widely Targeted HPLC-MS/MS Metabolomics Analysis Reveals Natural Metabolic Insights in Insects. Metabolites. 2023; 13(6):735. https://doi.org/10.3390/metabo13060735
Chicago/Turabian StyleLi, Zhaoxin, Yunlong Cheng, Jinxin Chen, Weijun Xu, Wentao Ma, Sheng Li, and Erxia Du. 2023. "Widely Targeted HPLC-MS/MS Metabolomics Analysis Reveals Natural Metabolic Insights in Insects" Metabolites 13, no. 6: 735. https://doi.org/10.3390/metabo13060735